Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients

Abstract
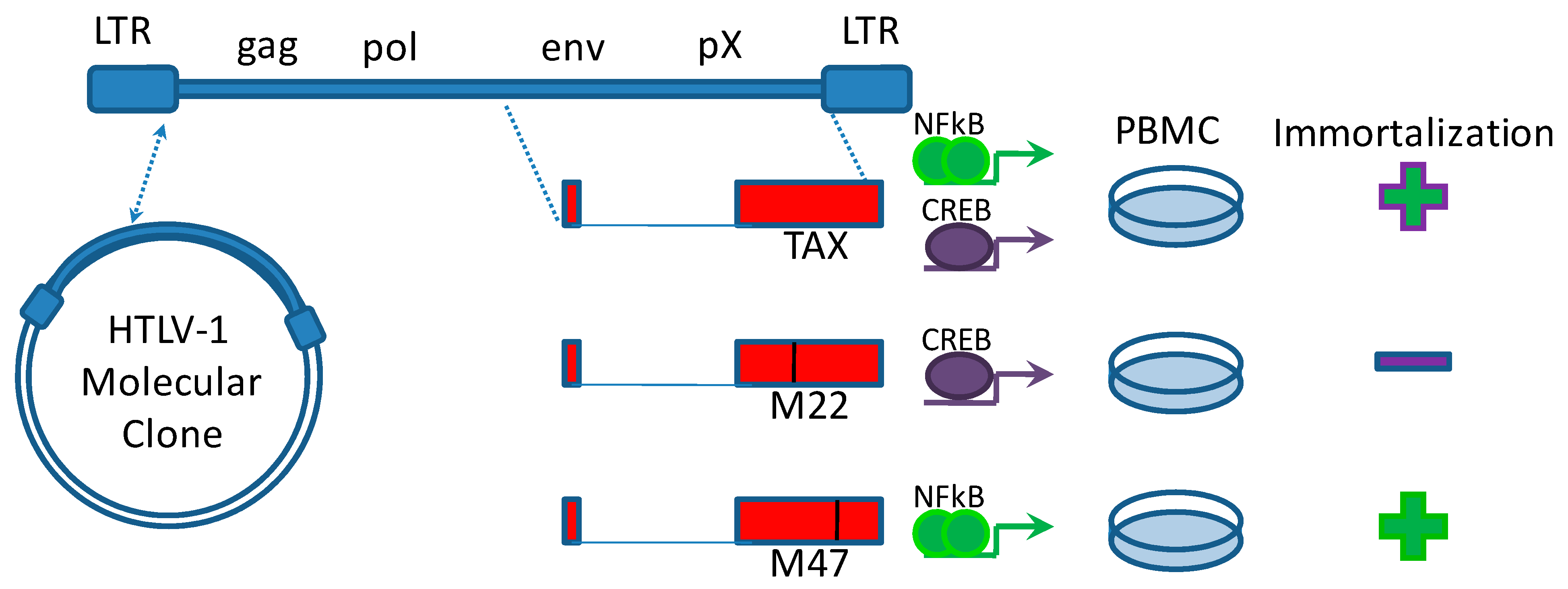
:1. Introduction
2. HTLV-1 Oncogenesis Depends on NFκB
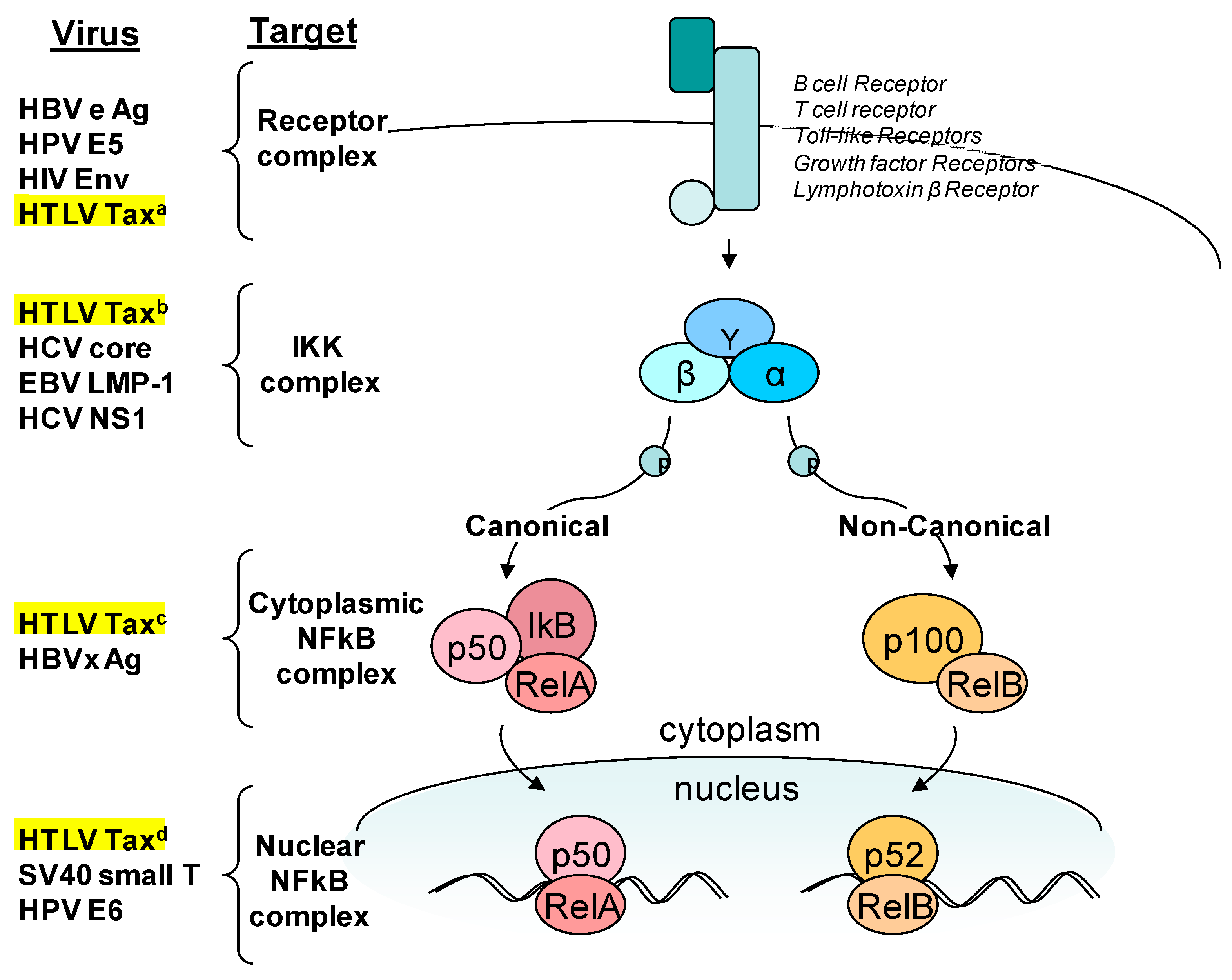
3. The NFκB Signaling Pathway
4. Tax Targets the NFκB Pathway
5. NFκB and Apoptosis
6. NFκB and Inflammation
7. NFκB and T-Cells
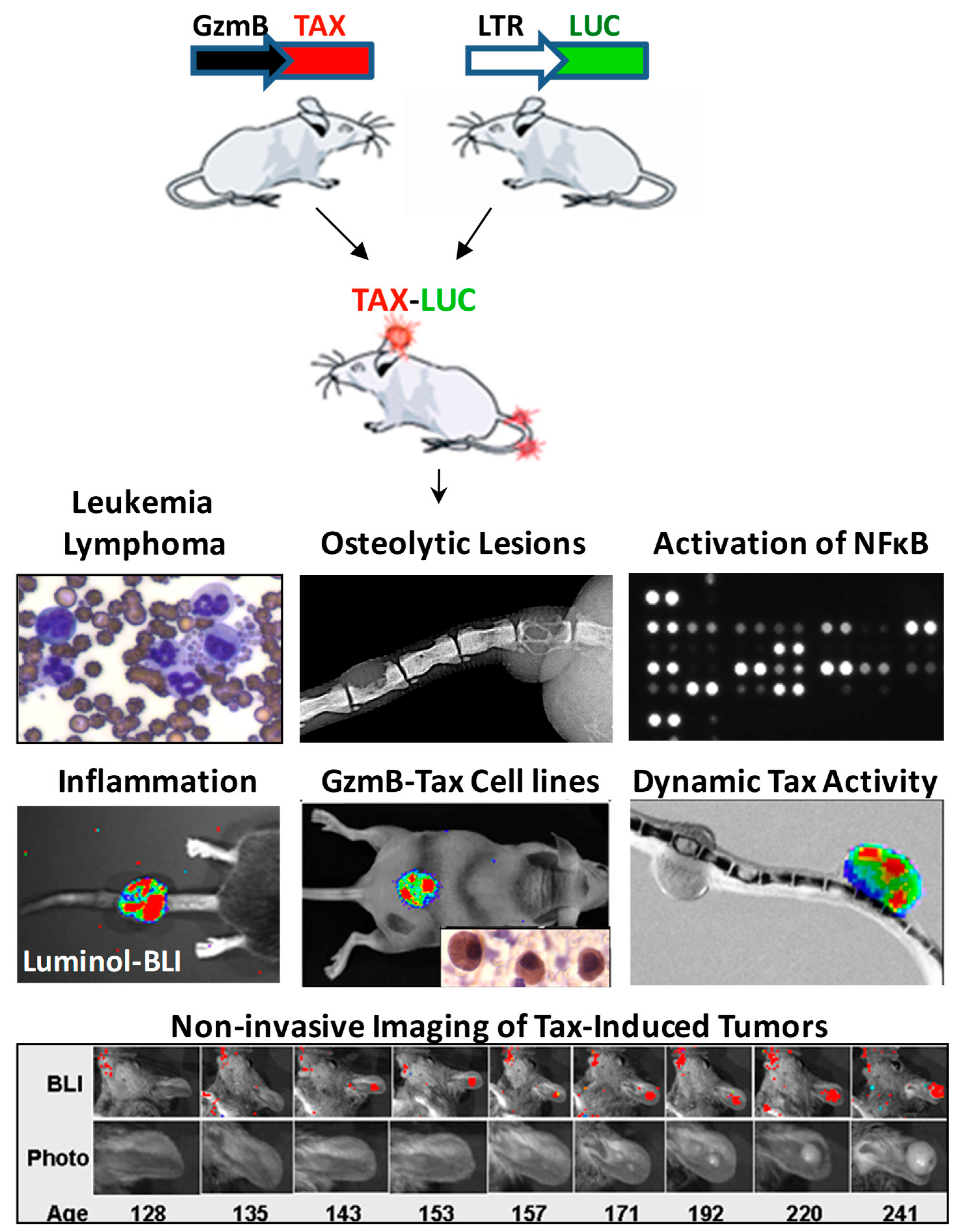
8. Targeting NFκB in vivo
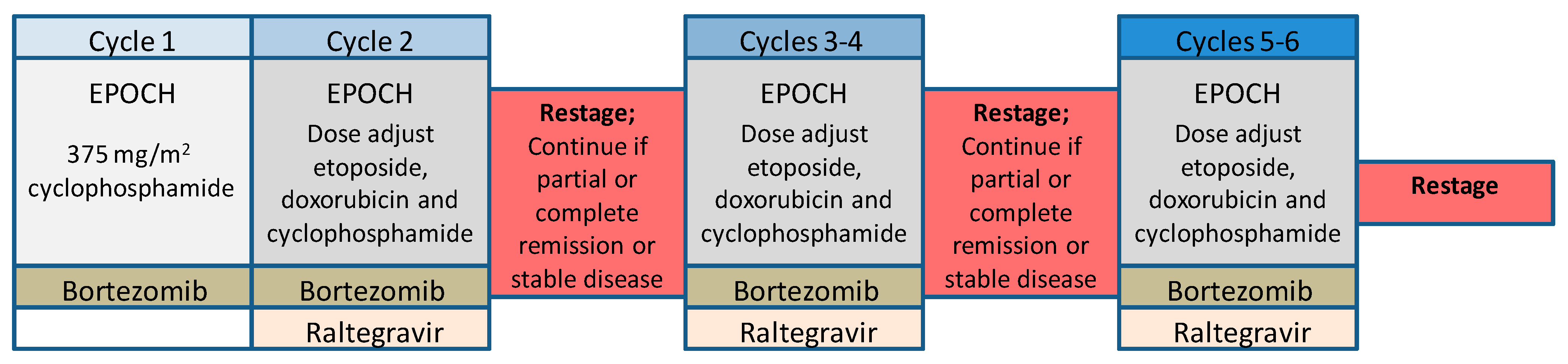
9. Targeting NFκB in ATLL Patients
10. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Peloponese, J.; Yeung, M.; Jeang, K. Modulation of nuclear factor-kB by human T-cell leukemia virus type 1 tax protein: Implications for oncogenesis and inflammation. Immunol. Res. 2006, 34, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Horie, R. NF-kB in pathogenesis and treatment of adult T-cell leukemia/lymphoma. Int. Rev. Immunol. 2007, 26, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Nerenberg, M.I. An HTLV-I Transgenic Mouse Model: Role of the tax gene in pathogenesis in multiple organ systems. Curr. Top. Microbiol. Immunol. 1990, 160, 121–128. [Google Scholar] [PubMed]
- Iwakura, Y.; Tosu, M.; Yoshida, E.; Takiguchi, M.; Sato, K.; Kitajima, I.; Nishioka, K.; Yamamoto, K.; Takeda, T.; Hatanaka, M.; et al. Induction of inflammatory arthropathy resembling rheumatoid arthritis in mice transgenic for HTLV-I. Science 1991, 253, 1026–1028. [Google Scholar] [CrossRef] [PubMed]
- Iwakura, Y.; Saijo, S.; Kioka, Y.; Nakayama-Yamada, J.; Itagaki, K.; Tosu, M.; Asano, M.; Kanai, Y.; Kakimoto, K. Autoimmunity induction by human T-cell leukemia virus type 1 in transgenic mice that develop chronic inflammatory arthropathy resembling rheumatoid arthritis in humans. J. Immunol. 1995, 155, 1588–1598. [Google Scholar] [CrossRef]
- Yamamoto, H.; Sekiguchi, T.; Itagaki, K.; Saijo, S.; Iwakura, Y. Inflammatory polyarthritis in mice transgenic for human T-cell leukemia virus type I. Arthritis Rheum. 1993, 36, 1612–1620. [Google Scholar] [CrossRef]
- Fujisawa, K.; Asahara, H.; Okamoto, K.; Aono, H.; Hasunuma, T.; Kobata, T.; Iwakura, Y.; Yonehara, S.; Sumida, T.; Nishioka, K. Therapeutic effect of the anti-Fas antibody on arthritis in HTLV-1 tax transgenic mice. J. Clin. Invest. 1996, 98, 271–278. [Google Scholar] [CrossRef]
- Grossman, W.; Kimata, J.; Wong, F.; Zutter, M.; Ley, T.; Ratner, L. Development of leukemia in mice transgenic for the tax gene of human T-cell leukemia virus type I. Proc. Natl. Acad. Sci. U. S. A. 1995, 92, 1057–1061. [Google Scholar] [CrossRef]
- Hasegawa, H.; Sawa, H.; Lewis, M.; Orba, Y.; Sheehy, N.; Yamamoto, Y.; Ichinohe, T.; Tsunetsugu-Yokota, Y.; Katano, H.; Takahashi, H.; et al. Thymus-derived leukemia-lymphoma in mice transgenic for the Tax gene of human T-lymphotropic virus type I. Nat. Med. 2006, 12, 466–472. [Google Scholar] [CrossRef]
- Watters, K.; Dean, J.; Hasegawa, H.; Sawa, H.; Hall, W.; Sheehy, N. Cytokine and growth factor expression by HTLV-1 Lck-tax transgenic cells in SCID mice. AIDS Res. Hum. Retroviruses 2010, 26, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Kimata, J.; Wong, F.; Wang, J.; Ratner, L. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology 1994, 204, 656–664. [Google Scholar] [CrossRef]
