
Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments

 , ,
, ,
Abstract
:1. Introduction

2. The WSSV Genome and miRNAS
2.1. WSSV Genome
2.2. miRNAs, WSSV Infection, and Pathogenesis
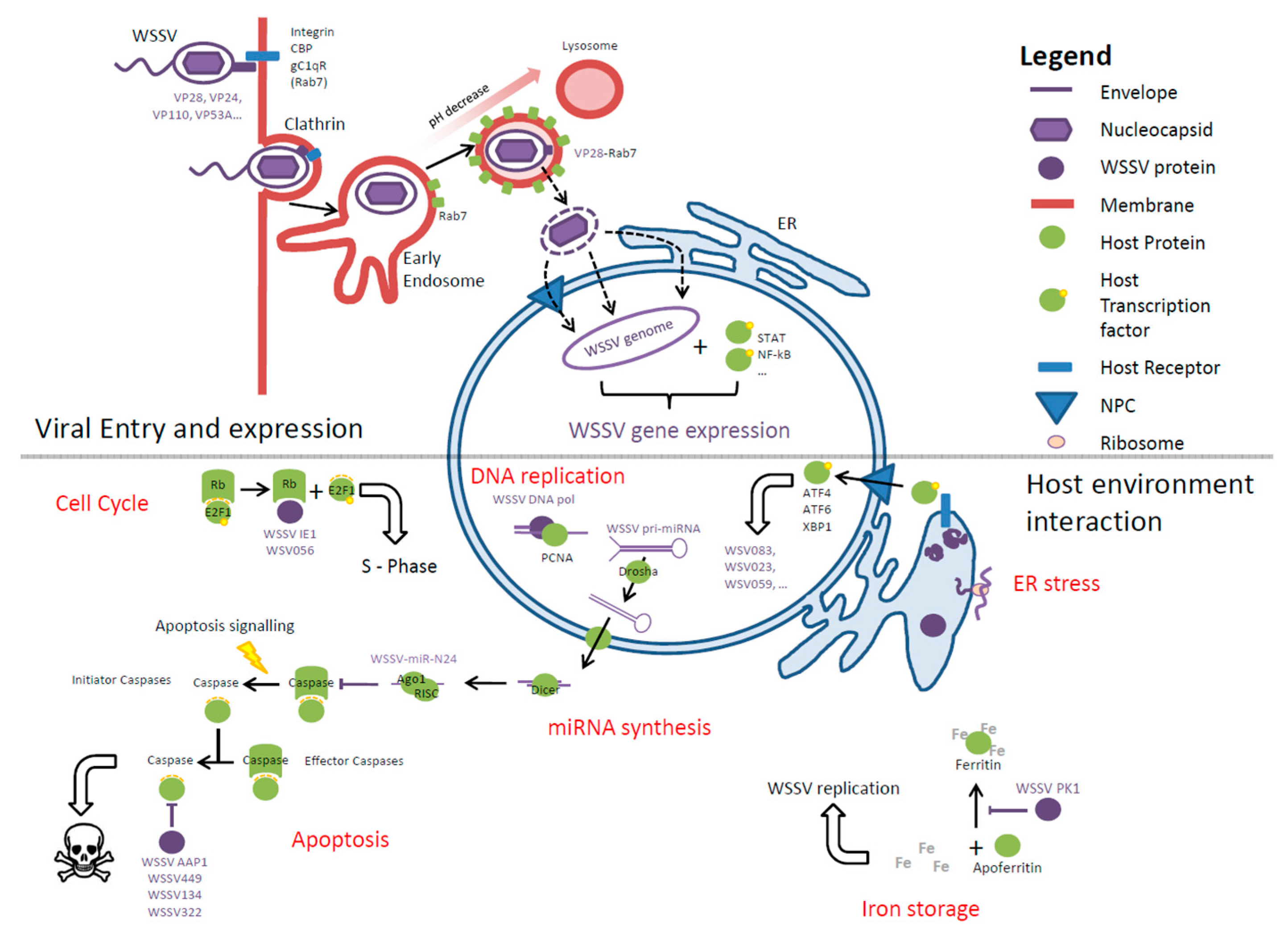
3. WSSV Infection

3.1. Viral Receptors, Interactions, and Entry

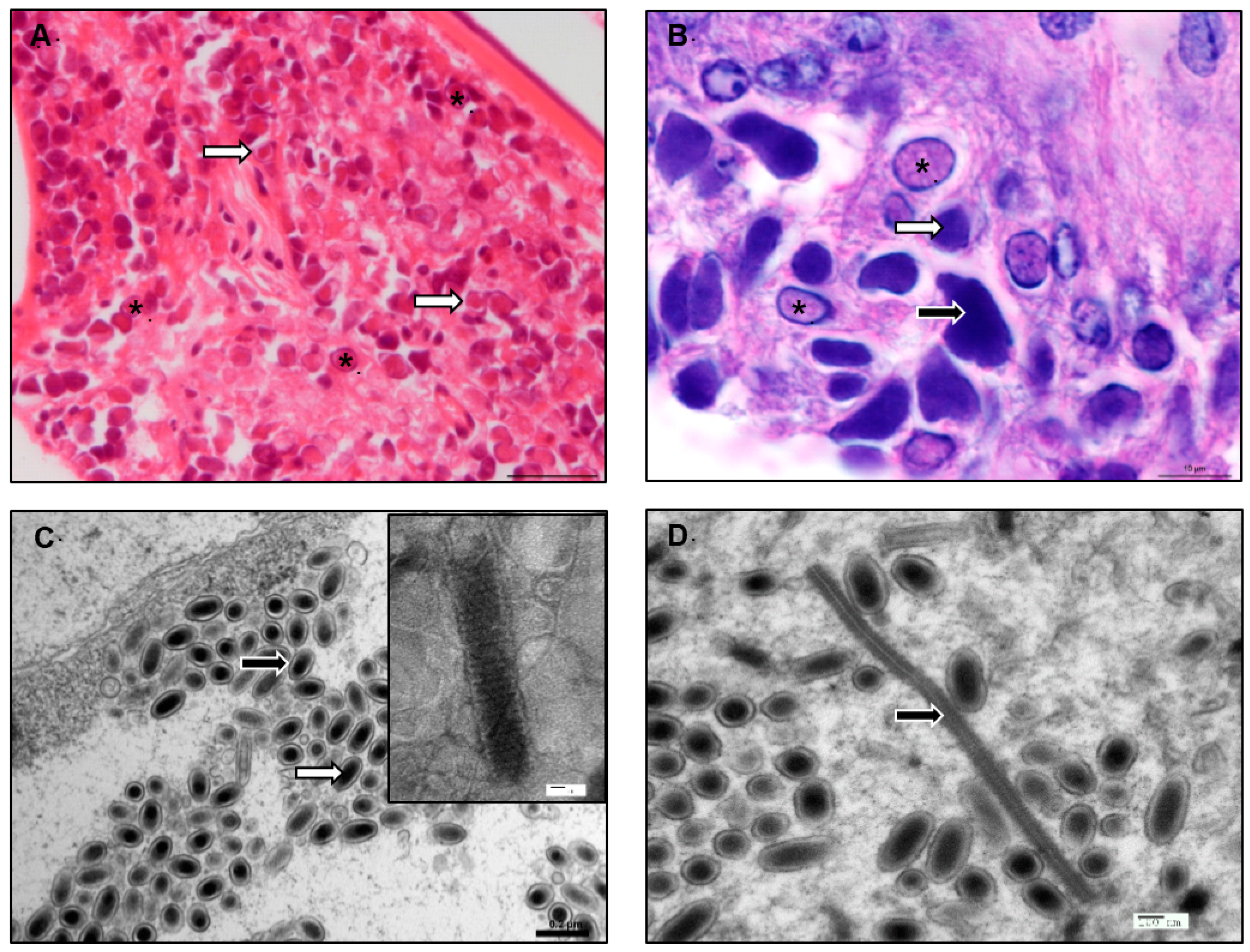{kind=link}
{kind=link}
| Viral Protein | Host Protein | Species | Reference |
|---|---|---|---|
| VP24, VP32, VP39B, VP41A, VP51B, VP53A, VP53B, VP60A, VP110, VP124, VP337 | Chitin-binding protein (PmCBP) | Penaeus monodon | [89,90] |
| VP53A | Glut1 | P. monodon | [91] |
| VP15, VP26, VP28 | gC1qR (PlgC1qR) | Pacifastacus leniusculus | [92] |
| VP95, VP28, VP26, VP24, VP19, VP14 | C-type lectin (LvCTL1) | Litopenaeus vannamei | [93] |
| VP28 | C-type lectin (FcLec3) | Fenneropenaeus chinensis | [94] |
| VP26, VP28 | C-type lectins (MjLecA, MjLecB, MjLecC) | Marsupenaeus japonicus | [95] |
| VP28 | C-type lectins (MjsvCL) | M. japonicus | [96] |
| VP28 | C-type lectins (LdlrLec1, LdlrLec2) | M. japonicus | [97] |
| VP187 | β-Integrin | P. japonicus | [98] |
| P. clarkii | |||
| VP26, VP31, VP37, VP90, VP136 | β-Integrin | L. vannamei | [99] |
| WSSV-CLP | α-integrin, β-integrin, Syndecan | F. chinensis | [100] (Bioinformatic prediction) |
| VP15, VP28 | Calreticulin (PlCRT) | P. leniusculus | [101] |
| VP466 | Rab (PjRab) | P. japonicus | [102] |
| VP28 | Rab7 (PmRab7) | P. monodon | [103] |
| ORF514 | PCNA (lvPCNA) | L. vannamei | [104] (Bioinformatic prediction) |
| WSSV PK1 | Ferritin (lvFerritin) | L. vannamei | [105] |
| Wsv083 | FAK (MjFAK) | M. japonicus | [106] |
| AAP1 (WSSV449) | Caspase (PmCaspase) | P. monodon | [107] |
| WSSV134, WSSV332 | Caspase (PmCasp) | P. monodon | [108] |
| WSSV249 | Ubc (PvUbc) | L. vannamei | [109] |
| ICP11 | Histones | P. monodon | [110] |
| VP9 | RACK1 (PmRACK1) | P. monodon | [111] |
| VP15 | FKBP46 (PmFKBP46) | P. monodon | [112] |
| VP15 | CRT (PlgCRT) | P. leniusculus | [101] |
| WSSV-miRNA | Dorsha, Dicer, Ago1 | – | [77] |
| VP14 | Arginine kinase (LvAK) | L. vannamei | [113] |
| VP26 | Actin | Procambarus clarkii | [114] |
| ORF427 | PPs | L. vannamei | [115] |
| WSSV IE1 | TATA box-binding protein (PmTBP) | P. monodon | [116] |
| WSSV IE1, WSV056 | Retinoblastoma protein (Lv-RBL) | L. vannamei | [117] |
3.2. Escaping from Endosomes
3.3. Viral Replication—The Molecular Processes
| Transcription Factor (Host, Virus) | Target (Host, Virus) | Species | Reference |
|---|---|---|---|
| STAT (PmSTAT) | ie1 | Penaeus monodon | [136] |
| PHB2 (Sf-PHB2) | ie1 | Spodoptera frugiperda | [137] |
| Nf-κB (LvRelish, LvDorsal) | ie1, WSSV303, WSSV371 | Litopenaeus Vannamei | [138,139,140] |
| c-JUN | ie1 | L. vannamei | [141] (Bioinformatic prediction) |
| XBP1 (LvXBP1) | wsv083 | L. vannamei | [142] |
| ATF4 (LvATF4) | wsv023 | L. vannamei | [142] |
| KLF (PmKLF) | WSSV108 | P. monodon | [143] |
| ATFβ (LvATFβ) | wsv059, wsv166 | L. vannamei | [144] |
| VP38, VP41B | Caspase (PjCaspase) | M. japonicus | [145] |
| WSSV-miR-N24 | Caspase 8 | M. japonicus | [78] |
3.4. Maintaining the Host Cell Environment
3.4.1. Metabolism
3.4.2. Iron
3.4.3. Endoplasmic Reticulum Stress Responses
3.4.4. Apoptosis
3.4.5. Particle Assembly and Release
4. Current Treatment Options for WSD
5. Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stentiford, G.D.; Oidtmann, B.; Scott, A.; Peeler, E.J. Crustacean diseases in European legislation: Implications for importing and exporting nations. Aquaculture 2010, 306, 27–34. [Google Scholar] [CrossRef]
- Stentiford, G.D.; Bonami, J.R.; Alday-Sanz, V. A critical review of susceptibility of crustaceans to taura syndrome, yellowhead disease and white spot disease and implications of inclusion of these diseases in European legislation. Aquaculture 2009, 291, 1–17. [Google Scholar] [CrossRef]
- Nakano, H.; Koube, H.; Umezawa, S.; Momoyama, K.; Hiraoka, M.; Inouye, K.; Oseko, N. Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in Japan in 1993: Epizootiological survey and infection trails. Fish Pathol. 1994, 29, 135–139. [Google Scholar] [CrossRef]
- Flegel, T.W. Special topic review: Major viral diseases of the black tiger prawn (Penaeus monodon) in thailand. World J. Microbiol. Biotechnol. 1997, 13, 433–442. [Google Scholar] [CrossRef]
- Mohan, C.V.; Shankar, K.M.; Kulkarni, S.; Sudha, P.M. Histopathology of cultured shrimp showing gross signs of yellow head syndrome and white spot syndrome during 1994 Indian epizootics. Dis. Aquat. Org. 1998, 34, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Zhan, W.B.; Wang, Y.H.; Fryer, J.L.; Yu, K.K.; Fukuda, H.; Meng, Q.X. White spot syndrome virus infection of cultured shrimp in China. J. Aquat. Anim. Health 1998, 10, 405–410. [Google Scholar] [CrossRef]
- Wang, Q.; Poulos, B.T.; Lightner, D.V. Protein analysis of geographic isolates of shrimp white spot syndrome virus. Arch. Virol. 2000, 145, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Stentiford, G.D.; Lightner, D.V. Cases of white spot disease (WSD) in European shrimp farms. Aquaculture 2011, 319, 302–306. [Google Scholar] [CrossRef]
- Lightner, D.V. Global transboundary disease politics: The OIE perspective. J. Invertebr. Pathol. 2012, 110, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Flegel, T.W.; Lightner, D.V.; Owens, L. Shrimp disease control: Past, present and future. Dis. Asian Aquacult. 2008, 6, 355–378. [Google Scholar]
- Stentiford, G.D.; Neil, D.M.; Peeler, E.J.; Shields, J.D.; Small, H.J.; Flegel, T.W.; Vlak, J.M.; Jones, B.; Morado, F.; Moss, S.; et al. Disease will limit future food supply from the global crustacean fishery and aquaculture sectors. J. Invertebr. Pathol. 2012, 110, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.D. Research priorities for diseases of the blue crab Callinectes sapidus. Bull. Mar. Sci. 2003, 72, 505–517. [Google Scholar]
- Wang, C.H.; Lo, C.F.; Leu, J.H.; Chou, C.M.; Yeh, P.Y.; Chou, H.Y.; Tung, M.C.; Chang, C.F.; Su, M.S.; Kou, G.H. Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dis. Aquat. Org. 1995, 23, 239–242. [Google Scholar] [CrossRef]
- Mayo, M. A summary of taxonomic changes recently approved by ictv. Arch. Virol. 2002, 147. [Google Scholar] [CrossRef] [PubMed]
- Mayo, M. Virus taxonomy—Houston 2002. Arch. Virol. 2002, 147. [Google Scholar] [CrossRef]
- Van Hulten, M.C.; Witteveldt, J.; Peters, S.; Kloosterboer, N.; Tarchini, R.; Fiers, M.; Sandbrink, H.; Lankhorst, R.K.; Vlak, J.M. The white spot syndrome virus DNA genome sequence. Virology 2001, 286, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Vlak, J.M.; Bonami, J.R.; Flegel, T.W.; Kou, G.H.; Lightner, D.V.; Lo, C.F.; Loh, P.C.; Walker, P.W. Nimaviridae. In Eighth Report of the International Committee on Taxonomy of Viruses; Elsevier/Academic Press: Cambridge, MA, USA, 2005; pp. 187–192. [Google Scholar]
