
Maporal Hantavirus Causes Mild Pathology in Deer Mice (Peromyscus maniculatus)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Infections
2.3. Tissue Processing
2.4. ELISA
2.5. Serum Neutralization Assay
2.6. Detection of Viral RNA in Tissues
2.7. Cytokine PCR
2.8. Generation of Deer Mouse Pulmonary Microvascular Endothelial Cells
2.9. PMVEC Infections
3. Results
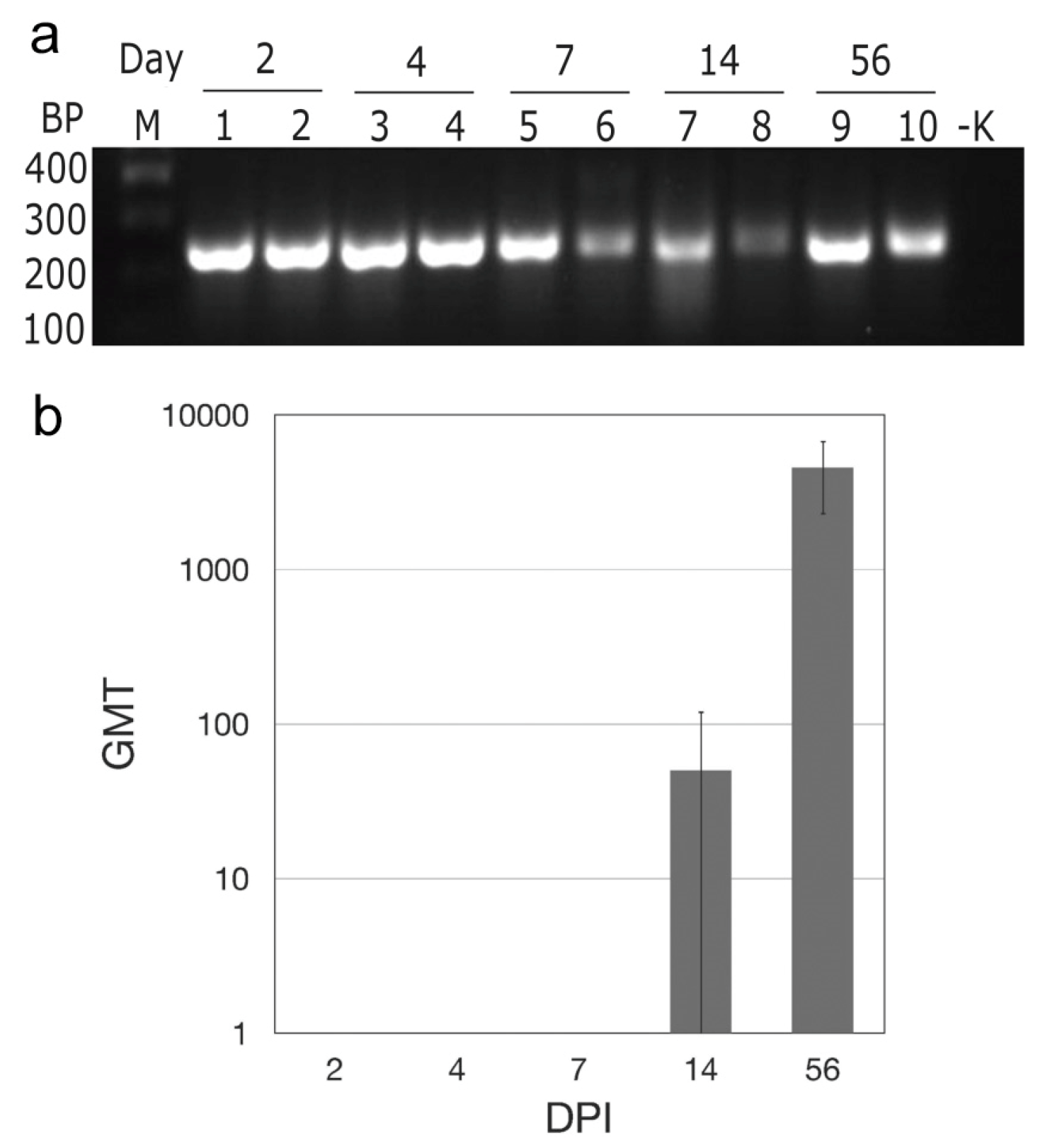
3.1. Maporal Virus Persistently Infects Deer Mice
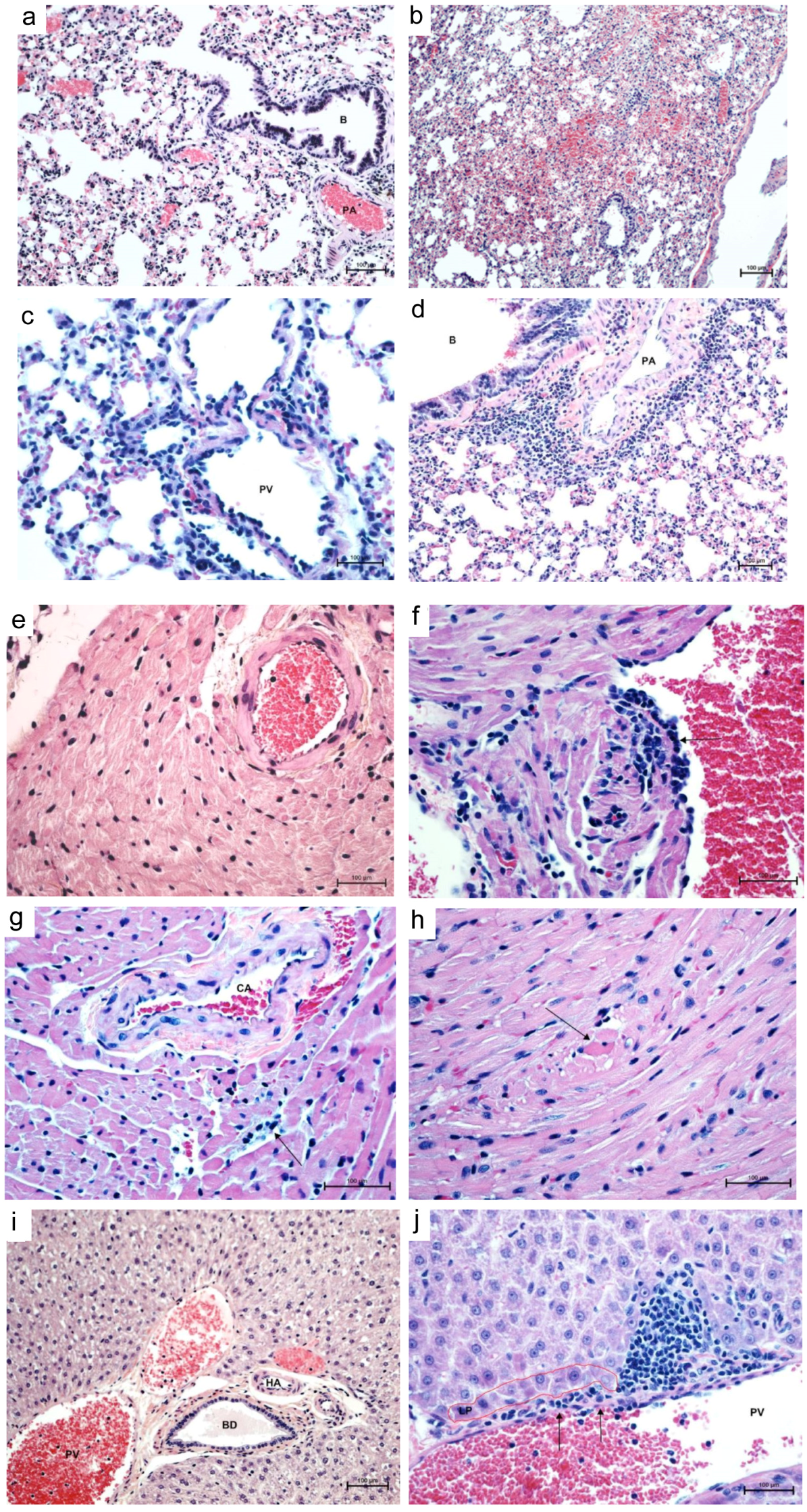
3.2. Histopathology and Immunohistochemistry
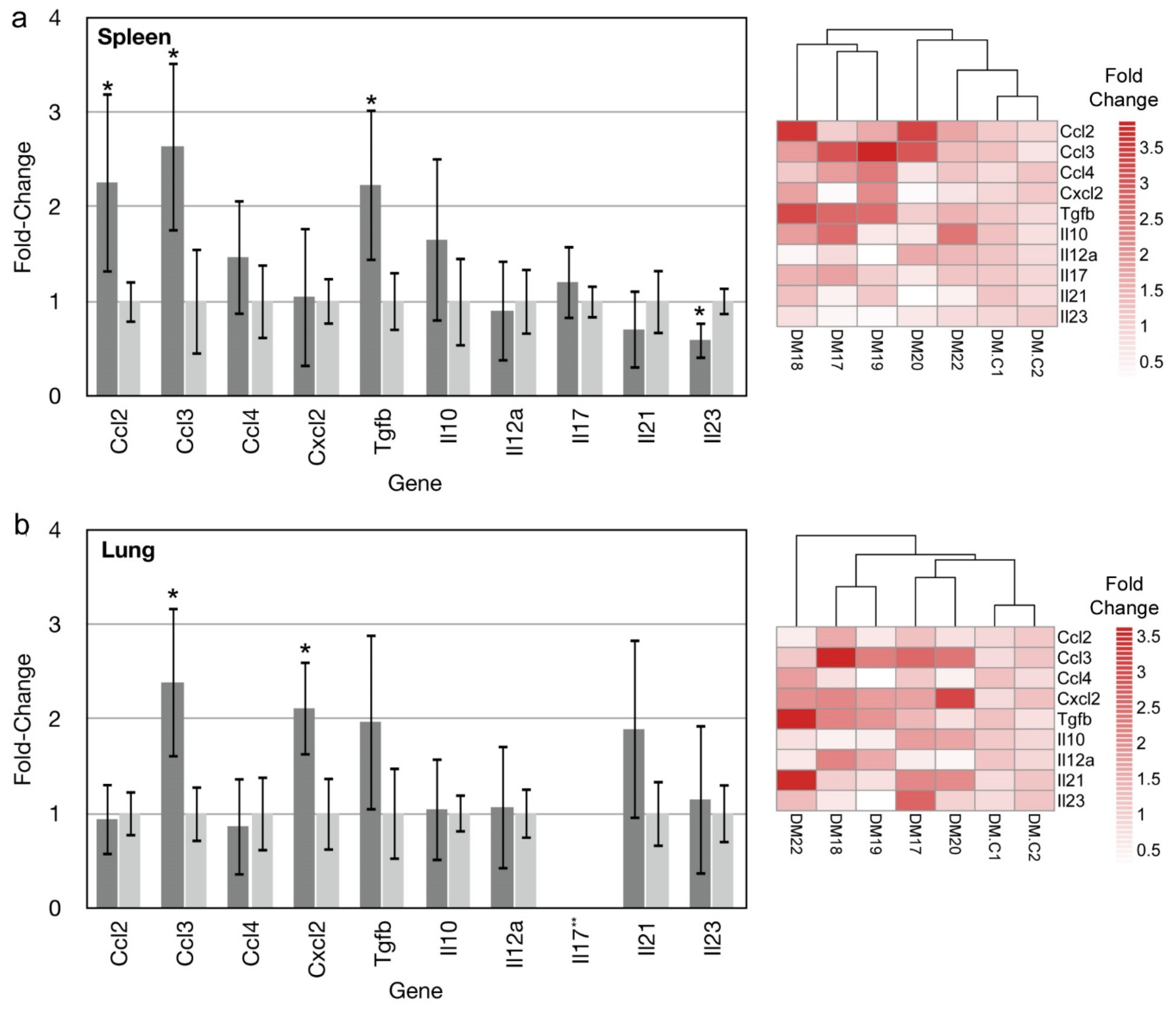
3.3. Gene Expression Profiling
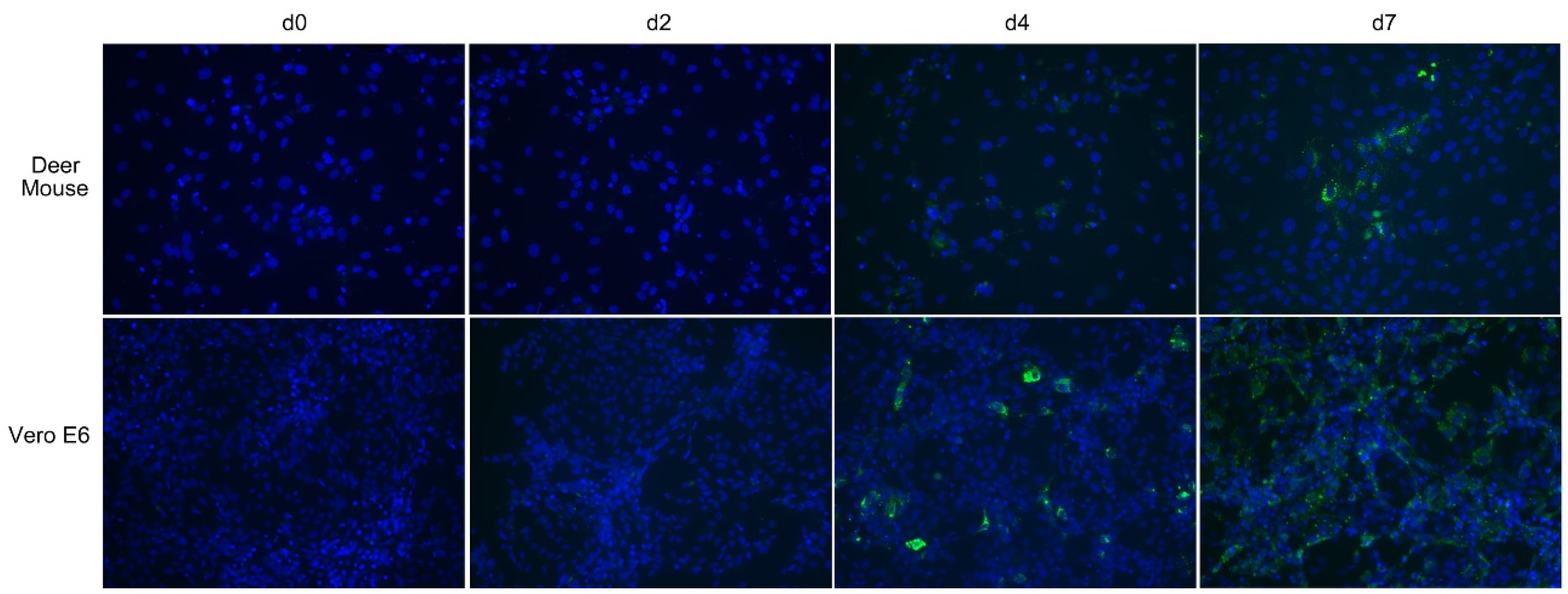
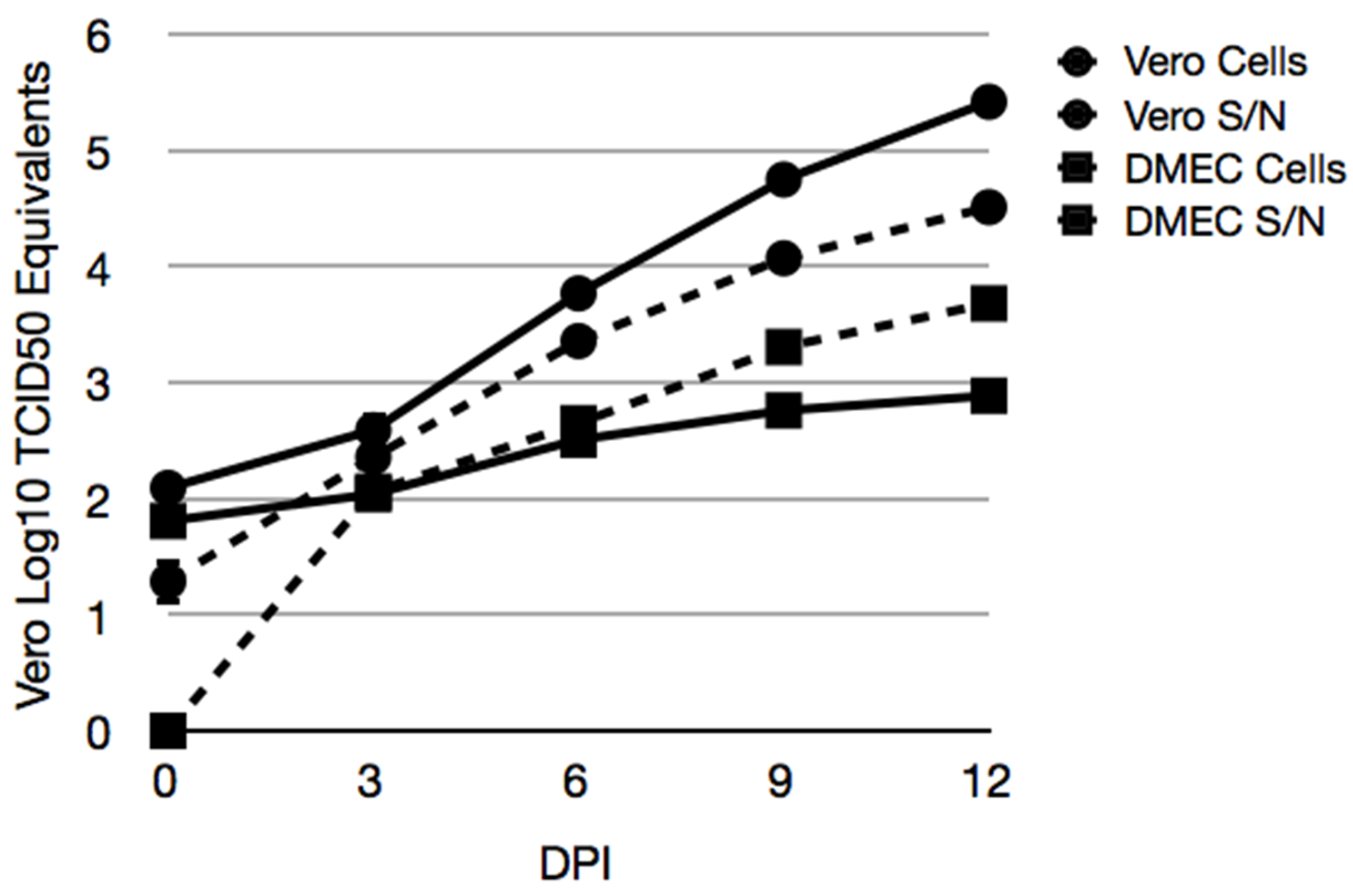
3.4. Maporal Virus Infects Deer Mouse Pulmonary Cells
3.5. Maporal Virus RNA Accumulates in the Cellular Fraction of Deer Mouse PMVEC
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schmaljohn, C.; Hjelle, B. Hantaviruses: A global disease problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Prescott, J. Hantavirus immunology of rodent reservoirs: Current status and future directions. Viruses 2014, 6, 1317–1335. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.M. Hantaviruses: History and overview. Curr. Top. Microbiol. Immunol. 2001, 256, 1–14. [Google Scholar] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef] [PubMed]
- Clement, J.; Maes, P.; Lagrou, K.; van Ranst, M.; Lameire, N. A unifying hypothesis and a single name for a complex globally emerging infection: Hantavirus disease. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Gizzi, M.; Delaere, B.; Weynand, B.; Clement, J.; Maes, P.; Vergote, V.; Laenen, L.; Hjelle, B.; Verroken, A.; Dive, A.; et al. Another case of “European hantavirus pulmonary syndrome” with severe lung, prior to kidney, involvement, and diagnosed by viral inclusions in lung macrophages. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1341–1345. [Google Scholar] [CrossRef] [PubMed]
