Hantaviruses in the Americas and Their Role as Emerging Pathogens

1
Center for Infectious Diseases and Immunity, Department of Pathology, University of New Mexico Health Sciences Center, MSC08 4640, Albuquerque, New Mexico 87131, USA
2
Biology Department, University of New Mexico, MSC03 2020, Albuquerque, New Mexico 87131, USA
*
Author to whom correspondence should be addressed.
Viruses 2010, 2(12), 2559-2586; https://doi.org/10.3390/v2122559
Submission received: 25 October 2010
/
Revised: 15 November 2010
/
Accepted: 24 November 2010
/
Published: 25 November 2010
(This article belongs to the Special Issue Pathogenesis of Emerging and Re-Emerging RNA Viruses)
Abstract
:The continued emergence and re-emergence of pathogens represent an ongoing, sometimes major, threat to populations. Hantaviruses (family Bunyaviridae) and their associated human diseases were considered to be confined to Eurasia, but the occurrence of an outbreak in 1993–94 in the southwestern United States led to a great increase in their study among virologists worldwide. Well over 40 hantaviral genotypes have been described, the large majority since 1993, and nearly half of them pathogenic for humans. Hantaviruses cause persistent infections in their reservoir hosts, and in the Americas, human disease is manifest as a cardiopulmonary compromise, hantavirus cardiopulmonary syndrome (HCPS), with case-fatality ratios, for the most common viral serotypes, between 30% and 40%. Habitat disturbance and larger-scale ecological disturbances, perhaps including climate change, are among the factors that may have increased the human caseload of HCPS between 1993 and the present. We consider here the features that influence the structure of host population dynamics that may lead to viral outbreaks, as well as the macromolecular determinants of hantaviruses that have been regarded as having potential contribution to pathogenicity.
1. Introduction
Emerging pathogens cause new or previously unrecognized diseases, and among them, emerging zoonotic diseases are a major concern among scientists studying infectious diseases at different spatial and temporal scales [1,2]. Changes in biotic and abiotic conditions may alter population disease dynamics and lead to the emergence of zoonotic infections [3,4,5,6]. During the last decades, several outbreaks of emerging and re-emerging viral pathogens have occurred, affecting both purely-local and worldwide/pandemic involvement of human populations. Among the conspicuous examples are influenza A, Ebola virus, hepatitis C virus, severe adult respiratory distress (SARS), coronavirus, and human immunodeficiency virus, which challenge prevention and control measures of public health systems [7]. In the Americas, the recent outbreak of pandemic influenza A subtype H1N1 became a major target for control due to its rapid spread, and uncertainties in virulence and transmissibility, yet vaccine availability was limited when significant activity occurred in advance of the traditional influenza season [8]. However, in the last century outbreaks of several viral-related diseases have emerged or re-emerged involving arenaviruses and dengue viruses, and more recently, hantaviruses, and the expansion of the geographic range of West Nile virus. Among zoonotic diseases, small mammals are hosts of several pathogenic RNA viruses, especially Arenaviridae and Bunyaviridae: Hantavirus [9,10,11].
Hantavirus infections became a concern in the Americas after the description of an outbreak of acute respiratory distress occurred in the Four Corners area in 1993 [12]. The newly recognized disease, hantavirus cardiopulmonary syndrome, HCPS (or hantavirus pulmonary syndrome), was linked to infection by the newly-discovered Sin Nombre virus (SNV), and the rodent Peromyscus maniculatus (deer mouse) was identified as the reservoir [13]. However, hantavirus infections have a much longer history. A review of ancient Chinese writings, dating back to approximately 960 AD, revealed descriptions closely resembling hemorrhagic fever with renal syndrome (HFRS), the syndrome caused by Old World hantaviruses [14]. During the twentieth century, cases of acute febrile disease with renal compromise were described from several Eurasian countries and Japan, often in association with military engagements [15]. HFRS as a distinct syndrome, however, was first brought to the attention of western medicine in association with an outbreak that occurred among United Nations troops during the Korean conflict between 1951 and 1954, where more than 3,200 soldiers were afflicted [16]. It took more than two decades until the etiologic agent, Hantaan virus (HTNV), was isolated from the striped field mouse Apodemus agrarius, detected in part by the binding of antibodies from patient serum samples to the lung tissues of healthy, wild-caught field mice [17,18]. The virus was later found to represent the type species of a new genus Hantavirus of the family Bunyaviridae, although it was later apparent that the first hantavirus to be isolated was the shrew-borne Thottapalayam virus [19]. The categorization of hantaviruses as belonging to the family Bunyaviridae is due in part to the consistent presence of three RNA genomes that are circularized in vivo as a result of the presence of terminal complementary nucleotides that help fold the genome into a “hairpin” morphology, first described for the Uukuniemi phlebovirus [19,20]. Table 1 is a list of the predominant, serologically distinct pathogenic hantaviruses. Many other named genotypes are described, but such other pathogenic forms are generally closely related to Andes or, in some cases, Sin Nombre virus.
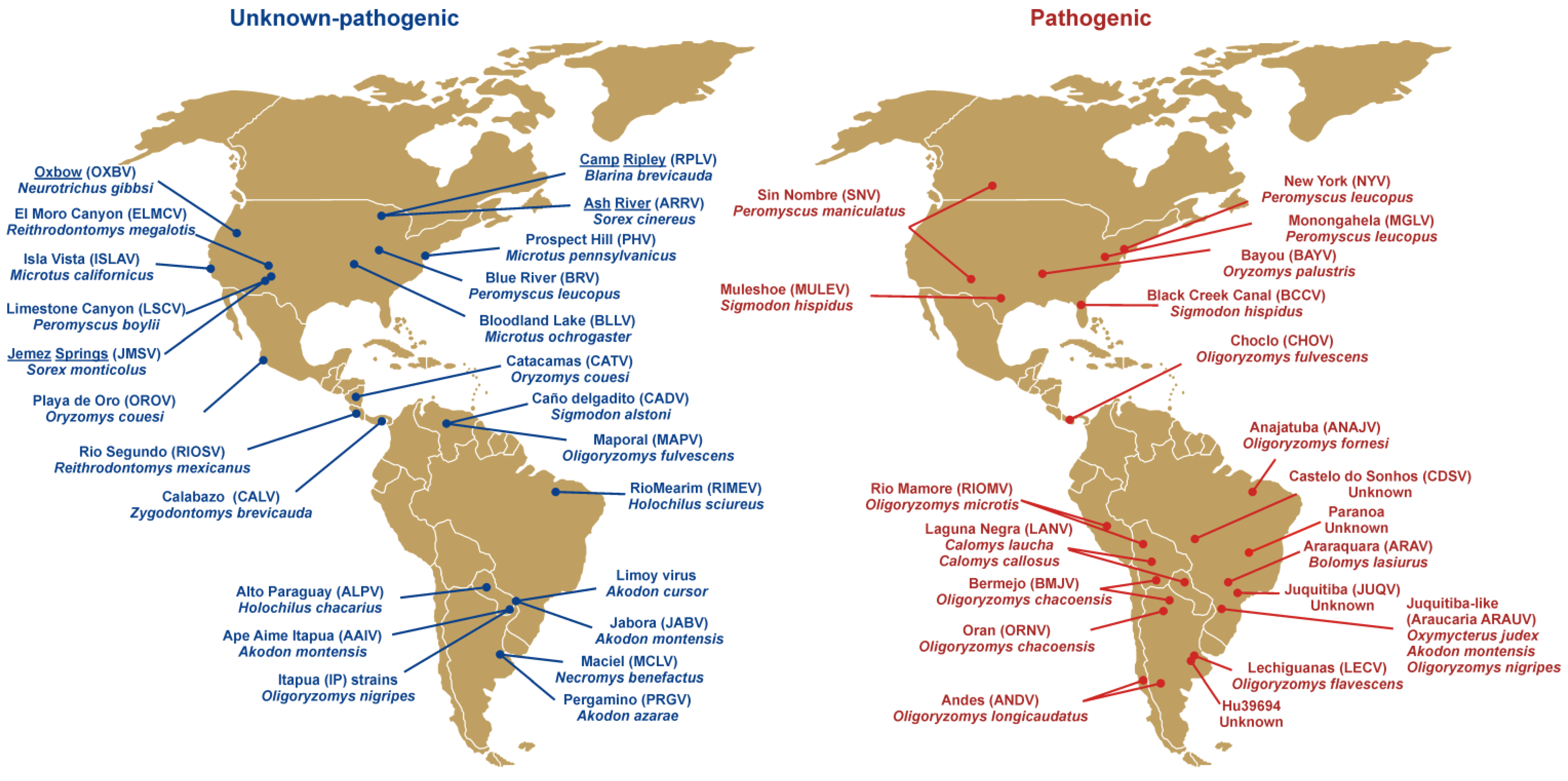
Hantavirus discovery continues at a rapid pace. After the identification of SNV in 1993, approximately 43 genotypes have been reported in the Americas alone (Figure 1), with 20 of those associated with clinical cases of HCPS. Most hantaviruses worldwide have been identified in murid or cricetid rodents of the subfamilies Murinae, Arvicolinae, Neotominae, and Sigmodontinae as their main reservoir, but the discovery of hantaviruses in insectivores (Soricidae, Talpidae) on at least three continents [21,22] shows that the genetic diversity of hantaviruses is far higher than expected.
2. Molecular Features and Cell Interactions
Member viruses of the family Bunyaviridae, including those of the genus Hantavirus are spherical, enveloped viruses of 100–200 nm diameter with a tripartite helical nucleocapsid. The hantavirus genome is composed of three negative sense single stranded RNA segments: (i) The large (L) segment of approximately 6,500 nucleotides (nt) encodes a RNA-dependent RNA polymerase of ~2,160 amino acids (aa); (ii) The middle (M) segment is 3,700-nt long, and encodes the envelope glycoprotein precursor (GPC), which is processed into two transmembrane glycoproteins, Gn (652 aa, formerly “G1”) and Gc (488 aa, previously “G2”) through a cotranslational proteolytic mechanism in the endoplasmic reticulum [23,24,25,26]; (iii) The small (S) segment length varies from ~1,700 to 2,100 nt, and encodes an RNA-binding nucleocapsid (N) protein that ranges between 428 and 436-aa for hantaviruses.
Figure 1.
Map of hantavirus genotypes in the Americas reported to be non-pathogenic or of unknown pathogenicity (blue) and pathogenic (red) for humans. Hantaviruses reported in Soricomorpha (shrews and moles) are underlined.
Figure 1.
Map of hantavirus genotypes in the Americas reported to be non-pathogenic or of unknown pathogenicity (blue) and pathogenic (red) for humans. Hantaviruses reported in Soricomorpha (shrews and moles) are underlined.

2.1. Envelope Glycoproteins
During virus maturation, the precursor form GPC is processed using a membrane-bound protease into Gn and Gc, a cleavage that occurs, and appears to be signaled, after the conserved peptide signal WAASA at the C-terminal of Gn [24]. Although the two proteins can be expressed independently through transfection, they can be retained in the wrong cellular compartment (ER or aggresome); they thus must be co-expressed to allow them stability so that the two can be assembled correctly in the Golgi [25,27,28,29].
A number of activities and properties have been identified for the hantavirus envelope glycoproteins, including some features that are suspected to be involved in the pathogenicity of the disease-causing serotypes, a possibility that has engendered experimental attention. The glycoproteins are the known or presumed ligands for at least two distinct cellular receptors, the β3 integrin chain and decay accelerating factor, or DAF [30,31]; with gC1qR/p32 also identified as another potential entry receptor [32]. Comparisons with the tick-borne encephalitis virus E protein, led Tischler et al. to consider the Gc glycoprotein as a potential class II fusion protein, perhaps imparting fusion activity to the virion, and this hypothesis has gained support in other studies [33,34].
Additional activities have been identified with, or claimed to be related to, Gn. For many of these studies, an underlying premise has held that there are differences between the glycoproteins of “pathogenic” hantaviruses relative to viruses in the genus that are dubbed to be “non-pathogenic”. While it is true that it has not yet been possible to link Prospect Hill virus (PHV) to human disease, the absence of evidence for its pathogenicity should perhaps not be equated with the evidence of its absence. One might only consider that the level of disease (e.g., lethargy, fever, proteinuria, and azotemia) associated with infection of nonhuman primates by PHV is not significantly different from that recorded for nonhuman primate models using the known-pathogen Puumala virus (PUUV) [35,36]. For the purpose of this discussion we will presume that apathogenic hantaviruses are indeed apathogenic.
