
Cloning the Horse RNA Polymerase I Promoter and Its Application to Studying Influenza Virus Polymerase Activity

{kind=link}
{kind=link}
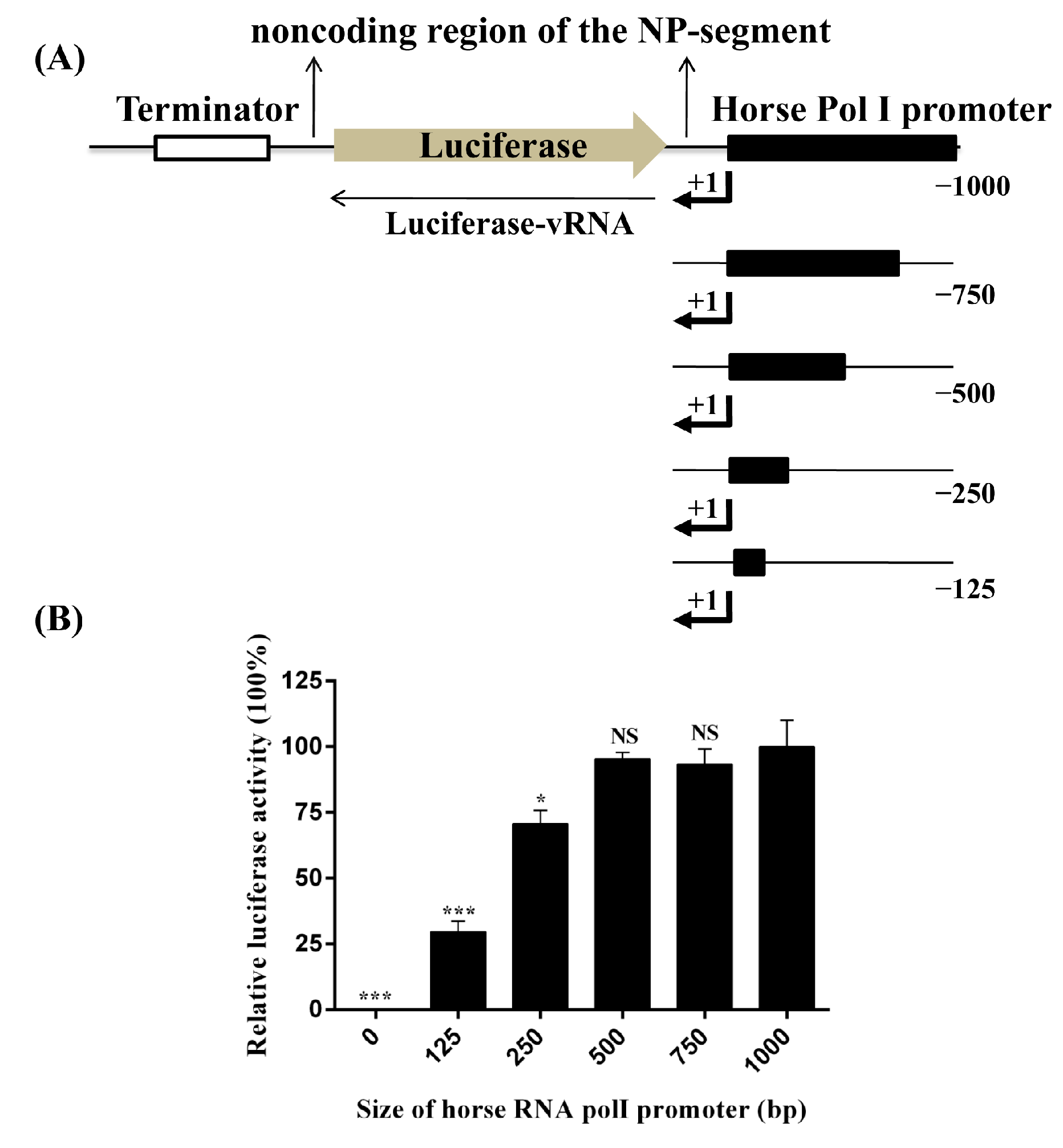{kind=link}
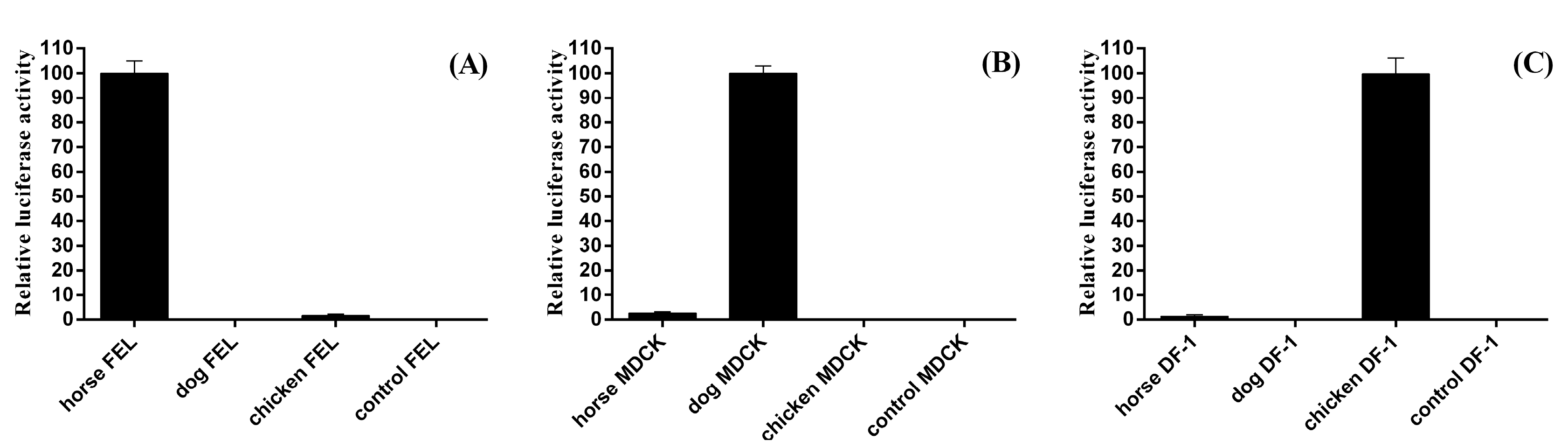{kind=link}
{kind=link}
Abstract
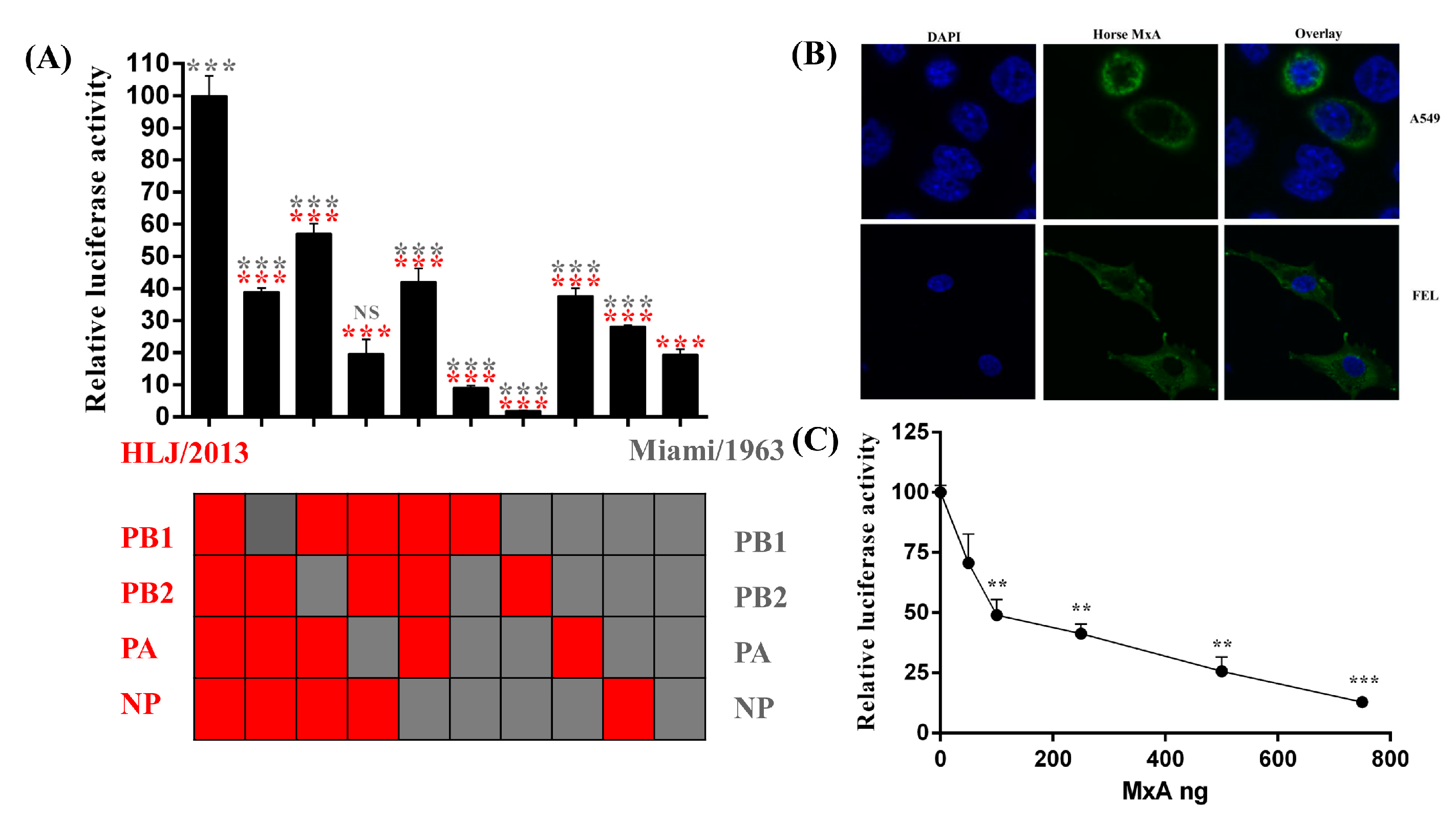
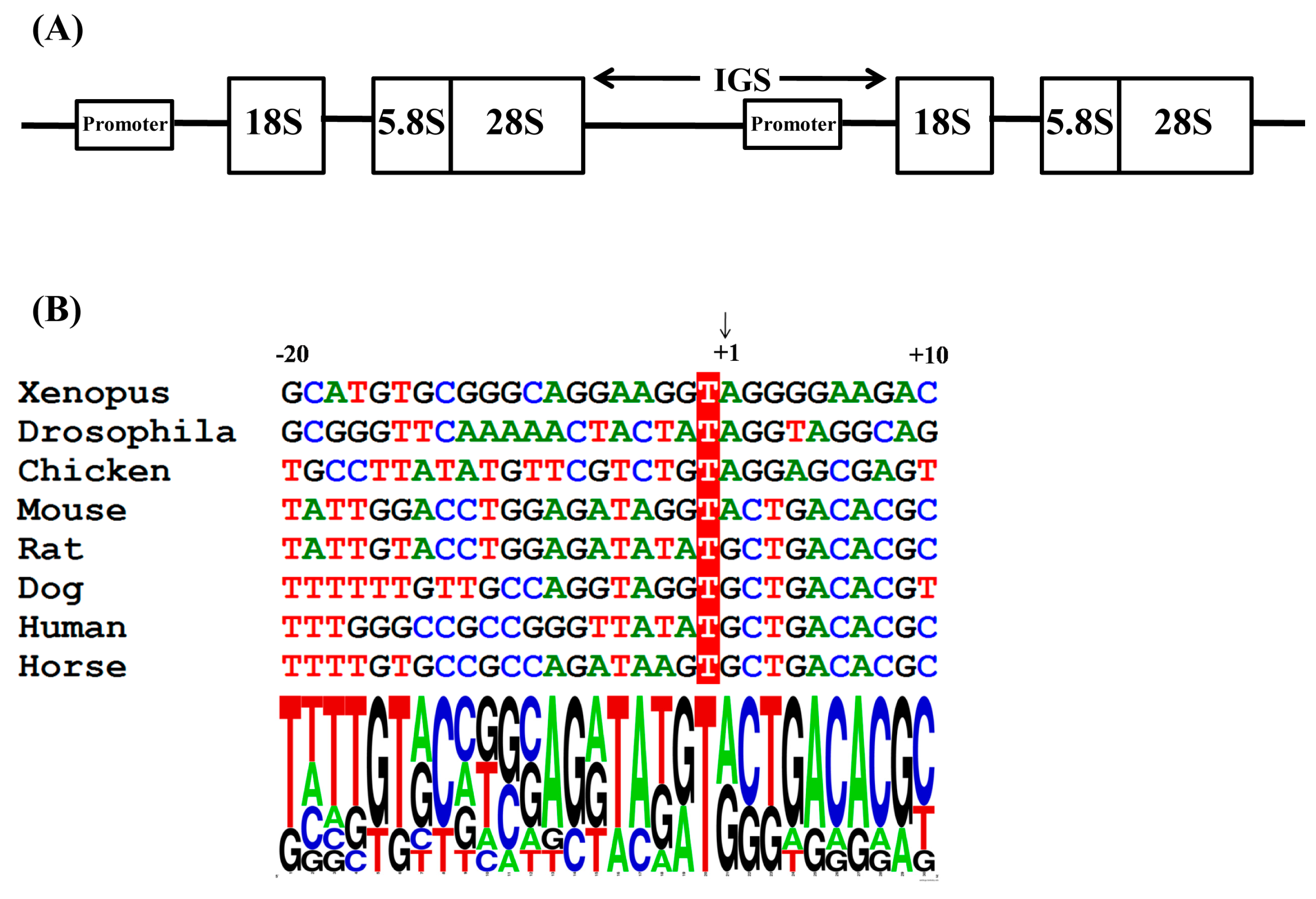
:1. Introduction
2. Results and Discussion
3. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Eisfeld, A.J.; Neumann, G.; Kawaoka, Y. At the centre: Influenza A virus ribonucleoproteins. Nat. Rev. Microbiol. 2015, 13, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Watanabe, T.; Ito, H.; Watanabe, S.; Goto, H.; Gao, P.; Hughes, M.; Perez, D.R.; Donis, R.; Hoffmann, E.; et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. USA 1999, 96, 9345–9350. [Google Scholar] [CrossRef] [PubMed]
- Fodor, E.; Devenish, L.; Engelhardt, O.G.; Palese, P.; Brownlee, G.G.; Garcia-Sastre, A. Rescue of influenza A virus from recombinant DNA. J. Virol. 1999, 73, 9679–9682. [Google Scholar] [PubMed]
- Hoffmann, E.; Neumann, G.; Kawaoka, Y.; Hobom, G.; Webster, R.G. A DNA transfection system for generation of influenza A virus from eight plasmids. Proc. Natl. Acad. Sci. USA 2000, 97, 6108–6113. [Google Scholar] [CrossRef] [PubMed]