- Robek, M.; Ratner, L. Immortalization of CD4(+) and CD8(+) T lymphocytes by human T-cell leukemia virus type 1 Tax mutants expressed in a functional molecular clone. J. Virol. 1999, 73, 4856–4865. [Google Scholar] [CrossRef] [PubMed]
- Shih, V.; Tsui, R.; Caldwell, A.; Hoffmann, A. A single NFκB system for both canonical and non-canonical signaling. Cell Res. 2011, 21, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Saggioro, D.; Silic-Benussi, M.; Biasiotto, R.; D'Agostino, D.; Ciminale, V. Control of cell death pathways by HTLV-1 proteins. Front. Biosci. 2009, 14, 3338–3351. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Ballard, D. Persistent activation of NFκB by the tax transforming protein of HTLV-1; hijacking cellular IkappaB kinases. Oncogene 1999, 18, 6948–6958. [Google Scholar] [CrossRef]
- Jeong, S.; Pise-Masison, C.; Radonovich, M.; Park, H.; Brady, J. Activated AKT regulates NF-kappaB activation, p53 inhibition and cell survival in HTLV-1-transformed cells. Oncogene 2005, 24, 6719–6728. [Google Scholar] [CrossRef]
- Tanaka, H.; Fujita, N.; Tsuruo, T. 3-Phosphoinositide-dependent protein kinase-1-mediated IkappaB kinase beta (IkkB) phosphorylation activates NF-kappaB signaling. J. Biol. Chem. 2005, 280, 40965–40973. [Google Scholar] [CrossRef]
- O'Mahony, A.; Montano, M.; Van Beneden, K.; Chen, L.; Greene, W. Human T-cell lymphotropic virus type 1 tax induction of biologically active NF-kappaB requires IkappaB kinase-1-mediated phosphorylation of RelA/p65. J. Biol. Chem. 2004, 279, 18137–18145. [Google Scholar] [CrossRef]
- Xiao, G.; Cvijic, M.; Fong, A.; Harhaj, E.; Uhlik, M.; Waterfield, M.; Sun, S. Retroviral oncoprotein Tax induces processing of NF-kappaB2/p100 in T cells: Evidence for the involvement of IKKalpha. EMBO J. 2001, 20, 6805–6815. [Google Scholar] [CrossRef]
- Bernal-Mizrachi, L.; Lovly, C.; Ratner, L. The role of NF-{kappa}B-1 and NF-{kappa}B-2-mediated resistance to apoptosis in lymphomas. Proc. Natl. Acad. Sci. U S A. 2006, 103, 9220–9225. [Google Scholar] [CrossRef]
- Shoji, T.; Higuchi, M.; Kondo, R.; Takahashi, M.; Oie, M.; Tanaka, Y.; Aoyagi, Y.; Fujii, M. Identification of a novel motif responsible for the distinctive transforming activity of human T-cell leukemia virus (HTLV) type 1 Tax1 protein from HTLV-2 Tax2. Retrovirology 2009, 6, 83. [Google Scholar] [CrossRef] [PubMed]
- Portis, T.; Grossman, W.; Harding, J.; Hess, J.; Ratner, L. Analysis of p53 inactivation in a human T-cell leukemia virus type 1 Tax transgenic mouse model. J. Virol. 2001, 75, 2185–2193. [Google Scholar] [CrossRef] [PubMed]
- Pise-Masison, C.; Mahieux, R.; Jiang, H.; Ashcroft, M.; Radonovich, M.; Duvall, J.; Guillerm, C.; Brady, J. Inactivation of p53 by human T-cell lymphotropic virus type 1 Tax requires activation of the NF-kappaB pathway and is dependent on p53 phosphorylation. Mol. Cell. Biol. 2000, 10, 3377–3386. [Google Scholar] [CrossRef] [PubMed]