While some studies have suggested that Gn glycoproteins are directed more rapidly into the ubiquitin-proteosome pathway than are apathogenic forms, others have interpreted differences in the handling of Gn glycoproteins across hantavirus species by the ubiquitin-proteosomal system as independent of pathogenicity [37,38,39]. Some investigators have directed their efforts toward identifying a differential capacity, either kinetic or in absolute magnitude, in the ability of pathogenic and apathogenic hantaviruses to elicit an interferon response in cells. One premise that emerges is that apathogenic forms would tend to induce an earlier innate response that would render it more likely that the virus would be quickly cleared or rendered less competent in its replication so as to blunt any pathological response in the host [40,41,42]. The anti-hantavirus innate response can in some cases be attributed to viral interaction as a ligand of TLR-3, but not in others, and in endothelial cells, it appears not to require more than the viral particle itself, even when introduced in replication-incompetent form [43,44]. Proteins and mRNAs prominently induced by hantaviruses include MxA and IFIT-1 (ISG-56) and others including some with known or suspected anti-viral activity. Those hantaviruses, often highly pathogenic strains, that fail to induce a potent antiviral response, are suspected or presumed to have a (more) potent interferon-pathway antagonism mechanism relative to other viruses, a mechanism that acts positively to prevent an effective innate response from forming, at least early in infection [42,45]. Yet some instances are reported wherein highly pathogenic hantaviruses, such as SNV, are also able to induce expression of interferon-stimulated gene mRNAs, even very early in infection, with ISG proteins, as expected, taking longer to appear in the cell [44]. Anti-interferon activities have also been attributed to the NSs protein that may be elaborated in cells infected by serotypes that encode this protein [46]. Other investigators have examined the activities of hantavirus glycoproteins and other proteins that might themselves directly affect some aspects of the pathogenic progression associated with hantavirus infection of humans, such as vascular permeability changes. While early attempts to directly cause increases in permeability of endothelial monolayers with viral particles or viral infection were largely disappointing, hantaviruses have been identified as adversely affecting endothelial migration over substrata and in potentiating VEG-F-induced endothelial permeability [47,48].
2.2. The Hantavirus Nucleocapsid Protein
The shorter (≈50-kD) nucleocapsid or N protein is a structural component of the viral nucleocapsid, along with the genomic viral RNA segments. As an RNA-binding protein that engages the hairpin termini of the genomic segments with high affinity [49,50], it limits the access of the RNA to host nucleases and helps to render viral replication a closed process within the cytoplasm. It also acts as a peripheral membrane protein, as does the L protein [51], an activity that could play a role in its presumed, but not yet demonstrated function as matrix [52]. Until recently, it had not been appreciated that N has a wide variety of other activities, some of which can be linked, not only to fundamental requirements of replication, but also to the interference with an array of the intracellular processes of the normal cell. Thus, an interaction between the amino terminus of the hantavirus N protein and the cellular protein Daxx has been proposed, with the suggestion of potential pro-apoptotic consequences [51]. N is also reported to interact with actin microfilaments, and the SUMO-1 protein [53,54]. Using reporter-gene based assays, Connie Schmaljohn and her colleagues have reported that Hantaan virus’ nucleocapsid protein has an inhibitory role in inflammatory responses mediated by NF kappa B (NF-κB). The effects on NF-κB expression appeared to be confined to prevention of its nuclear translocation after its attempted activation with lipopolysaccharide, LPS [55]. In the cytoplasm of infected cells, N protein can be found in cellular P bodies where it sequesters and protects 5’ caps. It may locate the caps through its interaction with DCP1, a key constituent of P bodies. During hantavirus infection, the viral RNAs become concentrated in P bodies, through their interaction with N and DCP1. The N protein demonstrates preferential protection of mRNAs engineered to prematurely terminate their encoded protein in comparison to native mRNAs [56].
N protein has been increasingly linked to viral replication and translation, sometimes in previously unanticipated ways. It is among a growing family of diverse viral proteins that can serve as a nonspecific “RNA chaperone”, an activity that should facilitate the L polymerase’s access to vRNA for transcription and replication, in that it can transiently dissociate misfolded RNA structures [57]. Some of N protein’s effects on translation might not immediately be recognized to be adaptive in nature. It can replace the entire EIF4F translational initiation complex, simultaneously presenting the ribosome with a replacement for the cap-binding activity of eIF 4E, binding to the 43S pre-initiation complex as does eIF 4G, while replacing the helicase activity of eIF 4A, which is presumed to be needed to dissociate higher-order RNA structure [56,58]. These three factors normally work together to achieve translational initiation. In P bodies, N protein’s ability to bind at high affinity to capped native cellular oligoribonucleotides, along with its activity in protecting capped RNAs from degradation likely facilitates the access of capped oligonucleotides for use in transcriptional initiation by L polymerase (“cap snatching”).
Trafficking of N for viral assembly: Classically, N protein in infected cells appears to be clustered or particulate in nature, with a heavy concentration at a single perinuclear location, widely considered to be the Golgi [27]. The N proteins of hantaviruses are found in association with particulate fractions, and confocal microscopy and biochemical-inhibitor studies have shown that N tracks along microtubules but not with actin filaments [52]. The ultimate destination for N, for its assembly into viral particles is the Golgi, and it traffics there via the endoplasmic reticulum-Golgi intermediate complex (ERGIC), also known as vesicular-tubular cluster [52]. A dominant negative inhibitor, dynamitin, associated with dynein-mediated transport, reduced N’s accumulation in the Golgi. Later studies suggested that the specific dependence on microtubular transport is specific to Old World hantaviruses such as HTNV, but that the New World hantavirus ANDV is instead associated with actin filaments [59]. However, recent data indicates that microtubular transport is indeed utilized for the New World hantavirus SNV [60].
3. Pathology and Diagnosis
Hantavirus diseases of man have long been suspected of having an immunopathogenic basis in part because of their relatively long incubation period of 2–3 weeks and the observed temporal association between immunologic derangements and the first appearance of signs and symptoms of hantavirus illness. HFRS and HCPS share many clinical features, leading many investigators to consider them to be, in essence, different manifestations of a similar pathogenic process, differing mainly in the primary target organs of disease expression (Table 2). The pathogenesis of hantavirus infections is the topic of a continuously-updated review in the series UpToDate [61].
By the time symptoms appear in HCPS, both strong antiviral responses, and, for the more virulent viral genotypes, viral RNA can be detected in blood plasma or nucleated blood cells respectively [63,64]. At least three studies have correlated plasma viral RNA with disease severity for HCPS and HFRS, suggesting that the replication of the virus plays an ongoing and real-time role in viral pathogenesis [65,66,67]. Several hallmark pathologic changes have been identified that occur in both HFRS and HCPS. A critical feature of both is a transient (~ 1–5 days) capillary leak involving the kidney and retroperitoneal space in HFRS and the lungs in HCPS. The resulting leakage is exudative in character, with chemical composition high in protein and resembling plasma.
3.1. Pathology
The continued experience indicating the strong tissue tropism for endothelial cells, specifically, is among the several factors that make β3 integrin an especially attractive candidate as an important in vivo receptor for hantaviruses. It is likely that hantaviruses arrive at their target tissues through uptake by regional lymph nodes, perhaps with or within an escorting lung histiocyte. The virus seeds local endothelium, where the first few infected cells give rise, ultimately, to a primary viremia, a process that appears to take a long time for hantavirus infections [62,63].

{kind=link}
Table 2.
Similarities and differences between HFRS and HCPS.
| HFRS | HCPS | |
| Transmission | Aerosolized wild-rodent excreta, no arthropod vector | Aerosolized wild-rodent excreta, no arthropod vector |
| Stages | Incubation period: 1–5 wk, prodrome 5–10 d, hypotensive phase 1–3 d oliguric phage 3–5 d convalescence | Incubation period: 1–5 wk, prodrome (febrile phase) 3–10 d, cardiorespiratoryphase 1–6 d diuretic phase 1–3 d convalescence |
| Primary organ of attack | Kidney, lymphoreticular system | Lungs, lymphoreticular system |
| Location of effusion | Retroperitoneal space | Thoracic bed; pulmonary interstitium |
| Pathologic hallmarks | Capillary leak. Mononuclear infiltrate. | Capillary leak. Interstitial mononuclear cell infiltrate. |
| Clinical findings | Fever, chills, nausea and vomiting, headache, lethargy, oliguria, tachycardia, facial and truncal flushing, hemorrhage, occasionally noncardiogenic pulmonary edema with tachypnea, shortness of breath, shock, death in 0.1 to 5%. | Fever, chills, nausea and vomiting, headache, lethargy, dyspnea, tachycardia, tachypnea, shortness of breath, noncardiogenic pulmonary edema, hemorrhage/petechiae (ANDV), shock, death in 10 to 50%. |
| Laboratory findings | Hypoalbuminemia, azotemia, proteinuria, hematuria, leukopenia/leukocytosis | Hypoxia, Hypoalbuminemia, thrombocytopenia, leukopenia/leukocytosis, hemoconcentration |
| Treatment | Specific: ribavirin. Nonspecific: cardioventilatory support, dialysis | Specific: none. Nonspecific: extracorporeal membrane oxygenation (ECMO), cardioventilatory support |
By the time that secondary viremia emerges, the agents of the more severe forms of HFRS and HCPS have begun to achieve sufficient mass as to induce, through PAMP-PRR interactions and other means, the expression of proinflammatory cytokines [64]. For HCPS, that expression favors the pulmonary bed and lymphoid organs, yet, for unknown reasons, spares the retroperitoneum and, in general, the kidney. In HFRS the situation is reversed, and yet it is often not appreciated that the expected preferential tissue tropism of HFRS-associated viruses and their HCPS-associated counterparts for the renal and pulmonary beds, respectively, is not as one would predict through the manifestations of the two diseases.
Local elaboration of inflammatory and chemotactic mediators is considered to be a requirement for the development of systemic disease symptoms, with those abnormalities sometimes culminating in shock and death. Yet it is not hypoxemia, due to the prominent pulmonary edema, that leads to death in most fatal cases of HCPS, but rather intoxication of the heart by as-yet-undefined mediators that leads to the low cardiac output state and the associated shock syndrome [64,65]. It is tempting to speculate that mediators produced in the lung in connection with the inflammatory infiltrate can percolate through the coronary circulation with minimal dilution in HCPS, a disadvantageous consequence of the close anatomic juxtaposition of the two organs. Thus, at least three classes of potential mechanisms, some overlapping and all certainly nonexclusive of the others, could be presumed to underlie the pathogenesis of HCPS. These include:
(1) Innate immune mechanisms. The nature of interactions between hantavirus pathogen-associated molecular patterns (PAMP) with the pattern recognition receptors (PRR) of susceptible endothelial cells are beginning to be clarified. The prototypical HTNV appears to be recognized by TLR-3 [43]. Such an infection has consequences such as increased expression of HLA-DR in dendritic cells [66] and differentiation of monocytes toward dendritic cells [67].
(2) Direct viral effects. The observed correlation between viral load and disease severity leaves the possibility open that hantavirus particles or RNA can themselves have toxic effects on cells or on signaling. Some investigators have favored direct viral toxicity, acting through the inhibition of endothelial cell barrier function, as an explanation for much of the capillary leak, although there is widespread agreement that multiple mechanisms that mediate pathogenesis likely operate simultaneously in the affected patient [68]. A potentially important clue toward the mechanism by which hantavirus infections deplete blood platelets and, in some cases cause hemorrhagic manifestations, was advanced by the recent discovery that pathogenic hantaviruses are able to recruit platelets to adhere to endothelial cell surfaces, with β3 integrin used as a critical binding element [69].
(3) Pathogenic effects caused by the activities of specific viral macromolecules. We have reviewed some of the activities associated with the Gn, Gc and N, virally-encoded polypeptides in previous sections.
Testing models of pathogenesis can be done more effectively when there is an animal model that mimics key aspects of the disease. There is no such model that closely mimics HFRS, but animal models exist for both the asymptomatic carriage of PUUV and SNV by their native carrier rodents, the bank vole Myodes glareolus and the deer mouse P. maniculatus; as well as a Syrian hamster model using ANDV or the related Maporal virus from Venezuela, for which an HCPS-mimetic disease is observed [70,71,72,73].
The ANDV-Syrian hamster model has a number of features in common with the human disease, as well as some differences. Unlike the neurologic diseases that have been possible to elicit with HTNV, the hamster model for HCPS appears to be caused by capillary leak that results in pulmonary edema and the production of a pleural effusion with exudative characteristics. Typically the hamsters die between 11 and 14-d post-inoculation, reflecting a slightly accelerated incubation period in comparison to human infections. As with human HCPS, the microscopic examination of the lung reveals abundant fibrin deposition, thickened alveolar septa, and viral antigen expressed abundantly in the microvascular endothelium. ANDV-infected hamsters fitted with physiologic monitoring devices exhibited diminished pulse pressures, tachycardia, and hypotension that appear to closely mimic the shock that is believed to be the proximate cause of demise in patients who succumb to HCPS [65,74].
Compared to the human disease, ANDV-infected hamsters exhibit exceptionally high titers of live ANDV in their tissues, with much of the viral replication occurring in hepatocytes, which are spared in the human disease. Titers of live ANDV in some cases exceed 108/g, whereas hantavirus isolates from human tissues have been notoriously difficult to obtain. Despite the universal occurrence of mildly-elevated hepatic enzymes in patients with HCPS, hepatic enzymes do not appear to be present at elevated levels in the blood of diseased hamsters even immediately before death [75].