- Massin, P.; Rodrigues, P.; Marasescu, M.; van der Werf, S.; Naffakh, N. Cloning of the chicken RNA polymerase I promoter and use for reverse genetics of influenza A viruses in avian cells. J. Virol. 2005, 79, 13811–13816. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Horimoto, T.; Yamada, S.; Kakugawa, S.; Goto, H.; Kawaoka, Y. Establishment of canine RNA polymerase I-driven reverse genetics for influenza A virus: Its application for H5N1 vaccine production. J. Virol. 2008, 82, 1605–1609. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Duke, G.M. Cloning of the canine RNA polymerase I promoter and establishment of reverse genetics for influenza A and B in MDCK cells. Virol. J. 2007, 4. [Google Scholar] [CrossRef] [PubMed]
- Doelling, J.H.; Pikaard, C.S. Species-specificity of rRNA gene transcription in plants manifested as a switch in RNA polymerase specificity. Nucleic Acids Res. 1996, 24, 4725–4732. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Curtiss, R., III. Efficient generation of influenza virus with a mouse RNA polymerase I-driven all-in-one plasmid. Virol. J. 2015, 12. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc. Natl. Acad. Sci. USA 2005, 102, 18590–18595. [Google Scholar] [CrossRef] [PubMed]
- Konig, R.; Stertz, S.; Zhou, Y.; Inoue, A.; Hoffmann, H.H.; Bhattacharyya, S.; Alamares, J.G.; Tscherne, D.M.; Ortigoza, M.B.; Liang, Y.; et al. Human host factors required for influenza virus replication. Nature 2010, 463, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Li, Y.; Rivailler, P.; Conrardy, C.; Castillo, D.A.; Chen, L.M.; Recuenco, S.; Ellison, J.A.; Davis, C.T.; York, I.A.; et al. A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. USA 2012, 109, 4269–4274. [Google Scholar] [CrossRef] [PubMed]