- Oh, U.; McCormick, M.; Datta, D.; Turner, R.; Bobb, K.; Monie, D.; Sliskovic, D.; Tanaka, Y.; Zhang, J.; Meshulam, J.; et al. Inhibition of immune activation by a novel nuclear factor-kappa B inhibitor in HTLV-I-associated neurologic disease. Blood 2011, 117, 3363–3369. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Sica, A.; Balkwill, F. Cancer-related inflammation. Nature 2008, 454, 436–444. [Google Scholar] [CrossRef]
- Schäfer, M.; Werner, S. Cancer as an overhealing wound: An old hypothesis revisited. Nat. Rev. Mol. Cell. Biol. 2008, 9, 628–638. [Google Scholar] [CrossRef]
- Gao, L.; Deng, H.; Zhao, H.; Hirbe, A.; Harding, J.; Ratner, L.; Weilbaecher, K. HTLV-1 Tax transgenic mice develop spontaneous osteolytic bone metastases prevented by osteoclast inhibition. Blood 2005, 106, 4294–4302. [Google Scholar] [CrossRef]
- Portis, T.; Harding, J.; Ratner, L. The contribution of NF-kappa B activity to spontaneous proliferation and resistance to apoptosis in human T-cell leukemia virus type 1 Tax-induced tumors. Blood 2001, 98, 1200–1208. [Google Scholar] [CrossRef]
- Grossman, W.; Ratner, L. Cytokine expression and tumorigenicity of large granular lymphocytic leukemia cells from mice transgenic for the tax gene of human T-cell leukemia virus type I. Blood 1997, 90, 783–794. [Google Scholar] [CrossRef]
- Gross, S.; Gammon, S.; Moss, B.; Rauch, D.; Harding, J.; Heinecke, J.; Ratner, L.; Piwnica-Worms, D. Bioluminescence imaging of myeloperoxidase activity in vivo. Nat. Med. 2009, 15, 455–461. [Google Scholar] [CrossRef]
- Rauch, D.; Gross, S.; Harding, J.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Ratner, L. Imaging spontaneous tumorigenesis: inflammation precedes development of peripheral NK tumors. Blood 2009, 113, 1493–1500. [Google Scholar] [CrossRef] [PubMed]
- Mitra-Kaushik, S.; Harding, J.; Hess, J.; Schreiber, R.; Ratner, L. Enhanced tumorigenesis in HTLV-1 tax-transgenic mice deficient in interferon-gamma. Blood 2004, 104, 3305–3311. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, Y.; Osame, M.; Kubota, R.; Tara, M.; Yoshida, M. Human T-cell leukemia virus type-1 (HTLV-1) Tax is expressed at the same level in infected cells of HTLV-1-associated myelopathy or tropical spastic paraparesis patients as in asymptomatic carriers but at a lower level in adult T-cell leukemia cells. Blood 1995, 85, 1865–1870. [Google Scholar] [CrossRef] [PubMed]
- Bangham, C.; Osame, M. Cellular immune response to HTLV-1. Oncogene 2005, 24, 6035–6046. [Google Scholar] [CrossRef] [PubMed]
- Rauch, D.; Gross, S.; Harding, J.; Bokhari, S.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Ratner, L. T-cell activation promotes tumorigenesis in inflammation-associated cancer. Retrovirology 2009, 6, 116. [Google Scholar] [CrossRef] [PubMed]