- Braun, M.; Bjorkstrom, N.K.; Gupta, S.; Sundstrom, K.; Ahlm, C.; Klingstrom, J.; Ljunggren, H.G. NK cell activation in human hantavirus infection explained by virus-induced IL-15/IL15Rα expression. PLoS Pathog. 2014, 10, e1004521. [Google Scholar] [CrossRef] [PubMed]
- Ennis, F.A.; Cruz, J.; Spiropoulou, C.F.; Waite, D.; Peters, C.J.; Nichol, S.T.; Kariwa, H.; Koster, F.T. Hantavirus pulmonary syndrome: CD8+ and CD4+ cytotoxic T lymphocytes to epitopes on Sin Nombre virus nucleocapsid protein isolated during acute illness. Virology 1997, 238, 380–390. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Rothman, A.L.; Kurane, I.; Montoya, J.M.; Nolte, K.B.; Norman, J.E.; Waite, D.C.; Koster, F.T.; Ennis, F.A. High levels of cytokine-producing cells in the lung tissues of patients with fatal hantavirus pulmonary syndrome. J. Infect. Dis. 1999, 179, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Hammerbeck, C.D.; Hooper, J.W. T cells are not required for pathogenesis in the Syrian hamster model of hantavirus pulmonary syndrome. J. Virol. 2011, 85, 9929–9944. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; Safronetz, D.; Haddock, E.; Robertson, S.; Scott, D.; Feldmann, H. The adaptive immune response does not influence hantavirus disease or persistence in the Syrian hamster. Immunology 2013, 140, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Zaki, S.R.; Greer, P.W.; Coffield, L.M.; Goldsmith, C.S.; Nolte, K.B.; Foucar, K.; Feddersen, R.M.; Zumwalt, R.E.; Miller, G.L.; Khan, A.S.; et al. Hantavirus pulmonary syndrome. Pathogenesis of an emerging infectious disease. Am. J. Pathol. 1995, 146, 552–579. [Google Scholar] [PubMed]