3.2. Adaptive Immune Responses
The protracted incubation period associated with hantavirus disease gives the host considerable time to mount a mature immune response against the virus. Thus, in contradistinction to infections of comparable severity and related symptomatology associated with arenaviruses and filoviruses, hantavirus infections of humans are associated with antibody responses of significant titer by the time symptoms commence. Despite this observation, it appears to be possible that natural variation in individual neutralizing antibody responses among patients with SNV infections can be linked to disease severity, suggesting that administration of antiviral antibodies could prove effective therapeutically [76]. In the case of ANDV infection, new evidence has emerged indicating that the apparent clearance of the virus from the blood does not result in the complete removal of antigenic stimulus by the virus, suggesting that the virus may persist, perhaps in some as-yet undetermined immunologically privileged site [77].
A role for T cell-mediated pathological responses in HFRS and HCPS has been the source of speculation for a variety of reasons. The severity of SNV-associated HCPS may have made it more apparent that the onset of pulmonary edema, tachycardia and hypertension seemed to be all but universally temporally associated with the appearance of a spectrum of highly-activated cells of the lymphoid lineage in the peripheral blood. Cells with a close morphologic similarity to these “immunoblasts” were detected in the congested, heavy lungs of patients who came to autopsy, as well as in lymphoid organs and in the portal triads [63,78,79,80]. These observations led to speculation that some component of hantavirus pathogenesis could be linked to the appearance of antiviral T cells that could stimulate or contribute to the appearance of a “storm” of mediators and the associated capillary leak phenotype. Subsequent studies have borne out the expectation that a significant fraction of the immunoblast population in patients with HCPS are T cells with specificity for specific class I HLA-presented epitopes of viral antigens, including Gn, Gc and N [77,81,82,83]. Presumably, the antiviral activities of such cells, manifested in part through their elaboration of mediators in the affected interstitium, can contribute to the endothelial/capillary leak that lies at the heart of hantavirus pathogenesis.
Because early cases of HCPS often came to autopsy, it became possible to examine necropsied tissues for expression of cytokines. The study by Mori et al. (1999) revealed high relative expression of proinflammatory cytokines including TNFα, IL-1β, IL-6, providing evidence in favor of a “cytokine storm” model for pathogenesis [64]. The authors believed, based on the morphology of cytokine-secreting cells, that both monocytes and lymphocytes were contributing to the production of cytokines. That proinflammatory mediators are found in elevated levels in the plasma as well as the renal interstitium of patients with acute hantaviral illness has been recognized for some time as well [84,85].
While diagnosis of HCPS as well as HFRS is best accomplished with IgM serology, in the acute stage of SNV infection, RT-PCR can also be used if blood cells or blood clot are used instead of plasma or serum, where sensitivity even using nested PCR primers drops to about 70% [86,87,88]. In a facility at which many cases of HCPS are treated, the University of New Mexico medical center in Albuquerque, a diagnostic service has long been offered in which the patient’s hematologic findings are analyzed to establish the probability that a patient has HCPS. The combination of thrombocytopenia, elevated abundance of “immunoblast” lymphocytes, left-shifted polymorphonuclear cell population without strong morphologic evidence for their activation, and elevated hemoglobin or hematocrit values is highly specific for HCPS and allows clinicians the ability to put presumptive-HCPS patients on extracorporeal membrane oxygenation (ECMO), which is believed to have saved many patients from a lethal outcome [89].
4. Epidemiology and Epizootiology
4.1. Epidemiology and Transmission Mechanisms from Rodents to Humans
Human infection by hantaviruses is thought to follow contact with secretions or excretions produced by infected rodents. In the United States, 538 human infections by hantavirus were reported through late December 2009 [90], with New Mexico, Arizona and Colorado exhibiting the highest case-loads. While the prototypical central American hantavirus in central America was Rio Segundo virus of Reithrodontomys mexicanus from Costa Rica, the first human disease appeared some years later in Panama, where Choclo virus (CHOV) arose as the etiologic agent and is believed to be responsible for all known cases of HCPS. The fulvous pygmy rice rat Oligoryzomys fulvescens has been identified as the rodent reservoir [91]. In Panama, the first cases of HCPS, albeit with little or no evident cardiac involvement, were reported in 1999, and since then, 106 human infections have occurred with a 26% mortality rate [92]. Serosurveys of mammals in Mexico and Costa Rica have found anti-hantavirus antibodies [93,94,95,96], and seroprevalences ranging between 0.6 to 1.6% in human populations were reported despite the absence of known HCPS cases [97]. In South America, HCPS cases have been indentified in Argentina, Bolivia, Brazil, Chile, Paraguay and Uruguay, and evidence for human exposure to hantaviruses have also been reported in Venezuela [98] and Perú [99]. In southern South America, ANDV is the main etiologic agent with cases in Chile and Argentina reported since 1995. In Chile, 671 cases of HCPS due to ANDV have occurred during the period 2001–2009 [100]. Since 1995, more than 1,000 HCPS cases have been reported in Argentina [101]; in Brazil, approximately 1,100 HCPS cases have been identified between 1993 and 2008 [102]. Case-fatality ratios in those three countries have been similar, ranging from 30% (Argentina), 36% (Chile) and 39% (Brazil).
Hantavirus infections occur more frequently in men than women, although the male/female ratio is highly variable. For example, Panamanian communities showed a ratio of 55 men to 45 women [103], while in Chile the ratio is more biased to males (71%) [104]. In the Paraguayan Chaco the male-female ratio approaches 50% [105]. In North America, by December 2009 63% of case-patients were males [90]. All ethnic and racial groups seem to be susceptible to hantavirus infections, and the differences between certain groups (as indigenous and non-indigenous) are more likely correlated with the type habitat where the population resides (e.g., rural versus urban areas). In fact, rural communities account for the highest hantavirus incidences overall and are therefore at higher risk [92,105,106,107,108,109,110,111], although the importance of peridomestic settings as a major area of exposure has also been emphasized [112,113].
The main mechanism by which humans acquire hantavirus infection is by exposure to aerosols of contaminated rodent feces, urine, and saliva [114,115]. This can occur when humans reside in areas in close proximity to those that rodents inhabit, live in areas infested with rodents, or when rodents invade human settings, which are more frequent in rural habitats. There is a long history of human co-existence with rodents, raising questions about the apparent recent increases in hantavirus-related illnesses, especially HCPS. Other than an apparent association with El Niño southern oscillation (ENSO) events in some regions [116,117], the recent increases in incidence of HCPS do not seem to follow a readily-defined temporal or spatial pattern. However, some landscape features such as habitat fragmentation or human-disturbed areas may influence rodent population dynamics and impact viral incidence [118,119,120,121]. Despite the stochasticity associated with contraction of hantavirus infection, certain scenarios have been recognized as posing higher risk. Human activities in poorly ventilated buildings that aerosolize particulates that are then inhaled (i.e., cleaning, shaking rugs, dusting) are frequently identified among patients admitted for HCPS [11,122]. Outdoor activities are thought to convey lower risk due to lability of hantaviruses to UV radiation and the presumed tendency to be dispersed in wind, although certain environmental conditions seem to maintain the virus for longer periods outside its natural host allowing for indirect transmission [123]. An alternative but uncommon route of virus transmission is by rodent bites [124,125,126]. Field workers handling mammals are potentially at higher risk of exposure with hantavirus infections, although when quantified through serosurveys the absolute risk appears rather slight [127]. A new study in Colorado suggests the possibility that a rodent bite may have been the proximate vehicle for outdoor transmission of SNV [128], which re-emphasizes the use of personal protective equipment during field work activities [129]. As a particular case within hantaviruses, person-to-person transmission has exclusively been documented for the South American Andes virus [130,131,132,133,134,135]. The identification of this transmission route has been made using both molecular tools and epidemiological surveys, but the mechanism of interpersonal transmission is not well established. Recent findings show that family clusters and specifically sexual partners share the greater risk of interpersonal transmission, although sexual transmission per se can be neither inferred nor refuted presently [130,135]. Interestingly, ANDV may also be shed by humans through other biological fluids such as urine [136], illustrating the particular properties that differentiate this virus from other hantaviruses. Although interpersonal transmission seems to be unique for ANDV, viral RNA of PUUV has been detected in saliva of patients with HFRS, and some patients with SNV-HCPS have viral RNA in tracheal secretions [88,137].
4.2. Rodent Reservoirs and Cross-species Transmission
Hantaviruses in the Americas are naturally hosted by rodents (Muridae and Cricetidae) as well as shrews (Soricidae) and moles (Talpidae) (Figure 1). Three shrew and one mole species have been reported to host hantaviruses and their pathogenicity for humans remains unknown [22,138,139]. At least 15 rodent species have been identified as carriers of different pathogenic hantaviruses, with some South American genotypes such as Castelo do Sonhos (CDSV) or Hu39694 only identified after human infections (Figure 1). Hantaviruses typically show high species-specificity and no intermediate host [140]. However, some hantavirus genotypes have been described in the same rodent species. Such is the case of Playa de Oro (OROV) and Catacamas (CATV) identified in Oryzomys couesi [141,142], or Maporal (MAPV) and Choclo (CHOV) hosted by O. fulvescens [91,143]. In North America both Muleshoe and Black Creek Canal hantaviruses have been detected in geographically-distant Sigmodon hispidus [144,145]. Also, one hantavirus genotype (e.g., Juquitiba-like virus) may be carried by more than one rodent species (O. nigripes, Oxymycterus judex, Akodon montesis). Another example is Laguna Negra virus (LANV) which after being identified in Calomys laucha [146] has also been reported in C. callosus [147]. The rapid increase in the discovery of new hantaviruses and the identification of their hosts does not seem likely to end soon as new small mammal species are screened [95]. This subject is complicated by continued controversy in the criteria for the classification of distinct hantaviruses [148,149], which is also tied to host taxonomic classification and taxonomic rearrangements.
Cross-species transmission is a major process during spread, emergence, and evolution of RNA viruses [6,150]. Particularly within hantaviruses, spillover to secondary hosts are increasingly identified as more extensive studies are performed [151,152,153,154,155,156]. For example, ANDV is the predominant etiologic agent of HCPS in South America, and O. longicaudatus the main rodent reservoir. Spillover in at least four other rodent species that co-occur with the reservoir have been identified, with Abrothrix longipilis showing the second higher prevalence to ANDV-antibodies, and there is presently no question that the virus is extremely similar genetically between the two host rodents [157,158]. In North America, spillover of Bayou virus (BAYV) may have occurred from the main reservoir O. palustris to S. hispidus, R. fulvescens, P. leucopus, and B. taylori [159,160,161]. Hantavirus spillover is more likely to occur with host populations inhabiting sympatric or syntopic regions [151,162], and cross-species transmission would presumably have greater chances of success if the host species are closely related [163]. An interesting exception is found between Oxbow virus (OXBV) and Asama virus (ASAV) in which a host-switch process seemed to have occurred between mammals belonging to two families (Talpidae and Soricidae), likely as a result of alternating and recurrent co-divergence of certain taxa through evolutionary time [138].
4.3. Outbreaks
Hantaviruses are horizontally transmitted between rodents and are not transmitted by arthropods (unlike other viruses of the family Bunyaviridae). Spillover infection to nonhuman mammals usually results in no onward (or “dead-end”) transmission, but if humans are infected may result in high morbidity and mortality [122,164]. During the spring of 1993, an outbreak of patients with HCPS due to SNV occurred in the Four Corners states resulting in more than 60% case-fatality among the initial cases, many involving members of the Navajo tribe [12,121]. In Panama, an outbreak was reported during 1999–2000 in Los Santos, and 12 cases where identified with three fatalities [165,166]. This represented the first report of human hantavirus infections in Central America. In South America, the first largest identified outbreak occurred in the Chaco region in northwestern Paraguay during 1995–1996. Seventeen individuals were identified with SNV antibody (ELISA) or were antigen (IHC) positive out of 52 suspected cases [167]. Major outbreaks due to ANDV occurred in 1996 in southern Argentina [131,134]; in southern Chile clusters of patients presented with hantavirus illness in 1997 [158]. In Brazil, the first outbreak was identified in the Brazilian Amazon (Maranhão State) in 2000, and involved small villages that resulted in a 13.3% prevalence of those tested (398 total residents) [168].
