
The Impact of COVID-19 on People Living with HIV-1 and HIV-1-Associated Neurological Complications




Abstract
:1. Introduction
2. Similarities and Differences between SARS-CoV-2 and HIV-1
- (a)
- Fear among the public is the most common attribute of both SARS-CoV-2 and HIV-1 viral infection. This public fear makes people psychologically ill, leading to stress and anxiety [42].
- (b)
- Both SARS-CoV-2 and HIV-1 are enveloped viruses with single-stranded RNA as the genome.
- (c)
- Both the genomes of SARS-CoV-2 and HIV-1 are prone to mutation, and the accumulation of mutations within the host under selection pressure results in the emergence of new variants. Furthermore, immunocompromised PLWH harbor SARS-CoV-2 for a longer time, providing ample time for mutant accumulation and resulting in the SARS-CoV-2 variant stemming [44].
- (d)
- SARS-CoV-2 and HIV-1 have zoonotic origins and were transmitted to humans from animal reservoirs, HIV-1 from non-human primates (NHPs) and SARS-CoV-2 from bats.
- (e)
- In respective natural reservoirs, SARS-CoV-2 and HIV-1 infections produce mild to no symptoms but incite disease upon human infection.
- (f)
- One reason for the widespread COVID-19 and AIDS pandemics is the transmission of SARS-CoV-2 and HIV-1 via asymptomatically infected individuals.
- (g)
- Lymphopenia as a result of drastic loss of CD4+T cells occurs due to HIV-1 and SARS-CoV-2 infection and is considered a prognostic marker [45,46,47]. There is a substantial drop in CD4+T cell counts in the acute phase of HIV-1 infection in contrast to the chronic phase, where a continued decline in CD4+T cells occurs and leads to AIDS. Lymphocytopenia is the hallmark of COVID-19 severity, but elevated levels of CD4+T and CD8+T cells were associated with milder disease conditions [45].
- (h)
- SARS-CoV-2 and HIV-1 induce neutrophil extracellular traps (NETs) and cause NETosis, a neutrophil death mechanism. NETosis also may cause increased secretion of chemokines and cytokines, leading to increased inflammation.
- (i)
- Both SARS-CoV-2 and HIV-1 induce higher proinflammatory cytokine secretion and inflammation.
- (j)
- Increased serum levels of proinflammatory cytokines in SARS-CoV-2 and HIV-1 infected patients are considered to be biomarkers and are predictive variables associated with morbidity and mortality.
- (k)
- Both these viruses, SARS-CoV-2 and HIV-1 infection, lead to immune dysregulation and are mediated by inflammasome activation. These viruses can activate NLRP3 inflammasome in different cells, including monocytes/macrophages and microglia. SARS-CoV-2-induced Microglial and macrophage NLRP3 inflammasome activation in the CNS results in neuroinflammation, causing myriad neurological manifestations.
- (l)
- A portion of SARS-CoV-2 and HIV-1 infected individuals develop neurocognitive impairments.
3. HIV-1-Induced Neurological Manifestations
4. Impact of SARS-CoV-2 on the CNS
5. Impact of COVID-19 on PLWH
5.1. Risk Factors Associated with COVID-19 in PLWH
- i.
- Age: It is well known that older people are more susceptible to the COVID-19 pandemic. At the beginning of the pandemic, biological age was recognized as a significant risk factor. Several studies revealed that older people are more likely to become infected with SARS-CoV-2 and COVID-19-related hospitalization and mortality. According to the Centers for Disease Control and Prevention (CDC), people aged 85 or older have a 13-fold higher risk of hospitalization and 630-fold death than those aged 18–29. However, people between 50 and 85 are at a 4–8-fold higher risk of hospitalization and 30–220-fold of death [122,123]. As a result of stringent cART regiments, half of PLWH in the United States are 50 or older [118,119,124]. These older PLWH with several age-related comorbidities and compromised immune systems are at increased risk of COVID-19 complications. They usually experience physical and cognitive impairments, chronic immune activation and multimorbidity even without COVID-19 at ages younger than HIV-1 negative people [125,126,127,128,129]. Thus, aging and age-associated comorbidity make PLWH succumb to SARS-CoV-2 infection and COVID-19 severity [130]. Since COVID-19 also causes neurological syndromes, SARS-CoV-2 may imply a cumulative effect on preexisting HAND or may lead to the development of HAND. Nonetheless, investigating the impact of COVID-19 on aged PLWH with HAND is imperative in order to mitigate the devastating impact of the syndemic.
- ii.
- Sex: There are gender biases recorded for SARS-CoV-2 infection and COVID-19-mediated disease severity. It was found that SARS-CoV-2-infected men aged between 40 and 70 years proceed to COVID-19 severity with double the risk of mortality than the same age group of women [131,132,133,134]. The sex-biasedness in the risk of SARS-CoV-2 infection and COVID-19-induced disease severity is most likely due to sex-based differential immune responses and immunomodulatory effectors such as sex hormones and sex-specific comorbidity [135]. The sex-based biases are possibly due to male-induced nonclassical monocytes and increased cytokines (IL-8 and IL-18) production in contrast to women-induced robust CD8+T cell response [136]. Additionally, lesser vulnerability and COVID-19 severity in women may be associated with enhanced neutrophil activity and increased type I interferon (IFN-I), generating robust innate immune response via TLRs [137].Additionally, in males, the ACE2 and TMPRSS2 responsible for SARS-CoV-2 infection and pathogenesis were elevated compared to females [89,138,139]. Both ACE2 and TMPRSS2 are androgen-responsive [140,141,142,143]. Men with lower testosterone levels were found to be associated with severe COVID-19 outcomes [140,141,142,143]. The lower testosterone levels in men were also linked to higher proinflammatory cytokine [144]. Thus, androgen deficiency and testosterone dysregulation could modulate ACE2 and TMPRSS2 expression, further influencing COVID-19 outcomes in PLWH [145]. In contrast, the impact of COVID-19 in PLWH was the opposite regarding sex bias, with increased morbidity and mortality recorded in women than in men [146,147]. Such a discrepancy could be due to stronger immune activation and increased inflammatory markers in women, despite similar viral suppression among men and women under strict cART regimens [148]. Although these initial studies have concluded the opposite incidence of SARS-CoV-2 infection and COVID-19 outcomes in PLWH, the sex-biased differential impact warrants further investigation and critical interpretation.
- iii.
- Comorbidities: As mentioned above, older people are at higher risk of COVID-19 severity; this may be due to the presence of comorbidities in people of this age group. PLWH are often with several comorbidities, including dysregulated immunity, chronic diseases in the lung, kidney and liver, and obesity, diabetes, hypertension, hyperlipidemia, cardiovascular disease and other health issues, which may exacerbate the COVID-19 severity [21,22,149,150,151,152,153]. PLWH with multiple comorbidities are victims of disease severity and possess an increased chance of mortality [154,155]. However, studies have revealed that PLWH adhering to cART, with adequate CD4+T cell count, viral suppression and without comorbidities do not have a higher risk of COVID-19 severity and mortality than non-PLWH [156]. Future studies are warranted to delineate how these comorbidities affect COVID-19 severity and mortality in PLWH and PLWH with impaired neurocognitive function.
- iv.
- Immunity: The role of the immune system is to protect us from infections and diseases. Any alteration or dysregulation in the immune system makes us vulnerable to infection and severe disease outcomes. People with compromised immunity are at higher risk of SARS-CoV-2 infection and COVID-19 severity. In PLWH, the immune system is dysregulated despite the cART regimen due to a lack of immune reconstitution or loss of immunological memory [157,158,159]. In a recent study, the cART regimen exhibited potential protective effects on the incidence and severity of COVID-19 in PLWH [160]. However, after clinical trials of several antiretrovirals used to treat HIV-1, only partial protection was observed, and more clinical trials may provide us with a better understanding of using antiretrovirals to treat SARS-CoV-2 infection [161]. Another study on PLWH showed that neither HIV-1 plasma viral load nor CD4+T cell count at the time of diagnosis determined COVID-19 outcomes [121], despite SARS-CoV-2 and HIV-1 infection-mediated T cell lymphopenia [46]. On the contrary, recent studies presented on CROI 2022 reported the worst COVID-19 clinical outcomes in PLWH with recent CD4+T cell counts of <200. More dedicated meta-analysis is required to evaluate the effects of antivirals on SARS-CoV-2 infection and COVID-19 severity. Viruses can inhibit T cell receptor signaling and immune response, resulting in immune system dysregulation and imposing severe outcomes of COVID-19 in PLWH. Furthermore, the emergence of SARS-CoV-2 variants make PLWH who are immunocompromised more susceptible to accumulating mutations during viral replication. These immunocompromised individuals may harbor viruses for a longer time, providing an opportunity for variant emergence. HIV may “trigger” the emergence of SARS-CoV-2 variants reported by a study carried out in South Africa because of the high number of PLWH in this region [44,162].
- v.
- Socioeconomic inequalities: In this world, millions of people suffer from the risk of the social determinants of health inequalities, including the economic conditions for affordable healthcare due to poverty [163]. PLWH are often with socioeconomic burden due to health conditions and loss of income sources. Poor people with financial burdens are unhealthy due to poor diet and inability to afford appropriate healthcare; thus, the people in this group are more vulnerable to infection and disease progression to severity. The living hygiene condition deteriorates with food insecurity and the absence of proper care. Further, the economic uncertainty is disproportionate and evident in some communities, such as the black and Hispanic populations, and people in these communities represent most sufferers of these conditions. There are several worldwide studies on determination of socioeconomic factors that affect disease progression and related mortality. It is believed that socioeconomic burden disproportionately impacts the risk of SARS-CoV-2 infection and disease severity. Thus, poor people with socioeconomic disadvantage are in the high-risk category for SARS-CoV-2 infection and disease severity due to their poor health conditions and immunocompromised status.
- vi.
- Substance abuse: Early in the COVID-19 pandemic, a steep rise in substance abuse, drug overdose and mental health challenges was reported [164]. Among PLWH, drug abuse is common, further burdening COVID-19 severity and mortality as COVID-19 imposed social isolation resulting in mental health deterioration and strained PLWH toward substance use disorders (SUDs) [165]. People with SUD (PWSUD) might have greater exposure and health challenges leading to higher susceptibility to SARS-CoV-2 infection [166]. PWSUD are poor in adhering to social distancing and following COVID-19 guidelines; this may be due to sharing syringes for substance use, especially in large congregate settings [167,168,169]. People with previously diagnosed SUDs have an 8-fold higher possibility of SARS-CoV-2 infection than people without SUD [170]. Thus, it is evident that PWSUD have a higher possibility of SARS-CoV-2 infection and COVID-19 severity. Different SUD types also determine exposure and illness, such as people with opioid use disorders (OUD) and cannabis use disorder (CAUD), who are at 10.2% and 5.3% higher likelihood of SARS-CoV-2 infection and COVID-19 outcomes [170]. In addition, alcohol use disorder (AUD) and Methamphetamine (MA) use disorder also increased due to the lockdown and stay home policy. There is an increased incidence of COVID-19 diagnosis, hospitalization and fatal outcomes in PWSUD compared to people without SUD [170,171]. Some PWSUD may have dysregulated immune responses and comorbidities, including HIV-1 infection, which can further exacerbate COVID-19-mediated complications [172]. There are reports of psychiatric association with SUD and increased incidence of COVID-19-related hospitalization and extended hospital stay [173]. Overall, PWSUD have higher vulnerability for SARS-CoV-2 infection and COVID-19 outcomes.
5.2. SARS-CoV-2 Infection and COVID-19 Severity in PLWH
5.3. COVID-19 Imposed Neurological Outcomes on PLWH
6. Mechanisms of COVID-19-Induced Neurological Manifestations in PLWH
6.1. Inflammasome Activation
6.2. Neurotoxic Activities of SARS-CoV-2 Proteins
6.3. Other
7. SARS-CoV-2/HIV-1 Syndemic Challenges
- (a).
- Fear among PLWH for SARS-CoV-2 infection has decreased their engagement in care. They are even scared to visit pharmacies to collect cART. This fear is heightened in the SARS-CoV-2/HIV-1 syndemic, and individuals remain unaware of how the future changes due to COVID-19 may impact the ongoing antiretroviral treatment against HIV-1.
- (b).
- Broad-scale COVID-19-imposed lockdown measures to contain the SARS-CoV-2 infection culminated in restrictions to movements and the suspension of public and private transportation. These restrictions have hindered engagement with HIV testing services and access [272].
- (c).
- (d).
- The COVID-19 pandemic has overwhelmed the worldwide healthcare system. The high demand and shortage of healthcare workers enforced the engagement of HIV-1 physicians in COVID-19 care, resulting in a lack of care and routine testing for PLWH [275].
- (e).
- The diversion of healthcare workers and facilities disrupted testing and the identification of drug resistance and opportunistic infections. This unavailability of care and enforced disruption in cART furthers the emergence of drug resistance due to the accumulation of mutations and leads to death due to opportunistic infections [36].