- Calisher, C.H.; Root, J.J.; Mills, J.N.; Rowe, J.E.; Reeder, S.A.; Jentes, E.S.; Wagoner, K.; Beaty, B.J. Epizootiology of Sin Nombre and El Moro Canyon hantaviruses, southeastern Colorado, 1995–2000. J. Wildl. Dis. 2005, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Sweeney, W.; Mills, J.N.; Beaty, B.J. Natural history of Sin Nombre virus in western Colorado. Emerg. Infect. Dis. 1999, 5, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar] [CrossRef] [PubMed]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Bharadwaj, M.; Yee, J.; Ricci, R.; Feddersen, R.M.; Hjelle, B. Experimental infection model for Sin Nombre hantavirus in the deer mouse (Peromyscus maniculatus). Proc. Natl. Acad. Sci. USA 2000, 97, 10578–10583. [Google Scholar] [CrossRef] [PubMed]
- Botten, J.; Mirowsky, K.; Kusewitt, D.; Ye, C.; Gottlieb, K.; Prescott, J.; Hjelle, B. Persistent Sin Nombre virus infection in the deer mouse (Peromyscus maniculatus) model: Sites of replication and strand-specific expression. J. Virol. 2003, 77, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Acuna-Retamar, M.; Feinstein, S.; Prescott, J.; Torres-Perez, F.; Podell, B.; Peters, S.; Ye, C.; Black, W.C.T.; Hjelle, B. Kinetics of immune responses in deer mice experimentally infected with Sin Nombre virus. J. Virol. 2012, 86, 10015–10027. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Prescott, J.; Cogswell, A.C.; Oko, L.; Mirowsky-Garcia, K.; Galvez, A.P.; Hjelle, B. Regulatory T cell-like responses in deer mice persistently infected with Sin Nombre virus. Proc. Natl. Acad. Sci. USA 2007, 104, 15496–15501. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Quackenbush, S.; Rovnak, J.; Haddock, E.; Black, W.C.T.; Feldmann, H.; Prescott, J. Differential lymphocyte and antibody responses in deer mice infected with Sin Nombre hantavirus or Andes hantavirus. J. Virol. 2014, 88, 8319–8331. [Google Scholar] [CrossRef] [PubMed]
- Spengler, J.R.; Haddock, E.; Gardner, D.; Hjelle, B.; Feldmann, H.; Prescott, J. Experimental Andes virus infection in deer mice: Characteristics of infection and clearance in a heterologous rodent host. PLoS ONE 2013, 8, e55310. [Google Scholar] [CrossRef] [PubMed]
- Artsob, H.; LeDuc, J.; Beaty, B.J.; McKinney, R.; Gerone, P.; Purcell, R.; Saluzzo, J.-F.; Gonzalez, J.-P.; Jahrling, P.; Kiley, M. Laboratory management of agents associated with Hantavirus pulmonary syndrome: Interim biosafety guidelines. MMWR 1994, 43, 1–7. [Google Scholar]
- Fulhorst, C.F.; Cajimat, M.N.; Utrera, A.; Milazzo, M.L.; Duno, G.M. Maporal virus, a hantavirus associated with the fulvous pygmy rice rat (Oligoryzomys fulvescens) in western Venezuela. Virus Res. 2004, 104, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Hanson, J.D.; Utrera, A.; Fulhorst, C.F. The delicate pygmy rice rat (Oligoryzomys delicatus) is the principal host of Maporal virus (family Bunyaviridae, genus Hantavirus). Vector Borne Zoonotic Dis. 2011, 11, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Milazzo, M.L.; Eyzaguirre, E.J.; Molina, C.P.; Fulhorst, C.F. Maporal viral infection in the Syrian golden hamster: A model of hantavirus pulmonary syndrome. J. Infect. Dis. 2002, 186, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Cautivo, K.; Schountz, T.; Acuna-Retamar, M.; Ferres, M.; Torres-Perez, F. Rapid enzyme-linked immunosorbent assay for the detection of hantavirus-specific antibodies in divergent small mammals. Viruses 2014, 6, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N.; Johnson, J.M.; Ksiazek, T.G.; Ellis, B.A.; Rollin, P.E.; Yates, T.L.; Mann, M.O.; Johnson, M.R.; Campbell, M.L.; Miyashiro, J.; et al. A survey of hantavirus antibody in small-mammal populations in selected United States National Parks. Am. J. Trop. Med. Hyg. 1998, 58, 525–532. [Google Scholar] [PubMed]
- Schountz, T.; Green, R.; Davenport, B.; Buniger, A.; Richens, T.; Root, J.J.; Davidson, F.; Calisher, C.H.; Beaty, B.J. Cloning and characterization of deer mouse (Peromyscus maniculatus) cytokine and chemokine cDNAs. BMC Immunol. 2004. [Google Scholar] [CrossRef] [Green Version]
- Oko, L.; Aduddell-Swope, B.; Willis, D.; Hamor, R.; Coons, T.A.; Hjelle, B.; Schountz, T. Profiling helper T cell subset gene expression in deer mice. BMC Immunol. 2006. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Shaw, T.I.; Glenn, T.C.; Feldmann, H.; Prescott, J. Expression profiling of lymph node cells from deer mice infected with Andes virus. BMC Immunol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kariwa, H.; Kimura, M.; Yoshizumi, S.; Arikawa, J.; Yoshimatsu, K.; Takashima, I.; Hashimoto, N. Modes of Seoul virus infections: Persistency in newborn rats and transiency in adult rats. Arch. Virol. 1996, 141, 2327–2338. [Google Scholar] [CrossRef] [PubMed]