- Mitra-Kaushik, S.; Harding, J.; Hess, J.; Ratner, L. Effects of the proteasome inhibitor PS-341 on tumor growth in HTLV-1 Tax transgenic mice and Tax tumor transplants. Blood 2004, 104, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Yamada, Y.; Ikeda, S.; Yamasaki, Y.; Tsukasaki, K.; Tanaka, Y.; Tomonaga, M.; Yamamoto, N.; Fujii, M. Bay 11–7082 inhibits transcription factor NF-kappaB and induces apoptosis of HTLV-I-infected T-cell lines and primary adult T-cell leukemia cells. Blood 2002, 100, 1828–1834. [Google Scholar] [CrossRef]
- Horie, R. NF-kappaB in pathogenesis and treatment of adult T-cell leukemia/lymphoma. Int. Rev. Immunol. 2007, 26, 269–281. [Google Scholar] [CrossRef]
- Ikezoe, T.; Yang, Y.; Bandobashi, K.; Saito, T.; Takemoto, S.; Machida, H.; Togitani, K.; Koeffler, H.; Taguchi, H. Oridonin, a diterpenoid purified from Rabdosia rubescens, inhibits the proliferation of cells from lymphoid malignancies in association with blockade of the NF-kappa B signal pathways. Mol. Cancer Ther. 2005, 4, 578–586. [Google Scholar] [CrossRef]
- Okudaira, T.; Tomita, M.; Uchihara, J.; Matsuda, T.; Ishikawa, C.; Kawakami, H.; Masuda, M.; Tanaka, Y.; Ohshiro, K.; Takasu, N.; et al. NIK-333 inhibits growth of human T-cell leukemia virus type I-infected T-cell lines and adult T-cell leukemia cells in association with blockade of nuclear factor-kappaB signal pathway. Mol. Cancer Ther. 2006, 5, 704–712. [Google Scholar] [CrossRef]
- Tomita, M.; Kawakami, H.; Uchihara, J.; Okudaira, T.; Masuda, M.; Takasu, N.; Matsuda, T.; Ohta, T.; Tanaka, Y.; Ohshiro, K.; et al. Curcumin (diferuloylmethane) inhibits constitutive active NF-kappaB, leading to suppression of cell growth of human T-cell leukemia virus type I-infected T-cell lines and primary adult T-cell leukemia cells. Int. J. Cancer 2006, 118, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Haneji, K.; Matsuda, T.; Tomita, M.; Kawakami, H.; Ohshiro, K.; Uchihara, J.; Masuda, M.; Takasu, N.; Tanaka, Y.; Ohta, T.; et al. Fucoidan extracted from Cladosiphon okamuranus Tokida induces apoptosis of human T-cell leukemia virus type 1-infected T-cell lines and primary adult T-cell leukemia cells. Nutr. Cancer 2005, 52, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Nishioka, C.; Ikezoe, T.; Yang, J.; Komatsu, N.; Bandobashi, K.; Taniguchi, A.; Kuwayama, Y.; Togitani, K.; Koeffler, H.; Taguchi, H. Histone deacetylase inhibitors induce growth arrest and apoptosis of HTLV-1-infected T-cells via blockade of signaling by nuclear factor kappaB. Leuk. Res. 2008, 32, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Horie, R.; Kumasaka, T.; Ishida, A.; Ishida, T.; Yamaguchi, K.; Watanabe, T.; Umezawa, K.; Urano, T. In vivo antitumor activity of the NF-kappaB inhibitor dehydroxymethylepoxyquinomicin in a mouse model of adult T-cell leukemia. Carcinogenesis 2005, 26, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Shimoyama, M. Diagnostic criteria and classification of clinical subtypes of adult T-cell leukaemia-lymphoma. A report from the Lymphoma Study Group (1984–87). Br. J. Haematol. 1991, 79, 428–437. [Google Scholar] [CrossRef] [PubMed]