- (f).
- It was speculated from multiple mathematical models that the disruption of cART may increase HIV-1/AIDS-related deaths [276].
- (g).
- HIV-1 vaccine discovery is severely hindered due to the COVID-19 pandemic. The lockdowns, restrictions and emergency regulations owing to the pandemic clinical trials of HIV-1 vaccines are derailed, resulted in a delay in reduction ofHIV-1 vaccine discovery.
- (h).
- The issue of vaccine hesitancy among PLWH is a primary concern. Unvaccinated people are at higher risk of SARS-CoV-2 infection and COVID-19 severity, which may worsen HAND [277]. There must be awareness and priority to vaccinate PLWH. Additionally, a lower level of anti-SARS-CoV-2 spike protein antibody was reported in PLWH compared to non-PLWH control after vaccination with mRNA-1273 or BNT126b2 [278]. The suggestion is that higher dosages and frequent booster dosages are essential for PLWH to maintain long-term immunity at the level of an average person.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lucas, S. Pandemics and pathology: A reflection on influenza, HIV/AIDS and SARS (COVID-19) pandemic infections. Diagn. Histopathol. 2021, 27, 128–133. [Google Scholar] [CrossRef]
- Olufadewa, I.; Oduguwa, I.; Adesina, M.; Ibiang, K.; Eke, N.; Adewumi, B.; Ebong, I.; Abudu, F.; Adeyelu, N. COVID-19: Learning from the HIV/AIDS pandemic response in Africa. Int. J. Health Plan. Manag. 2021, 36, 610–617. [Google Scholar] [CrossRef]
- Demoliou, C.; Papaneophytou, C.; Nicolaidou, V. SARS-CoV-2 and HIV-1: So Different yet so Alike. Immune Response at the Cellular and Molecular Level. Int. J. Med. Sci. 2022, 19, 1787–1795. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N. Engl. J. Med. 2020, 382, 1199–1207. [Google Scholar] [CrossRef]
- Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef]
- Weiss, R.A. How does HIV cause AIDS? Science 1993, 260, 1273–1279. [Google Scholar] [CrossRef]
- Fauci, A.S.; Macher, A.M.; Longo, D.L.; Lane, H.C.; Rook, A.H.; Masur, H.; Gelmann, E.P. NIH conference. Acquired immunodeficiency syndrome: Epidemiologic, clinical, immunologic, and therapeutic considerations. Ann. Intern. Med. 1984, 100, 92–106. [Google Scholar] [CrossRef]
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 8 December 2022).
- UNAIDS. AIDS BY THE NUMBERS. UNAIDS: Geneva, Switzerland. Available online: https://www.unaids.org/en (accessed on 8 December 2022).
- World Health Organization. HIV Data and Statistics. Available online: https://www.who.int/teams/global-hiv-hepatitis-and-stis-programmes/hiv/strategic-information/hiv-data-and-statistics (accessed on 9 December 2022).
- Shiau, S.; Krause, K.D.; Valera, P.; Swaminathan, S.; Halkitis, P.N. The Burden of COVID-19 in People Living with HIV: A Syndemic Perspective. AIDS Behav. 2020, 24, 2244–2249. [Google Scholar] [CrossRef] [PubMed]
- Gatechompol, S.; Avihingsanon, A.; Putcharoen, O.; Ruxrungtham, K.; Kuritzkes, D.R. COVID-19 and HIV infection co-pandemics and their impact: A review of the literature. AIDS Res. Ther. 2021, 18, 28. [Google Scholar] [CrossRef]
- Dutschke, A.; Wejse, C.; Nanque, J.P.; Medina, C.; Honge, B.L.; Jespersen, S.; Bissau HIV Cohort Study Group. SARS-CoV-2 seroprevalence among people living with HIV in Guinea-Bissau. Public Health 2022, 209, 36–38. [Google Scholar] [CrossRef]
- Kalichman, S.C.; Eaton, L.A.; Berman, M.; Kalichman, M.O.; Katner, H.; Sam, S.S.; Caliendo, A.M. Intersecting Pandemics: Impact of SARS-CoV-2 (COVID-19) Protective Behaviors on People Living with HIV, Atlanta, Georgia. J. Acquir. Immune Defic. Syndr. 2020, 85, 66–72. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, G.; Springer, G.; Gustafson, D.; Kassaye, S.; Alcaide, M.L.; Ramirez, C.; Sharma, A.; Palella, F.J.; Tien, P.C.; Detels, R.; et al. COVID-19 symptoms and SARS-CoV-2 infection among people living with HIV in the US: The MACS/WIHS combined cohort study. HIV Res. Clin. Pract. 2020, 21, 130–139. [Google Scholar] [CrossRef] [PubMed]
- UNAIDS. People Living with HIV at Higher Risk of COVID-19 Illness but Have Lower Access to COVID-19 Vaccines. Available online: https://www.unaids.org/en/site-search#?cludoquery=COVID-19%20HIV%20double%20jeopardy%20&cludopage=1&cludorefurl=https%3A%2F%2Fwww.unaids.org%2Fen&cludorefpt=UNAIDS&cludoinputtype=standard (accessed on 7 December 2022).
- World Health Organization. Coronavirus Disease (COVID-19) and People Living with HIV. Available online: https://www.who.int/news-room/questions-and-answers/item/coronavirus-disease-(COVID-19)-COVID-19-and-people-living-with-hiv (accessed on 28 December 2022).
- Gervasoni, C.; Meraviglia, P.; Riva, A.; Giacomelli, A.; Oreni, L.; Minisci, D.; Atzori, C.; Ridolfo, A.; Cattaneo, D. Clinical Features and Outcomes of Patients with Human Immunodeficiency Virus with COVID-19. Clin. Infect. Dis. 2020, 71, 2276–2278. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, K.; Rentsch, C.T.; MacKenna, B.; Schultze, A.; Mehrkar, A.; Bates, C.J.; Eggo, R.M.; Morton, C.E.; Bacon, S.C.J.; Inglesby, P.; et al. HIV infection and COVID-19 death: A population-based cohort analysis of UK primary care data and linked national death registrations within the OpenSAFELY platform. Lancet HIV 2021, 8, e24–e32. [Google Scholar] [CrossRef]
- Shalev, N.; Scherer, M.; LaSota, E.D.; Antoniou, P.; Yin, M.T.; Zucker, J.; Sobieszczyk, M.E. Clinical Characteristics and Outcomes in People Living with Human Immunodeficiency Virus Hospitalized for Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2294–2297. [Google Scholar] [CrossRef] [PubMed]
- Hadi, Y.B.; Naqvi, S.F.Z.; Kupec, J.T.; Sarwari, A.R. Characteristics and outcomes of COVID-19 in patients with HIV: A multicentre research network study. AIDS 2020, 34, F3–F8. [Google Scholar] [CrossRef]
- Ho, H.E.; Peluso, M.J.; Margus, C.; Matias Lopes, J.P.; He, C.; Gaisa, M.M.; Osorio, G.; Aberg, J.A.; Mullen, M.P. Clinical Outcomes and Immunologic Characteristics of Coronavirus Disease 2019 in People with Human Immunodeficiency Virus. J. Infect. Dis. 2021, 223, 403–408. [Google Scholar] [CrossRef]
- Xu, Y.; Zhuang, Y.; Kang, L. A Review of Neurological Involvement in Patients with SARS-CoV-2 Infection. Med. Sci. Monit. 2021, 27, e932962. [Google Scholar] [CrossRef]
- Guadarrama-Ortiz, P.; Choreno-Parra, J.A.; Sanchez-Martinez, C.M.; Pacheco-Sanchez, F.J.; Rodriguez-Nava, A.I.; Garcia-Quintero, G. Neurological Aspects of SARS-CoV-2 Infection: Mechanisms and Manifestations. Front. Neurol. 2020, 11. [Google Scholar] [CrossRef]
- Spudich, S.; Nath, A. Nervous system consequences of COVID-19. Science 2022, 375, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Maurya, S.K.; Baghel, M.S.; Gaurav Chaudhary, V.; Kaushik, A.; Gautam, A. Putative role of mitochondria in SARS-CoV-2 mediated brain dysfunctions: A prospect. Biotechnol. Genet. Eng. Rev. 2022, 8, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Nepal, G.; Rehrig, J.H.; Shrestha, G.S.; Shing, Y.K.; Yadav, J.K.; Ojha, R.; Pokhrel, G.; Tu, Z.L.; Huang, D.Y. Neurological manifestations of COVID-19: A systematic review. Crit. Care 2020, 24, 421. [Google Scholar] [CrossRef]
- Whittaker, A.; Anson, M.; Harky, A. Neurological Manifestations of COVID-19: A systematic review and current update. Acta Neurol. Scand. 2020, 142, 14–22. [Google Scholar] [CrossRef]
- Gonzalez-Scarano, F.; Martin-Garcia, J. The neuropathogenesis of AIDS. Nat. Rev. Immunol. 2005, 5, 69–81. [Google Scholar] [CrossRef] [PubMed]
- An, S.F.; Groves, M.; Gray, F.; Scaravilli, F. Early entry and widespread cellular involvement of HIV-1 DNA in brains of HIV-1 positive asymptomatic individuals. J. Neuropathol. Exp. Neurol. 1999, 58, 1156–1162. [Google Scholar] [CrossRef]
- Davis, L.E.; Hjelle, B.L.; Miller, V.E.; Palmer, D.L.; Llewellyn, A.L.; Merlin, T.L.; Young, S.A.; Mills, R.G.; Wachsman, W.; Wiley, C.A. Early viral brain invasion in iatrogenic human immunodeficiency virus infection. Neurology 1992, 42, 1736–1739. [Google Scholar] [CrossRef]
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S.; Lu, P.; Weizman, O.E.; Liu, F.; Dai, Y.; et al. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef]
- Olajide, O.A.; Iwuanyanwu, V.U.; Adegbola, O.D.; Al-Hindawi, A.A. SARS-CoV-2 Spike Glycoprotein S1 Induces Neuroinflammation in BV-2 Microglia. Mol. Neurobiol. 2022, 59, 445–458. [Google Scholar] [CrossRef]
- Albornoz, E.A.; Amarilla, A.A.; Modhiran, N.; Parker, S.; Li, X.X.; Wijesundara, D.K.; Aguado, J.; Zamora, A.P.; McMillan, C.L.D.; Liang, B.; et al. SARS-CoV-2 drives NLRP3 inflammasome activation in human microglia through spike protein. Mol. Psychiatry 2022, 1–16. [Google Scholar] [CrossRef]
- Prabhu, S.; Poongulali, S.; Kumarasamy, N. Impact of COVID-19 on people living with HIV: A review. J. Virus Erad. 2020, 6, 100019. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.E.; Fielding, B.C. Insult to Injury-Potential Contribution of Coronavirus Disease-19 to Neuroinflammation and the Development of HIV-Associated Neurocognitive Disorders. AIDS Res. Hum. Retroviruses 2021, 37, 601–609. [Google Scholar] [CrossRef]
- Swanson, P.A., 2nd; McGavern, D.B. Viral diseases of the central nervous system. Curr. Opin. Virol. 2015, 11, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Haidar, M.A.; Shakkour, Z.; Reslan, M.A.; Al-Haj, N.; Chamoun, P.; Habashy, K.; Kaafarani, H.; Shahjouei, S.; Farran, S.H.; Shaito, A.; et al. SARS-CoV-2 involvement in central nervous system tissue damage. Neural Regen. Res. 2022, 17, 1228–1239. [Google Scholar] [CrossRef] [PubMed]