- Stoltz, M.; Sundstrom, K.B.; Hidmark, A.; Tolf, C.; Vene, S.; Ahlm, C.; Lindberg, A.M.; Lundkvist, A.; Klingstrom, J. A model system for in vitro studies of bank vole borne viruses. PLoS ONE 2011, 6, e28992. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, R.; Amyx, H.L.; Gajdusek, D.C. Experimental infection with Puumala virus, the etiologic agent of nephropathia epidemica, in bank voles (Clethrionomys glareolus). J. Virol. 1985, 55, 34–38. [Google Scholar] [PubMed]
- Hutchinson, K.L.; Rollin, P.E.; Peters, C.J. Pathogenesis of a North American hantavirus, Black Creek Canal virus, in experimentally infected Sigmodon hispidus. Am. J. Trop. Med. Hyg. 1998, 59, 58–65. [Google Scholar] [PubMed]
- Abbott, K.D.; Ksiazek, T.G.; Mills, J.N. Long-term hantavirus persistence in rodent populations in central Arizona. Emerg. Infect. Dis. 1999, 5, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Bagamian, K.H.; Towner, J.S.; Kuenzi, A.J.; Douglass, R.J.; Rollin, P.E.; Waller, L.A.; Mills, J.N. Transmission ecology of Sin Nombre hantavirus in naturally infected North American deermouse populations in outdoor enclosures. PLoS ONE 2012, 7, e47731. [Google Scholar] [CrossRef] [PubMed]
- Kuenzi, A.J.; Morrison, M.L.; Swann, D.E.; Hardy, P.C.; Downard, G.T. A longitudinal study of Sin Nombre virus prevalence in rodents, southeastern Arizona. Emerg. Infect. Dis. 1999, 5, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Schountz, T.; Calisher, C.H.; Richens, T.R.; Rich, A.A.; Doty, J.B.; Hughes, M.T.; Beaty, B.J. Rapid field immunoassay for detecting antibody to Sin Nombre virus in deer mice. Emerg. Infect. Dis. 2007, 13, 1604–1607. [Google Scholar] [CrossRef] [PubMed]