- Pradeep, B.; Rai, P.; Mohan, S.A.; Shekhar, M.S.; Karunasagar, I. Biology, host range, pathogenesis and diagnosis of white spot syndrome virus. Indian J. Virol. 2012, 23, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.Y.; Huang, C.Y.; Wang, C.H.; Chiang, H.C.; Lo, C.F. Pathogenicity of a baculovirus infection causing white spot syndrome in cultured penaeid shrimp in Taiwan. Dis. Aquat. Org. 1995, 23, 165–173. [Google Scholar] [CrossRef]
- Rajan, P.R.; Ramasamy, P.; Purushothaman, V.; Brennan, G.P. White spot baculovirus syndrome in the indian shrimp Penaeus monodon and P. Indicus. Aquaculture 2000, 184, 31–44. [Google Scholar] [CrossRef]
- Wang, Y.G.; Hassan, M.D.; Shariff, M.; Zamri, S.M.; Chen, X. Histopathology and cytopathology of white spot syndrome virus (WSSV) in cultured Penaeus monodon from peninsular malaysia with emphasis on pathogenesis and the mechanism of white spot formation. Dis. Aquat. Org. 1999, 39, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lightner, D.V. A Handbook of Shrimp Pathology and Diagnostic Procedures for Diseases of Cultured Penaeid Shrimp; World Aquaculture Society: Baton Roughe, LA, USA, 1996; p. 304. [Google Scholar]
- Durand, S.; Lightner, D.V.; Redman, R.M.; Bonami, J.R. Ultrastructure and morphogenesis of white spot syndrome baculovirus (WSSV). Dis. Aquat. Org. 1997, 29, 205–211. [Google Scholar] [CrossRef]
- Nadala, E.C.B.; Loh, P.C. A comparative study of three different isolates of white spot virus. Dis. Aquat. Org. 1998, 33, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Oidtmann, B.; Stentiford, G.D. White spot syndrome virus (WSSV) concentrations in crustacean tissues: A review of data relevant to assess the risk associated with commodity trade. Transbound. Emerg. Dis. 2011, 58, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.J.; Mohan, C.V. Viral disease emergence in shrimp aquaculture: Origins, impact and the effectiveness of health management strategies. Rev. Aquacult. 2009, 1, 125–154. [Google Scholar] [CrossRef]
- Momoyama, K.; Hiraoka, M.; Nakano, H.; Koube, H.; Inouye, K.; Oseko, N. Mass mortalities of cultured kuruma shrimp, Penaeus japonicus, in japan in 1993: Histopathological study. Fish Pathol. 1994, 29, 141–148. [Google Scholar] [CrossRef]
- Takahashi, Y.; Itami, T.; Kondom, M.; Maeda, M.; Fuji, R.; Tomonaga, S.; Supamattaya, K.; Boonyaratpalin, S. Electron microscopic evidence of baciliform virus infection in kuruma shrimp (Penaeus japonicus). Fish Pathol. 1994, 29, 121–125. [Google Scholar] [CrossRef]
- Cai, S.; Huang, J.; Wang, C.; Song, X.; Sun, X.; Yu, J.; Zhang, Y.; Yang, C. Epidemiological studies on the explosive epidemic disease of prawn in 1993–1994. J. Fishery Sci. China 1995, 19, 112–117. [Google Scholar]
- Chen, L.L.; Lo, C.F.; Chiu, Y.L.; Chang, C.F.; Kou, G.H. Natural and experimental infection of white spot syndrome virus (WSSV) in benthic larvae of mud crab Scylla setrata. Dis. Aquat. Org. 2000, 40, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Sahul Hameed, A.S.; Charles, M.X.; Anilkumar, M. Tolerance of Macrobrachium rosenbergii to white spot syndrome virus. Aquaculture 2000, 183, 207–213. [Google Scholar] [CrossRef]
- Hossain, M.S.; Chakraborty, A.; Joseph, B.; Otta, S.K.; Karunasagar, I. Detection of new hosts for white spot syndrome virus of shrimp using nested polymerase chain reaction. Aquaculture 2001, 198, 1–11. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Bangyeekhun, E.; Soderhall, K.; Soderhall, I. Experimental infection of white spot syndrome virus in freshwater crayfish Pacifastacus leniusculus. Dis. Aquat. Org. 2001, 47, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Bayot, B.; Amano, Y.; Panchana, F.; de Blas, I.; Alday, V.; Calderon, J. White spot syndrome virus infection in cultured Penaeus vannamei (boone) in ecuador with emphasis on histopathology and ultrastructure. J. Fish Dis. 2003, 26, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Yoganandhan, K.; Thirupathi, S.; Sahul Hameed, A.S. Biochemical, physiological and hematological changes in white spot syndrome virus-infected shrimp, Penaeus indicus. Aquaculture 2003, 221, 1–11. [Google Scholar] [CrossRef]
- Bateman, K.S.; Tew, I.; French, C.; Hicks, R.J.; Martin, P.; Munro, J.; Stentiford, G.D. Susceptibility to infection and pathogenicity of white spot disease (WSD) in non-model crustacean host taxa from temperate regions. J. Invertebr. Pathol. 2012, 110, 340–351. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.F.; Ho, C.H.; Chen, C.H.; Liu, K.F.; Chiu, Y.L.; Yeh, P.Y.; Peng, S.E.; Hsu, H.C.; Liu, H.C.; Chang, C.F.; et al. Detection and tissue tropism of white spot syndrome baculovirus (WSBV) in captured brooders of Penaeus monodon with a special emphasis on reproductive organs. Dis. Aquat. Org. 1997, 30, 53–72. [Google Scholar] [CrossRef]
- Chou, H.Y.; Huang, C.Y.; Lo, C.F.; Kou, G.H. Studies on transmission of white spot syndrome associated baculovirus (WSBV) in Penaeus monodon and P. japonicus via waterborne contact and oral ingestion. Aquaculture 1998, 164, 263–276. [Google Scholar] [CrossRef]
- Lotz, J.M.; Soto, M.A. Model of white spot syndrome virus (WSSV) epidemics in Litopenaeus vannamei. Dis. Aquat. Org. 2002, 50, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Tuyen, N.X.; Verreth, J.; Vlak, J.M.; de Jong, M.C. Horizontal transmission dynamics of white spot syndrome virus by cohabitation trials in juvenile Penaeus monodon and P. vannamei. Prev. Vet. Med. 2014, 117, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.F.; Ho, C.H.; Peng, S.E.; Chen, C.H.; Hsu, H.C.; Chiu, Y.L.; Chang, C.F.; Liu, K.F.; Su, M.S.; Wang, C.H.; et al. White spot syndrome baculovirus (WSBV) detected in cultured and captured shrimp, crabs and other arthropods. Dis. Aquat. Org. 1996, 27, 215–225. [Google Scholar] [CrossRef]
- Waikhom, G.; John, K.R.; George, M.R.; Jeyaseelan, M.J.P. Differential host passaging alters pathogenicity and induces genomic variation in white spot syndrome virus. Aquaculture 2006, 261, 54–63. [Google Scholar] [CrossRef]
- Nunan, L.M.; Poulos, B.T.; Lightner, D.V. The detection of white spot syndrome virus (WSSV) and yellow head virus (YHV) in imported commodity shrimp. Aquaculture 1998, 160, 19–30. [Google Scholar] [CrossRef]
- Durand, S.V.; Tang, K.F.J.; Lightner, D.V. Frozen commodity shrimp: Potential avenue for introduction of white spot syndrome virus and yellow head virus. J. Aquat. Anim. Health 2000, 12, 128–135. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Signaling pathways regulating innate immune responses in shrimp. Fish Shellfish Immunol. 2013, 34, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, M.S.; Ponniah, A.G. Recent insights into host-pathogen interaction in white spot syndrome virus infected penaeid shrimp. J. Fish Dis. 2014, 38, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Paz, A. White spot syndrome virus: An overview on an emergent concern. Vet. Res. 2010, 41, 43. [Google Scholar] [CrossRef] [PubMed]