- Moncorge, O.; Mura, M.; Barclay, W.S. Evidence for avian and human host cell factors that affect the activity of influenza virus polymerase. J. Virol. 2010, 84, 9978–9986. [Google Scholar] [CrossRef] [PubMed]
- Bussey, K.A.; Bousse, T.L.; Desmet, E.A.; Kim, B.; Takimoto, T. PB2 residue 271 plays a key role in enhanced polymerase activity of influenza A viruses in mammalian host cells. J. Virol. 2010, 84, 4395–4406. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, G.; Abram, M.; Keiner, B.; Wagner, R.; Klenk, H.D.; Stech, J. Differential polymerase activity in avian and mammalian cells determines host range of influenza virus. J. Virol. 2007, 81, 9601–9604. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Feng, H.; Xu, J.; Zhao, D.; Shi, J.; Li, Y.; Deng, G.; Jiang, Y.; Li, X.; Zhu, P.; et al. The PA protein directly contributes to the virulence of H5N1 avian influenza viruses in domestic ducks. J. Virol. 2011, 85, 2180–2188. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Guo, W.; Qi, T.; Ma, J.; Zhao, S.; Tian, Z.; Pan, J.; Zhu, C.; Wang, X.; Xiang, W. Genetic analysis of the PB1-F2 gene of equine influenza virus. Virus Genes 2013, 47, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Lu, G.; Guo, W.; Qi, T.; Ma, J.; Zhu, C.; Zhao, S.; Pan, J.; Xiang, W. Identification of equine influenza virus infection in asian wild horses (Equus przewalskii). Arch. Virol. 