- Safronetz, D.; Ebihara, H.; Feldmann, H.; Hooper, J.W. The Syrian hamster model of hantavirus pulmonary syndrome. Antivir. Res. 2012, 95, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Davis, I.C.; Zajac, A.J.; Nolte, K.B.; Botten, J.; Hjelle, B.; Matalon, S. Elevated generation of reactive oxygen/nitrogen species in hantavirus cardiopulmonary syndrome. J. Virol. 2002, 76, 8347–8359. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; Hall, P.; Acuna-Retamar, M.; Ye, C.; Wathelet, M.G.; Ebihara, H.; Feldmann, H.; Hjelle, B. New World hantaviruses activate IFNλ production in type I IFN-deficient vero E6 cells. PLoS ONE 2010, 5, e11159. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, P.; Schmidt-Chanasit, J.; Gunther, S. The N terminus of Andes virus L protein suppresses mRNA and protein expression in mammalian cells. J. Virol. 2013, 87, 6975–6985. [Google Scholar] [CrossRef] [PubMed]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J. Med. Virol. 2007, 79, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Matthys, V.S.; Cimica, V.; Dalrymple, N.; Glennon, N.B.; Bianco, C.; Mackow, E.R. Hantavirus GnT elements mediate TRAF3 binding and inhibit RIG-I/TBK1 directed IFNbeta transcription by blocking IRF3 phosphorylation. J. Virol. 2014. [Google Scholar] [CrossRef]
- Vera-Otarola, J.; Solis, L.; Soto-Rifo, R.; Ricci, E.P.; Pino, K.; Tischler, N.D.; Ohlmann, T.; Darlix, J.L.; Lopez-Lastra, M. The Andes hantavirus NSs protein is expressed from the viral small mRNA by a leaky scanning mechanism. J. Virol. 2012, 86, 2176–2187. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse # | DPI | Lungs | Heart | ||||
|---|---|---|---|---|---|---|---|
| Cong./Ede | MIAH | IP | PBLH | CD/apop | LIM | ||
| DM 1 | 2 | + | + | - | - | - | + |
| DM 2 | 2 | ++ | + | +/− | - | +/− | +/− |
| DM 3 | 4 | ++ | + | +/− | + | +/− | +/− |
| DM 4 | 4 | + | - | - | - | +/− | +/− |
| DM 5 | 7 | ++ | - | + | + | +/− | + |