- Sritunyalucksana, K.; Utairungsee, T.; Sirikharin, R.; Srisala, J. Virus-binding proteins and their roles in shrimp innate immunity. Fish Shellfish Immunol. 2012, 33, 1269–1275. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; He, J.; Lin, X.; Li, Q.; Pan, D.; Zhang, X.; Xu, X. Complete genome sequence of the shrimp white spot bacilliform virus. J. Virol. 2001, 75, 11811–11820. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.Y.; Yoon, J.; Lee, Y.S.; Kim, Y.B.; Choi, T.J. Analysis of the complete nucleotide sequence of a white spot syndrome virus isolated from Pacific white shrimp. J. Microbiol. 2013, 51, 695–699. [Google Scholar] [CrossRef] [PubMed]
- Marks, H.; Goldbach, R.W.; Vlak, J.M.; van Hulten, M.C. Genetic variation among isolates of white spot syndrome virus. Arch. Virol. 2004, 149, 673–697. [Google Scholar] [CrossRef] [PubMed]
- Sindhupriya, M.; Saravanan, M.; Otta, S.K.; Bala Amarnath, C.; Arulraj, R.; Bhuvaneswari, T.; Ezhil Praveena, P.; Jithendran, K.P.; Ponniah, A.G. White spot syndrome virus (WSSV) genome stability maintained over six passages through three different penaeid shrimp species. Dis. Aquat. Org. 2014, 111, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Shekar, M.; Pradeep, B.; Karunasagar, I. White spot syndrome virus: Genotypes, epidemiology and evolutionary studies. Indian J. Virol. 2012, 23, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Marks, H.; Vorst, O.; van Houwelingen, A.M.; van Hulten, M.C.; Vlak, J.M. Gene-expression profiling of white spot syndrome virus in vivo. J. Gen. Virol. 2005, 86, 2081–2100. [Google Scholar] [CrossRef] [PubMed]
- Sablok, G.; Sanchez-Paz, A.; Wu, X.; Ranjan, J.; Kuo, J.; Bulla, I. Genome dynamics in three different geographical isolates of white spot syndrome virus (WSSV). Arch. Virol. 2012, 157, 2357–2362. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.M.; Wang, H.C.; Leu, J.H.; Hsiao, H.H.; Wang, A.H.; Kou, G.H.; Lo, C.F. Genomic and proteomic analysis of thirty-nine structural proteins of shrimp white spot syndrome virus. J. Virol. 2004, 78, 11360–11370. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.M.; Wang, H.C.; Leu, J.H.; Wang, A.H.; Zhuang, Y.; Walker, P.J.; Kou, G.H.; Lo, C.F. Identification of the nucleocapsid, tegument, and envelope proteins of the shrimp white spot syndrome virus virion. J. Virol. 2006, 80, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.-H.; Tsai, J.-M.; Wang, H.-C.; Wang, A.H.J.; Wang, C.-H.; Kou, G.-H.; Lo, C.-F. The unique stacked rings in the nucleocapsid of the white spot syndrome virus virion are formed by the major structural protein VP664, the largest viral structural protein ever found. J. Virol. 2005, 79, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Yu, H.T.; Peng, S.E.; Chang, Y.S.; Pien, H.W.; Lin, C.J.; Huang, C.J.; Tsai, M.F.; Huang, C.J.; Wang, C.H.; et al. Cloning, characterization, and phylogenetic analysis of a shrimp white spot syndrome virus gene that encodes a protein kinase. Virology 2001, 289, 362–377. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.F.; Lo, C.F.; van Hulten, M.C.; Tzeng, H.F.; Chou, C.M.; Huang, C.J.; Wang, C.H.; Lin, J.Y.; Vlak, J.M.; Kou, G.H. Transcriptional analysis of the ribonucleotide reductase genes of shrimp white spot syndrome virus. Virology 2000, 277, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Leu, J.H.; Huang, C.J.; Chou, C.M.; Chen, S.M.; Wang, C.H.; Lo, C.F.; Kou, G.H. Identification of a nucleocapsid protein (vp35) gene of shrimp white spot syndrome virus and characterization of the motif important for targeting vp35 to the nuclei of transfected insect cells. Virology 2002, 293, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.T.; Wang, H.C.; Yang, Y.T.; Kou, G.H.; Lo, C.F. The DNA virus white spot syndrome virus uses an internal ribosome entry site for translation of the highly expressed nonstructural protein ICP35. J. Virol. 2013, 87, 13263–13278. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.T.; Leu, J.H.; Wang, H.C.; Chen, L.L.; Kou, G.H.; Lo, C.F. Polycistronic mrnas and internal ribosome entry site elements (IRES) are widely used by white spot syndrome virus (WSSV) structural protein genes. Virology 2009, 387, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhang, X. Internal initiation of mrna translation in insect cell mediated by an internal ribosome entry site (IRES) from shrimp white spot syndrome virus (WSSV). Biochem. Biophys. Res. Commun. 2006, 344, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Gale, M.; Tan, S.L.; Katze, M.G. Translational control of viral gene expression in eukaryotes. Microbiol. Mol. Biol. Rev. 2000, 64, 239–280. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P.; Chen, C.-Z. Micromanagers of gene expression: The potentially widespread influence of metazoan microRNAs. Nat. Rev. Genet. 2004, 5, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S. Role of microRNAs in insect host–microorganism interactions. Front. Physiol. 2011, 2, 48. [Google Scholar] [CrossRef] [PubMed]
- Skalsky, R.L.; Cullen, B.R. Viruses, microRNAs, and host interactions. Annu. Rev. Microbiol. 2010, 64, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Ørom, U.A.; Nielsen, F.C.; Lund, A.H. MicroRNA-10a binds the 5′UTR of ribosomal protein mRNAs and enhances their translation. Mol. Cell 2008, 30, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Liu, X.; Li, D.; Wang, P.; Li, N.; Lu, L.; Cao, X. MicroRNA-466l upregulates IL-10 expression in TLR-triggered macrophages by antagonizing RNA-binding protein tristetraprolin-mediated IL-10 mRNA degradation. J. Immunol. 2010, 184, 6053–6059. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Frentiu, F.D.; Moreira, L.A.; O’Neill, S.L.; Asgari, S. Wolbachia uses host microRNAs to manipulate host gene expression and facilitate colonization of the dengue vector Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 9250–9255. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.; Asgari, S. Functional analysis of a cellular microRNA in insect host-ascovirus interaction. J. Virol. 2010, 84, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Z.; Weber, F.; Croce, C.; Liu, C.G.; Liao, X.; Pellett, P.E. Human cytomegalovirus infection alters the expression of cellular microRNA species that affect its replication. J. Virol. 2008, 82, 9065–9074. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, I.M.; Cheng, G.; Wieland, S.; Volinia, S.; Croce, C.M.; Chisari, F.V.; David, M. Interferon modulation of cellular microRNAs as an antiviral mechanism. Nature 2007, 449, 919–922. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Xu, D.; Zhang, X. Characterization of host microRNAs that respond to DNA virus infection in a crustacean. BMC Genet. 