- Saleemi, M.A.; Ahmad, B.; Benchoula, K.; Vohra, M.S.; Mea, H.J.; Chong, P.P.; Palanisamy, N.K.; Wong, E.H. Emergence and molecular mechanisms of SARS-CoV-2 and HIV to target host cells and potential therapeutics. Infect. Genet. Evol. 2020, 85, 104583. [Google Scholar] [CrossRef] [PubMed]
- Fung, S.Y.; Yuen, K.S.; Ye, Z.W.; Chan, C.P.; Jin, D.Y. A tug-of-war between severe acute respiratory syndrome coronavirus 2 and host antiviral defence: Lessons from other pathogenic viruses. Emerg. Microbes Infect. 2020, 9, 558–570. [Google Scholar] [CrossRef]
- Illanes-Alvarez, F.; Marquez-Ruiz, D.; Marquez-Coello, M.; Cuesta-Sancho, S.; Giron-Gonzalez, J.A. Similarities and differences between HIV and SARS-CoV-2. Int. J. Med. Sci. 2021, 18, 846–851. [Google Scholar] [CrossRef]
- Levine, A.; Sacktor, N.; Becker, J.T. Studying the neuropsychological sequelae of SARS-CoV-2: Lessons learned from 35 years of neuroHIV research. J. Neurovirol. 2020, 26, 809–823. [Google Scholar] [CrossRef]
- Burki, T. The origin of SARS-CoV-2 variants of concern. Lancet Infect. Dis. 2022, 22, 174–175. [Google Scholar] [CrossRef]
- Peng, X.; Ouyang, J.; Isnard, S.; Lin, J.; Fombuena, B.; Zhu, B.; Routy, J.P. Sharing CD4+ T Cell Loss: When COVID-19 and HIV Collide on Immune System. Front. Immunol. 2020, 11, 596631. [Google Scholar] [CrossRef]
- Sigalov, A.B. SARS-CoV-2 may affect the immune response via direct inhibition of T cell receptor: Mechanistic hypothesis and rationale. Biochimie 2022, 195, 86–89. [Google Scholar] [CrossRef]
- Cooper, T.J.; Woodward, B.L.; Alom, S.; Harky, A. Coronavirus disease 2019 (COVID-19) outcomes in HIV/AIDS patients: A systematic review. HIV Med. 2020, 21, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.E.; Zink, M.C. Molecular biology and pathogenesis of animal lentivirus infections. Clin. Microbiol. Rev. 1996, 9, 100–117. [Google Scholar] [CrossRef]
- Ho, D.D.; Rota, T.R.; Schooley, R.T.; Kaplan, J.C.; Allan, J.D.; Groopman, J.E.; Resnick, L.; Felsenstein, D.; Andrews, C.A.; Hirsch, M.S. Isolation of HTLV-III from cerebrospinal fluid and neural tissues of patients with neurologic syndromes related to the acquired immunodeficiency syndrome. N. Engl. J. Med. 1985, 313, 1493–1497. [Google Scholar] [CrossRef] [PubMed]
- Robinson-Papp, J.; Saylor, D. HIV in the Brain: From Devastating Dementia to White Matter Hyperintensities. Neurology 2021, 96, 645–646. [Google Scholar] [CrossRef]
- Resnick, L.; Berger, J.R.; Shapshak, P.; Tourtellotte, W.W. Early penetration of the blood-brain-barrier by HIV. Neurology 1988, 38, 9–14. [Google Scholar] [CrossRef]
- Wiley, C.A.; Schrier, R.D.; Nelson, J.A.; Lampert, P.W.; Oldstone, M.B. Cellular localization of human immunodeficiency virus infection within the brains of acquired immune deficiency syndrome patients. Proc. Natl. Acad. Sci. USA 1986, 83, 7089–7093. [Google Scholar] [CrossRef]
- Kumar, A.M.; Fernandez, J.B.; Singer, E.J.; Commins, D.; Waldrop-Valverde, D.; Ownby, R.L.; Kumar, M. Human immunodeficiency virus type 1 in the central nervous system leads to decreased dopamine in different regions of postmortem human brains. J. Neurovirol. 2009, 15, 257–274. [Google Scholar] [CrossRef]
- Persidsky, Y.; Gendelman, H.E. Mononuclear phagocyte immunity and the neuropathogenesis of HIV-1 infection. J. Leukoc. Biol. 2003, 74, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Persidsky, Y.; Stins, M.; Way, D.; Witte, M.H.; Weinand, M.; Kim, K.S.; Bock, P.; Gendelman, H.E.; Fiala, M. A model for monocyte migration through the blood-brain barrier during HIV-1 encephalitis. J. Immunol. 1997, 158, 3499–3510. [Google Scholar] [CrossRef] [PubMed]
- Nottet, H.S.; Persidsky, Y.; Sasseville, V.G.; Nukuna, A.N.; Bock, P.; Zhai, Q.H.; Sharer, L.R.; McComb, R.D.; Swindells, S.; Soderland, C.; et al. Mechanisms for the transendothelial migration of HIV-1-infected monocytes into brain. J. Immunol. 1996, 156, 1284–1295. [Google Scholar] [CrossRef] [PubMed]
- Albright, A.V.; Soldan, S.S.; Gonzalez-Scarano, F. Pathogenesis of human immunodeficiency virus-induced neurological disease. J. Neurovirol. 2003, 9, 222–227. [Google Scholar] [CrossRef]
- Killingsworth, L.; Spudich, S. Neuropathogenesis of HIV-1: Insights from across the spectrum of acute through long-term treated infection. Semin. Immunopathol. 2022, 44, 709–724. [Google Scholar] [CrossRef] [PubMed]
- Clifford, D.B.; Ances, B.M. HIV-associated neurocognitive disorder. Lancet Infect. Dis. 2013, 13, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799. [Google Scholar] [CrossRef]
- Wallet, C.; De Rovere, M.; Van Assche, J.; Daouad, F.; De Wit, S.; Gautier, V.; Mallon, P.W.G.; Marcello, A.; Van Lint, C.; Rohr, O.; et al. Microglial Cells: The Main HIV-1 Reservoir in the Brain. Front. Cell. Infect. Microbiol. 2019, 9, 362. [Google Scholar] [CrossRef]
- Borrajo Lopez, A.; Penedo, M.A.; Rivera-Baltanas, T.; Perez-Rodriguez, D.; Alonso-Crespo, D.; Fernandez-Pereira, C.; Olivares, J.M.; Agis-Balboa, R.C. Microglia: The Real Foe in HIV-1-Associated Neurocognitive Disorders? Biomedicines 2021, 9, 925. [Google Scholar] [CrossRef] [PubMed]
- Nachega, J.B.; Marconi, V.C.; van Zyl, G.U.; Gardner, E.M.; Preiser, W.; Hong, S.Y.; Mills, E.J.; Gross, R. HIV treatment adherence, drug resistance, virologic failure: Evolving concepts. Infect. Disord. Drug. Targets 2011, 11, 167–174. [Google Scholar] [CrossRef] [PubMed]
- De Francesco, D.; Underwood, J.; Post, F.A.; Vera, J.H.; Williams, I.; Boffito, M.; Sachikonye, M.; Anderson, J.; Mallon, P.W.; Winston, A.; et al. Defining cognitive impairment in people-living-with-HIV: The POPPY study. BMC Infect. Dis. 2016, 16, 617. [Google Scholar] [CrossRef]
- Saylor, D.; Dickens, A.M.; Sacktor, N.; Haughey, N.; Slusher, B.; Pletnikov, M.; Mankowski, J.L.; Brown, A.; Volsky, D.J.; McArthur, J.C. HIV-associated neurocognitive disorder—Pathogenesis and prospects for treatment. Nat. Rev. Neurol. 2016, 12, 309. [Google Scholar] [CrossRef]
- Eggers, C.; Arendt, G.; Hahn, K.; Husstedt, I.W.; Maschke, M.; Neuen-Jacob, E.; Obermann, M.; Rosenkranz, T.; Schielke, E.; Straube, E.; et al. HIV-1-associated neurocognitive disorder: Epidemiology, pathogenesis, diagnosis, and treatment. J. Neurol. 2017, 264, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Al-Harthi, L.; Campbell, E.; Schneider, J.A.; Bennett, D.A. What HIV in the Brain Can Teach Us About SARS-CoV-2 Neurological Complications? AIDS Res. Hum. Retrovir. 2021, 37, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Machhi, J.; Herskovitz, J.; Senan, A.M.; Dutta, D.; Nath, B.; Oleynikov, M.D.; Blomberg, W.R.; Meigs, D.D.; Hasan, M.; Patel, M.; et al. The Natural History, Pathobiology, and Clinical Manifestations of SARS-CoV-2 Infections. J. Neuroimmune Pharmacol. 2020, 15, 359–386. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.; Martinez, E.; Petrosillo, N.; Nichols, J.; Islam, E.; Pruitt, K.; Almodovar, S. SARS-CoV-2 and Human Immunodeficiency Virus: Pathogen Pincer Attack. HIV AIDS 2021, 13, 361–375. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus-Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Liotta, E.M.; Batra, A.; Clark, J.R.; Shlobin, N.A.; Hoffman, S.C.; Orban, Z.S.; Koralnik, I.J. Frequent neurologic manifestations and encephalopathy-associated morbidity in COVID-19 patients. Ann. Clin. Transl. Neurol. 2020, 7, 2221–2230. [Google Scholar] [CrossRef]
- Puelles, V.G.; Lutgehetmann, M.; Lindenmeyer, M.T.; Sperhake, J.P.; Wong, M.N.; Allweiss, L.; Chilla, S.; Heinemann, A.; Wanner, N.; Liu, S.; et al. Multiorgan and Renal Tropism of SARS-CoV-2. N. Engl. J. Med. 2020, 383, 590–592. [Google Scholar] [CrossRef]
- Solomon, I.H.; Normandin, E.; Bhattacharyya, S.; Mukerji, S.S.; Keller, K.; Ali, A.S.; Adams, G.; Hornick, J.L.; Padera, R.F., Jr.; Sabeti, P. Neuropathological Features of COVID-19. N. Engl. J. Med. 2020, 383, 989–992. [Google Scholar] [CrossRef]
- Moriguchi, T.; Harii, N.; Goto, J.; Harada, D.; Sugawara, H.; Takamino, J.; Ueno, M.; Sakata, H.; Kondo, K.; Myose, N.; et al. A first case of meningitis/encephalitis associated with SARS-Coronavirus-2. Int. J. Infect. Dis. 2020, 94, 55–58. [Google Scholar] [CrossRef]
- Matschke, J.; Lutgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schroder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef]
- Rutkai, I.; Mayer, M.G.; Hellmers, L.M.; Ning, B.; Huang, Z.; Monjure, C.J.; Coyne, C.; Silvestri, R.; Golden, N.; Hensley, K.; et al. Neuropathology and virus in brain of SARS-CoV-2 infected non-human primates. Nat. Commun. 2022, 13, 1745. [Google Scholar] [CrossRef] [PubMed]
- Bullen, C.K.; Hogberg, H.T.; Bahadirli-Talbott, A.; Bishai, W.R.; Hartung, T.; Keuthan, C.; Looney, M.M.; Pekosz, A.; Romero, J.C.; Sille, F.C.M.; et al. Infectability of human BrainSphere neurons suggests neurotropism of SARS-CoV-2. ALTEX 2020, 37, 665–671. [Google Scholar] [CrossRef]
- Jacob, F.; Pather, S.R.; Huang, W.K.; Zhang, F.; Wong, S.Z.H.; Zhou, H.; Cubitt, B.; Fan, W.; Chen, C.Z.; Xu, M.; et al. Human Pluripotent Stem Cell-Derived Neural Cells and Brain Organoids Reveal SARS-CoV-2 Neurotropism Predominates in Choroid Plexus Epithelium. Cell. Stem Cell. 2020, 27, 937–950 e939. [Google Scholar] [CrossRef]
- Pellegrini, L.; Bonfio, C.; Chadwick, J.; Begum, F.; Skehel, M.; Lancaster, M.A. Human CNS barrier-forming organoids with cerebrospinal fluid production. Science 2020, 369, eaaz5626. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Jin, H.; Wang, M.; Hu, Y.; Chen, S.; He, Q.; Chang, J.; Hong, C.; Zhou, Y.; Wang, D.; et al. Neurologic Manifestations of Hospitalized Patients with Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol. 2020, 77, 683–690. [Google Scholar] [CrossRef]