The factors that trigger hantavirus outbreaks are still poorly understood, probably because they result from several interacting biotic and abiotic features whose key parameters are difficult to model. However, the use of new modeling approaches that involve geographical and environmental features seem to be promising in predicting potential hantavirus outbreaks and/or areas of higher risk [169,170,171,172]. Because hantaviruses are known to be directly transmitted from infected to susceptible hosts, the first natural approach is to relate outbreaks to the ecology of the viral hosts. Hantavirus transmission and persistence in rodent populations depends on several factors that interact to affect ecological dynamics of the host, which in turn is strongly influenced by the behavioral characteristics of individual rodent species, to landscape structure, and environmental features [173,174]. Viral transmission depends on contact rates among susceptible hosts, and despite the prevailing notion that a higher density increases encounters and hence secondary infected hosts, contrasting patterns relating rodent population size and virus prevalence can be found [175]. In addition, it has been shown that SNV transmission follows a contact heterogeneity pattern, where individuals in the population have different probability of transmitting the infection [176]. The understanding of viral transmission proves to be far more complex when species other than the main reservoir host are incorporated in the model. In fact, recent studies have shown that higher hosts species diversity is correlated with lower infection prevalence in North America for P. maniculatus [177], in Central America for O. fulvescens (reservoir of Choclo virus) and Zygodontomys brevicauda (reservoir of Calabazo virus) [178], and in South America for Akodon montensis (reservoir of Jabora virus) [162]. Contact rates vary according to the spatial distribution of populations and seem to be strongly influenced by landscape structure. For example, SNV prevalence in P. maniculatus was higher in landscapes with a higher level of fragmentation of the preferred habitat [179]. In addition, certain properties of the landscape such as elevation, slope, and land cover seem to be useful in detecting areas with persistent SNV infections, and therefore thought to be refugial areas where the virus can be maintained for years [169]. Changes in the natural environment of reservoir species, such as forest fragmentation and habitat loss, may alter population abundance and distribution and lead to hantavirus outbreaks, as observed in the Azurero Peninsula of Panama [118,119]. Also, differences in the microhabitat, including overstory cover, may lead to differences in the ecological dynamics within populations and affect the rate of exposure to the virus [180]. Differences in hantavirus infections through contrasting landscapes in the latitudinal span have been found in rodent populations of O. longicaudatus in Chile, suggesting that humans are differentially exposed to the virus [107,181].
Rodent population dynamics are affected by seasonal changes of weather and climate [182,183]. In the case of the ENSO-associated outbreaks, a complex cascade of events triggered by highly unusual rains in the precedent year have been postulated to result in an increase of primary production and rodent densities, also increasing the likelihood of transmission of the virus to humans, but it has proved difficult to precisely demonstrate the suggested intermediate events such as increased rodent densities in the increased caseload [116,121,184]. In South America, effects of climate change and hantavirus outbreaks have not been well studied, despite the knowledge that several rodents species that are reservoirs of emerging diseases have dramatically been affected by events like El Niño [185]. Changes in host population dynamics are also affected by seasonality, which may lead to disease outbreaks when processes that equilibrate rodent populations from season to season are interrupted [186].
Viral emergence may continue to be promoted as human-introduced changes continue to increase in the environment at different geographical scales. Human incursions into previously uncultivated environments may lead to new contacts between rodent reservoirs and humans, increasing the likelihood of contracting infections [187]. These changes may also alter rodent’s population structure and dynamics and interspecies interactions creating conditions that may lead to viral outbreaks, viral establishment in new hosts, and emergence of HCPS [102,162], even with seemingly slight ecological disturbance to the virus-host system [188].
5. Concluding Remarks
Certain pathophysiologic characteristics, including thrombocytopenia and shock, of hantavirus diseases of humans, bear substantial similarity to the hemorrhagic fevers induced by other viruses such arenaviruses, filoviruses and flaviviruses, despite sharing essentially no sequence similarities therewith. Such observations raise questions about whether such commonalities in pathogenesis are chance similarities of phenotype, or instead report the presence of common molecular mechanisms among the viruses.
In this review we discuss the general properties, discoveries and epidemiology/ecology of the New World forms of pathogenic hantaviruses, and also seek to identify some of the characteristics of the viral macromolecules and immunologic mechanisms that have been proposed as potential direct mediators of the pathogenic events that characterize the human disease HCPS. While it is unlikely that expression of any particular viral protein or RNAs in isolation can be relied upon to replicate key phenotypes of infection by the complete virus, some of the findings have been sufficiently consistent with what is known of the pathogenesis in vivo that they offer plausible first-pass leads in the search for therapeutic targets. We look forward to the mechanistic revelations that will follow the inevitably expanded usage of powerful methods such as deep sequencing, ever-more advanced imaging, and microscopic methods, and animal models that can at last be said to be close mimics of human hantavirus disease.
Acknowledgments
Financial support was provided by the Fogarty International Center Research Grant # D43 TW007131, and the NIH-ICIDR Chilean Hantavirus Grant 1 U19 AI45452-01.
References and Notes
- Holmes, E.; Drummond, A. The Evolutionary Genetics of Viral Emergence. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Childs, J.E., Mackenzie, J.S., Richt, J.A., Eds.; Springer-Verlag Berlin Heidelberg: New York, NY, USA, 2007; pp. 51–66. [Google Scholar]
- Childs, J.E.; Mackenzie, J.S.; Richt, J.A. Introduction: Conceptualizing and Partitioning the Emergence Process of Zoonotic Viruses from Wildlife to Humans. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Childs, J.E., Mackenzie, J.S., Richt, J.A., Eds.; Springer-Verlag Berlin Heidelberg: New York, NY, USA, 2007; pp. 1–31. [Google Scholar]
- Morse, S.S. Factors in the emergence of infectious diseases. Emerg. Infect. Dis. 1995, 1, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Woolhouse, M.E.; Gowtage-Sequeria, S. Host range and emerging and reemerging pathogens. Emerg. Infect. Dis. 2005, 11, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.H.; Latham, S.M.; Woolhouse, M.E. Risk factors for human disease emergence. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 983–989. [Google Scholar] [CrossRef]
- Holmes, E.C. The Evolutionary Genetics of Emerging Viruses. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 353–372. [Google Scholar] [CrossRef]
- Racaniello, V.R. Emerging infectious diseases. J. Clin. Invest. 2004, 113, 796–798. [Google Scholar] [CrossRef]
- Fraser, C.; Donnelly, C.A.; Cauchemez, S.; Hanage, W.P.; Van Kerkhove, M.D.; Hollingsworth, T.D.; Griffin, J.; Baggaley, R.F.; Jenkins, H.E.; Lyons, E.J.; et al. Pandemic Potential of a Strain of Influenza A (H1N1): Early Findings. Science 2009, 324, 1557–1561. [Google Scholar] [CrossRef]
- Mills, J.N.; Childs, J.E. Ecologic studies of rodent reservoirs: Their relevance for human health. Emerg. Infect. Dis. 1998, 4, 529–537. [Google Scholar] [CrossRef]
- Zizi, M.; Heyman, P.; Vandenvelde, C. The assessment of human health risks from rodent-borne diseases by means of ecological studies of rodent reservoirs. Mil. Med. 2002, 167, 70–73. [Google Scholar]
- Hjelle, B.; Torres-Pérez, F. Rodent-Borne Viruses. In Clinical Virology Manual, 4th ed.; Specter, S., Hodinka, R., Wiedbrauk, D., Young, S., Eds.; America Society for Microbiology Press: Washington, DC, USA, 2009; pp. 641–658. [Google Scholar]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar] [CrossRef]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef]
- Lee, H.W. Hemorrhagic fever with renal syndrome. History of Hantaan virus and epidemiological features. Scand. J. Infect. Dis. 1982, 36, 82–85. [Google Scholar]
- Lee, H.W. Hemorrhagic fever with renal syndrome in Korea. Rev. Infect. Dis. 1989, 11 (Suppl. 4), S864–S876. [Google Scholar] [CrossRef] [PubMed]
- Smadel, J.E. Epidemic hemorrhagic fever. Am. J. Public Health Nations Health 1953, 43, 1327–1330. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Lee, P.W. Korean hemorrhagic fever. I. Demonstration of causative antigen and antibodies. Korean J. Intern. Med. 1976, 19, 371–394. [Google Scholar]
- Lee, H.W.; Lee, P.W.; Johnson, K.M. Isolation of the etiologic agent of Korean Hemorrhagic fever. J. Infect. Dis. 1978, 137, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Schmaljohn, C.S.; Hasty, S.E.; Dalrymple, J.M.; Leduc, J.W.; Lee, H.W.; von Bonsdorff, C.H.; Brummer-Korvenkontio, M.; Vaheri, A.; Tsai, T.F.; Regnery, H.L. Antigenic and genetic properties of viruses linked to hemorrhagic fever with renal syndrome. Science 1985, 227, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, R.F.; von Bonsdorff, C.H. Ribonucleoproteins of Uukuniemi virus are circular. J. Virol. 1975, 15, 386–392. [Google Scholar] [CrossRef]
- Arai, S.; Ohdachi, S.D.; Asakawa, M.; Kang, H.J.; Mocz, G.; Arikawa, J.; Okabe, N.; Yanagihara, R. Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides). Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 16296–16301. [Google Scholar] [CrossRef]
- Arai, S.; Song, J.W.; Sumibcay, L.; Bennett, S.N.; Nerurkar, V.R.; Parmenter, C.; Cook, J.A.; Yates, T.L.; Yanagihara, R. Hantavirus in northern short-tailed shrew, United States. Emerg. Infect. Dis. 2007, 13, 1420–1423. [Google Scholar] [CrossRef]
- Schmaljohn, C.S.; Schmaljohn, A.L.; Dalrymple, J.M. Hantaan virus M RNA: Coding strategy, nucleotide sequence, and gene order. Virology 1987, 157, 31–39. [Google Scholar] [CrossRef]
- Lober, C.; Anheier, B.; Lindow, S.; Klenk, H.D.; Feldmann, H. The Hantaan virus glycoprotein precursor is cleaved at the conserved pentapeptide WAASA. Virology 2001, 289, 224–229. [Google Scholar] [CrossRef]
- Pensiero, M.N.; Hay, J. The Hantaan virus M-segment glycoproteins G1 and G2 can be expressed independently. J. Virol. 1992, 66, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Antic, D.; Wright, K.E.; Kang, C.Y. Maturation of Hantaan virus glycoproteins G1 and G2. Virology 1992, 189, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Ruusala, A.; Persson, R.; Schmaljohn, C.S.; Pettersson, R.F. Coexpression of the membrane glycoproteins G1 and G2 of Hantaan virus is required for targeting to the Golgi complex. Virology 1992, 186, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Elliott, R.M. Golgi localization of Hantaan virus glycoproteins requires coexpression of G1 and G2. Virology 2002, 300, 31–38. [Google Scholar] [CrossRef]
- Spiropoulou, C.F.; Goldsmith, C.S.; Shoemaker, T.R.; Peters, C.J.; Compans, R.W. Sin Nombre virus glycoprotein trafficking. Virology 2003, 308, 48–63. [Google Scholar] [CrossRef]
- Gavrilovskaya, I.N.; Brown, E.J.; Ginsberg, M.H.; Mackow, E.R. Cellular entry of hantaviruses which cause hemorrhagic fever with renal syndrome is mediated by beta(3) integrins. J. Virol. 1999, 73, 3951–3959. [Google Scholar] [CrossRef]
- Krautkramer, E.; Zeier, M. Hantavirus causing hemorrhagic fever with renal syndrome enters from the apical surface and requires decay-accelerating factor (DAF/CD55). J. Virol. 2008, 82, 4257–4264. [Google Scholar] [CrossRef]
- Choi, Y.; Kwon, Y.C.; Kim, S.I.; Park, J.M.; Lee, K.H.; Ahn, B.Y. A hantavirus causing hemorrhagic fever with renal syndrome requires gC1qR/p32 for efficient cell binding and infection. Virology 2008, 381, 178–183. [Google Scholar] [CrossRef]
- Hepojoki, J.; Strandin, T.; Vaheri, A.; Lankinen, H. Interactions and oligomerization of hantavirus glycoproteins. J. Virol. 2010, 84, 227–242. [Google Scholar] [CrossRef]
- Tischler, N.D.; Gonzalez, A.; Perez-Acle, T.; Rosemblatt, M.; Valenzuela, P.D.T. Hantavirus Gc glycoprotein: Evidence for a class II fusion protein. J. Gen. Virol. 2005, 86, 2937–2947. [Google Scholar] [CrossRef] [PubMed]
