Role of Human Endogenous Retroviral Long Terminal Repeats (LTRs) in Maintaining the Integrity of the Human Germ Line

{kind=link}
Abstract
:1. ERV-9 LTR Driven Human Transcripts
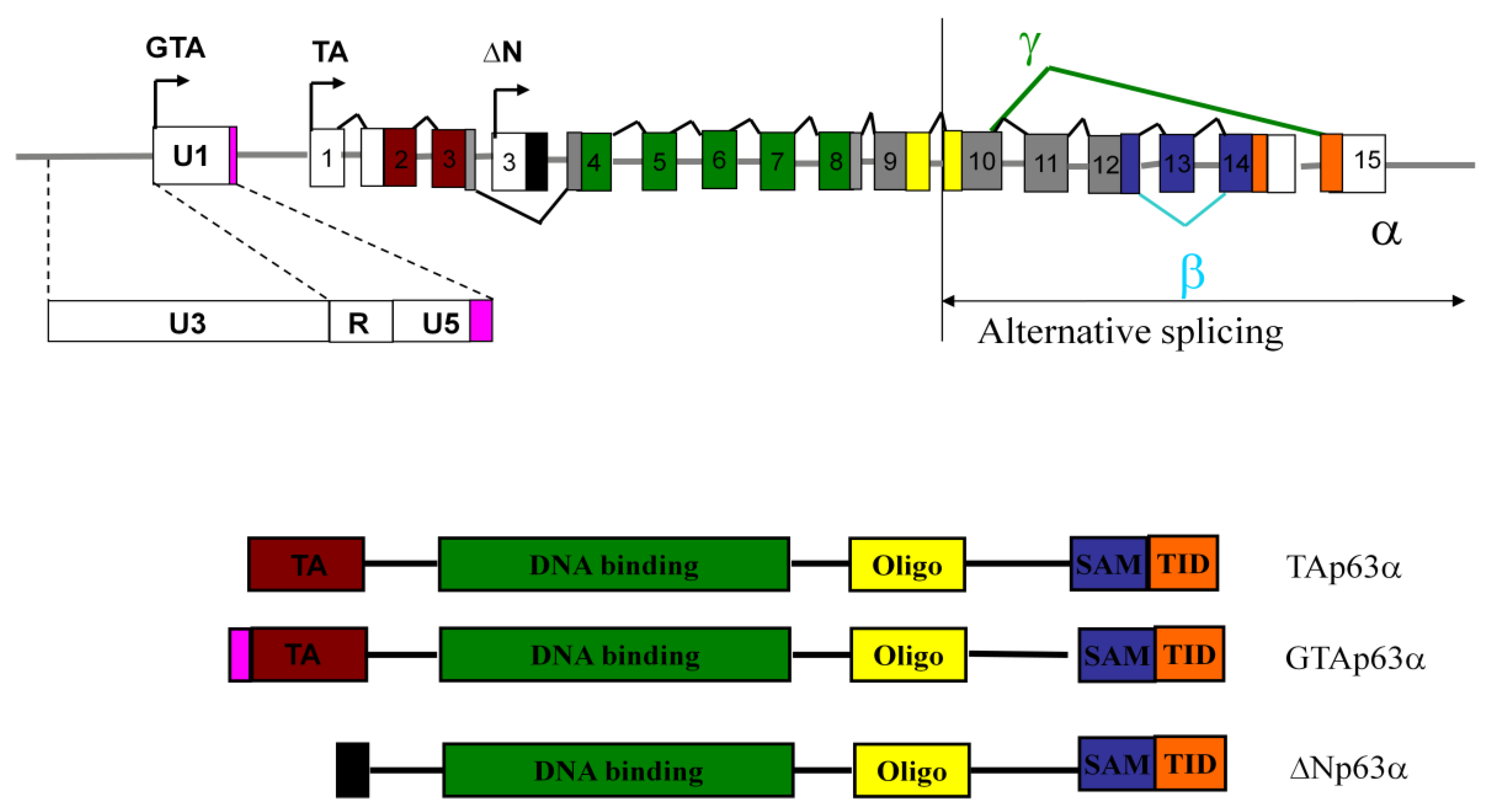
2. TAp63 Isoforms Present in the Human p63 Locus
3. GTAp63 is Expressed in Male Germ Cells
Acknowledgements
References and Notes
- Beyer, U.; Moll-Rocek, J.; Moll, U.M.; Dobbelstein, M. Endogenous retrovirus drives hitherto unknown proapoptotic p63 isoforms in the male germ line of humans and great apes. Proc. Natl. Acad. Sci. U. S. A. 2011, 108, 3624–3629. [Google Scholar] [CrossRef] [PubMed]
- Crum, C.P.; McKeon, F.D. p63 in epithelial survival, germ cell surveillance, and neoplasia. Annu. Rev. Pathol. 2010, 5, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Ting, C.N.; Rosenberg, M.P.; Snow, C.M.; Samuelson, L.C.; Meisler, M.H. Endogenous retroviral sequences are required for tissue-specific expression of a human salivary amylase gene. Genes Dev. 1992, 6, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Medstrand, P.; Landry, J.R.; Mager, D.L. Long terminal repeats are used as alternative promoters for the endothelin B receptor and apolipoprotein C-I genes in humans. J. Biol. Chem. 2001, 276, 1896–1903. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.J.; Lock, W.M.; Mager, D.L. Endogenous retroviral LTRs as promoters for human genes: a critical assessment. Gene 2009, 448, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Pi, W.; Bollag, R.; Zeng, S.; Keskintepe, M.; Saliman, H.; Krantz, S.; Whitney, B.; Tuan, D. The solitary long terminal repeats of ERV-9 endogenous retrovirus are conserved during primate evolution and possess enhancer activities in embryonic and hematopoietic cells. J. Virol. 2002, 76, 2410–2423. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhu, X.; Pi, W.; Ling, J.; Ko, L.; Takeda, Y.; Tuan, D. The long terminal repeat (LTR) of ERV-9 human endogenous retrovirus binds to NF-Y in the assembly of an active LTR enhancer complex NF-Y/MZF1/GATA-2. J. Biol. Chem. 2005, 280, 35184–35194. [Google Scholar] [CrossRef]
- Siggers, P.; Smith, L.; Greenfield, A. Sexually dimorphic expression of Gata-2 during mouse gonad development. Mech. Dev. 2002, 111, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Zon, L.I.; Mather, C.; Burgess, S.; Bolce, M.E.; Harland, R.M.; Orkin, S.H. Expression of GATA- binding proteins during embryonic development in Xenopus laevis. Proc. Natl. Acad. Sci. U. S. A. 1991, 88, 10642–10646. [Google Scholar] [CrossRef]