- Khan, S.; Gomes, J. Neuropathogenesis of SARS-CoV-2 infection. Elife 2020, 9, e59136. [Google Scholar] [CrossRef] [PubMed]
- Deigendesch, N.; Stenzel, W. Acute and chronic viral infections. Handb. Clin. Neurol. 2017, 145, 227–243. [Google Scholar] [CrossRef]
- Solomon, T. Neurological infection with SARS-CoV-2—The story so far. Nat. Rev. Neurol. 2021, 17, 65–66. [Google Scholar] [CrossRef]
- Brann, D.H.; Tsukahara, T.; Weinreb, C.; Lipovsek, M.; Van den Berge, K.; Gong, B.; Chance, R.; Macaulay, I.C.; Chou, H.J.; Fletcher, R.B.; et al. Non-neuronal expression of SARS-CoV-2 entry genes in the olfactory system suggests mechanisms underlying COVID-19-associated anosmia. Sci. Adv. 2020, 6, eabc5801. [Google Scholar] [CrossRef]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brunink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Seo, G.; Lee, G.; Kim, M.J.; Baek, S.H.; Choi, M.; Ku, K.B.; Lee, C.S.; Jun, S.; Park, D.; Kim, H.G.; et al. Rapid Detection of COVID-19 Causative Virus (SARS-CoV-2) in Human Nasopharyngeal Swab Specimens Using Field-Effect Transistor-Based Biosensor. ACS Nano 2020, 14, 5135–5142. [Google Scholar] [CrossRef]
- Dutta, D.; Naiyer, S.; Mansuri, S.; Soni, N.; Singh, V.; Bhat, K.H.; Singh, N.; Arora, G.; Mansuri, M.S. COVID-19 Diagnosis: A Comprehensive Review of the RT-qPCR Method for Detection of SARS-CoV-2. Diagnostics 2022, 12, 1503. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell. 2020, 181, 271–280 e278. [Google Scholar] [CrossRef]
- Doobay, M.F.; Talman, L.S.; Obr, T.D.; Tian, X.; Davisson, R.L.; Lazartigues, E. Differential expression of neuronal ACE2 in transgenic mice with overexpression of the brain renin-angiotensin system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R373–R381. [Google Scholar] [CrossRef] [PubMed]
- Netland, J.; Meyerholz, D.K.; Moore, S.; Cassell, M.; Perlman, S. Severe acute respiratory syndrome coronavirus infection causes neuronal death in the absence of encephalitis in mice transgenic for human ACE2. J. Virol. 2008, 82, 7264–7275. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, S.; Chua, R.L.; Trefzer, T.; Kahn, N.C.; Schneider, M.A.; Muley, T.; Winter, H.; Meister, M.; Veith, C.; Boots, A.W.; et al. SARS-CoV-2 receptor ACE2 and TMPRSS2 are primarily expressed in bronchial transient secretory cells. EMBO J. 2020, 39, e105114. [Google Scholar] [CrossRef]
- Butowt, R.; Bilinska, K. SARS-CoV-2: Olfaction, Brain Infection, and the Urgent Need for Clinical Samples Allowing Earlier Virus Detection. ACS Chem. Neurosci. 2020, 11, 1200–1203. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef]
- Kumari, P.; Rothan, H.A.; Natekar, J.P.; Stone, S.; Pathak, H.; Strate, P.G.; Arora, K.; Brinton, M.A.; Kumar, M. Neuroinvasion and Encephalitis Following Intranasal Inoculation of SARS-CoV-2 in K18-hACE2 Mice. Viruses 2021, 13, 132. [Google Scholar] [CrossRef]
- Jiao, L.; Yang, Y.; Yu, W.; Zhao, Y.; Long, H.; Gao, J.; Ding, K.; Ma, C.; Li, J.; Zhao, S.; et al. The olfactory route is a potential way for SARS-CoV-2 to invade the central nervous system of rhesus monkeys. Signal. Transduct. Target. Ther. 2021, 6, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Kang, H.; Li, S.; Zhao, X. Understanding the neurotropic characteristics of SARS-CoV-2: From neurological manifestations of COVID-19 to potential neurotropic mechanisms. J. Neurol. 2020, 267, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Zingaropoli, M.A.; Iannetta, M.; Piermatteo, L.; Pasculli, P.; Latronico, T.; Mazzuti, L.; Campogiani, L.; Duca, L.; Ferraguti, G.; De Michele, M.; et al. Neuro-Axonal Damage and Alteration of Blood-Brain Barrier Integrity in COVID-19 Patients. Cells 2022, 11, 2480. [Google Scholar] [CrossRef] [PubMed]
- Kielian, T. Toll-like receptors in central nervous system glial inflammation and homeostasis. J. Neurosci. Res. 2006, 83, 711–730. [Google Scholar] [CrossRef]
- Louveau, A.; Harris, T.H.; Kipnis, J. Revisiting the Mechanisms of CNS Immune Privilege. Trends Immunol. 2015, 36, 569–577. [Google Scholar] [CrossRef]
- Peluso, R.; Haase, A.; Stowring, L.; Edwards, M.; Ventura, P. A Trojan Horse mechanism for the spread of visna virus in monocytes. Virology 1985, 147, 231–236. [Google Scholar] [CrossRef]
- Haase, A.T. Pathogenesis of lentivirus infections. Nature 1986, 322, 130–136. [Google Scholar] [CrossRef]
- Aljawai, Y.; Richards, M.H.; Seaton, M.S.; Narasipura, S.D.; Al-Harthi, L. β-Catenin/TCF-4 signaling regulates susceptibility of macrophages and resistance of monocytes to HIV-1 productive infection. Curr. HIV Res. 2014, 12, 164–173. [Google Scholar] [CrossRef]
- Xu, J.; Zhong, S.; Liu, J.; Li, L.; Li, Y.; Wu, X.; Li, Z.; Deng, P.; Zhang, J.; Zhong, N.; et al. Detection of severe acute respiratory syndrome coronavirus in the brain: Potential role of the chemokine mig in pathogenesis. Clin. Infect. Dis. 2005, 41, 1089–1096. [Google Scholar] [CrossRef]
- Brewer, R.C.; Robinson, W.H.; Lanz, T.V. SARS-CoV-2 infection of monocytes: Balancing acts of antibodies and inflammasomes. Signal. Transduct. Target. Ther. 2022, 7, 250. [Google Scholar] [CrossRef] [PubMed]
- Padovani, A.; Pilotto, A. Looking for lights in the fog of long-term neurological COVID. Nat. Rev. Neurol. 2023, 19, 7–8. [Google Scholar] [CrossRef]
- Xu, E.; Xie, Y.; Al-Aly, Z. Long-term neurologic outcomes of COVID-19. Nat. Med. 2022, 28, 2406–2415. [Google Scholar] [CrossRef]
- Davis, H.E.; McCorkell, L.; Vogel, J.M.; Topol, E.J. Long COVID: Major findings, mechanisms and recommendations. Nat. Rev. Microbiol. 2023, 21, 133–146. [Google Scholar] [CrossRef]
- Gao, W.; Lv, J.; Pang, Y.; Li, L.M. Role of asymptomatic and pre-symptomatic infections in COVID-19 pandemic. BMJ 2021, 375, n2342. [Google Scholar] [CrossRef]
- Huang, L.; Li, X.; Gu, X.; Zhang, H.; Ren, L.; Guo, L.; Liu, M.; Wang, Y.; Cui, D.; Wang, Y.; et al. Health outcomes in people 2 years after surviving hospitalisation with COVID-19: A longitudinal cohort study. Lancet Respir. Med. 2022, 10, 863–876. [Google Scholar] [CrossRef]
- The UNAIDS Global AIDS Update 2021. UNAIDS Report Shows That People Living with HIV Face a Double Jeopardy, HIV and COVID-19, While Key Populations and Children Continue to Be Left behind in Access to HIV Services. Available online: https://www.unaids.org/en/resources/presscentre/pressreleaseandstatementarchive/2021/july/20210714_global-aids-update (accessed on 7 December 2022).
- Wang, Y.; Xie, Y.; Hu, S.; Ai, W.; Tao, Y.; Tang, H.; Jing, F.; Tang, W. Systematic Review and Meta-Analyses of THE Interaction between HIV Infection and COVID-19: Two Years’ Evidence Summary. Front. Immunol. 2022, 13, 864838. [Google Scholar] [CrossRef]
- Spinelli, M.A.; Jones, B.L.H.; Gandhi, M. COVID-19 Outcomes and Risk Factors among People Living with HIV. Curr. HIV/AIDS Rep. 2022, 19, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Varshney, K.; Ghosh, P.; Stiles, H.; Iriowen, R. Risk Factors for COVID-19 Mortality among People Living with HIV: A Scoping Review. AIDS Behav. 2022, 26, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Antiretroviral Therapy Cohort, C. Life expectancy of individuals on combination antiretroviral therapy in high-income countries: A collaborative analysis of 14 cohort studies. Lancet 2008, 372, 293–299. [Google Scholar] [CrossRef]
- Mills, E.J.; Bakanda, C.; Birungi, J.; Chan, K.; Ford, N.; Cooper, C.L.; Nachega, J.B.; Dybul, M.; Hogg, R.S. Life expectancy of persons receiving combination antiretroviral therapy in low-income countries: A cohort analysis from Uganda. Ann. Intern. Med. 2011, 155, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Samji, H.; Cescon, A.; Hogg, R.S.; Modur, S.P.; Althoff, K.N.; Buchacz, K.; Burchell, A.N.; Cohen, M.; Gebo, K.A.; Gill, M.J.; et al. Closing the gap: Increases in life expectancy among treated HIV-positive individuals in the United States and Canada. PLoS ONE 2013, 8, e81355. [Google Scholar] [CrossRef]
- Katz, I.T.; Maughan-Brown, B. Improved life expectancy of people living with HIV: Who is left behind? Lancet HIV 2017, 4, e324–e326. [Google Scholar] [CrossRef] [PubMed]
- Marcus, J.L.; Leyden, W.A.; Alexeeff, S.E.; Anderson, A.N.; Hechter, R.C.; Hu, H.; Lam, J.O.; Towner, W.J.; Yuan, Q.; Horberg, M.A.; et al. Comparison of Overall and Comorbidity-Free Life Expectancy Between Insured Adults with and Without HIV Infection, 2000–2016. JAMA Netw. Open. 2020, 3, e207954. [Google Scholar] [CrossRef] [PubMed]
- Bachelard, A.; Sautereau, A.; Digumber, M.; Isernia, V.; Phung, B.; Lehur, A.C.; Gac, S.L.; Landman, R.; Yazdanpanah, Y.; Ghosn, J. Risk Factors Associated with Severe/Critical COVID-19 in People Living with HIV-1. Int. J. Infect. Dis. 2022, 122, 152–154. [Google Scholar] [CrossRef]
- Montano, M. Pressing Questions and Challenges in the HIV-1 and SARS-CoV-2 Syndemic. AIDS Res. Hum. Retrovir. 2021, 37, 589–600. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Risk for COVID-19 Infection, Hospitalization, and Death by Age Group. Available online: https://www.cdc.gov/coronavirus/2019-ncov/COVID-data/investigations-discovery/hospitalization-death-by-age.html (accessed on 14 February 2023).
- Centers for Disease Control and Prevention. HIV and COVID-19 Basics. Available online: https://www.cdc.gov/hiv/basics/COVID-19.html (accessed on 14 February 2023).