- Tischler, N.D.; Rosemblatt, M.; Valenzuela, P.D. Characterization of cross-reactive and serotype-specific epitopes on the nucleocapsid proteins of hantaviruses. Virus Res. 2008, 135, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Groen, J.; Gerding, M.; Koeman, J.P.; Roholl, P.J.; van Amerongen, G.; Jordans, H.G.; Niesters, H.G.; Osterhaus, A.D. A macaque model for hantavirus infection. J. Infect. Dis. 1995, 172, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Sen, A.; Mackow, E.R. Degrons at the C terminus of the pathogenic but not the nonpathogenic hantavirus G1 tail direct proteasomal degradation. J. Virol. 2007, 81, 4323–4330. [Google Scholar] [CrossRef]
- Geimonen, E.; Fernandez, I.; Gavrilovskaya, I.N.; Mackow, E.R. Tyrosine residues direct the ubiquitination and degradation of the NY-1 hantavirus G1 cytoplasmic tail. J. Virol. 2003, 77, 10760–10768. [Google Scholar] [CrossRef]
- Wang, H.; Strandin, T.; Hepojoki, J.; Lankinen, H.; Vaheri, A. Degradation and aggresome formation of the Gn tail of the apathogenic Tula hantavirus. J. Gen. Virol. 2009, 90, 2995–3001. [Google Scholar] [CrossRef]
- Geimonen, E.; Neff, S.; Raymond, T.; Kocer, S.S.; Gavrilovskaya, I.N.; Mackow, E.R. Pathogenic and nonpathogenic hantaviruses differentially regulate endothelial cell responses. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 13837–13842. [Google Scholar] [CrossRef]
- Kraus, A.A.; Raftery, M.J.; Giese, T.; Ulrich, R.; Zawatzky, R.; Hippenstiel, S.; Suttorp, N.; Kruger, D.H.; Schonrich, G. Differential antiviral response of endothelial cells after infection with pathogenic and nonpathogenic hantaviruses. J. Virol. 2004, 78, 6143–6150. [Google Scholar] [CrossRef]
- Spiropoulou, C.F.; Albarino, C.G.; Ksiazek, T.G.; Rollin, P.E. Andes and Prospect Hill hantaviruses differ in early induction of interferon although both can downregulate interferon signaling. J. Virol. 2007, 81, 2769–2776. [Google Scholar] [CrossRef]
- Handke, W.; Oelschlegel, R.; Franke, R.; Kruger, D.H.; Rang, A. Hantaan virus triggers TLR3-dependent innate immune responses. J. Immunol. 2009, 182, 2849–2858. [Google Scholar] [CrossRef]
- Prescott, J.; Ye, C.Y.; Sen, G.; Hjelle, B. Induction of innate immune response genes by Sin Nombre hantavirus does not require viral replication. J. Virol. 2005, 79, 15007–15015. [Google Scholar] [CrossRef] [PubMed]
- Alff, P.J.; Gavrilovskaya, I.N.; Gorbunova, E.; Endriss, K.; Chong, Y.S.; Geimonen, E.; Sen, N.; Reich, N.C.; Mackow, E.R. The pathogenic NY-1 hantavirus G1 cytoplasmic tail inhibits RIG-I- and TBK-1-directed interferon responses. J. Virol. 2006, 80, 9676–9686. [Google Scholar] [CrossRef]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef]
- Gavrilovskaya, I.N.; Peresleni, T.; Geimonen, E.; Mackow, E.R. Pathogenic hantaviruses selectively inhibit beta(3) integrin directed endothelial cell migration. Arch. Virol. 2002, 147, 1913–1931. [Google Scholar] [CrossRef] [PubMed]
- Gavrilovskaya, I.N.; Gorbunova, E.E.; Mackow, N.A.; Mackow, E.R. Hantaviruses Direct Endothelial Cell Permeability by Sensitizing Cells to the Vascular Permeability Factor, VEGF, while Angiopoietin-1 and Sphingosine 1-Phosphate inhibit Hantavirus-directed Permeability. J. Virol. 2008, 82, 5797–5806. [Google Scholar] [CrossRef]
- Severson, W.; Partin, L.; Schmaljohn, C.S.; Johnsson, C.B. Characterization of the hantaan nucleocapsid protein-ribonucleic acid interaction. J. Biol. Chem. 1999, 274, 33732–33739. [Google Scholar] [CrossRef]
- Mir, M.A.; Panganiban, A.T. Trimeric hantavirus nucleocapsid protein binds specifically to the viral RNA panhandle. J. Virol. 2004, 78, 8281–8288. [Google Scholar] [CrossRef] [PubMed]
- Kaukinen, P.; Vaheri, A.; Plyusnin, A. Hantavirus nucleocapsid protein: A multifunctional molecule with both housekeeping and ambassadorial duties. Arch. Virol. 2005, 150, 1693–1713. [Google Scholar] [CrossRef]
- Ramanathan, H.N.; Chung, D.H.; Plane, S.J.; Sztul, E.; Chu, Y.K.; Guttieri, M.C.; McDowell, M.; Ali, G.; Jonsson, C.B. Dynein-dependent transport of the hantaan virus nucleocapsid protein to the endoplasmic reticulum-Golgi intermediate compartment. J. Virol. 2007, 81, 8634–8647. [Google Scholar] [CrossRef]
- Ravkov, E.V.; Nichol, S.T.; Peters, C.J.; Compans, R.W. Role of actin microfilaments in Black Creek Canal virus morphogenesis. J. Virol. 1998, 72, 2865–2870. [Google Scholar] [CrossRef]
- Kaukinen, P.; Vaheri, A.; Plyusnin, A. Non-covalent interaction between nucleocapsid protein of Tula hantavirus and small ubiquitin-related modifier-1, SUMO-1. Virus Res. 2003, 92, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.L.; Frias-Staheli, N.; Garcia-Sastre, A.; Schmaljohn, C.S. Hantaan virus nucleocapsid protein binds to importin alpha proteins and inhibits tumor necrosis factor alpha-induced activation of nuclear factor kappa B. J. Virol. 2009, 83, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Panganiban, A.T. A protein that replaces the entire cellular eIF4F complex. EMBO J. 2008, 27, 3129–3139. [Google Scholar] [CrossRef] [PubMed]
- Mir, M.A.; Panganiban, A.T. Characterization of the RNA chaperone activity of hantavirus nucleocapsid protein. J. Virol. 2006, 80, 6276–6285. [Google Scholar] [CrossRef] [PubMed]
- Panganiban, A.T.; Mir, M.A. Bunyavirus N: eIF4F surrogate and cap-guardian. Cell Cycle 2009, 8, 1332–1337. [Google Scholar] [CrossRef]
- Ramanathan, H.N.; Jonsson, C.B. New and Old World hantaviruses differentially utilize host cytoskeletal components during their life cycles. Virology 2008, 374, 138–150. [Google Scholar] [CrossRef]
- Buranda, T.; Wu, Y.; Perez, D.; Jett, S.D.; Bondu-Hawkins, V.; Ye, C.; Edwards, B.; Hall, P.; Larson, R.S.; Lopez, G.P.; Sklar, L.A.; Hjelle, B. Recognition of DAF and alpha(v)beta(3) by inactivated Hantaviruses, towards the development of HTS flow cytometry assays. Anal. Biochem. 2010, 402, 151–160. [Google Scholar] [CrossRef]
- Hjelle, B. Pathogenesis of hantavirus infections. In UpToDate; Rose, B.D., Ed.; UpToDate: Wellesley, MA, USA, 2008. [Google Scholar]
- Duchin, J.S.; Koster, F.T.; Peters, C.J.; Simpson, G.L.; Tempest, B.; Zaki, S.R.; Ksiazek, T.G.; Rollin, P.E.; Nichol, S.; Umland, E.T.; Moolenaar, R.L.; Reef, S.E.; Nolte, K.B.; Gallaher, M.M.; Butler, J.C.; Breiman, R.F.; Group, f.T.H.S. Hantavirus pulmonary syndrome: A clinical description of 17 patients with a newly recognized disease. N. Engl. J. Med. 1994, 330, 949–955. [Google Scholar] [CrossRef]
- Nolte, K.B.; Feddersen, R.M.; Foucar, K.; Zaki, S.R.; Koster, F.T.; Madar, D.; Merlin, T.L.; McFeeley, P.J.; Umland, E.T.; Zumwalt, R.E. Hantavirus pulmonary syndrome in the United States: A pathological description of a disease caused by a new agent. Hum. Pathol. 1995, 26, 110–120. [Google Scholar] [CrossRef]
- Mori, M.; Rothman, A.L.; Kurane, I.; Montoya, J.M.; Nolte, K.B.; Norman, J.E.; Waite, D.C.; Koster, F.T.; Ennis, F.A. High levels of cytokine-producing cells in the lung tissues of patients with fatal hantavirus pulmonary syndrome. J. Infect. Dis. 1999, 179, 295–302. [Google Scholar] [CrossRef]
- Campen, M.J.; Milazzo, M.L.; Fulhorst, C.F.; Obot Akata, C.J.; Koster, F. Characterization of shock in a hamster model of hantavirus infection. Virology 2006, 356, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Raftery, M.J.; Kraus, A.A.; Ulrich, R.; Kruger, D.H.; Schonrich, G. Hantavirus infection of dendritic cells. J. Virol. 2002, 76, 10724–10733. [Google Scholar] [CrossRef] [PubMed]
- Markotic, A.; Hensley, L.; Daddario, K.; Spik, K.; Anderson, K.; Schmaljohn, C. Pathogenic hantaviruses elicit different immunoreactions in THP-1 cells and primary monocytes and induce differentiation of human monocytes to dendritic-like cells. Coll. Antropol. 2007, 31, 1159–1167. [Google Scholar] [PubMed]
- Mackow, E.R.; Gavrilovskaya, I.N. Hantavirus regulation of endothelial cell functions. Thromb. Haemost. 2009, 102, 1030–1041. [Google Scholar]
- Gavrilovskaya, I.N.; Gorbunova, E.E.; Mackow, E.R. Pathogenic Hantaviruses Direct the Adherence of Quiescent Platelets to Infected Endothelial Cells. J. Virol. 2010, 84, 4832–4839. [Google Scholar] [CrossRef]
- Milazzo, M.L.; Eyzaguirre, E.J.; Molina, C.P.; Fulhorst, C.F. Maporal viral infection in the Syrian golden hamster: A model of hantavirus pulmonary syndrome. J. Infect. Dis. 2002, 186, 1390–1395. [Google Scholar] [CrossRef]
- Hooper, J.W.; Larsen, T.; Custer, D.M.; Schmaljohn, C.S. A lethal disease model for hantavirus pulmonary syndrome. Virology 2001, 289, 6–14. [Google Scholar] [CrossRef]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Bharadwaj, M.; Yee, J.; Ricci, R.; Feddersen, R.M.; Hjelle, B. Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus). Proc. Natl. Acad. Sci. U. S. A. 2000, 97, 10578–10583. [Google Scholar] [CrossRef]
- Lundkvist, A.; Cheng, Y.; Sjolander, K.B.; Niklasson, B.; Vaheri, A.; Plyusnin, A. Cell culture adaptation of Puumala hantavirus changes the infectivity for its natural reservoir, Clethrionomys glareolus, and leads to accumulation of mutants with altered genomic RNA S segment. J. Virol. 1997, 71, 9515–9523. [Google Scholar] [CrossRef]
- Mertz, G.J.; Hjelle, B.; Crowley, M.; Iwamoto, G.; Tomicic, V.; Vial, P.A. Diagnosis and treatment of new world hantavirus infections. Curr. Opin. Infect. Dis. 2006, 19, 437–442. [Google Scholar] [CrossRef]
- Wahl-Jensen, V.; Chapman, J.; Asher, L.; Fisher, R.; Zimmerman, M.; Larsen, T.; Hooper, J.W. Temporal Analysis of Andes Virus and Sin Nombre Virus Infection of Syrian Hamsters. J Virol. 2007, 81, 7449–7462. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, M.; Nofchissey, R.; Goade, D.; Koster, F.; Hjelle, B. Humoral immune responses in the hantavirus cardiopulmonary syndrome. J. Infect. Dis. 2000, 182, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Manigold, T.; Mori, A.; Graumann, R.; Llop, E.; Simon, V.; Ferres, M.; Valdivieso, F.; Castillo, C.; Hjelle, B.; Vial, P. Highly differentiated, resting gn-specific memory CD8 T cells persist years after infection by andes hantavirus. PLoS Pathog. 2010, 6, e1000779. [Google Scholar] [CrossRef] [PubMed]
- Zaki, S.R.; Greer, P.W.; Coffield, L.M.; Goldsmith, C.S.; Nolte, K.B.; Foucar, K.; Feddersen, R.M.; Zumwalt, R.E.; Miller, G.L.; Khan, A.S. Hantavirus pulmonary syndrome. Pathogenesis of an emerging infectious disease. Am. J. Pathol. 1995, 146, 552–579. [Google Scholar] [PubMed]
- Jenison, S.; Hjelle, B.; Simpson, S.; Hallin, G.; Feddersen, R.; Koster, F. Hantavirus pulmonary syndrome: Clinical, diagnostic, and virologic aspects. Semin. Respir. Infect. 1995, 10, 259–269. [Google Scholar] [PubMed]
- Koster, F.; Foucar, K.; Hjelle, B.; Scott, A.; Chong, Y.Y.; Larson, R.; McCabe, M. Rapid presumptive diagnosis of hantavirus cardiopulmonary syndrome by peripheral blood smear review. Am. J. Clin. Pathol. 2001, 116, 665–672. [Google Scholar] [CrossRef]
- Ennis, F.A.; Cruz, J.; Spiropoulou, C.F.; Waite, D.; Peters, C.J.; Nichol, S.T.; Kariwa, H.; Koster, F.T. Hantavirus pulmonary syndrome: CD8(+) and CD4(+) cytotoxic T lymphocytes to epitopes on Sin Nombre virus nucleocapsid protein isolated during acute illness. Virology 1997, 238, 380–390. [Google Scholar] [CrossRef]
- Kilpatrick, E.D.; Terajima, M.; Koster, F.T.; Catalina, M.D.; Cruz, J.; Ennis, F.A. Role of specific CD8(+) T cells in the severity of a fulminant zoonotic viral hemorrhagic fever, hantavirus pulmonary syndrome. J. Immunol. 2004, 172, 3297–3304. [Google Scholar] [CrossRef]
- Terajima, M.; Hayasaka, D.; Maeda, K.; Ennis, F.A. Immunopathogenesis of hantavirus pulmonary syndrome and hemorrhagic fever with renal syndrome: Do CD8(+) T cells trigger capillary leakage in viral hemorrhagic fevers? Immunol. Lett. 2007, 113, 117–120. [Google Scholar] [CrossRef]
- Linderholm, M.; Ahlm, C.; Settergren, B.; Waage, A.; Tarnvik, A. Elevated plasma levels of tumor necrosis factor (TNF)-alpha, soluble TNF receptors, interleukin (IL)-6, and IL-10 in patients with hemorrhagic fever with renal syndrome. J. Infect. Dis. 1996, 173, 38–43. [Google Scholar] [CrossRef]
- Temonen, M.; Mustonen, J.; Helin, H.; Pasternack, A.; Vaheri, A.; Holthofer, H. Cytokines, adhesion molecules, and cellular infiltration in nephropathia epidemica kidneys: An immunohistochemical study. Clin. Immunol. Immunopathol. 1996, 78, 47–55. [Google Scholar] [CrossRef]
- Hjelle, B.; Spiropoulou, C.F.; Torrez-Martinez, N.; Morzunov, S.; Peters, C.J.; Nichol, S.T. Detection of Muerto Canyon virus RNA in peripheral blood mononuclear cells from patients with hantavirus pulmonary syndrome. J. Infect. Dis. 1994, 170, 1013–1017. [Google Scholar] [CrossRef]
- Terajima, M.; Hendershot, J.D., III; Kariwa, H.; Koster, F.T.; Hjelle, B.; Goade, D.; DeFronzo, M.C.; Ennis, F.A. High levels of viremia in patients with the Hantavirus pulmonary syndrome. J. Infect. Dis. 1999, 180, 2030–2034. [Google Scholar] [CrossRef] [PubMed]
- Xiao, R.; Yang, S.; Koster, F.; Ye, C.; Stidley, C.; Hjelle, B. Sin Nombre viral RNA load in patients with hantavirus cardiopulmonary syndrome. J. Infect. Dis. 2006, 194, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Dietl, C.A.; Wernly, J.A.; Pett, S.B.; Yassin, S.F.; Sterling, J.P.; Dragan, R.; Milligan, K.; Crowley, M.R. Extracorporeal membrane oxygenation support improves survival of patients with severe Hantavirus cardiopulmonary syndrome. J. Thorac. Cardiovasc. Surg. 2008, 135, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Case Information: Hantavirus Pulmonary Syndrome Case Count and Descriptive Statistics. Available online: http://www.cdc.gov/ncidod/diseases/hanta/hps/noframes/caseinfo.htm (accessed on 12 November 2010).