2014, 159, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Van Maanen, C.; Cullinane, A. Equine influenza virus infections: An update. Vet. Q. 2002, 24, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Lai, A.C.; Chambers, T.M.; Holland, R.E., Jr.; Morley, P.S.; Haines, D.M.; Townsend, H.G.; Barrandeguy, M. Diverged evolution of recent equine-2 influenza (H3N8) viruses in the western hemisphere. Arch. Virol. 2001, 146, 1063–1074. [Google Scholar] [CrossRef] [PubMed]
- Grummt, I. Life on a planet of its own: Regulation of RNA polymerase I transcription in the nucleolus. Genes Dev. 2003, 17, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Bilofsky, H.S.; Burks, C. The genbank genetic sequence data bank. Nucleic Acids Res. 1988, 16, 1861–1863. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, T.; Barker, D.; Birney, E.; Cameron, G.; Chen, Y.; Clark, L.; Cox, T.; Cuff, J.; Curwen, V.; Down, T.; et al. The ensembl genome database project. Nucleic Acids Res. 2002, 30, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLOGO: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, R.H.; Kehler, W.H.; Henthorne, J.C. The 1963 equine influenza epizootic. J. Am. Vet. Med. Assoc. 1963, 143, 1108–1110. [Google Scholar] [PubMed]
- Prism, GraphPad, version 5.01; GraphPad Software Inc.: San Diego, CA, USA, 2007.