- Schrack, J.A.; Jacobson, L.P.; Althoff, K.N.; Erlandson, K.M.; Jamieson, B.D.; Koletar, S.L.; Phair, J.; Brown, T.T.; Margolick, J.B.; Multicenter, A.C.S. Effect of HIV-infection and cumulative viral load on age-related decline in grip strength. AIDS 2016, 30, 2645–2652. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, D.P.; Iudicello, J.E.; Bondi, M.W.; Doyle, K.L.; Morgan, E.E.; Massman, P.J.; Gilbert, P.E.; Woods, S.P. Elevated rates of mild cognitive impairment in HIV disease. J. Neurovirol. 2015, 21, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Deeks, S.G.; Tracy, R.; Douek, D.C. Systemic effects of inflammation on health during chronic HIV infection. Immunity 2013, 39, 633–645. [Google Scholar] [CrossRef]
- Guaraldi, G.; Milic, J.; Martinez, E.; Kamarulzaman, A.; Mussini, C.; Waters, L.; Pozniak, A.; Mallon, P.; Rockstroh, J.K.; Lazarus, J.V. Human Immunodeficiency Virus (HIV) Care Models During the Coronavirus Disease 2019 (COVID-19) Era. Clin. Infect. Dis. 2021, 73, e1222–e1227. [Google Scholar] [CrossRef]
- Premeaux, T.A.; Javandel, S.; Hosaka, K.R.J.; Greene, M.; Therrien, N.; Allen, I.E.; Corley, M.J.; Valcour, V.G.; Ndhlovu, L.C. Associations Between Plasma Immunomodulatory and Inflammatory Mediators with VACS Index Scores Among Older HIV-Infected Adults on Antiretroviral Therapy. Front. Immunol. 2020, 11, 1321. [Google Scholar] [CrossRef] [PubMed]
- Boukhvalova, M.S.; Mortensen, E.; Caple, J.; Joseph, J.; Sylla, F.; Kamali, A.; Stylos, D.; Lopez, D.; March, T.; Byrd, K.M.; et al. SARS-CoV-2 infection augments species- and age-specific predispositions in cotton rats. Sci. Rep. 2023, 13, 757. [Google Scholar] [CrossRef] [PubMed]
- Scully, E.P.; Haverfield, J.; Ursin, R.L.; Tannenbaum, C.; Klein, S.L. Considering how biological sex impacts immune responses and COVID-19 outcomes. Nat. Rev. Immunol. 2020, 20, 442–447. [Google Scholar] [CrossRef]
- Nielsen, J.; Norgaard, S.K.; Lanzieri, G.; Vestergaard, L.S.; Moelbak, K. Sex-differences in COVID-19 associated excess mortality is not exceptional for the COVID-19 pandemic. Sci. Rep. 2021, 11, 20815. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Volgman, A.S.; Michos, E.D. Sex Differences in Mortality From COVID-19 Pandemic: Are Men Vulnerable and Women Protected? JACC Case Rep. 2020, 2, 1407–1410. [Google Scholar] [CrossRef]
- Bhopal, S.S.; Bhopal, R. Sex differential in COVID-19 mortality varies markedly by age. Lancet 2020, 396, 532–533. [Google Scholar] [CrossRef]
- Capuano, A.; Rossi, F.; Paolisso, G. COVID-19 Kills More Men Than Women: An Overview of Possible Reasons. Front. Cardiovasc. Med. 2020, 7, 131. [Google Scholar] [CrossRef]
- Takahashi, T.; Ellingson, M.K.; Wong, P.; Israelow, B.; Lucas, C.; Klein, J.; Silva, J.; Mao, T.; Oh, J.E.; Tokuyama, M.; et al. Sex differences in immune responses that underlie COVID-19 disease outcomes. Nature 2020, 588, 315–320. [Google Scholar] [CrossRef]
- Gupta, S.; Nakabo, S.; Blanco, L.P.; O’Neil, L.J.; Wigerblad, G.; Goel, R.R.; Mistry, P.; Jiang, K.; Carmona-Rivera, C.; Chan, D.W.; et al. Sex differences in neutrophil biology modulate response to type I interferons and immunometabolism. Proc. Natl. Acad. Sci. USA 2020, 117, 16481–16491. [Google Scholar] [CrossRef]
- Salah, H.M.; Mehta, J.L. Hypothesis: Sex-Related Differences in ACE2 Activity May Contribute to Higher Mortality in Men Versus Women With COVID-19. J. Cardiovasc. Pharmacol. Ther. 2021, 26, 114–118. [Google Scholar] [CrossRef]
- Haitao, T.; Vermunt, J.V.; Abeykoon, J.; Ghamrawi, R.; Gunaratne, M.; Jayachandran, M.; Narang, K.; Parashuram, S.; Suvakov, S.; Garovic, V.D. COVID-19 and Sex Differences: Mechanisms and Biomarkers. Mayo Clin. Proc. 2020, 95, 2189–2203. [Google Scholar] [CrossRef]
- Samuel, R.M.; Majd, H.; Richter, M.N.; Ghazizadeh, Z.; Zekavat, S.M.; Navickas, A.; Ramirez, J.T.; Asgharian, H.; Simoneau, C.R.; Bonser, L.R.; et al. Androgen Signaling Regulates SARS-CoV-2 Receptor Levels and Is Associated with Severe COVID-19 Symptoms in Men. Cell. Stem Cell. 2020, 27, 876–889 e812. [Google Scholar] [CrossRef] [PubMed]
- Leach, D.A.; Mohr, A.; Giotis, E.S.; Cil, E.; Isac, A.M.; Yates, L.L.; Barclay, W.S.; Zwacka, R.M.; Bevan, C.L.; Brooke, G.N. The antiandrogen enzalutamide downregulates TMPRSS2 and reduces cellular entry of SARS-CoV-2 in human lung cells. Nat. Commun. 2021, 12, 4068. [Google Scholar] [CrossRef]
- Rastrelli, G.; Di Stasi, V.; Inglese, F.; Beccaria, M.; Garuti, M.; Di Costanzo, D.; Spreafico, F.; Greco, G.F.; Cervi, G.; Pecoriello, A.; et al. Low testosterone levels predict clinical adverse outcomes in SARS-CoV-2 pneumonia patients. Andrology 2021, 9, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Beltrame, A.; Salguero, P.; Rossi, E.; Conesa, A.; Moro, L.; Bettini, L.R.; Rizzi, E.; D’Angio, M.; Deiana, M.; Piubelli, C.; et al. Association Between Sex Hormone Levels and Clinical Outcomes in Patients With COVID-19 Admitted to Hospital: An Observational, Retrospective, Cohort Study. Front. Immunol. 2022, 13, 834851. [Google Scholar] [CrossRef]
- Grandys, M.; Majerczak, J.; Zapart-Bukowska, J.; Duda, K.; Kulpa, J.K.; Zoladz, J.A. Lowered Serum Testosterone Concentration Is Associated with Enhanced Inflammation and Worsened Lipid Profile in Men. Front. Endocrinol. 2021, 12, 735638. [Google Scholar] [CrossRef] [PubMed]
- Rochira, V.; Guaraldi, G. Hypogonadism in the HIV-infected man. Endocrinol. Metab. Clin. North. Am. 2014, 43, 709–730. [Google Scholar] [CrossRef]
- Hogg, R.S.; Eyawo, O.; Collins, A.B.; Zhang, W.; Jabbari, S.; Hull, M.W.; Lima, V.D.; Ahmed, T.; Kendall, C.E.; Althoff, K.N.; et al. Health-adjusted life expectancy in HIV-positive and HIV-negative men and women in British Columbia, Canada: A population-based observational cohort study. Lancet HIV 2017, 4, e270–e276. [Google Scholar] [CrossRef]
- Guaraldi, G.; Silva, A.R.; Stentarelli, C. Multimorbidity and functional status assessment. Curr. Opin. HIV AIDS 2014, 9, 386–397. [Google Scholar] [CrossRef]
- Krebs, S.J.; Slike, B.M.; Sithinamsuwan, P.; Allen, I.E.; Chalermchai, T.; Tipsuk, S.; Phanuphak, N.; Jagodzinski, L.; Kim, J.H.; Ananworanich, J.; et al. Sex differences in soluble markers vary before and after the initiation of antiretroviral therapy in chronically HIV-infected individuals. AIDS 2016, 30, 1533–1542. [Google Scholar] [CrossRef]
- Skrzynski, A.K.; Darmstadter, B.L.; Aahivp Miner, S.P.; Agpcnp, B.C.; Subedi, K.; Kahal, D. Characteristics and Outcomes of SARS-CoV-2 Infection Among Adults Living with HIV In Delaware: The Story of a Syndemic During the First 12 Months of the SARS-CoV-2 Pandemic. Dela J. Public Health 2021, 7, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, M.E.; Ross, P.; Lasso, M.; Dominguez, I.; Puente, M.; Valenzuela, P.; Enberg, M.; Serri, M.; Munoz, R.; Pinos, Y.; et al. Clinical characteristics and outcomes of people living with HIV hospitalized with COVID-19: A nationwide experience. Int. J. STD AIDS 2021, 32, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Lorenc, A.; Ananthavarathan, P.; Lorigan, J.; Jowata, M.; Brook, G.; Banarsee, R. The prevalence of comorbidities among people living with HIV in Brent: A diverse London Borough. London J. Prim. Care 2014, 6, 84–90. [Google Scholar] [CrossRef]
- Shaikh, F.S.; Aldhafferi, N.; Buker, A.; Alqahtani, A.; Dey, S.; Abdulhamid, S.; AlBuhairi, D.A.M.; Alkabour, R.S.A.; Atiyah, W.S.O.; Bachar Chrouf, S.; et al. Comorbidities and Risk Factors for Severe Outcomes in COVID-19 Patients in Saudi Arabia: A Retrospective Cohort Study. J. Multidiscip. Healthc. 2021, 14, 2169–2183. [Google Scholar] [CrossRef] [PubMed]
- MacNeill, J.J.; Linnes, J.C.; Hubach, R.D.; Rodriguez, N.M. From crisis to crisis: Impacts of the COVID-19 pandemic on people living with HIV and HIV/AIDS service organizations in Indiana. BMC Health Serv. Res. 2022, 22, 622. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yang, Q.; Chi, J.; Dong, B.; Lv, W.; Shen, L.; Wang, Y. Comorbidities and the risk of severe or fatal outcomes associated with coronavirus disease 2019: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 99, 47–56. [Google Scholar] [CrossRef]
- Augello, M.; Bono, V.; Rovito, R.; Tincati, C.; Marchetti, G. Immunologic Interplay Between HIV/AIDS and COVID-19: Adding Fuel to the Flames? Curr. HIV/AIDS Rep. 2023, 20, 51–75. [Google Scholar] [CrossRef]
- Mazzitelli, M.; Trunfio, M.; Sasset, L.; Leoni, D.; Castelli, E.; Lo Menzo, S.; Gardin, S.; Putaggio, C.; Brundu, M.; Garzotto, P.; et al. Factors Associated with Severe COVID-19 and Post-Acute COVID-19 Syndrome in a Cohort of People Living with HIV on Antiretroviral Treatment and with Undetectable HIV RNA. Viruses 2022, 14, 493. [Google Scholar] [CrossRef]
- Kerneis, S.; Launay, O.; Turbelin, C.; Batteux, F.; Hanslik, T.; Boelle, P.Y. Long-term immune responses to vaccination in HIV-infected patients: A systematic review and meta-analysis. Clin. Infect. Dis. 2014, 58, 1130–1139. [Google Scholar] [CrossRef]
- Elrefaei, M.; McElroy, M.D.; Preas, C.P.; Hoh, R.; Deeks, S.; Martin, J.; Cao, H. Central memory CD4+ T cell responses in chronic HIV infection are not restored by antiretroviral therapy. J. Immunol. 2004, 173, 2184–2189. [Google Scholar] [CrossRef]
- Thomas, A.; Hammarlund, E.; Gao, L.; Holman, S.; Michel, K.G.; Glesby, M.; Villacres, M.C.; Golub, E.T.; Roan, N.R.; French, A.L.; et al. Loss of Preexisting Immunological Memory Among Human Immunodeficiency Virus-Infected Women Despite Immune Reconstitution with Antiretroviral Therapy. J. Infect. Dis. 2020, 222, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Del Amo, J.; Polo, R.; Moreno, S.; Diaz, A.; Martinez, E.; Arribas, J.R.; Jarrin, I.; Hernan, M.A.; The Spanish HIV/COVID-19 Collaboration. Incidence and Severity of COVID-19 in HIV-Positive Persons Receiving Antiretroviral Therapy: A Cohort Study. Ann. Intern. Med. 2020, 173, 536–541. [Google Scholar] [CrossRef]
- Johnston, R. The first 6 months of HIV-SARS-CoV-2 coinfection: Outcomes for 6947 individuals. Curr. Opin. HIV AIDS 2021, 16, 54–62. [Google Scholar] [CrossRef]
- Freer, J.; Mudaly, V. HIV and COVID-19 in South Africa. BMJ 2022, 376, e069807. [Google Scholar] [CrossRef]