2012, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhang, X. Functional analysis of a crustacean microRNA in host-virus interactions. J. Virol. 2012, 86, 12997–13004. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, X. Comprehensive characterization of viral miRNAs involved in white spot syndrome virus (WSSV) infection. RNA Biol. 2012, 9, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Cui, Y.; Zhang, X. Involvement of viral microRNA in the regulation of antiviral apoptosis in shrimp. J. Virol. 2014, 88, 2544–2554. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, K.; Zhang, X. Viral microRNAs targeting virus genes promote virus infection in shrimp in vivo. J. Virol. 2014, 88, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Escobedo-Bonilla, C.M.; Alday-Sanz, V.; Wille, M.; Sorgeloos, P.; Pensaert, M.B.; Nauwynck, H.J. A review on the morphology, molecular characterization, morphogenesis and pathogenesis of white spot syndrome virus. J. Fish Dis. 2008, 31, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Mercer, J.; Schelhaas, M.; Helenius, A. Virus entry by endocytosis. Annu. Rev. Biochem. 2010, 79, 803–833. [Google Scholar] [CrossRef] [PubMed]
- Sodeik, B. Mechanisms of viral transport in the cytoplasm. Trends Microbiol. 2000, 8, 465–472. [Google Scholar] [CrossRef]
- Marsh, M.; Bron, R. Sfv infection in CHO cells: Cell-type specific restrictions to productive virus entry at the cell surface. J. Cell Sci. 1997, 110, 95–103. [Google Scholar] [PubMed]
- Kalia, M.; Jameel, S. Virus entry paradigms. Amino Acids 2011, 41, 1147–1157. [Google Scholar] [PubMed]
- Van Hulten, M.C.W.; Witteveldt, J.; Snippe, M.; Vlak, J.M. White spot syndrome virus envelope protein VP28 is involved in the systemic infection of shrimp. Virology 2001, 285, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Yuan, J.F.; Cai, C.A.; Gu, W.G.; Shi, Z.L. Multiple envelope proteins are involved in white spot syndrome virus (WSSV) infection in crayfish. Arch. Virol. 2006, 151, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.; Wang, Z.; Qi, Y.; Yao, L.; Qian, J.; Hu, L. Vp28 of shrimp white spot syndrome virus is involved in the attachment and penetration into shrimp cells. J. Biochem. Mol. Biol. 2004, 37, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.; Xu, L.; Yang, F. Vp26 of white spot syndrome virus functions as a linker protein between the envelope and nucleocapsid of virions by binding with VP51. J. Virol. 2008, 82, 12598–12601. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.L.; Lu, L.C.; Wu, W.J.; Lo, C.F.; Huang, W.P. White spot syndrome virus envelope protein VP53A interacts with Penaeus monodon chitin-binding protein (PmCBP). Dis. Aquat. Org. 2007, 74, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.Y.; Hsu, T.C.; Huang, P.Y.; Kang, S.T.; Lo, C.F.; Huang, W.P.; Chen, L.L. Penaeus monodon chitinbinding protein (PMCBP) is involved in white spot syndrome virus (WSSV) infection. Fish Shellfish Immunol. 2009, 27, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-T.; Leu, J.-H.; Huang, P.-Y.; Chen, L.-L. A putative cell surface receptor for white spot syndrome virus is a member of a transporter superfamily. PLoS ONE 2012, 7, e33216. [Google Scholar] [CrossRef] [PubMed]
- Watthanasurorot, A.; Jiravanichpaisal, P.; Soderhall, I.; Soderhall, K. A gC1qR prevents white spot syndrome virus replication in the freshwater crayfish Pacifastacus leniusculus. J. Virol. 2010, 84, 10844–10851. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Y.; Yin, Z.X.; Xu, X.P.; Weng, S.P.; Rao, X.Y.; Dai, Z.X.; Luo, Y.; Yang, G.; Li, Z.H.; Guan, H.J.; et al. A novel C-type lectin from the shrimp Litopenaeus vannamei possesses anti-white spot syndrome virus activity. J. Virol. 2009, 83, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, W.T.; Zhang, X.W.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. A c-type lectin is involved in the innate immune response of Chinese white shrimp. Fish Shellfish Immunol. 2009, 27, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Song, K.K.; Li, D.F.; Zhang, M.C.; Yang, H.J.; Ruan, L.W.; Xu, X. Cloning and characterization of three novel WSSV recognizing lectins from shrimp Marsupenaeus japonicus. Fish Shellfish Immunol. 2010, 28, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Xu, Y.H.; Xu, J.D.; Zhao, X.F.; Wang, J.X. Collaboration between a soluble C-type lectin and calreticulin facilitates white spot syndrome virus infection in shrimp. J. Immunol. 2014, 193, 2106–2117. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Bi, W.J.; Wang, X.W.; Zhao, Y.R.; Zhao, X.F.; Wang, J.X. Two novel C-type lectins with a low-density lipoprotein receptor class a domain have antiviral function in the shrimp Marsupenaeus japonicus. Dev. Comp. Immunol. 2014, 42, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Li, D.F.; Zhang, M.C.; Yang, H.J.; Zhu, Y.B.; Xu, X. B-integrin mediates WSSV infection. Virology 2007, 368, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Liu, Q.H.; Huang, J. Multiple proteins of white spot syndrome virus involved in recognition of β-integrin. J. Biosci. 2014, 39, 1–8. [Google Scholar] [CrossRef]
- Sun, Z.; Li, S.; Li, F.; Xiang, J. Bioinformatic prediction of WSSV-host protein-protein interaction. BioMed Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Watthanasurorot, A.; Guo, E.; Tharntada, S.; Lo, C.F.; Soderhall, K.; Soderhall, I. Hijacking of host calreticulin is required for the white spot syndrome virus replication cycle. J. Virol. 2014, 88, 8116–8128. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zong, R.; Xu, J.; Zhang, X. Antiviral phagocytosis is regulated by a novel Rab-dependent complex in shrimp Penaeus japonicus. J. Proteom. Res. 2007, 7, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Sritunyalucksana, K.; Wannapapho, W.; Lo, C.F.; Flegel, T.W. PMRAB7 is a VP28-binding protein involved in white spot syndrome virus infection in shrimp. J. Virol. 2006, 80, 10734–10742. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Miranda, J.S.; Lopez-Zavala, A.A.; Arvizu-Flores, A.A.; Garcia-Orozco, K.D.; Stojanoff, V.; Rudiño-Piñera, E.; Brieba, L.G.; Sotelo-Mundo, R.R. Crystal structure of the shrimp proliferating cell nuclear antigen: Structural complementarity with WSSV DNA polymerase pip-box. PLoS ONE 2014, 9, e94369. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Lee, D.Y.; Wang, H.C.; Kang, S.T.; Hwang, P.P.; Kou, G.H.; Huang, M.F.; Chang, G.D.; Lo, C.F. White spot syndrome virus protein kinase 1 defeats the host cell’s iron-withholding defense mechanism by interacting with host ferritin. J. Virol. 