- Tsukasaki, K.; Hermine, O.; Bazarbachi, A.; Ratner, L.; Ramos, J.; Harrington, W., Jr.; O'Mahony, D.; Janik, J.; Bittencourt, A.; Taylor, G.; et al. Definition, prognostic factors, treatment, and response criteria of adult T-cell leukemia-lymphoma: A proposal from an international consensus meeting. J. Clin. Oncol. 2009, 27, 453–459. [Google Scholar] [CrossRef]
- Seegulam, M.; Ratner, L. Integrase Inhibitors Effective Against Human T-Cell Leukemia Virus Type 1. Antimicrob. Agents Chemother. 2011, 55, 2011–2017. [Google Scholar] [CrossRef]
- Ratner, L.; Harrington, W.; Feng, X.; Grant, C.; Jacobson, S.; Noy, A.; Sparano, J.; Lee, J.; Ambinder, R.; Campbell, N.; et al. AIDS Malignancy Consortium. Human T-cell leukemia virus reactivation with progression of adult T-cell leukemia-lymphoma. Plos ONE 2009, 4, e4420. [Google Scholar] [CrossRef]
- Nerenberg, M.; Hinrichs, S.; Reynolds, R.; Khoury, G.; Jay, G. The tat Gene of Human T-Lymphotropic Virus Type 1 Induces Mesenchymal Tumors in Transgenic Mice. Science 1987, 237, 1324–1329. [Google Scholar] [CrossRef]
- Kim, S.; Winokur, T.; Lee, H.; Danielpour, D.; Kim, K.; Geiser, A.; Chen, L.; Sporn, M.; Roberts, A.; Jay, G. Overexpression of transforming growth factor-beta in transgenic mice carrying the human T-cell lymphotropic virus type I tax gene. Mol. Cell. Biol. 1991, 11, 5222–5228. [Google Scholar] [CrossRef]
- Kitajima, I.; Shinohara, T.; Bilakovics, J.; Brown, D.; Xu, X.; Nerenberg, M. Ablation of transplanted HTLV-I Tax-transformed tumors in mice by antisense inhibition of NF-kappa B. Science 1992, 258, 1792–1795. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, S.; Nerenberg, M.; Reynolds, R.; Khoury, G.; Jay, G. A transgenic mouse model for human neurofibromatosis. Science 1987, 237, 1340–1343. [Google Scholar] [CrossRef] [PubMed]
- Green, J.; Baird, A.; Hinrichs, S.; Klintworth, G.; Jay, G. Adrenal medullary tumors and iris proliferation in a transgenic mouse model of neurofibromatosis. Am. J. Pathol. 1992, 140, 1401–1410. [Google Scholar] [PubMed]
- Feigenbaum, L.; Fujita, K.; Collins, F.; Jay, G. Repression of the NF1 gene by Tax may explain the development of neurofibromas in human T-lymphotropic virus type 1 transgenic mice. J. Virol. 1996, 70, 3280–3285. [Google Scholar] [CrossRef]
- Green, J. Trans activation of nerve growth factor in transgenic mice containing the human T-cell lymphotropic virus type I tax gene. Mol. Cell. Biol. 1991, 11, 4635–4641. [Google Scholar]
- Nerenberg, M.; Wiley, C. Degeneration of oxidative muscle fibers in HTLV-1 tax transgenic mice. Am. J. Pathol. 1989, 135, 1025–1033. [Google Scholar]
- Green, J.; Hinrichs, S.; Vogel, J.; Jay, G. Exocrinopathy resembling Sjogren's syndrome in HTLV-1 tax transgenic mice. Nature 1989, 341, 72–74. [Google Scholar] [CrossRef]
- Peebles, R.; Maliszewski, C.; Sato, T.; Hanley-Hyde, J.; Maroulakou, I.; Hunziker, R.; Schneck, J.; Green, J. Abnormal B-cell function in HTLV-I-tax transgenic mice. Oncogene 1995, 10, 1045–1051. [Google Scholar]