- Vincent, M.J.; Quiroz, E.; Gracia, F.; Sanchez, A.J.; Ksiazek, T.G.; Kitsutani, P.T.; Ruedas, L.A.; Tinnin, D.S.; Caceres, L.; Garcia, A.; Rollin, P.E.; Mills, J.N.; Peters, C.J.; Nichol, S.T. Hantavirus pulmonary syndrome in Panama: Identification of novel Hantaviruses and their likely reservoirs. Virology 2000, 277, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Armien, A.G.; Armien, B.; Koster, F.; Pascale, J.M.; Avila, M.; Gonzalez, P.; de la Cruz, M.; Zaldivar, Y.; Mendoza, Y.; Gracia, F.; Hjelle, B.; Lee, S.J.; Yates, T.L.; Salazar-Bravo, J. Hantavirus infection and habitat associations among rodent populations in agroecosystems of Panama: Implications for human disease risk. Am. J. Trop. Med. Hyg. 2009, 81, 59–66. [Google Scholar] [CrossRef]
- Suzan, G.; Ceballos, G.; Mills, J.; Ksiazek, T.G.; Yates, T. Serologic evidence of hantavirus infection in sigmodontine rodents in Mexico. J. Wildl. Dis. 2001, 37, 391–393. [Google Scholar] [CrossRef]
- Mantooth, S.J.; Milazzo, M.L.; Bradley, R.D.; Hice, C.L.; Ceballos, G.; Tesh, R.B.; Fulhorst, C.F. Geographical distribution of rodent-associated hantaviruses in Texas. J. Vector Ecol. 2001, 26, 7–14. [Google Scholar]
- Castro-Arellano, I.; Suzan, G.; Leon, R.F.; Jimenez, R.M.; Lacher, T.E., Jr. Survey for antibody to hantaviruses in Tamaulipas, Mexico. J. Wildl. Dis. 2009, 45, 207–212. [Google Scholar] [CrossRef]
- Hjelle, B.; Anderson, B.; Torrez-Martinez, N.; Song, W.; Gannon, W.L.; Yates, T.L. Prevalence and geographic genetic variation of hantaviruses of New World harvest mice (Reithrodontomys): Identification of a divergent genotype from a Costa Rican Reithrodontomys mexicanus. Virology 1995, 207, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Vado-Solís, I.; Pérez-Osorio, C.; Lara-Lara, J.; Ruiz-Piña, H.A.; Cárdenas-Marrufo, M.; Milazzo, M.L.; Fulhorst, C.F.; Zavala-Velázquez, J. Evidencia serológica de infección por Hantavirus en población humana del estado de Yucatán, México. Rev. Biomed. 2003, 14, 221–225. [Google Scholar] [CrossRef]
- Rivas, Y.J.; Moros, Z.; Moron, D.; Uzcategui, M.G.; Duran, Z.; Pujol, F.H.; Liprandi, F.; Ludert, J.E. The seroprevalences of anti-hantavirus IgG antibodies among selected Venezuelan populations. Ann. Trop. Med. Parasitol. 2003, 97, 61–67. [Google Scholar] [CrossRef]
- Ramos, M.; Mola, C.L.d.; Salmon, G.; Guevara, C.; Pacheco, V.; Vasquez, A.; Kochel, T.; Albujar, C.; Rollin, P.; Comer, J.A.; Montgomery, J. Evidence of occupational exposure to Hantavirus and other vector-borne viral pathogens among mammalogist and other field workers in Peru. In 58th Annual Meeting of the American Society of Tropical Medicine and Hygiene, Washington, DC, USA, 18–22 November 2009. [Google Scholar]
- Chilean Ministry of Health. Boletín Epidemiológico de Hantavirus Situación al 14 octubre de 2010. Available online: http://epi.minsal.cl/epi/html/bolets/reportes/Hantavirus/Hantavirus.pdf (accessed on 12 November 2010).
- Bellomo, C.; Nudelman, J.; Kwaszka, R.; Vazquez, G.; Cantoni, G.; Weinzettel, B.; Larrieu, E.; Padula, P. Expansión geográfica del síndrome pulmonar por hantavirus en la Argentina: Informe del caso más austral. Medicina (B Aires) 2009, 69, 647–650. [Google Scholar] [PubMed]
- Figueiredo, L.T.; Moreli, M.L.; de-Sousa, R.L.; Borges, A.A.; de-Figueiredo, G.G.; Machado, A.M.; Bisordi, I.; Nagasse-Sugahara, T.K.; Suzuki, A.; Pereira, L.E.; de-Souza, R.P.; de-Souza, L.T.; Braconi, C.T.; Harsi, C.M.; de-Andrade-Zanotto, P.M. Hantavirus pulmonary syndrome, central plateau, southeastern, and southern Brazil. Emerg. Infect. Dis. 2009, 15, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Armien, B.; Pascale, J.M.; Bayard, V.; Munoz, C.; Mosca, I.; Guerrero, G.; Armien, A.; Quiroz, E.; Castillo, Z.; Zaldivar, Y.; Gracia, F.; Hjelle, B.; Koster, F. High seroprevalence of hantavirus infection on the Azuero peninsula of Panama. Am. J. Trop. Med. Hyg. 2004, 70, 682–687. [Google Scholar] [CrossRef]
- Sotomayor, V.; Olea, A.M.; Labrana, M. Diagnosis and treatment of cardiopulmonary hantavirus syndrome. Chile-2007. Rev. Chil. Infectol. 2009, 26, 68–84. [Google Scholar]
- Ferrer, J.F.; Galligan, D.; Esteban, E.; Rey, V.; Murua, A.; Gutierrez, S.; Gonzalez, L.; Thakuri, M.; Feldman, L.; Poiesz, B.; Jonsson, C. Hantavirus infection in people inhabiting a highly endemic region of the Gran Chaco territory, Paraguay: Association with Trypanosoma cruzi infection, epidemiological features and haematological characteristics. Ann. Trop. Med. Parasitol. 2003, 97, 269–280. [Google Scholar] [CrossRef]
- Ferrer, J.F.; Jonsson, C.B.; Esteban, E.; Galligan, D.; Basombrio, M.A.; Peralta-Ramos, M.; Bharadwaj, M.; Torrez-Martinez, N.; Callahan, J.; Segovia, A.; Hjelle, B. High prevalence of hantavirus infection in Indian communities of the Paraguayan and Argentinean Gran Chaco. Am. J. Trop. Med. Hyg. 1998, 59, 438–444. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Navarrete-Droguett, J.; Aldunate, R.; Yates, T.L.; Mertz, G.J.; Vial, P.A.; Ferres, M.; Marquet, P.A.; Palma, R.E. Peridomestic small mammals associated with confirmed cases of human hantavirus disease on southcentral Chile. Am. J. Trop. Med. Hyg. 2004, 70, 305–309. [Google Scholar] [CrossRef]
- Frey, M.T.; Vial, P.C.; Castillo, C.H.; Godoy, P.M.; Hjelle, B.; Ferres, M.G. Hantavirus prevalence in the IX Region of Chile. Emerg. Infect. Dis. 2003, 9, 827–832. [Google Scholar]
- Pini, N.; Levis, S.; Calderon, G.; Ramirez, J.; Bravo, D.; Lozano, E.; Ripoll, C.; St Jeor, S.; Ksiazek, T.G.; Barquez, R.M.; Enria, D. Hantavirus infection in humans and rodents, northwestern Argentina. Emerg. Infect. Dis. 2003, 9, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Limongi, J.E.; da Costa, F.C.; Pinto, R.M.; de Oliveira, R.C.; Bragagnolo, C.; Lemos, E.R.; de Paula, M.B.; Pajuaba Neto, A.A.; Ferreira, M.S. Cross-sectional Survey of Hantavirus Infection, Brazil. Emerg. Infect. Dis. 2009, 15, 1981–1983. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, L.M.; Lindsey, A.E.; Hjelle, B.; Dominguez, D.; Brown, J.; Goade, D.; Jonsson, C.B. Prevalence of antibodies to Sin Nombre virus in humans living in rural areas of southern New Mexico and western Texas. Virus Res. 2001, 74, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Douglass, R.J.; Semmens, W.J.; Matlock-Cooley, S.J.; Kuenzi, A.J. Deer mouse movements in peridomestic and sylvan settings in relation to Sin Nombre virus antibody prevalence. J. Wildl. Dis. 2006, 42, 813–818. [Google Scholar] [CrossRef]
- Kuenzi, A.J.; Douglass, R.J.; White, D.; Bond, C.W.; Mills, J.N. Antibody to Sin Nombre virus in rodents associated with peridomestic habitats in west central Montana. Am. J. Trop. Med. Hyg. 2001, 64, 137–146. [Google Scholar] [CrossRef]
- Botten, J.; Mirowsky, K.; Ye, C.Y.; Gottlieb, K.; Saavedra, M.; Ponce, L.; Hjelle, B. Shedding and intracage transmission of Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus) model. J. Virol. 2002, 76, 7587–7594. [Google Scholar] [CrossRef]
- Padula, P.; Figueroa, R.; Navarrete, M.; Pizarro, E.; Cadiz, R.; Bellomo, C.; Jofre, C.; Zaror, L.; Rodriguez, E.; Murua, R. Transmission study of Andes hantavirus infection in wild sigmodontine rodents. J. Virol. 2004, 78, 11972–11979. [Google Scholar] [CrossRef]
- Yates, T.L.; Mills, J.N.; Parmenter, C.A.; Ksiazek, T.G.; Parmenter, R.R.; Vande Castle, J.R.; Calisher, C.H.; Nichol, S.T.; Abbott, K.D.; Young, J.C.; Morrison, M.L.; Beaty, B.J.; Dunnum, J.L.; Baker, R.J.; Salazar-Bravo, J.; Peters, C.J. The ecology and evolutionary history of an emergent disease: Hantavirus pulmonary syndrome. Bioscience 2002, 52, 989–998. [Google Scholar] [CrossRef]
- Parmenter, R.R.; Brunt, J.W.; Moore, D.I.; Ernest, S. The Hantavirus epidemic in the Southwest: Rodent population dynamics and the implications for transmission of Hantavirus-associated Adult Respiratory Distress Syndrome HARDS) in the Four Corners Region. Report to the Federal Centers for Disease Control and Prevention; Publication No. 41; Sevilleta Long-Term Ecological Research Program (LTER): Atlanta, GA, USA, July 1993; pp. 1–45. [Google Scholar]
- Suzan, G.; Armien, A.; Mills, J.N.; Marce, E.; Ceballos, G.; Avila, M.; Salazar-Bravo, J.; Ruedas, L.; Armien, B.; Yates, T.L. Epidemiological considerations of rodent community composition in fragmented landscapes in Panama. J. Mammal. 2008, 89, 684–690. [Google Scholar] [CrossRef]
- Suzan, G.; Marce, E.; Giermakowski, J.T.; Armien, B.; Pascale, J.; Mills, J.; Ceballos, G.; Gomez, A.; Aguirre, A.A.; Salazar-Bravo, J.; Armien, A.; Parmenter, R.; Yates, T. The effect of habitat fragmentation and species diversity loss on hantavirus prevalence in panama. Ann. N. Y. Acad. Sci. 2008, 1149, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Goodin, D.G.; Koch, D.E.; Owen, R.D.; Chu, Y.K.; Hutchinson, J.M.S.; Jonsson, C.B. Land cover associated with hantavirus presence in Paraguay. Global Ecol. Biogeogr. 2006, 15, 519–527. [Google Scholar] [CrossRef]
- Hjelle, B.; Glass, G.E. Outbreak of hantavirus infection in the four corners region of the United States in the wake of the 1997–1998 El Nino-southern oscillation. J. Infect. Dis. 2000, 181, 1569–1573. [Google Scholar] [CrossRef]
- Klein, S.; Calisher, C. Emergence and Persistence of Hantaviruses. In Wildlife and Emerging Zoonotic Diseases: The Biology, Circumstances and Consequences of Cross-Species Transmission; Childs, J.E., Mackenzie, J.S., Richt, J.A., Eds.; Springer-Verlag Berlin Heidelberg: New York, NY, USA, 2007; pp. 217–252. [Google Scholar]