- Pellowski, J.A.; Kalichman, S.C.; Matthews, K.A.; Adler, N. A pandemic of the poor: Social disadvantage and the U.S. HIV epidemic. Am. Psychol. 2013, 68, 197–209. [Google Scholar] [CrossRef]
- Czeisler, M.E.; Lane, R.I.; Petrosky, E.; Wiley, J.F.; Christensen, A.; Njai, R.; Weaver, M.D.; Robbins, R.; Facer-Childs, E.R.; Barger, L.K.; et al. Mental Health, Substance Use, and Suicidal Ideation During the COVID-19 Pandemic—United States, June 24-30, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Hochstatter, K.R.; Akhtar, W.Z.; Dietz, S.; Pe-Romashko, K.; Gustafson, D.H.; Shah, D.V.; Krechel, S.; Liebert, C.; Miller, R.; El-Bassel, N.; et al. Potential Influences of the COVID-19 Pandemic on Drug Use and HIV Care Among People Living with HIV and Substance Use Disorders: Experience from a Pilot mHealth Intervention. AIDS Behav. 2021, 25, 354–359. [Google Scholar] [CrossRef]
- Board, A.R.; Kim, S.; Park, J.; Schieber, L.; Miller, G.F.; Pike, J.; Cremer, L.J.; Asher, A. Risk factors for COVID-19 among persons with substance use disorder (PWSUD) with hospital visits—United States, April 2020-December 2020. Drug. Alcohol. Depend. 2022, 232, 109297. [Google Scholar] [CrossRef] [PubMed]
- Melamed, O.C.; Hauck, T.S.; Buckley, L.; Selby, P.; Mulsant, B.H. COVID-19 and persons with substance use disorders: Inequities and mitigation strategies. Subst. Abus. 2020, 41, 286–291. [Google Scholar] [CrossRef]
- Vasylyeva, T.I.; Smyrnov, P.; Strathdee, S.; Friedman, S.R. Challenges posed by COVID-19 to people who inject drugs and lessons from other outbreaks. J. Int. AIDS Soc. 2020, 23, e25583. [Google Scholar] [CrossRef]
- Holloway, I.W.; Spaulding, A.C.; Ochoa, A.M.; Randall, L.A.; King, A.R.; Frew, P.M.; Hbou Study Team. COVID-19 vulnerability among people who use drugs: Recommendations for global public health programmes and policies. J. Int. AIDS Soc. 2020, 23, e25551. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Kaelber, D.C.; Xu, R.; Volkow, N.D. COVID-19 risk and outcomes in patients with substance use disorders: Analyses from electronic health records in the United States. Mol. Psychiatry 2021, 26, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Baillargeon, J.; Polychronopoulou, E.; Kuo, Y.F.; Raji, M.A. The Impact of Substance Use Disorder on COVID-19 Outcomes. Psychiatr. Serv. 2021, 72, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, J.; Manini, A.F. Opioid Use Disorder and COVID-19: Biological Plausibility for Worsened Outcomes. Subst. Use Misuse 2020, 55, 1900–1901. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, D.; Sureshanand, S.; Pittman, B.; Radhakrishnan, R. Impact of Cannabis Use, Substance Use Disorders, and Psychiatric Diagnoses on COVID-19 Outcomes: A Retrospective Cohort Study. J. Clin. Psychiatry 2022, 83, 21m14332. [Google Scholar] [CrossRef]
- Compton, W.M.; Jones, C.M. Substance Use among Men Who Have Sex with Men. N. Engl. J. Med. 2021, 385, 352–356. [Google Scholar] [CrossRef]
- Fletcher, J.B.; Swendeman, D.; Reback, C.J. Mental Health and Substance Use Disorder Comorbidity among Methamphetamine-Using Men Who have Sex with Men. J. Psychoact. Drugs 2018, 50, 206–213. [Google Scholar] [CrossRef]
- Cohen, J.M.; Li, M.J.; Javanbakht, M.; Gorbach, P.M.; Shoptaw, S.J. Methamphetamine use and adoption of preventive behaviors early in the COVID-19 pandemic among men who have sex with men in Los Angeles, California. Drug. Alcohol. Depend. Rep. 2022, 5, 100097. [Google Scholar] [CrossRef]
- Kanwugu, O.N.; Adadi, P. HIV/SARS-CoV-2 coinfection: A global perspective. J. Med. Virol. 2021, 93, 726–732. [Google Scholar] [CrossRef]
- D’Souza, G.; Tong, W.; Gustafson, D.; Alcaide, M.L.; Lahiri, C.D.; Sharma, A.; French, A.L.; Palella, F.J.; Kempf, M.C.; Mimiaga, M.J.; et al. SARS-CoV-2 Infection Among People Living with HIV Compared With People Without HIV: Survey Results From the MACS-WIHS Combined Cohort Study. J. Acquir. Immune Defic. Syndr. 2022, 89, 1–8. [Google Scholar] [CrossRef]
- Barbera, L.K.; Kamis, K.F.; Rowan, S.E.; Davis, A.J.; Shehata, S.; Carlson, J.J.; Johnson, S.C.; Erlandson, K.M. HIV and COVID-19: Review of clinical course and outcomes. HIV Res. Clin. Pract. 2021, 22, 102–118. [Google Scholar] [CrossRef]
- Ssentongo, P.; Heilbrunn, E.S.; Ssentongo, A.E.; Advani, S.; Chinchilli, V.M.; Nunez, J.J.; Du, P. Epidemiology and outcomes of COVID-19 in HIV-infected individuals: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 6283. [Google Scholar] [CrossRef]
- Maggiolo, F.; Zoboli, F.; Arosio, M.; Valenti, D.; Guarneri, D.; Sangiorgio, L.; Ripamonti, D.; Callegaro, A. SARS-CoV-2 infection in persons living with HIV: A single center prospective cohort. J. Med. Virol. 2021, 93, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- d’Ettorre, G.; Recchia, G.; Ridolfi, M.; Siccardi, G.; Pinacchio, C.; Innocenti, G.P.; Santinelli, L.; Frasca, F.; Bitossi, C.; Ceccarelli, G.; et al. Analysis of type I IFN response and T cell activation in severe COVID-19/HIV-1 coinfection: A case report. Medicine 2020, 99, e21803. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.K.; Arora, S.; Ashta, K.; Singh, T. Clinical outcome of patients with HIV/SARS-CoV-2 co-infection. Med. J. Armed Forces India 2022. ahead of print. [Google Scholar] [CrossRef]
- Nagarakanti, S.R.; Okoh, A.K.; Grinberg, S.; Bishburg, E. Clinical outcomes of patients with COVID-19 and HIV coinfection. J. Med. Virol. 2021, 93, 1687–1693. [Google Scholar] [CrossRef]
- Akyala, A.I.; Iwu, C.J. Novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) co-infection with HIV: Clinical case series analysis in North Central Nigeria. Pan Afr. Med. J. 2020, 37, 47. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.L.; Ambrosioni, J.; Garcia, F.; Martinez, E.; Soriano, A.; Mallolas, J.; Miro, J.M.; on behaf of the COVID-19 in HIV Investigators. COVID-19 in patients with HIV: Clinical case series. Lancet HIV 2020, 7, e314–e316. [Google Scholar] [CrossRef] [PubMed]
- Diao, B.; Wang, C.; Tan, Y.; Chen, X.; Liu, Y.; Ning, L.; Chen, L.; Li, M.; Liu, Y.; Wang, G.; et al. Reduction and Functional Exhaustion of T Cells in Patients with Coronavirus Disease 2019 (COVID-19). Front. Immunol. 2020, 11, 827. [Google Scholar] [CrossRef]
- Amimo, F.; Lambert, B.; Magit, A. What does the COVID-19 pandemic mean for HIV, tuberculosis, and malaria control? Trop. Med. Health 2020, 48, 32. [Google Scholar] [CrossRef]
- Pinto, R.M.; Park, S. COVID-19 Pandemic Disrupts HIV Continuum of Care and Prevention: Implications for Research and Practice Concerning Community-Based Organizations and Frontline Providers. AIDS Behav. 2020, 24, 2486–2489. [Google Scholar] [CrossRef] [PubMed]
- Adadi, P.; Kanwugu, O.N. Living with HIV in the time of COVID-19: A glimpse of hope. J. Med. Virol. 2021, 93, 59–60. [Google Scholar] [CrossRef] [PubMed]
- The Lancet, H. Lockdown fears for key populations. Lancet HIV 2020, 7, e373. [Google Scholar] [CrossRef]
- Guo, W.; Weng, H.L.; Bai, H.; Liu, J.; Wei, X.; Zhou, K.; Amakobe, S. A rapid assessment of the impact of the new coronavirus pneumonia epidemic on the health needs of HIV-infected patients. Chin. J. Epidemiol. 2020, 41, 662–666. [Google Scholar]
- MacIntyre, C.R.; Heslop, D.J. Public health, health systems and palliation planning for COVID-19 on an exponential timeline. Med. J. Aust. 2020, 212, 440–442 e441. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, C.; Surda, P.; Whitehead, E.; Kumar, B.N. Early recovery following new onset anosmia during the COVID-19 pandemic—An observational cohort study. J. Otolaryngol. Head. Neck Surg. 2020, 49, 26. [Google Scholar] [CrossRef]
- Giacomelli, A.; Pezzati, L.; Conti, F.; Bernacchia, D.; Siano, M.; Oreni, L.; Rusconi, S.; Gervasoni, C.; Ridolfo, A.L.; Rizzardini, G.; et al. Self-reported Olfactory and Taste Disorders in Patients with Severe Acute Respiratory Coronavirus 2 Infection: A Cross-sectional Study. Clin. Infect. Dis. 2020, 71, 889–890. [Google Scholar] [CrossRef]
- The Joint United Nations Programme on HIV/AIDS (UNAIDS). Global HIV & AIDS Statistics—Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 14 February 2023).
- African Health Organization. HIV and AIDS Fact Sheet. Available online: https://aho.org/fact-sheets/hiv-and-aids-fact-sheet/ (accessed on 14 February 2023).
- Steardo, L.; Steardo, L., Jr.; Zorec, R.; Verkhratsky, A. Neuroinfection may contribute to pathophysiology and clinical manifestations of COVID-19. Acta Physiol. 2020, 229, e13473. [Google Scholar] [CrossRef]
- Geng, J.; Wang, L.; Zhang, L.; Qin, C.; Song, Y.; Ma, Y.; Chen, Y.; Chen, S.; Wang, Y.; Zhang, Z.; et al. Blood-Brain Barrier Disruption Induced Cognitive Impairment Is Associated with Increase of Inflammatory Cytokine. Front. Aging Neurosci. 2018, 10, 129. [Google Scholar] [CrossRef]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Li, X.; Geng, M.; Peng, Y.; Meng, L.; Lu, S. Molecular immune pathogenesis and diagnosis of COVID-19. J. Pharm. Anal. 2020, 10, 102–108. [Google Scholar] [CrossRef]
- Li, Y.; Fu, L.; Gonzales, D.M.; Lavi, E. Coronavirus neurovirulence correlates with the ability of the virus to induce proinflammatory cytokine signals from astrocytes and microglia. J. Virol. 2004, 78, 3398–3406. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, X.R.; Ju, Z.Y.; He, W.F. Advances in the research of mechanism and related immunotherapy on the cytokine storm induced by coronavirus disease 2019. Zhonghua Shao Shang Za Zhi 2020, 36, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.E.; Ipser, J.C.; Stein, D.J.; Joska, J.A.; Naude, P.J.W. Peripheral immune dysregulation in the ART era of HIV-associated neurocognitive impairments: A systematic review. Psychoneuroendocrinology 2020, 118, 104689. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Cohen, R.; Gongvatana, A.; Ross, S.; Olchowski, J.; Devlin, K.; Tashima, K.; Navia, B.; Delamonte, S. Relationship of plasma cytokines and clinical biomarkers to memory performance in HIV. J. Neuroimmunol. 2013, 265, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Liu, A.; Qiao, L.; Sheng, B.; Xu, M.; Li, W.; Chen, D. The relationship of CSF and plasma cytokine levels in HIV infected patients with neurocognitive impairment. Biomed. Res. Int. 2015, 2015, 506872. [Google Scholar] [CrossRef]