2015, 89, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Ruan, L.; Xu, X. An immediate-early protein of white spot syndrome virus modulates the phosphorylation of focal adhesion kinase of shrimp. Virology 2011, 419, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.H.; Chen, L.L.; Lin, Y.R.; Kou, G.H.; Lo, C.F. Molecular mechanism of the interactions between white spot syndrome virus anti-apoptosis protein AAP-1 (WSSV449) and shrimp effector caspase. Dev. Comp. Immunol. 2010, 34, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Lertwimol, T.; Sangsuriya, P.; Phiwsaiya, K.; Senapin, S.; Phongdara, A.; Boonchird, C.; Flegel, T.W. Two new anti-apoptotic proteins of white spot syndrome virus that bind to an effector caspase (PmCasp) of the giant tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 2014, 38, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chua, H.K.; Gusti, A.A.; He, F.; Fenner, B.; Manopo, I.; Wang, H.; Kwang, J. RING-H2 protein WSSV249 from white spot syndrome virus sequesters a shrimp ubiquitin-conjugating enzyme, PvUbc, for viral pathogenesis. J. Virol. 2005, 79, 8764–8772. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Wang, H.C.; Ko, T.P.; Lee, Y.M.; Leu, J.H.; Ho, C.H.; Huang, W.P.; Lo, C.F.; Wang, A.H. White spot syndrome virus protein ICP11: A histone-binding DNA mimic that disrupts nucleosome assembly. Proc. Natl. Acad. Sci. USA 2008, 105, 20758–20763. [Google Scholar] [CrossRef] [PubMed]
- Tonganunt, M.; Saelee, N.; Chotigeat, W.; Phongdara, A. Identification of a receptor for activated protein kinase C1 (Pm-RACK1), a cellular gene product from black tiger shrimp (Penaeus monodon) interacts with a protein, VP9 from the white spot syndrome virus. Fish Shellfish Immunol. 2009, 26, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Sangsuriya, P.; Senapin, S.; Huang, W.-P.; Lo, C.-F.; Flegel, T.W. Co-interactive DNA-binding between a novel, immunophilin-like shrimp protein and VP15 nucleocapsid protein of white spot syndrome virus. PLoS ONE 2011, 6, e25420. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.F.; Liu, Q.H.; Guan, G.K.; Li, C.; Huang, J. Arginine kinase of Litopenaeus vannamei involved in white spot syndrome virus infection. Gene 2014, 539, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Yang, F. Interaction of white spot syndrome virus VP26 protein with actin. Virology 2005, 336, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Kwang, J. Identification of a novel shrimp protein phosphatase and its association with latency-related ORF427 of white spot syndrome virus. FEBS Lett. 2004, 577, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chang, Y.S.; Huang, W.T.; Chen, I.T.; Wang, K.C.; Kou, G.H.; Lo, C.F. Penaeus monodon tata box-binding protein interacts with the white spot syndrome virus transactivator ie1 and promotes its transcriptional activity. J. Virol. 2011, 85, 6535–6547. [Google Scholar] [CrossRef] [PubMed]
- Ran, X.; Bian, X.; Ji, Y.; Yan, X.; Yang, F.; Li, F. White spot syndrome virus ie1 and wsv056 modulate the G1/S transition by binding to the host retinoblastoma protein. J. Virol. 2013, 87, 12576–12582. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-S.; Liu, W.-J.; Lee, C.-C.; Chou, T.-L.; Lee, Y.-T.; Wu, T.-S.; Huang, J.-Y.; Huang, W.-T.; Lee, T.-L.; Kou, G.-H.; et al. A 3D model of the membrane protein complex formed by the white spot syndrome virus structural proteins. PLoS ONE 2010, 5, e10718. [Google Scholar] [CrossRef] [PubMed]
- Spear, P.G.; Longnecker, R. Herpesvirus entry: An update. J. Virol. 2003, 77, 10179–10185. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.P.; Tarr, A.W. Human lectins and their roles in viral infections. Molecules 2015, 20, 2229–2271. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.-J.; Kang, S.-T.; Leu, J.-H.; Chen, L.-L. Endocytic pathway is indicated for white spot syndrome virus (WSSV) entry in shrimp. Fish Shellfish Immunol. 2013, 35, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.; Jin, S.; Zhang, Y.; Li, F.; Xiang, J. Granulocytes of the red claw crayfish Cherax quadricarinatus can endocytose beads, E. coli and wssv, but in different ways. Dev. Comp. Immunol. 2014, 46, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Li, F.; Wu, J.; Yang, F. White spot syndrome virus enters crayfish hematopoietic tissue cells via Clathrin-mediated endocytosis. Virology 2015, 486, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Mayor, S.; Presley, J.F.; Maxfield, F.R. Sorting of membrane components from endosomes and subsequent recycling to the cell surface occurs by a bulk flow process. J. Cell Biol. 1993, 121, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Lozach, P.-Y.; Huotari, J.; Helenius, A. Late-penetrating viruses. Curr. Opin. Virol. 2011, 1, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, L.; Wang, S.; Wang, T. Rab7: Roles in membrane trafficking and disease. Biosci. Rep. 2009, 29, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Kobiler, O.; Drayman, N.; Butin-Israeli, V.; Oppenheim, A. Virus strategies for passing the nuclear envelope barrier. Nucleus 2012, 3, 526–539. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed]
- Del Conte-Zerial, P.; Brusch, L.; Rink, J.C.; Collinet, C.; Kalaidzidis, Y.; Zerial, M.; Deutsch, A. Membrane identity and GTPase cascades regulated by toggle and cut-out switches. Mol. Syst. Biol. 2008, 4, 206. [Google Scholar] [CrossRef] [PubMed]
- Píndaro, Á.-R.; Humberto, M.-R.C.; Javier, M.-B.F.; Marcial, E.-B.C. Silencing pacific white shrimp Litopenaeus vannamei LVRAB7 reduces mortality in brooders challenged with white spot syndrome virus. Aquacult. Res. 2013, 44, 772–782. [Google Scholar] [CrossRef]
- Attasart, P.; Kaewkhaw, R.; Chimwai, C.; Kongphom, U.; Namramoon, O.; Panyim, S. Inhibition of white spot syndrome virus replication in Penaeus monodon by combined silencing of viral rr2 and shrimp PmRab7. Virus Res. 2009, 145, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ongvarrasopone, C.; Chanasakulniyom, M.; Sritunyalucksana, K.; Panyim, S. Suppression of PmRab7 by dsRNA inhibits WSSV or YHV infection in shrimp. Ma. Biotechnol. 2008, 10, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; He, N.; Yang, Z.; Liu, H.; Xu, X. Differential gene expression profile in hepatopancreas of WSSV-resistant shrimp (Penaeus japonicus) by suppression subtractive hybridization. Dev. Comp. Immunol. 2005, 29, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.; Splinter, D.; Keijzer, N.; Wulf, P.S.; Demmers, J.; Ohtsuka, T.; Modesti, M.; Maly, I.V.; Grosveld, F.; Hoogenraad, C.C.; et al. Rab6 regulates transport and targeting of exocytotic carriers. Dev. Cell 2007, 13, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chang, Y.S.; Wang, A.H.; Kou, G.H.; Lo, C.F. White spot syndrome virus annexes a shrimp stat to enhance expression of the immediate-early gene ie1. J. Virol. 2007, 81, 1461–1471. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Yu, L.; Wang, Q.; Liu, W.; Cui, Y.; Kwang, J. Sf-PHB2, a new transcription factor, drives WSSV ie1 gene expression via a 12-bp DNA element. Virol. J. 2012, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Zhang, S.; Chen, Y.G.; Wang, P.H.; Xu, X.P.; Li, C.Z.; Chen, Y.H.; Fan, W.Z.; Yan, H.; Weng, S.P.; et al. Litopenaeus vannamei NF-κB is required for WSSV replication. Dev. Comp. Immunol. 2014, 45, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.D.; Zhao, L.; Zhang, H.Q.; Xu, X.P.; Jia, X.T.; Chen, Y.H.; Wang, P.H.; Weng, S.P.; Yu, X.Q.; Yin, Z.X.; et al. Shrimp NF-κB binds to the immediate-early gene ie1 promoter of white spot syndrome virus and upregulates its activity. Virology 2010, 406, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.H.; Gu, Z.H.; Wan, D.H.; Zhang, M.Y.; Weng, S.P.; Yu, X.Q.; He, J.G. The shrimp NF-κB pathway is activated by white spot syndrome virus (WSSV) 449 to facilitate the expression of WSSV069 (ie1), WSSV303 and WSSV371. PLoS ONE 2011, 6, e24773. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yan, X.; Ruan, L.; Xu, X. A novel jnk from Litopenaeus vannamei involved in white spot syndrome virus infection. Dev. Comp. Immunol. 2012, 37, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Pang, L.R.; Chen, Y.G.; Weng, S.P.; Yue, H.T.; Zhang, Z.Z.; Chen, Y.H.; He, J.G. Activating transcription factor 4 and x box binding protein 1 of Litopenaeus vannamei transcriptional regulated white spot syndrome virus genes wsv023 and wsv083. PLoS ONE 2013, 8, e62603. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Lo, C.F.; Kou, G.H.; Leu, J.H.; Lai, Y.J.; Chang, L.K.; Chang, Y.S. The promoter of the white spot syndrome virus immediate-early gene wssv108 is activated by the cellular KLF transcription factor. Dev. Comp. Immunol. 2015, 49, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Yue, H.T.; Zhang, Z.Z.; Bi, H.T.; Chen, Y.G.; Weng, S.P.; Chan, S.; He, J.G.; Chen, Y.H. An activating transcription factor of Litopenaeus vannamei involved in wssv genes wsv059 and wsv166 regulation. Fish Shellfish Immunol. 2014, 41, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.; Chen, C.; Gao, Y.; Lin, J.; Jin, C.; Wang, W. Regulation of shrimp PjCaspase promoter activity by WSSV VP38 and VP41B. Fish Shellfish Immunol. 2011, 30, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, M.; Ke, W.; Ji, Y.; Bian, X.; Yan, X. Identification of the immediate-early genes of white spot syndrome virus. Virology 2009, 385, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Chang, Y.S.; Wang, C.H.; Kou, G.H.; Lo, C.F. Microarray and RT-PCR screening for white spot syndrome virus immediate-early genes in cycloheximide-treated shrimp. Virology 2005, 334, 327–341. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Ho, Y.; Yu, L.; Kwang, J. WSSV ie1 promoter is more efficient than CMV promoter to express H5 hemagglutinin from influenza virus in baculovirus as a chicken vaccine. BMC Microbiol. 2008, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, H.; Wang, Y.; Li, N.; Peng, W.-P.; Sun, Y.; Tong, G.-Z.; Qiu, H.-J. Efficient gene delivery into mammalian cells mediated by a recombinant baculovirus containing a whispovirus ie1 promoter, a novel shuttle promoter between insect cells and mammalian cells. J. Biotechnol. 2007, 131, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Li, C.; Su, Z.; Liang, Q.; Li, H.; Liang, S.; Weng, S.; He, J.; Xu, X. Identification of a jak/stat pathway receptor domeless from pacific white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2015, 44, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Vallabhapurapu, S.; Karin, M. Regulation and function of NF-κB transcription factors in the immune system. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef] [PubMed]
- Matys, V.; Kel-Margoulis, O.V.; Fricke, E.; Liebich, I.; Land, S.; Barre-Dirrie, A.; Reuter, I.; Chekmenev, D.; Krull, M.; Hornischer, K.; et al. Transfac® and its module transcompel®: Transcriptional gene regulation in eukaryotes. Nucleic Acids Res. 2006, 34, D108–D110. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.J.; Gao, L.; Wang, X.W.; Zhao, X.F.; Wang, J.X. SUMO-conjugating enzyme E2 UBC9 mediates viral immediate-early protein sumoylation in crayfish to facilitate reproduction of white spot syndrome virus. J. Virol. 2013, 87, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Flemington, E.K. Herpesvirus lytic replication and the cell cycle: Arresting new developments. J. Virol. 2001, 75, 4475–4481. [Google Scholar] [CrossRef] [PubMed]
- Kudoh, A.; Fujita, M.; Kiyono, T.; Kuzushima, K.; Sugaya, Y.; Izuta, S.; Nishiyama, Y.; Tsurumi, T. Reactivation of lytic replication from b cells latently infected with Epstein-Barr virus occurs with high s-phase cyclin-dependent kinase activity while inhibiting cellular DNA replication. J. Virol. 2003, 77, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-L.; Wang, H.-C.; Huang, C.-J.; Peng, S.-E.; Chen, Y.-G.; Lin, S.-J.; Chen, W.-Y.; Dai, C.-F.; Yu, H.-T.; Wang, C.-H.; et al. Transcriptional analysis of the DNA polymerase gene of shrimp white spot syndrome virus. Virology 2002, 301, 136–147. [Google Scholar] [CrossRef] [PubMed]
- De-la-Re-Vega, E.; Garcia-Orozco, K.D.; Arvizu-Flores, A.A.; Yepiz-Plascencia, G.; Muhlia-Almazan, A.; Hernández, J.; Brieba, L.G.; Sotelo-Mundo, R.R. White spot syndrome virus ORF514 encodes a bona fide DNA polymerase. Molecules 2011, 16, 532–542. [Google Scholar] [CrossRef] [PubMed]
- De-la-Re-Vega, E.; Muhlia-Almazan, A.; Arvizu-Flores, A.A.; Islas-Osuna, M.A.; Yepiz-Plascencia, G.; Brieba, L.G.; Sotelo-Mundo, R.R. Molecular modeling and expression of the Litopenaeus vannamei proliferating cell nuclear antigen (PCNA) after white spot syndrome virus shrimp infection. Res. Immunol. 2011, 1, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Lunt, S.Y.; van der Heiden, M.G. Aerobic glycolysis: Meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 2011, 27, 441–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warburg, O. On the origin of cancer cells. Science 1956, 123, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Su, M.A.; Huang, Y.T.; Chen, I.T.; Lee, D.Y.; Hsieh, Y.C.; Li, C.Y.; Ng, T.H.; Liang, S.Y.; Lin, S.Y.; Huang, S.W.; et al. An invertebrate Warburg effect: A shrimp virus achieves successful replication by altering the host metabolome via the pi3k-akt-mtor pathway. PLoS Pathog. 2014, 10, e1004196. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.T.; Aoki, T.; Huang, Y.T.; Hirono, I.; Chen, T.C.; Huang, J.Y.; Chang, G.D.; Lo, C.F.; Wang, H.C. White spot syndrome virus induces metabolic changes resembling the Warburg effect in shrimp hemocytes in the early stage of infection. J. Virol. 2011, 85, 12919–12928. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.B.; Hay, N. Is Akt the “warburg kinase”?—Akt-energy metabolism interactions and oncogenesis. Semin. Cancer Biol. 2009, 19, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Spangle, J.M.; Munger, K. The human papillomavirus type 16 E6 oncoprotein activates mtorc1 signaling and increases protein synthesis. J. Virol. 2010, 84, 9398–9407. [Google Scholar] [CrossRef]