- Kallio, E.R.; Klingstrom, J.; Gustafsson, E.; Manni, T.; Vaheri, A.; Henttonen, H.; Vapalahti, O.; Lundkvist, A. Prolonged survival of Puumala hantavirus outside the host: Evidence for indirect transmission via the environment. J. Gen. Virol. 2006, 87, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Douron, E.; Moriniere, B.; Matheron, S.; Girard, P.M.; Gonzalez, J.P.; Hirsch, F.; McCormick, J.B. HFRS after a wild rodent bite in the Haute-Savoie—and risk of exposure to Hantaan-like virus in a Paris laboratory. Lancet 1984, 1, 676–677. [Google Scholar] [CrossRef]
- Schultze, D.; Lundkvist, A.; Blauenstein, U.; Heyman, P. Tula virus infection associated with fever and exanthema after a wild rodent bite. Eur. J. Clin. Microbiol. Infect. Dis. 2002, 21, 304–306. [Google Scholar] [CrossRef]
- St Jeor, S.C. Three-week incubation period for hantavirus infection. Pediatr. Infect. Dis. J. 2004, 23, 974–975. [Google Scholar]
- Fulhorst, C.F.; Milazzo, M.L.; Armstrong, L.R.; Childs, J.E.; Rollin, P.E.; Khabbaz, R.; Peters, C.J.; Ksiazek, T.G. Hantavirus and arenavirus antibodies in persons with occupational rodent exposure, North America. Emerg. Infect. Dis. 2007, 13, 532–538. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Wilson, L.; Collinge, S.K.; Harmon, H.; Ray, C.; Medina, R.A.; Hjelle, B. Sin nombre virus infection in field workers, colorado, USA. Emerg. Infect. Dis. 2010, 16, 308–310. [Google Scholar] [CrossRef]
- Kelt, D.A.; Van Vuren, D.H.; Hafner, M.S.; Danielson, B.J.; Kelly, M.J. Threat of hantavirus pulmonary syndrome to field biologists working with small mammals. Emerg. Infect. Dis. 2007, 13, 1285–1287. [Google Scholar] [CrossRef]
- Ferres, M.; Vial, P.; Marco, C.; Yanez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.J.; Mertz, G.J. Prospective evaluation of household contacts of persons with hantavirus cardiopulmonary syndrome in Chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef]
- Lazaro, M.E.; Cantoni, G.E.; Calanni, L.M.; Resa, A.J.; Herrero, E.R.; Iacono, M.A.; Enria, D.A.; Cappa, S.M.G. Clusters of hantavirus infection, southern Argentina. Emerg. Infect. Dis. 2007, 13, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Padula, P.J.; Edelstein, A.; Miguel, S.D.L.; Lopez, N.M.; Rossi, C.M.; Rabinovich, R.D. Hantavirus Pulmonary Syndrome outbreak in Argentina: Molecular evidence for person-to-person transmission of Andes virus. Virology 1998, 241, 323–330. [Google Scholar] [CrossRef]
- Wells, R.M.; Estani, S.S.; Yadon, Z.E.; Enria, D.; Padula, P.; Pini, N.; Della Valle, M.G.; Mills, J.N.; Peters, C.J. Seroprevalence of antibodies to hantavirus in health care workers and other residents of southern Argentina. Clin. Infect. Dis. 1998, 27, 895–896. [Google Scholar] [CrossRef] [PubMed]
- Wells, R.M.; Estani, S.S.; Yadon, Z.E.; Enria, D.; Padula, P.; Pini, N.; Mills, J.N.; Peters, C.J.; Segura, E.L.; Guthmann, N.; Arguelo, E.; Klein, F.; Levy, R.; Nagel, C.; Calfin, R.; deRosas, F.; Lazaro, M.; Rosales, H.; Sandoval, P. An unusual hantavirus outbreak in southern Argentina: Person-to-person transmission? Emerg. Infect. Dis. 1997, 3, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.P.; Bellomo, C.; San Juan, J.; Pinna, D.; Forlenza, R.; Elder, M.; Padula, P.J. Person-to-person transmission of Andes virus. Emerg. Infect. Dis. 2005, 11, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.; Marsac, D.; Stefas, E.; Ferrer, P.; Tischler, N.D.; Pino, K.; Ramdohr, P.; Vial, P.; Valenzuela, P.D.; Ferres, M.; Veas, F.; Lopez-Lastra, M. Andes Virus Antigens are shed in urine of patients with acute Hantavirus Cardiopulmonary Syndrome. J. Virol. 2009, 83, 5046–5055. [Google Scholar] [CrossRef]
- Pettersson, L.; Klingstrom, J.; Hardestam, J.; Lundkvist, A.; Ahlm, C.; Evander, M. Hantavirus RNA in saliva from patients with hemorrhagic Fever with renal syndrome. Emerg. Infect. Dis. 2008, 14, 406–411. [Google Scholar] [CrossRef]
- Kang, H.J.; Bennett, S.N.; Dizney, L.; Sumibcay, L.; Arai, S.; Ruedas, L.A.; Song, J.W.; Yanagihara, R. Host switch during evolution of a genetically distinct hantavirus in the American shrew mole (Neurotrichus gibbsii). Virology 2009, 388, 8–14. [Google Scholar] [CrossRef]
- Arai, S.; Bennett, S.N.; Sumibcay, L.; Cook, J.A.; Song, J.W.; Hope, A.; Parmenter, C.; Nerurkar, V.R.; Yates, T.L.; Yanagihara, R. Phylogenetically distinct hantaviruses in the masked shrew (Sorex cinereus) and dusky shrew (Sorex monticolus) in the United States. Am. J. Trop. Med. Hyg. 2008, 78, 348–351. [Google Scholar] [CrossRef]
- Plyusnin, A.; Vapalahti, O.; Vaheri, A. Hantaviruses: Genome structure, expression and evolution. J. Gen. Virol. 1996, 77, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.K.; Owen, R.D.; Sanchez-Hernandez, C.; Romero-Almaraz, M.D.; Jonsson, C.B. Genetic characterization and phylogeny of a hantavirus from Western Mexico. Virus Res. 2007, 131, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Milazzo, M.L.; Cajimat, M.N.B.; Hanson, J.D.; Bradley, R.D.; Quintana, M.; Sherman, C.; Velasquez, R.T.; Fulhorst, C.F. Catacamas virus, a hantaviral species naturally associated with Oryzomys couesi (Coues’oryzomys) in Honduras. Am. J. Trop. Med. Hyg. 2006, 75, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Fulhorst, C.F.; Cajimat, M.N.B.; Utrera, A.; Milazzo, M.L.; Duno, G.M. Maporal virus, a hantavirus associated with the fulvous pygmy rice rat (Oligoryzomys fulvescens) in western Venezuela. Virus Res. 2004, 104, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, J.A.; Torrez-Martinez, N.; Neill, S.U.; Moore, G.M.; Hicks, B.N.; Pichuantes, S.; Nguyen, A.; Bharadwaj, M.; Hjelle, B. Cocirculation of multiple hantaviruses in Texas, with characterization of the small (S) genome of a previously undescribed virus of cotton rats (Sigmodon hispidus). Am. J. Trop. Med. Hyg. 1996, 55, 672–679. [Google Scholar] [CrossRef]
- Rollin, P.E.; Ksiazek, T.G.; Elliott, L.H.; Ravkov, E.V.; Martin, M.L.; Morzunov, S.; Livingstone, W.; Monroe, M.; Glass, G.; Ruo, S. Isolation of Black Creek Canal virus, a new hantavirus from Sigmodon hispidus in Florida. J. Med. Virol. 1995, 46, 35–39. [Google Scholar] [CrossRef]
- Johnson, A.M.; Bowen, M.D.; Ksiazek, T.G.; Williams, R.J.; Bryan, R.T.; Mills, J.N.; Peters, C.J.; Nichol, S.T. Laguna Negra virus associated with HPS in western Paraguay and Bolivia. Virology 1997, 238, 115–127. [Google Scholar] [CrossRef]
- Carroll, D.S.; Mills, J.N.; Montgomery, J.M.; Bausch, D.G.; Blair, I.J.; Burans, J.P.; Felices, V.; Gianella, A.; Iihoshi, N.; Nichol, S.T.; Olson, J.G.; Rogers, D.S.; Salazar, M.; Ksiazek, T.G. Hantavirus pulmonary syndrome in central Bolivia: Relationships between reservoir hosts, habitats, and viral genotypes. Am. J. Trop. Med. Hyg. 2005, 72, 42–46. [Google Scholar] [CrossRef]
- Maes, P.; Klempa, B.; Clement, J.; Matthijnssens, J.; Gajdusek, D.C.; Kruger, D.H.; Van Ranst, M. A proposal for new criteria for the classification of hantaviruses, based on S and M segment protein sequences. Infect. Genet. Evol. 2009, 9, 813–820. [Google Scholar] [CrossRef]
- Plyusnin, A. Genetics of hantaviruses: Implications to taxonomy. Arch. Virol. 2002, 147, 665–682. [Google Scholar] [CrossRef]
- Lloyd-Smith, J.O.; George, D.; Pepin, K.M.; Pitzer, V.E.; Pulliam, J.R.; Dobson, A.P.; Hudson, P.J.; Grenfell, B.T. Epidemic dynamics at the human-animal interface. Science 2009, 326, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Hjelle, B.; Yates, T. Modeling hantavirus maintenance and transmission in rodent communities. Curr. Top. Microbiol. Immunol. 2001, 256, 77–90. [Google Scholar]
- Schmidt-Chanasit, J.; Essbauer, S.; Petraityte, R.; Yoshimatsu, K.; Tackmann, K.; Conraths, F.J.; Sasnauskas, K.; Arikawa, J.; Thomas, A.; Pfeffer, M.; Scharninghausen, J.J.; Splettstoesser, W.; Wenk, M.; Heckel, G.; Ulrich, R.G. Extensive host sharing of central European Tula virus. J. Virol. 2010, 84, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Delfraro, A.; Tome, L.; D’Elia, G.; Clara, M.; Achaval, F.; Russi, J.C.; Rodonz, J.R. Juquitiba-like hantavirus from 2 nonrelated rodent species, Uruguay. Emerg. Infect. Dis. 2008, 14, 1447–1451. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, M.; Klempa, B.; Auste, B.; Bemmann, M.; Schmidt-Chanasit, J.; Buchner, T.; Groschup, M.H.; Meier, M.; Balkema-Buschmann, A.; Zoller, H.; Kruger, D.H.; Ulrich, R.G. Dobrava-belgrade virus spillover infections, Germany. Emerg. Infect. Dis. 2009, 15, 2017–2020. [Google Scholar] [CrossRef]
- Weidmann, M.; Schmidt, P.; Vackova, A.; Krivanec, K.; Munclinger, P.; Hufert, F.T. Identification of genetic evidence for Dobrava virus spillover in rodents by nested reverse transcription (RT)-PCR and TaqMan RT-PCR. J. Clin. Microbiol. 2005, 43, 808–812. [Google Scholar] [CrossRef]
- Vapalahti, O.; Lundkvist, A.; Fedorov, V.; Conroy, C.J.; Hirvonen, S.; Plyusnina, A.; Nemirov, K.; Fredga, K.; Cook, J.A.; Niemimaa, J.; Kaikusalo, A.; Henttonen, H.; Vaheri, A.; Plyusnin, A. Isolation and characterization of a hantavirus from Lemmus sibiricus: Evidence for host switch during hantavirus evolution. J. Virol. 1999, 73, 5586–5592. [Google Scholar] [CrossRef]
- Medina, R.A.; Torres-Pérez, F.; Galeno, H.; Navarrete, M.; Vial, P.A.; Palma, R.E.; Ferres, M.; Cook, J.A.; Hjelle, B. Ecology, genetic diversity, and phylogeographic structure of Andes virus in humans and rodents in Chile. J. Virol. 2009, 83, 2446–2459. [Google Scholar] [CrossRef]