- Gove, C.; Walmsley, M.; Nijjar, S.; Bertwistle, D.; Guille, M.; Partington, G.; Bomford, A.; Patient, R. Over-expression of GATA-6 in Xenopus embryos blocks differentiation of heart precursors. EMBO J. 1997, 16, 355–368. [Google Scholar] [CrossRef]
- Partington, G.A.; Bertwistle, D.; Nicolas, R.H.; Kee, W.J.; Pizzey, J.A.; Patient, R.K. GATA-2 is a maternal transcription factor present in Xenopus oocytes as a nuclear complex which is maintained throughout early development. Dev. Biol. 1997, 181, 144–155. [Google Scholar] [CrossRef]
- Pi, W.; Zhu, X.; Wu, M.; Wang, Y.; Fulzele, S.; Eroglu, A.; Ling, J.; Tuan, D. Long-range function of an intergenic retrotransposon. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 12992–12997. [Google Scholar] [CrossRef] [PubMed]
- Pi, W.; Yang, Z.; Wang, J.; Ruan, L.; Yu, X.; Ling, J.; Krantz, S.; Isales, C.; Conway, S.J.; Lin, S.; et al. The LTR enhancer of ERV-9 human endogenous retrovirus is active in oocytes and progenitor cells in transgenic zebrafish and humans. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 805–810. [Google Scholar] [CrossRef] [PubMed]
- May, P.; May, E. Twenty years of p53 research: Structural and functional aspects of the p53 protein. Oncogene 1999, 18, 7621–7636. [Google Scholar] [CrossRef] [PubMed]
- Suh, E.K.; Yang, A.; Kettenbach, A.; Bamberger, C.; Michaelis, A.H.; Zhu, Z.; Elvin, J.A.; Bronson, R.T.; Crum, C.P.; McKeon, F. p63 protects the female germ line during meiotic arrest. Nature 2006, 444, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Kaghad, M.; Caput, D.; McKeon, F. On the shoulders of giants: p63, p73 and the rise of p53. Trends Genet. 2002, 18, 90–95. [Google Scholar] [CrossRef] [PubMed]

© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, M.; Eiden, M.V. Role of Human Endogenous Retroviral Long Terminal Repeats (LTRs) in Maintaining the Integrity of the Human Germ Line. Viruses 2011, 3, 901-905. https://doi.org/10.3390/v3060901
Liu M, Eiden MV. Role of Human Endogenous Retroviral Long Terminal Repeats (LTRs) in Maintaining the Integrity of the Human Germ Line. Viruses. 2011; 3(6):901-905. https://doi.org/10.3390/v3060901
Chicago/Turabian StyleLiu, Meihong, and Maribeth V. Eiden. 2011. "Role of Human Endogenous Retroviral Long Terminal Repeats (LTRs) in Maintaining the Integrity of the Human Germ Line" Viruses 3, no. 6: 901-905. https://doi.org/10.3390/v3060901
APA StyleLiu, M., & Eiden, M. V. (2011). Role of Human Endogenous Retroviral Long Terminal Repeats (LTRs) in Maintaining the Integrity of the Human Germ Line. Viruses, 3(6), 901-905. https://doi.org/10.3390/v3060901