- Ye, T.; Wu, X.; Wu, W.; Dai, C.; Yuan, J. Ferritin protect shrimp Litopenaeus vannamei from WSSV infection by inhibiting virus replication. Fish Shellfish Immunol. 2015, 42, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kosuke, Z.; Nam, Y.K.; Kim, S.K.; Kim, K.H. Protection of shrimp (Penaeus chinensis) against white spot syndrome virus (WSSV) challenge by double-stranded RNA. Fish Shellfish Immunol. 2007, 23, 242–246. [Google Scholar] [CrossRef]
- Fung, T.S.; Liu, D.X. Coronavirus infection, ER stress, apoptosis and innate immunity. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A. A new paradigm: Innate immune sensing of viruses via the unfolded protein response. Front. Microbiol. 2014, 5, 222. [Google Scholar] [CrossRef] [PubMed]
- He, B. Viruses, endoplasmic reticulum stress, and interferon responses. Cell Death Differ. 2006, 13, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.M.; Raghuwanshi, S.K.; Rowe, D.T.; Wadowsky, R.M.; Rosendorff, A. Endoplasmic reticulum stress causes EBV lytic replication. Blood 2011, 118, 5528–5539. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Alonso, V.; Maruri-Avidal, L.; Arias, C.F.; López, S. Rotavirus infection induces the unfolded protein response of the cell and controls it through the nonstructural protein NSP3. J. Virol. 2011, 85, 12594–12604. [Google Scholar] [CrossRef] [PubMed]
- Burnett, H.F.; Audas, T.E.; Liang, G.; Lu, R.R. Herpes simplex virus-1 disarms the unfolded protein response in the early stages of infection. Cell Stress Chaperones 2012, 17, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Zhao, L.; Pang, L.R.; Li, X.Y.; Weng, S.P.; He, J.G. Identification and characterization of inositol-requiring enzyme-1 and x-box binding protein 1, two proteins involved in the unfolded protein response of Litopenaeus vannamei. Dev. Comp. Immunol. 2012, 38, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Luana, W.; Li, F.; Wang, B.; Zhang, X.; Liu, Y.; Xiang, J. Molecular characteristics and expression analysis of calreticulin in Chinese shrimp Fenneropenaeus chinensis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Visudtiphole, V.; Watthanasurorot, A.; Klinbunga, S.; Menasveta, P.; Kirtikara, K. Molecular characterization of calreticulin: A biomarker for temperature stress responses of the giant tiger shrimp Penaeus monodon. Aquaculture 2010, 308, S100–S108. [Google Scholar] [CrossRef]
- Luan, W.; Li, F.; Zhang, J.; Wang, B.; Xiang, J. Cloning and expression of glucose regulated protein 78 (GRP78) in Fenneropenaeus chinensis. Mol. Biol. Rep. 2009, 36, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-J.; Leu, J.-H.; Tsau, M.-T.; Chen, J.-C.; Chen, L.-L. Differential expression of LVHSP60 in shrimp in response to environmental stress. Fish Shellfish Immunol. 2011, 30, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-R.; Hung, H.-C.; Leu, J.-H.; Wang, H.-C.; Kou, G.-H.; Lo, C.-F. The role of aldehyde dehydrogenase and HSP70 in suppression of white spot syndrome virus replication at high temperature. J. Virol. 2011, 85, 3517–3525. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, N.; Gorman, A.; Gupta, S.; Samali, A. The EIF2α kinases: Their structures and functions. Cell. Mol. Life Sci. 2013, 70, 3493–3511. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ruan, L.; Shi, H. EIF2α of Litopenaeus vannamei involved in shrimp immune response to WSSV infection. Fish Shellfish Immunol. 2014, 40, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Koyama, A.H.; Fukumori, T.; Fujita, M.; Irie, H.; Adachi, A. Physiological significance of apoptosis in animal virus infection. Microbes Infect. 2000, 2, 1111–1117. [Google Scholar] [CrossRef]
- Best, S.M. Viral subversion of apoptotic enzymes: Escape from death row. Annu. Rev. Microbiol. 2008, 62, 171–192. [Google Scholar] [CrossRef] [PubMed]
- Henderson, T.; Stuck, K. Induction of apoptosis in response to white spot syndrome virus in the pacific white shrimp. In Penaeus Vannamei; Book of abstracts; Aquaculture America: Tampa, FL, USA, 1999; Abstract 67. [Google Scholar]
- Abeer, H.S.; Hassan, M.D.; Shariff, M. DNA fragmentation, an indicator of apoptosis, in cultured black tiger shrimp Penaeus monodon infected with white spot syndrome virus (WSSV). Dis. Aquat. Org. 2001, 44, 155–159. [Google Scholar]
- Kanokpan, W.; Kornnika, K.; Supatra Somapa, G.; Prasert, M.; Boonsirm, W. Time-course and levels of apoptosis in various tissues of black tiger shrimp Penaeus monodon infected with white-spot syndrome virus. Dis. Aquat. Org. 2003, 55, 3–10. [Google Scholar]
- Wu, J.L.; Muroga, K. Apoptosis does not play an important role in the resistance of “immune” Penaeus japonicus against white spot syndrome virus. J. Fish Dis. 2004, 27, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.H.; Lin, S.J.; Huang, J.Y.; Chen, T.C.; Lo, C.F. A model for apoptotic interaction between white spot syndrome virus and shrimp. Fish Shellfish Immunol. 2013, 34, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Mechanisms of caspase activation and inhibition during apoptosis. Mol. Cell 2002, 9, 459–470. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harbor Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhi, B.; Wu, W.; Zhang, X. Requirement for shrimp caspase in apoptosis against virus infection. Dev. Comp. Immunol. 2008, 32, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Xu, L.; Yang, F. Proteomic analysis of the major envelope and nucleocapsid proteins of white spot syndrome virus. J. Virol. 2006, 80, 10615–10623. [Google Scholar] [CrossRef] [PubMed]
- Jie, Z.; Xu, L.; Yang, F. The c-terminal region of envelope protein VP38 from white spot syndrome virus is indispensable for interaction with VP24. Arch. Virol. 2008, 153, 2103–2106. [Google Scholar] [CrossRef] [PubMed]
- Jesenberger, V.; Jentsch, S. Deadly encounter: Ubiquitin meets apoptosis. Nat. Rev. Mol. Cell Biol. 2002, 3, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Freemont, P.S. Ubiquitination: Ring for destruction? Current Biol. 2000, 10, R84–R87. [Google Scholar] [CrossRef]
- Chen, A.J.; Wang, S.; Zhao, X.F.; Yu, X.Q.; Wang, J.X. Enzyme E2 from Chinese white shrimp inhibits replication of white spot syndrome virus and ubiquitinates its ring domain proteins. J. Virol. 2011, 85, 8069–8079. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Fenner, B.J.; Godwin, A.K.; Kwang, J. White spot syndrome virus open reading frame 222 encodes a viral E3 ligase and mediates degradation of a host tumor suppressor via ubiquitination. J. Virol. 2006, 80, 3884–3892. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Wang, H.C.; Kou, G.C.; Lo, C.F.; Huang, W.P. Identification of ICP11, the most highly expressed gene of shrimp white spot syndrome virus (WSSV). Dis. Aquat. Org. 2007, 74, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, J.; Song, J.; Sivaraman, J.; Hew, C.L. Identification of a novel nonstructural protein, VP9, from white spot syndrome virus: Its structure reveals a ferredoxin fold with specific metal binding sites. J. Virol. 2006, 80, 10419–10427. [Google Scholar] [CrossRef] [PubMed]