- Ruddle, N.; Li, C.; Horne, W.; Santiago, P.; Troiano, N.; Jay, G.; Horowitz, M.; Baron, R. Mice transgenic for HTLV-I LTR-tax exhibit tax expression in bone, skeletal alterations, and high bone turnover. Virology 1993, 197, 196–204. [Google Scholar] [CrossRef]
- Bieberich, C.; King, C.; Tinkle, B.; Jay, G. A transgenic model of transactivation by the Tax protein of HTLV-I. Virology 1993, 196, 309–318. [Google Scholar] [CrossRef]
- Xu, X.; Brown, D.; Kitajima, I.; Bilakovics, J.; Fey, L.; Nerenberg, M. Transcriptional suppression of the human T-cell leukemia virus type I long terminal repeat occurs by an unconventional interaction of a CREB factor with the R region. Mol. Cell. Biol. 1994, 14, 5371–5383. [Google Scholar] [PubMed]
- Furuta, Y.; Aizawa, S.; Suda, Y.; Ikawa, Y.; Kishimoto, H.; Asano, Y.; Tada, T.; Hikikoshi, A.; Yoshida, M.; Seiki, M. Thymic atrophy characteristic in transgenic mice that harbor pX genes of human T-cell leukemia virus type I. J. Virol. 1989, 63, 3185–3189. [Google Scholar] [CrossRef] [PubMed]
- Habu, K.; Nakayama-Yamada, J.; Asano, M.; Saijo, S.; Itagaki, K.; Horai, R.; Yamamoto, H.; Sekiguchi, T.; Nosaka, T.; Hatanaka, M.; et al. The human T cell leukemia virus type I-tax gene is responsible for the development of both inflammatory polyarthropathy resembling rheumatoid arthritis and noninflammatory ankylotic arthropathy in transgenic mice. J. Immunol. 1999, 162, 2956–2963. [Google Scholar] [CrossRef] [PubMed]
- Kishi, S.; Saijo, S.; Arai, M.; Karasawa, S.; Ueda, S.; Kannagi, M.; Iwakura, Y.; Fujii, M.; Yonehara, S. Resistance to fas-mediated apoptosis of peripheral T cells in human T lymphocyte virus type I (HTLV-I) transgenic mice with autoimmune arthropathy. J. Exp. Med. 1997, 186, 57. [Google Scholar] [CrossRef]
- Coscoy, L.; Gonzalez-Dunia, D.; Tangy, F.; Syan, S.; Brahic, M.; Ozden, S. Molecular mechanism of tumorigenesis in mice transgenic for the human T cell leukemia virus Tax gene. Virology 1998, 248, 332–341. [Google Scholar] [CrossRef]
- Benvenisty, N.; Ornitz, D.; Bennett, G.; Sahagan, B.; Kuo, A.; Cardiff, R.; Leder, P. Brain tumours and lymphomas in transgenic mice that carry HTLV-I LTR/c-myc and Ig/tax genes. Oncogene 1992, 7, 2399–2405. [Google Scholar]
- Rauch, D.; Hurchla, M.; Harding, J.; Deng, H.; Shea, L.; Eagleton, M.; Niewiesk, S.; Lairmore, M.; Piwnica-Worms, D.; Rosol, T.; et al. The ARF tumor suppressor regulates bone remodeling and osteosarcoma development in mice. PLoS ONE 2010, 5, e15755. [Google Scholar] [CrossRef]
- Kwon, H.; Ogle, L.; Benitez, B.; Bohuslav, J.; Montano, M.; Felsher, D.; Greene, W. Lethal cutaneous disease in transgenic mice conditionally expressing type I human T cell leukemia virus Tax. J. Biol. Chem. 2005, 280, 35713–35722. [Google Scholar] [CrossRef]
- El Hajj, H.; El-Sabban, M.; Hasegawa, H.; Zaatari, G.; Ablain, J.; Saab, S.; Janin, A.; Mahfouz, R.; Nasr, R.; Kfoury, Y.; et al. Therapy-induced selective loss of leukemia-initiating activity in murine adult T cell leukemia. J. Exp. Med. 2010, 207, 2785–2792. [Google Scholar] [CrossRef]