- Sattler, F.R.; He, J.; Letendre, S.; Wilson, C.; Sanders, C.; Heaton, R.; Ellis, R.; Franklin, D.; Aldrovandi, G.; Marra, C.M.; et al. Abdominal obesity contributes to neurocognitive impairment in HIV-infected patients with increased inflammation and immune activation. J. Acquir. Immune Defic. Syndr. 2015, 68, 281–288. [Google Scholar] [CrossRef]
- Perrella, O.; Guerriero, M.; Izzo, E.; Soscia, M.; Carrieri, P.B. Interleukin-6 and granulocyte macrophage-CSF in the cerebrospinal fluid from HIV infected subjects with involvement of the central nervous system. Arq. Neuropsiquiatr. 1992, 50, 180–182. [Google Scholar] [CrossRef]
- Rubin, L.H.; Benning, L.; Keating, S.M.; Norris, P.J.; Burke-Miller, J.; Savarese, A.; Kumanan, K.N.; Awadalla, S.; Springer, G.; Anastos, K.; et al. Variability in C-reactive protein is associated with cognitive impairment in women living with and without HIV: A longitudinal study. J. Neurovirol. 2018, 24, 41–51. [Google Scholar] [CrossRef]
- Abassi, M.; Morawski, B.M.; Nakigozi, G.; Nakasujja, N.; Kong, X.; Meya, D.B.; Robertson, K.; Gray, R.; Wawer, M.J.; Sacktor, N.; et al. Cerebrospinal fluid biomarkers and HIV-associated neurocognitive disorders in HIV-infected individuals in Rakai, Uganda. J. Neurovirol. 2017, 23, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Kallianpur, A.R.; Gittleman, H.; Letendre, S.; Ellis, R.; Barnholtz-Sloan, J.S.; Bush, W.S.; Heaton, R.; Samuels, D.C.; Franklin, D.R., Jr.; Rosario-Cookson, D.; et al. Cerebrospinal Fluid Ceruloplasmin, Haptoglobin, and Vascular Endothelial Growth Factor Are Associated with Neurocognitive Impairment in Adults with HIV Infection. Mol. Neurobiol. 2019, 56, 3808–3818. [Google Scholar] [CrossRef] [PubMed]
- Abbasifard, M.; Khorramdelazad, H. The bio-mission of interleukin-6 in the pathogenesis of COVID-19: A brief look at potential therapeutic tactics. Life Sci. 2020, 257, 118097. [Google Scholar] [CrossRef]
- Moore, J.B.; June, C.H. Cytokine release syndrome in severe COVID-19. Science 2020, 368, 473–474. [Google Scholar] [CrossRef]
- Shekhawat, J.; Gauba, K.; Gupta, S.; Purohit, P.; Mitra, P.; Garg, M.; Misra, S.; Sharma, P.; Banerjee, M. Interleukin-6 Perpetrator of the COVID-19 Cytokine Storm. Indian J. Clin. Biochem. 2021, 36, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Tutal Gursoy, G.; Yuksel, H.; Mulkem Simsek, I.; Oral, S.; Erdogan Kucukdagli, F.; Karaman, A.; Akinci, E.; Bastug, A.; Guner, H.R.; Bektas, H. Neurological Presentations in Patients with COVID-19 in Cytokine Storm. Can. J. Neurol. Sci. 2021, 50, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, N.R.; Kunichoff, D.; Garshick, M.; Shah, B.; Pillinger, M.; Hochman, J.S.; Berger, J.S. C-reactive protein and clinical outcomes in patients with COVID-19. Eur. Heart J. 2021, 42, 2270–2279. [Google Scholar] [CrossRef]
- Yin, X.X.; Zheng, X.R.; Peng, W.; Wu, M.L.; Mao, X.Y. Vascular Endothelial Growth Factor (VEGF) as a Vital Target for Brain Inflammation during the COVID-19 Outbreak. ACS Chem. Neurosci. 2020, 11, 1704–1705. [Google Scholar] [CrossRef] [PubMed]
- Apostolou, E.; Rizwan, M.; Moustardas, P.; Sjogren, P.; Bertilson, B.C.; Bragee, B.; Polo, O.; Rosen, A. Saliva antibody-fingerprint of reactivated latent viruses after mild/asymptomatic COVID-19 is unique in patients with myalgic-encephalomyelitis/chronic fatigue syndrome. Front. Immunol. 2022, 13, 949787. [Google Scholar] [CrossRef]
- Stevenson, E.M.; Terry, S.; Copertino, D.; Leyre, L.; Danesh, A.; Weiler, J.; Ward, A.R.; Khadka, P.; McNeil, E.; Bernard, K.; et al. SARS-CoV-2 mRNA vaccination exposes latent HIV to Nef-specific CD8(+) T-cells. Nat. Commun. 2022, 13, 4888. [Google Scholar] [CrossRef]
- Teuwen, L.A.; Geldhof, V.; Pasut, A.; Carmeliet, P. COVID-19: The vasculature unleashed. Nat. Rev. Immunol. 2020, 20, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Perl, D.P.; Nair, G.; Li, W.; Maric, D.; Murray, H.; Dodd, S.J.; Koretsky, A.P.; Watts, J.A.; Cheung, V.; et al. Microvascular Injury in the Brains of Patients with COVID-19. N. Engl. J. Med. 2021, 384, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Proal, A.D.; VanElzakker, M.B. Long COVID or Post-acute Sequelae of COVID-19 (PASC): An Overview of Biological Factors That May Contribute to Persistent Symptoms. Front. Microbiol. 2021, 12, 698169. [Google Scholar] [CrossRef]
- Peluso, M.J.; Spinelli, M.A.; Deveau, T.M.; Forman, C.A.; Munter, S.E.; Mathur, S.; Tang, A.F.; Lu, S.; Goldberg, S.A.; Arreguin, M.I.; et al. Postacute sequelae and adaptive immune responses in people with HIV recovering from SARS-CoV-2 infection. AIDS 2022, 36, F7–F16. [Google Scholar] [CrossRef]
- Pujari, S.; Gaikwad, S.; Chitalikar, A.; Dabhade, D.; Joshi, K.; Bele, V. Long-coronavirus disease among people living with HIV in western India: An observational study. Immun. Inflamm. Dis. 2021, 9, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Kingery, J.R.; Safford, M.M.; Martin, P.; Lau, J.D.; Rajan, M.; Wehmeyer, G.T.; Li, H.A.; Alshak, M.N.; Jabri, A.; Kofman, A.; et al. Health Status, Persistent Symptoms, and Effort Intolerance One Year After Acute COVID-19 Infection. J. Gen. Intern. Med. 2022, 37, 1218–1225. [Google Scholar] [CrossRef]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.M.L.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J.; et al. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.H.; O’Connor, J.L.; Phillips, A.N.; Ronsholt, F.F.; Pett, S.; Vjecha, M.J.; French, M.A.; Lundgren, J.D.; Insight, S.; Groups, E.S.; et al. Factors Associated With Plasma IL-6 Levels During HIV Infection. J. Infect. Dis. 2015, 212, 585–595. [Google Scholar] [CrossRef]
- Serrano-Villar, S.; Sainz, T.; Lee, S.A.; Hunt, P.W.; Sinclair, E.; Shacklett, B.L.; Ferre, A.L.; Hayes, T.L.; Somsouk, M.; Hsue, P.Y.; et al. HIV-infected individuals with low CD4/CD8 ratio despite effective antiretroviral therapy exhibit altered T cell subsets, heightened CD8+ T cell activation, and increased risk of non-AIDS morbidity and mortality. PLoS Pathog. 2014, 10, e1004078. [Google Scholar] [CrossRef]
- Zicari, S.; Sessa, L.; Cotugno, N.; Ruggiero, A.; Morrocchi, E.; Concato, C.; Rocca, S.; Zangari, P.; Manno, E.C.; Palma, P. Immune Activation, Inflammation, and Non-AIDS Co-Morbidities in HIV-Infected Patients under Long-Term ART. Viruses 2019, 11, 200. [Google Scholar] [CrossRef]
- Pretorius, E.; Vlok, M.; Venter, C.; Bezuidenhout, J.A.; Laubscher, G.J.; Steenkamp, J.; Kell, D.B. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc. Diabetol. 2021, 20, 172. [Google Scholar] [CrossRef]
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple early factors anticipate post-acute COVID-19 sequelae. Cell 2022, 185, 881–895 e820. [Google Scholar] [CrossRef] [PubMed]
- Khoury, G.; Fromentin, R.; Solomon, A.; Hartogensis, W.; Killian, M.; Hoh, R.; Somsouk, M.; Hunt, P.W.; Girling, V.; Sinclair, E.; et al. Human Immunodeficiency Virus Persistence and T-Cell Activation in Blood, Rectal, and Lymph Node Tissue in Human Immunodeficiency Virus-Infected Individuals Receiving Suppressive Antiretroviral Therapy. J. Infect. Dis. 2017, 215, 911–919. [Google Scholar] [CrossRef]
- Zandman-Goddard, G.; Shoenfeld, Y. HIV and autoimmunity. Autoimmun. Rev. 2002, 1, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Cilloni, L.; Fu, H.; Vesga, J.F.; Dowdy, D.; Pretorius, C.; Ahmedov, S.; Nair, S.A.; Mosneaga, A.; Masini, E.; Sahu, S.; et al. The potential impact of the COVID-19 pandemic on the tuberculosis epidemic a modelling analysis. EClinicalMedicine 2020, 28, 100603. [Google Scholar] [CrossRef]
- Kelley, N.; Jeltema, D.; Duan, Y.; He, Y. The NLRP3 Inflammasome: An Overview of Mechanisms of Activation and Regulation. Int. J. Mol. Sci. 2019, 20, 3328. [Google Scholar] [CrossRef] [PubMed]
- Cassel, S.L.; Joly, S.; Sutterwala, F.S. The NLRP3 inflammasome: A sensor of immune danger signals. Semin. Immunol. 2009, 21, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xu, W.; Zhou, R. NLRP3 inflammasome activation and cell death. Cell. Mol. Immunol. 2021, 18, 2114–2127. [Google Scholar] [CrossRef]
- Dutta, D.; Liu, J.; Xiong, H. NLRP3 inflammasome activation and SARS-CoV-2-mediated hyperinflammation, cytokine storm and neurological syndromes. Int. J. Physiol. Pathophysiol. Pharmacol. 2022, 14, 138–160. [Google Scholar]
- Campbell, G.R.; To, R.K.; Hanna, J.; Spector, S.A. SARS-CoV-2, SARS-CoV-1, and HIV-1 derived ssRNA sequences activate the NLRP3 inflammasome in human macrophages through a non-classical pathway. iScience 2021, 24, 102295. [Google Scholar] [CrossRef]
- Cama, V.F.; Marin-Prida, J.; Acosta-Rivero, N.; Acosta, E.F.; Diaz, L.O.; Casadesus, A.V.; Fernandez-Marrero, B.; Gilva-Rodriguez, N.; Cremata-Garcia, D.; Cervantes-Llanos, M.; et al. The microglial NLRP3 inflammasome is involved in human SARS-CoV-2 cerebral pathogenicity: A report of three post-mortem cases. J. Neuroimmunol. 2021, 361, 577728. [Google Scholar] [CrossRef]
- Pan, P.; Shen, M.; Yu, Z.; Ge, W.; Chen, K.; Tian, M.; Xiao, F.; Wang, Z.; Wang, J.; Jia, Y.; et al. SARS-CoV-2 N protein promotes NLRP3 inflammasome activation to induce hyperinflammation. Nat. Commun. 2021, 12, 4664. [Google Scholar] [CrossRef] [PubMed]
- Yalcinkaya, M.; Liu, W.; Islam, M.N.; Kotini, A.G.; Gusarova, G.A.; Fidler, T.P.; Papapetrou, E.P.; Bhattacharya, J.; Wang, N.; Tall, A.R. Modulation of the NLRP3 inflammasome by SARS-CoV-2 Envelope protein. Sci. Rep. 2021, 11, 24432. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Lan, X.; Wen, H.; Lederman, R.; Chawla, A.; Attia, M.; Bongu, R.P.; Husain, M.; Mikulak, J.; Saleem, M.A.; et al. HIV Promotes NLRP3 Inflammasome Complex Activation in Murine HIV-Associated Nephropathy. Am. J. Pathol. 2016, 186, 347–358. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Yang, W.; Zeng, Z.; Wei, Y.; Gao, J.; Zhang, B.; Li, L.; Liu, L.; Wan, Y.; Zeng, Q.; et al. NLRP3-dependent pyroptosis is required for HIV-1 gp120-induced neuropathology. Cell. Mol. Immunol. 2020, 17, 283–299. [Google Scholar] [CrossRef]
- Chivero, E.T.; Guo, M.L.; Periyasamy, P.; Liao, K.; Callen, S.E.; Buch, S. HIV-1 Tat Primes and Activates Microglial NLRP3 Inflammasome-Mediated Neuroinflammation. J. Neurosci. 2017, 37, 3599–3609. [Google Scholar] [CrossRef]