- Boner, W.; Morgan, I.M. Novel cellular interacting partners of the human papillomavirus 16 transcription/replication factor E2. Virus Res. 2002, 90, 113–118. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, J.; Chen, H.; Hew, C.L.; Yan, J. DNA condensates organized by the capsid protein VP15 in white spot syndrome virus. Virology 2010, 408, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Witteveldt, J.; Vermeesch, A.M.G.; Langenhof, M.; de Lang, A.; Vlak, J.M.; van Hulten, M.C.W. Nucleocapsid protein VP15 is the basic DNA binding protein of white spot syndrome virus of shrimp. Arch. Virol. 2005, 150, 1121–1133. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.M.; Yao, Y.L.; Seto, E. The FK506-binding protein 25 functionally associates with histone deacetylases and with transcription factor YY1. EMBO J. 2001, 20, 4814–4825. [Google Scholar] [CrossRef] [PubMed]
- Kuzuhara, T.; Horikoshi, M. A nuclear FK506-binding protein is a histone chaperone regulating rDNA silencing. Nat. Struct. Mol. Biol. 2004, 11, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Uchiyama, S.; Sone, T.; Noda, M.; Lin, L.; Mizuno, H.; Matsunaga, S.; Fukui, K. Calreticulin as a new histone binding protein in mitotic chromosomes. Cytogenet. Genome Res. 2006, 115, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Holaska, J.M.; Black, B.E.; Love, D.C.; Hanover, J.A.; Leszyk, J.; Paschal, B.M. Calreticulin is a receptor for nuclear export. J. Cell Biol. 2001, 152, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.C.; Chen, Y.M.; Li, C.Y.; Chang, Y.H.; Liang, S.Y.; Lin, S.Y.; Lin, C.Y.; Chang, S.H.; Wang, Y.J.; Khoo, K.H.; et al. To complete its replication cycle, a shrimp virus changes the population of long chain fatty acids during infection via the PI3K-Akt-mTOR-HIF1α pathway. Dev. Comp. Immunol. 2015, 53, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Itami, T.; Asano, M.; Tokushige, K.; Kubono, K.; Nakagawa, A.; Takeno, N.; Nishimura, H.; Maeda, M.; Kondo, M.; Takahashi, Y. Enhancement of disease resistance of kuruma shrimp, Penaeus japonicus, after oral administration of peptidoglycan derived from Bifidobacterium thermophilum. Aquaculture 1998, 164, 277–288. [Google Scholar] [CrossRef]
- Thitamadee, S.; Srisala, J.; Taengchaiyaphum, S.; Sritunyalucksana, K. Double-dose β-glucan treatment in WSSV-challenged shrimp reduces viral replication but causes mortality possibly due to excessive Ros production. Fish Shellfish Immunol. 2014, 40, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Syed Musthaq, S.K.; Kwang, J. Reprint of “evolution of specific immunity in shrimp—A vaccination perspective against white spot syndrome virus”. Dev. Comp. Immunol. 2015, 48, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.S.B.; Manjusha, M.; Pai, S.S.; Rosamma, P. Fenneropenaeus indicus is protected from white spot disease by oral administration of inactivated white spot syndrome virus. Dis. Aquat. Org. 2005, 66, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Syed Musthaq, S.; Madhan, S.; Sahul Hameed, A.S.; Kwang, J. Localization of VP28 on the baculovirus envelope and its immunogenicity against white spot syndrome virus in Penaeus monodon. Virology 2009, 391, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, A.T.; Pham, C.K.; Pham, H.T.; Pham, H.L.; Nguyen, A.H.; Dang, L.T.; Huynh, H.A.; Cutting, S.M.; Phan, T.N. Bacillus subtilis spores expressing the VP28 antigen: A potential oral treatment to protect Litopenaeus vannamei against white spot syndrome. FEMS Microbiol. Lett. 2014, 358, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zhang, X. Host defense against DNA virus infection in shrimp is mediated by the siRNA pathway. European J. Immunol. 2013, 43, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, X. Protection of shrimp against white spot syndrome virus (WSSV) with beta-1,3-d-glucan-encapsulated VP28-sirna particles. Ma. Biotechnol. 2012, 14, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, G.; Sarathi, M.; Venkatesan, C.; Thomas, J.; Sahul Hameed, A.S. Oral administration of antiviral plant extract of Cynodon dactylon on a large scale production against white spot syndrome virus (WSSV) in Penaeus monodon. Aquaculture 2008, 279, 2–5. [Google Scholar] [CrossRef]
- Sudheer, N.S.; Philip, R.; Singh, I.S.B. In vivo screening of mangrove plants for anti WSSV activity in Penaeus monodon, and evaluation of ceriops tagal as a potential source of antiviral molecules. Aquaculture 2011, 311, 36–41. [Google Scholar] [CrossRef]
- Balasubramanian, G.; Sudhakaran, R.; Syed Musthaq, S.; Sarathi, M.; Sahul Hameed, A.S. Studies on the inactivation of white spot syndrome virus of shrimp by physical and chemical treatments, and seaweed extracts tested in marine and freshwater animal models. J. Fish Dis. 2006, 29, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, U.; Chakraborty, S.; Balasubramanian, T.; Das, P. Screening, isolation and optimization of anti–white spot syndrome virus drug derived from terrestrial plants. Asia-Pac. J. Trop. Biomed. 2014, 4, S118–S128. [Google Scholar] [CrossRef] [PubMed]
- Bindhu, F.; Velmurugan, S.; Donio, M.B.S.; Michaelbabu, M.; Citarasu, T. Influence of agathi grandiflora active principles inhibit viral multiplication and stimulate immune system in Indian white shrimp Fenneropenaeus indicus against white spot syndrome virus infection. Fish Shellfish Immunol. 2014, 41, 482–492. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verbruggen, B.; Bickley, L.K.; Van Aerle, R.; Bateman, K.S.; Stentiford, G.D.; Santos, E.M.; Tyler, C.R. Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments. Viruses 2016, 8, 23. https://doi.org/10.3390/v8010023
Verbruggen B, Bickley LK, Van Aerle R, Bateman KS, Stentiford GD, Santos EM, Tyler CR. Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments. Viruses. 2016; 8(1):23. https://doi.org/10.3390/v8010023
Chicago/Turabian StyleVerbruggen, Bas, Lisa K. Bickley, Ronny Van Aerle, Kelly S. Bateman, Grant D. Stentiford, Eduarda M. Santos, and Charles R. Tyler. 2016. "Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments" Viruses 8, no. 1: 23. https://doi.org/10.3390/v8010023
APA StyleVerbruggen, B., Bickley, L. K., Van Aerle, R., Bateman, K. S., Stentiford, G. D., Santos, E. M., & Tyler, C. R. (2016). Molecular Mechanisms of White Spot Syndrome Virus Infection and Perspectives on Treatments. Viruses, 8(1), 23. https://doi.org/10.3390/v8010023