- Kawaguchi, A.; Orba, Y.; Kimura, T.; Iha, H.; Ogata, M.; Tsuji, T.; Ainai, A.; Sata, T.; Okamoto, T.; Hall, W.; et al. Inhibition of the SDF-1alpha-CXCR4 axis by the CXCR4 antagonist AMD3100 suppresses the migration of cultured cells from ATL patients and murine lymphoblastoid cells from HTLV-I Tax transgenic mice. Blood 2009, 114, 2961–2968. [Google Scholar] [CrossRef]
- Yamazaki, J.; Mizukami, T.; Takizawa, K.; Kuramitsu, M.; Momose, H.; Masumi, A.; Ami, Y.; Hasegawa, H.; Hall, W.; Tsujimoto, H.; et al. Identification of cancer stem cells in a Tax-transgenic (Tax-Tg) mouse model of adult T-cell leukemia/lymphoma. Blood 2009, 114, 2709–2720. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Kumasaka, T.; Okada, S.; Urano, T. The Tax protein of HTLV-1 promotes oncogenesis in not only immature T cells but also mature T cells. Nat. Med. 2007, 13, 527–528. [Google Scholar] [CrossRef] [PubMed]
- Ohsugi, T.; Kumasaka, T. Low CD4/CD8 T-cell ratio associated with inflammatory arthropathy in human T-cell leukemia virus type I Tax transgenic mice. PLoS ONE 2011, 6, e18518. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.; Irvine, J.; Blyth, K.; Cameron, E.; Onions, D.; Campbell, M. Tumours derived from HTLV-I tax transgenic mice are characterized by enhanced levels of apoptosis and oncogene expression. J. Pathol. 1998, 186, 209–214. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Promoter | Gene | Strain | Diseases Observed | Relevance to ATLL | Merits of Model | Weaknesses of Model | Therapies Tested | Ref |
|---|---|---|---|---|---|---|---|---|
| HTLV LTR | Tax | C57BL/6 X DBA/2 X CD1 | Mesenchymal Tumors Thymic atrophy | Not Observed | Overexpression of TGF-beta, activation of NFκB | Tax transforms fibroblasts but not thymocytes | NFκB ODN | [49] [50] [51] |
| Neurofibromatosis Adrenal Tumors | Not Observed | Tax activation of Nerve growth factor and repression of NF1 | Not associated with HTLV associated diseases in humans | None | [52] [53] [54] [55] | |||
| Muscle degeneration | Myositis | High levels of Tax expression in muscle | Incomplete penetrance | None | [56] | |||
| Exocrinopathy Lymphadenopathy Splenomegaly | Sjogren syndrome ocular lesions | NfκB inflammatory disorders associated with Tax | Caused by B not T lymphocytes | None | [57] [58] | |||
| Bone Turnover | Lytic bone lesions | NFkB associated bone lesions | Incomplete penetrance | None | [59] | |||
| Tax βgal | Mesenchymal Tumors | Not Observed | Tax expression in response to tissue damage | Tissue damage not correlated with tumor | None | [60] [61] | ||
| HTLV LTR | pX | C57BL/6 X CD1 | Thymic Atrophy | Not Observed | Effects of pX on thymus independent of promoter used | pX gene expression not detectable | None | [62] |
| Ig-SV40 | ||||||||
| MMTV LTR | ||||||||
| HTLV LTR | pX | C3H/ HeN | Inflammatory Arthropathy Osteogenesis Autoimmunity | Arthritis | IL-1, IL-6, TNFα, TGFβ detected in joints. | No malignancy | Anti-Fas mAb (RK-8) | [4] [5] [7] [63] [64] |
| Tax | ||||||||
| CD4 | Tax | |||||||
| HTLV LTR | Tax | C3H | Mesenchymal Tumors | NFκB mediated malignancy | IκB degradation leads to constitutive NFκB activation | Expression restricted to CNS and testes | None | [65] |