| DM 6 | 7 | ++ | +/− | + | ++ | + | ++ |
| DM 7 | 14 | +/− | +/− | - | +/− | - | +/− |
| DM 8 | 14 | +/− | ++ | +/− | - | - | +/− |
| DM 9 | 56 | + | +/− | +/− | +/− | - | +/− |
| DM 10 | 56 | + | + | - | - | - | - |
| Feature | MAPV | SNV | ANDV |
|---|---|---|---|
| Clinical Disease | None | None | None |
| Outcome | Persistence | Persistence | Clearance |
| Nucleocapsid Ab day 14 | Some | Some | All |
| Neutralizing Ab titer | High | High | Low |
| Cytokine gene expression | Low | Low | High |
| Histopathology | Minimal-Mild | Minimal | None |
| Nucleocapsid IHC | None | Low | None |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGuire, A.; Miedema, K.; Fauver, J.R.; Rico, A.; Aboellail, T.; Quackenbush, S.L.; Hawkinson, A.; Schountz, T. Maporal Hantavirus Causes Mild Pathology in Deer Mice (Peromyscus maniculatus). Viruses 2016, 8, 286. https://doi.org/10.3390/v8100286
McGuire A, Miedema K, Fauver JR, Rico A, Aboellail T, Quackenbush SL, Hawkinson A, Schountz T. Maporal Hantavirus Causes Mild Pathology in Deer Mice (Peromyscus maniculatus). Viruses. 2016; 8(10):286. https://doi.org/10.3390/v8100286
Chicago/Turabian StyleMcGuire, Amanda, Kaitlyn Miedema, Joseph R. Fauver, Amber Rico, Tawfik Aboellail, Sandra L. Quackenbush, Ann Hawkinson, and Tony Schountz. 2016. "Maporal Hantavirus Causes Mild Pathology in Deer Mice (Peromyscus maniculatus)" Viruses 8, no. 10: 286. https://doi.org/10.3390/v8100286
APA StyleMcGuire, A., Miedema, K., Fauver, J. R., Rico, A., Aboellail, T., Quackenbush, S. L., Hawkinson, A., & Schountz, T. (2016). Maporal Hantavirus Causes Mild Pathology in Deer Mice (Peromyscus maniculatus). Viruses, 8(10), 286. https://doi.org/10.3390/v8100286