- Toro, J.; Vega, J.D.; Khan, A.S.; Mills, J.N.; Padula, P.; Terry, W.; Yadon, Z.; Valderrama, R.; Ellis, B.A.; Pavletic, C.; Cerda, R.; Zaki, S.; Wun-Ju, S.; Meyer, R.; Tapia, M.; Mansilla, C.; Baro, M.; Vergara, J.A.; Concha, M.; Calderon, G.; Enria, D.; Peters, C.J.; Ksiazek, T.G. An outbreak of hantavirus pulmonary syndrome, Chile, 1997. Emerg. Infect. Dis. 1998, 4, 687–694. [Google Scholar] [CrossRef]
- McIntyre, N.E.; Chu, Y.K.; Owen, R.D.; Abuzeineh, A.; De La Sancha, N.; Dick, C.W.; Holsomback, T.; Nisbett, R.A.; Jonsson, C. A longitudinal study of Bayou virus, hosts, and habitat. Am. J. Trop. Med. Hyg. 2005, 73, 1043–1049. [Google Scholar] [CrossRef]
- Holsomback, T.S.; McIntyre, N.E.; Nisbett, R.A.; Strauss, R.E.; Chu, Y.-K.; Abuzeineh, A.A.; Sancha, N.d.l.; Dick, C.W.; Jonsson, C.B.; Morris, B.E.L. Bayou virus detected in non-oryzomyine rodent hosts: An assessment of habitat composition, reservoir community structure, and marsh rice rat social dynamics. J. Vector Ecol. 2009, 34, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Torrez-Martinez, N.; Bharadwaj, M.; Goade, D.; Delury, J.; Moran, P.; Hicks, B.; Nix, B.; Davis, J.L.; Hjelle, B. Bayou virus-associated hantavirus pulmonary syndrome in eastern Texas: Identification of the rice rat, Oryzomys palustris, as reservoir host. Emerg. Infect. Dis. 1998, 4, 105–111. [Google Scholar] [CrossRef]
- Allen, L.J.; Wesley, C.L.; Owen, R.D.; Goodin, D.G.; Koch, D.; Jonsson, C.B.; Chu, Y.K.; Shawn Hutchinson, J.M.; Paige, R.L. A habitat-based model for the spread of hantavirus between reservoir and spillover species. J. Theor. Biol. 2009, 260, 510–522. [Google Scholar] [CrossRef] [PubMed]
- DeFilippis, V.R.; Villarreal, L.P. An introduction to the evolutionary ecology of viruses. In Viral Ecology; Hurst, C.J., Ed.; Academic Press: San Diego, USA, 2000; pp. 125–208. [Google Scholar]
- Schmaljohn, C.; Hjelle, B. Hantaviruses: A global disease problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Bayard, V.; Kitsutani, P.T.; Barria, E.O.; Ruedas, L.A.; Tinnin, D.S.; Munoz, C.; de Mosca, I.B.; Guerrero, G.; Kant, R.; Garcia, A.; Caceres, L.; Gracia, F.G.; Quiroz, E.; Castillo, Z.C.; Armien, B.; Libel, M.; Mills, J.N.; Khan, A.S.; Nichol, S.T.; Rollin, P.E.; Ksiazek, T.G.; Peters, C.J. Outbreak of hantavirus pulmonary syndrome, Los Santos, Panama, 1999–2000. Emerg. Infect. Dis. 2004, 10, 1635–1642. [Google Scholar] [CrossRef]
- Bayard, V.; Ortega, E.; Garcia, A.; Caceres, L.; Castillo, Z.; Quiroz, E.; Armien, B.; Gracia, F.; Serrano, J.; Guerrero, G.; Kant, R.; Pinifla, E.; Bravo, L.; Munoz, C.; de Mosca, I.B.; Rodriguez, A.; Campos, C.; Diaz, M.A.; Munoz, B.; Crespo, F.; Villalaz, I.; Rios, P.; Morales, E.; Sitton, J.M.T.; Reneau-Vernon, L.; Libel, M.; Castellanos, L.; Ruedas, L.; Tinnin, D.; Yates, T. Hantavirus pulmonary syndrome—Panama, 1999–2000. MMWR 2000, 49, 205–207, Reprinted J. Am. Med. Assoc. 2000, 283, 2232–2233. [Google Scholar]
- Williams, R.J.; Bryan, R.T.; Mills, J.N.; Palma, R.E.; Vera, I.; De Velasquez, F.; Baez, E.; Schmidt, W.E.; Figueroa, R.E.; Peters, C.J.; Zaki, S.R.; Khan, A.S.; Ksiazek, T.G. An outbreak of hantavirus pulmonary syndrome in western Paraguay. Am. J. Trop. Med. Hyg. 1997, 57, 274–282. [Google Scholar] [CrossRef]
- Mendes, W.S.; da Silva, A.A.M.; Aragao, L.F.C.; Aragao, N.J.L.; Raposo, M.D.; Elkhoury, M.R.; Suzuky, A.; Ferreira, I.B.; de Sousa, L.T.; Pannuti, C.S. Hantavirus infection in Anajatuba, Maranhao, Brazil. Emerg. Infect. Dis. 2004, 10, 1496–1498. [Google Scholar] [CrossRef]
- Glass, G.E.; Shields, T.; Cai, B.; Yates, T.L.; Parmenter, R. Persistently highest risk areas for hantavirus pulmonary syndrome: Potential sites for refugia. Ecol. Appl. 2007, 17, 129–139. [Google Scholar] [CrossRef]
- Glass, G.E.; Yates, T.L.; Fine, J.B.; Shields, T.M.; Kendall, J.B.; Hope, A.G.; Parmenter, C.A.; Peters, C.J.; Ksiazek, T.G.; Li, C.S.; Patz, J.A.; Mills, J.N. Satellite imagery characterizes local animal reservoir populations of Sin Nombre virus in the southwestern United States. Proc. Natl. Acad. Sci. U. S. A. 2002, 99, 16817–16822. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: An emerging (or re-emerging) discipline. Trends Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Carbajo, A.E.; Pardinas, U.F.J. Spatial distribution model of a Hantavirus reservoir, the long-tailed colilargo. J. Mammal. 2007, 88, 1555–1568. [Google Scholar] [CrossRef]
- Mills, J.N.; Amman, B.R.; Glass, G.E. Ecology of Hantaviruses and Their Hosts in North America. Vector Borne Zoonotic Dis. 2009, 10, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N. Regulation of rodent-borne viruses in the natural host: Implications for human disease. Arch. Virol. Suppl. 2005, 45–57. [Google Scholar]
- Adler, F.R.; Pearce-Duvet, J.M.C.; Dearing, M.D. How host population dynamics translate into time-lagged prevalence: An investigation of Sin Nombre virus in deer mice. Bull. Math. Biol. 2008, 70, 236–252. [Google Scholar] [CrossRef]
- Clay, C.A.; Lehmer, E.M.; Previtali, A.; St. Jeor, S.; Dearing, M.D. Contact heterogeneity in deer mice: Implications for Sin Nombre virus transmission. Proc. R. Soc. B. 2009, 276, 1305–1312. [Google Scholar] [CrossRef]
- Dizney, L.J.; Ruedas, L.A. Increased host species diversity and decreased prevalence of Sin Nombre virus. Emerg. Infect. Dis. 2009, 15, 1012–1018. [Google Scholar] [CrossRef]
- Suzan, G.; Marce, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armien, B.; Pascale, J.M.; Yates, T.L. Experimental Evidence for Reduced Rodent Diversity Causing Increased Hantavirus Prevalence. PLoS ONE 2009, 4, e5461. [Google Scholar] [CrossRef]
- Langlois, J.P.; Fahrig, L.; Merriam, G.; Artsob, H. Landscape structure influences continental distribution of hantavirus in deer mice. Landscape Ecol. 2001, 16, 255–266. [Google Scholar] [CrossRef]
- Goodin, D.G.; Paige, R.; Owen, R.D.; Ghimire, K.; Koch, D.E.; Chu, Y.-K.; Jonsson, C.B. Microhabitat characteristics of Akodon montensis, a reservoir for hantavirus, and hantaviral seroprevalence in an Atlantic forest site in eastern Paraguay. J. Vector Ecol. 2009, 34, 104–113. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Palma, R.E.; Hjelle, B.; Ferres, M.; Cook, J.A. Andes virus infections in the rodent reservoir and in humans vary across contrasting landscapes in Chile. Infect. Genet. Evol. 2010, 10, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Klempa, B. Hantaviruses and climate change. Clin Microbiol Infect 2009, 15, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.; Vercauteren, J.; Verstraeten, W.W.; Ducoffre, G.; Barrios, J.M.; Vandamme, A.M.; Maes, P.; Van Ranst, M. Relating increasing hantavirus incidences to the changing climate: The mast connection. Int. J. Health. Geogr. 2009, 8, 1. [Google Scholar] [CrossRef]
- Engelthaler, D.M.; Mosley, D.G.; Cheek, J.E.; Levy, C.E.; Komatsu, K.K.; Ettestad, P.; Davis, T.; Tanda, D.T.; Miller, L.; Frampton, J.W.; Porter, R.; Bryan, R.T. Climatic and environmental patterns associated with hantavirus pulmonary syndrome, Four Corners region, United States. Emerg. Infect. Dis. 1999, 5, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Jaksic, F.M.; Lima, M. Myths and facts on ratadas: Bamboo blooms, rainfall peaks and rodent outbreaks in South America. Austral Ecol. 2003, 28, 237–251. [Google Scholar] [CrossRef]
- Buceta, J.; Escudero, C.; de la Rubia, F.J.; Lindenberg, K. Outbreaks of Hantavirus induced by seasonality. Phys. Rev. E 2004, 69, 8. [Google Scholar] [CrossRef]
- Weiss, R.A.; McMichael, A.J. Social and environmental risk factors in the emergence of infectious diseases. Nat. Med. 2004, 10, S70–S76. [Google Scholar] [CrossRef]
- Sauvage, F.; Langlais, M.; Pontier, D. Predicting the emergence of human hantavirus disease using a combination of viral dynamics and rodent demographic patterns. Epidemiol. Infect. 2007, 135, 46–56. [Google Scholar] [CrossRef]
Table 1.
Major human-pathogenic hantaviruses.
| Virus strain | Acronym | Geographic Distribution | Host | Pathology | Case-fatality ratio (%) | |
| New World | Sin Nombre | SNV | North America | Peromyscus maniculatus | HCPS | –35 |
| Choclo | CHOV | Panamá | Oligoryzomys fulvescens | HCPS | ~21 | |
| Andes | ANDV | Argentina, Chile | Oligoryzomys longicaudatus | HCPS | 35 | |
| Laguna Negra | LANV | Argentina, Bolivia, Paraguay, | Calomys laucha, C. callosus | HCPS | 5–15 | |
| Old World | Hantaan | HTNV | China, Korea, Russia | Apodemus agrarius | HFRS | 5–0 |
| Seoul | SEOV | Worldwide | Rattus rattus, R. norvegicus | HFRS | 1–2 | |
| Puumala | PUUV | Scandinavia, western Europe, Russia | Myodes glareolus | HFRS/NE | 0.1–0.4 | |
| Dobrava-Belgrade | DOBV | Balkans | Apodemus flavicollis | HFRS | 5–10 |
HCPS: Hantavirus cardiopulmonary syndrome; HFRS: Hemorrhagic fever with renal syndrome; NE: Nephropathia epidemica
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Hjelle, B.; Torres-Pérez, F. Hantaviruses in the Americas and Their Role as Emerging Pathogens. Viruses 2010, 2, 2559-2586. https://doi.org/10.3390/v2122559
AMA Style
Hjelle B, Torres-Pérez F. Hantaviruses in the Americas and Their Role as Emerging Pathogens. Viruses. 2010; 2(12):2559-2586. https://doi.org/10.3390/v2122559
Chicago/Turabian StyleHjelle, Brian, and Fernando Torres-Pérez. 2010. "Hantaviruses in the Americas and Their Role as Emerging Pathogens" Viruses 2, no. 12: 2559-2586. https://doi.org/10.3390/v2122559