| Ig | Tax | FVB/N | Not Observed | Not Observed | Lymphoma with CNS involvement | Roles of c-Myc and Tax unclear | None | [66] |
| Ig HTLV LTR | Tax c-myc | CD4+ Lymphoma CNS Tumors | CD4+ Lymphoma | |||||
| GzmB | Tax | C57BL/6 | LGL lymphoma Leukemia Osteolytic lesions Splenomegaly Lymphadenopathy Hypercalcemia | Lymphoma Leukemia Lytic bone lesions Hypercalcemia | NFkB mediated leukemia lymphoma | Not a CD4+ T cell malignancy | Bortezomib | [8] [36] |
| Tax IL-2−/− | IL-2 not required for phenotype | Not a CD4+ T cell malignancy | None | [29] | ||||
| Tax IFNγ−/− | Accelerated tumor onset and death | May also affect tumor immunity | None | [32] | ||||
| Tax P53−/− | Accelerated disease progression | Only seen in P53+/− mice | None | [22] | ||||
| GzmB ApoE | Tax OPG | Reduced cancer and bone lesions | Causes osteopetrosis | Zoledronic Acid | [27] | |||
| GzmB HTLV LTR | Tax LUC | C57BL/6 X FVB | Bioluminescent tumors | Not a CD4+ T cell malignancy | None | [31] | ||
| GzmB HTLV LTR | Tax LUC ARF−/− | Lymphoma Bone Turnover Osteosarcoma | ARF−/− is not equivalent to p53−/− | Osteosarcoma not associated with ATLL | Zoledronic Acid | [67] | ||
| GzmB HTLV LTR TCR | Tax LUC TCRova | C57BL/6 X FVB X BALB/c | Leukemia Lymphoma Lymphadenopathy | Tax induced by wounding and T cell activation leads to enhanced tumorigenesis | Primary malignancy not a not a CD4+ T cell malignancy | None | [35] | |
| EμSRα TET TET | tTA Tax M47 | FVB/N | Alopecia Hyperkeratosis Splenomegaly | Skin Lesions | Tet-inducible model allows repression of Tax | No malignancy | None | [68] |
| EμSRα TET | tTA M22 | Not observed | Not Observed | Control establishes role of NFκB in disease | ||||
| Lck-prox | Tax | C57BL/6 | CD4- CD25+ pre-T cell Leukemia Lymphoma | Leukemia Lymphoma | Cancer stem cells derived from these mice recapitulate disease in SCID | Not a CD4+ T cell malignancy | As2O3 + IFN-α AMD3100 | [9] [69] [70] [71] |
| Lck-dis | Tax | C57BL/6 X DBA/2 | CD4+ CD25- mature T cell leukemia lymphoma Arthritis | Leukemia Lymphoma | Mature CD4+ or CD8+ T cell malignancy | Cells lack CD25 | None | [72] [73] |
| CD3-ε | Tax | C57BL/6 X CBA | Mesencymal tumors Mammary Adenoma | Not Observed | Tax associated with apoptosis and p53 | Not a CD4+ T cell malignancy | None | [74] |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rauch, D.A.; Ratner, L. Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients. Viruses 2011, 3, 886-900. https://doi.org/10.3390/v3060886
Rauch DA, Ratner L. Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients. Viruses. 2011; 3(6):886-900. https://doi.org/10.3390/v3060886
Chicago/Turabian StyleRauch, Daniel A., and Lee Ratner. 2011. "Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients" Viruses 3, no. 6: 886-900. https://doi.org/10.3390/v3060886
APA StyleRauch, D. A., & Ratner, L. (2011). Targeting HTLV-1 Activation of NFκB in Mouse Models and ATLL Patients. Viruses, 3(6), 886-900. https://doi.org/10.3390/v3060886