- Gumbs, S.B.H.; Berdenis van Berlekom, A.; Kubler, R.; Schipper, P.J.; Gharu, L.; Boks, M.P.; Ormel, P.R.; Wensing, A.M.J.; de Witte, L.D.; Nijhuis, M. Characterization of HIV-1 Infection in Microglia-Containing Human Cerebral Organoids. Viruses 2022, 14, 829. [Google Scholar] [CrossRef]
- Min, A.K.; Fortune, T.; Rodriguez, N.; Hedge, E.; Swartz, T.H. Inflammasomes as mediators of inflammation in HIV-1 infection. Transl. Res. 2023, 252, 1–8. [Google Scholar] [CrossRef]
- Gougeon, M.L. Alarmins and central nervous system inflammation in HIV-associated neurological disorders. J. Intern. Med. 2017, 281, 433–447. [Google Scholar] [CrossRef]
- Mamik, M.K.; Hui, E.; Branton, W.G.; McKenzie, B.A.; Chisholm, J.; Cohen, E.A.; Power, C. HIV-1 Viral Protein R Activates NLRP3 Inflammasome in Microglia: Implications for HIV-1 Associated Neuroinflammation. J. Neuroimmune Pharmacol. 2017, 12, 233–248. [Google Scholar] [CrossRef]
- Costela-Ruiz, V.J.; Illescas-Montes, R.; Puerta-Puerta, J.M.; Ruiz, C.; Melguizo-Rodriguez, L. SARS-CoV-2 infection: The role of cytokines in COVID-19 disease. Cytokine Growth Factor. Rev. 2020, 54, 62–75. [Google Scholar] [CrossRef]
- Chen, R.; Lan, Z.; Ye, J.; Pang, L.; Liu, Y.; Wu, W.; Qin, X.; Guo, Y.; Zhang, P. Cytokine Storm: The Primary Determinant for the Pathophysiological Evolution of COVID-19 Deterioration. Front. Immunol. 2021, 12, 589095. [Google Scholar] [CrossRef]
- Amruta, N.; Chastain, W.H.; Paz, M.; Solch, R.J.; Murray-Brown, I.C.; Befeler, J.B.; Gressett, T.E.; Longo, M.T.; Engler-Chiurazzi, E.B.; Bix, G. SARS-CoV-2 mediated neuroinflammation and the impact of COVID-19 in neurological disorders. Cytokine Growth Factor. Rev. 2021, 58, 1–15. [Google Scholar] [CrossRef]
- Buzhdygan, T.P.; DeOre, B.J.; Baldwin-Leclair, A.; Bullock, T.A.; McGary, H.M.; Khan, J.A.; Razmpour, R.; Hale, J.F.; Galie, P.A.; Potula, R.; et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier. Neurobiol. Dis. 2020, 146, 105131. [Google Scholar] [CrossRef] [PubMed]
- Lara, P.C.; Macias-Verde, D.; Burgos-Burgos, J. Age-induced NLRP3 Inflammasome Over-activation Increases Lethality of SARS-CoV-2 Pneumonia in Elderly Patients. Aging Dis. 2020, 11, 756–762. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738 e713. [Google Scholar] [CrossRef]
- Yang, H.; Rao, Z. Structural biology of SARS-CoV-2 and implications for therapeutic development. Nat. Rev. Microbiol. 2021, 19, 685–700. [Google Scholar] [CrossRef]
- Xu, H.; Akinyemi, I.A.; Chitre, S.A.; Loeb, J.C.; Lednicky, J.A.; McIntosh, M.T.; Bhaduri-McIntosh, S. SARS-CoV-2 viroporin encoded by ORF3a triggers the NLRP3 inflammatory pathway. Virology 2022, 568, 13–22. [Google Scholar] [CrossRef]
- Theobald, S.J.; Simonis, A.; Georgomanolis, T.; Kreer, C.; Zehner, M.; Eisfeld, H.S.; Albert, M.C.; Chhen, J.; Motameny, S.; Erger, F.; et al. Long-lived macrophage reprogramming drives spike protein-mediated inflammasome activation in COVID-19. EMBO Mol. Med. 2021, 13, e14150. [Google Scholar] [CrossRef]
- Olajide, O.A.; Iwuanyanwu, V.U.; Lepiarz-Raba, I.; Al-Hindawi, A.A. Induction of Exaggerated Cytokine Production in Human Peripheral Blood Mononuclear Cells by a Recombinant SARS-CoV-2 Spike Glycoprotein S1 and Its Inhibition by Dexamethasone. Inflammation 2021, 44, 1865–1877. [Google Scholar] [CrossRef]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. SARS coronavirus spike protein-induced innate immune response occurs via activation of the NF-kappaB pathway in human monocyte macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, T.; Alghanem, B.; Shaibah, H.; Mansour, F.A.; Alamri, H.S.; Akiel, M.A.; Alroqi, F.; Boudjelal, M. SARS-CoV-2 Coronavirus Spike Protein-Induced Apoptosis, Inflammatory, and Oxidative Stress Responses in THP-1-Like-Macrophages: Potential Role of Angiotensin-Converting Enzyme Inhibitor (Perindopril). Front. Immunol. 2021, 12, 728896. [Google Scholar] [CrossRef]
- Frank, M.G.; Nguyen, K.H.; Ball, J.B.; Hopkins, S.; Kelley, T.; Baratta, M.V.; Fleshner, M.; Maier, S.F. SARS-CoV-2 spike S1 subunit induces neuroinflammatory, microglial and behavioral sickness responses: Evidence of PAMP-like properties. Brain Behav. Immun. 2022, 100, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Cho, W.H.; Barcelon, E.; Kim, K.H.; Hong, J.; Lee, S.J. SARS-CoV-2 spike protein induces cognitive deficit and anxiety-like behavior in mouse via non-cell autonomous hippocampal neuronal death. Sci. Rep. 2022, 12, 5496. [Google Scholar] [CrossRef]
- Ma, J.; Zhu, F.; Zhao, M.; Shao, F.; Yu, D.; Ma, J.; Zhang, X.; Li, W.; Qian, Y.; Zhang, Y.; et al. SARS-CoV-2 nucleocapsid suppresses host pyroptosis by blocking Gasdermin D cleavage. EMBO J. 2021, 40, e108249. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sui, L.; Wu, P.; Wang, W.; Wang, Z.; Yu, Y.; Hou, Z.; Tan, G.; Liu, Q.; Wang, G. A dual-role of SARS-CoV-2 nucleocapsid protein in regulating innate immune response. Signal. Transduct. Target. Ther. 2021, 6, 331. [Google Scholar] [CrossRef]
- Potere, N.; Del Buono, M.G.; Caricchio, R.; Cremer, P.C.; Vecchie, A.; Porreca, E.; Dalla Gasperina, D.; Dentali, F.; Abbate, A.; Bonaventura, A. Interleukin-1 and the NLRP3 inflammasome in COVID-19: Pathogenetic and therapeutic implications. EBioMedicine 2022, 85, 104299. [Google Scholar] [CrossRef]
- Al-Aly, Z.; Xie, Y.; Bowe, B. High-dimensional characterization of post-acute sequelae of COVID-19. Nature 2021, 594, 259–264. [Google Scholar] [CrossRef]
- World Health Organization. COVID-19 Pandemic Triggers 25% Increase in Prevalence of Anxiety and Depression Worldwide; WHO: Geneva, Switzerland, 2022. [Google Scholar]
- Del Amo, J.; Polo, R.; Moreno, S.; Diaz, A.; Martinez, E.; Arribas, J.R.; Jarrin, I.; Hernan, M.A. Incidence and Severity of COVID-19 in HIV-Positive Persons Receiving Antiretroviral Therapy. Ann. Intern. Med. 2021, 174, 581–582. [Google Scholar] [CrossRef]
- Titanji, B.; Hernandez-Romieu, A.C.; Adelman, M.W. Incidence and Severity of COVID-19 in HIV-Positive Persons Receiving Antiretroviral Therapy. Ann. Intern. Med. 2021, 174, 580–581. [Google Scholar] [CrossRef]
- Ponticiello, M.; Mwanga-Amumpaire, J.; Tushemereirwe, P.; Nuwagaba, G.; King, R.; Sundararajan, R. “Everything is a Mess”: How COVID-19 is Impacting Engagement with HIV Testing Services in Rural Southwestern Uganda. AIDS Behav. 2020, 24, 3006–3009. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Dujaili, J.; Sandhu, A.K.; Hashmi, F.K. Concerns of HIV-positive migrant workers in COVID-19 pandemic: A call for action. J. Glob. Health 2020, 10, 020342. [Google Scholar] [CrossRef] [PubMed]
- Lodge, W., 2nd; Kuchukhidze, S. COVID-19, HIV, and Migrant Workers: The Double Burden of the Two Viruses. AIDS Patient Care STDS 2020, 34, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, J.D.; Skrzat-Klapaczynska, A.; Bursa, D.; Balayan, T.; Begovac, J.; Chkhartishvili, N.; Gokengin, D.; Harxhi, A.; Jilich, D.; Jevtovic, D.; et al. HIV care in times of the COVID-19 crisis—Where are we now in Central and Eastern Europe? Int. J. Infect. Dis. 2020, 96, 311–314. [Google Scholar] [CrossRef]
- Jewell, B.L.; Mudimu, E.; Stover, J.; Ten Brink, D.; Phillips, A.N.; Smith, J.A.; Martin-Hughes, R.; Teng, Y.; Glaubius, R.; Mahiane, S.G.; et al. Potential effects of disruption to HIV programmes in sub-Saharan Africa caused by COVID-19: Results from multiple mathematical models. Lancet HIV 2020, 7, e629–e640. [Google Scholar] [CrossRef]
- Wu, S.; Ming, F.; Xing, Z.; Zhang, Z.; Zhu, S.; Guo, W.; Zou, S.; Liu, J.; Liu, Y.; Liang, K. COVID-19 Vaccination Willingness Among People Living with HIV in Wuhan, China. Front. Public. Health 2022, 10, 883453. [Google Scholar] [CrossRef]
- Hensley, K.S.; Jongkees, M.J.; Geers, D.; GeurtsvanKessel, C.H.; Mueller, Y.M.; Dalm, V.; Papageorgiou, G.; Steggink, H.; Gorska, A.; Bogers, S.; et al. Immunogenicity and reactogenicity of SARS-CoV-2 vaccines in people living with HIV in the Netherlands: A nationwide prospective cohort study. PLoS Med. 2022, 19, e1003979. [Google Scholar] [CrossRef]



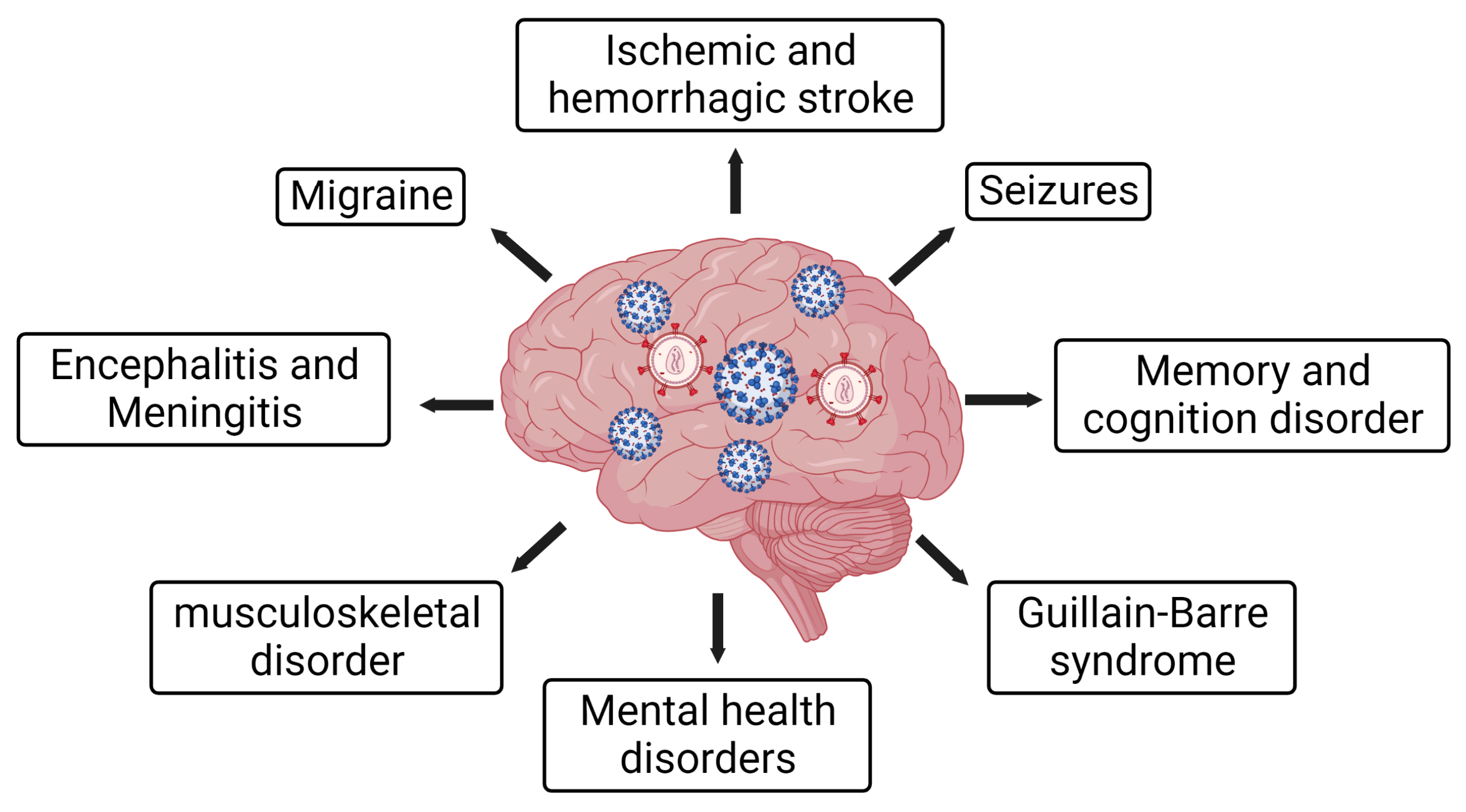{kind=link}
{kind=link}
| SARS-CoV-2 | HIV-1 | |
|---|---|---|
| Yes | Public fear | Yes |
| Yes | Enveloped virus | Yes |
| Yes | ssRNA genome | Yes |
| Yes | Natural origin | Yes |
| No | Symptoms in the natural reservoir | No |
| Yes | Asymptomatic spread | Yes |
| Yes | Inflammation | Yes |
| Yes | NLRP3 inflammasome activation | Yes |
| Yes | Lymphopenia | Yes |
| Yes | NETosis | Yes |
| Yes | Neurocognitive disorders | Yes |
| SARS-CoV-2 | HIV-1 | |
|---|---|---|
| Phylogeny | β-coronavirus | Lentivirus |
| Virion size | Spherical particle 50–200 nm in diameter | Spherical particle ~100 nm in diameter |
| Genome | One copy of single-stranded positive-sense RNA | Two copies of single-stranded positive-sense RNA |
| Genome size | ~29.2 kb | ~10 kb |
| Genome integration | No | Yes |
| Reservoir | No viral reservoir formation | This virus integrates into the human genome and forms reservoirs |
| Transmission | Air, aerosol | Sexual, body fluid |
| Receptor in use | ACE2, TMPRSS2 | CD4, CCR5 and CXCR4 |
| Symptoms | Breathing difficulty, fever, pneumonia and kidney failure | Flu or mononucleosis-like during early infection and opportunistic infections in late stages. Long time to progress to AIDS |
| Symptom timeline | 2–14 days after contact with the virus | 2–6 weeks after viral contact |
| Death percentage | 1–4% | ≥95% |
| Vaccine | Available | Not available |
| Medicine | Antivirals | Antiretroviral therapy (cART) |
| Cure | Curable (By antivirals or plasma therapy) | No cure (Can be controlled with cART) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutta, D.; Liu, J.; Xiong, H. The Impact of COVID-19 on People Living with HIV-1 and HIV-1-Associated Neurological Complications. Viruses 2023, 15, 1117. https://doi.org/10.3390/v15051117
Dutta D, Liu J, Xiong H. The Impact of COVID-19 on People Living with HIV-1 and HIV-1-Associated Neurological Complications. Viruses. 2023; 15(5):1117. https://doi.org/10.3390/v15051117
Chicago/Turabian StyleDutta, Debashis, Jianuo Liu, and Huangui Xiong. 2023. "The Impact of COVID-19 on People Living with HIV-1 and HIV-1-Associated Neurological Complications" Viruses 15, no. 5: 1117. https://doi.org/10.3390/v15051117