- Su, S.; Wang, L.; Fu, X.; He, S.; Hong, M.; Zhou, P.; Lai, A.; Gray, G.; Li, S. Equine influenza A(H3N8) virus infection in cats. Emerg. Infect. Dis. 2014, 20, 2096–2099. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Killip, M.J.; Staeheli, P.; Randall, R.E.; Jackson, D. The human interferon-induced MxA protein inhibits early stages of influenza A virus infection by retaining the incoming viral genome in the cytoplasm. J. Virol. 2013, 87, 13053–13058. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Stertz, S.; Kochs, G. The Mx gtpase family of interferon-induced antiviral proteins. Microbes Infect. 2007, 9, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.H.; Takada, A.; Mitsuhashi, T.; Agui, T.; Watanabe, T. Native antiviral specificity of chicken Mx protein depends on amino acid variation at position 631. Anim. Genet. 2004, 35, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, J.; Parthoens, E.; Schepens, B.; Fiers, W.; Saelens, X. Interferon-inducible protein Mx1 inhibits influenza virus by interfering with functional viral ribonucleoprotein complex assembly. J. Virol. 2012, 86, 13445–13455. [Google Scholar] [CrossRef] [PubMed]
- Miotto, O.; Heiny, A.T.; Albrecht, R.; Garcia-Sastre, A.; Tan, T.W.; August, J.T.; Brusic, V. Complete-proteome mapping of human influenza A adaptive mutations: Implications for human transmissibility of zoonotic strains. PLoS ONE 2010, 5, e9025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Liu, K.; Xue, C.Y.; Zhou, J.Q.; Xu, S.; Ren, Y.D.; Zheng, J.; Cao, Y.C. Impact of the segment-specific region of the 3'-untranslated region of the influenza A virus PB1 segment on protein expression. Virus Genes 2013, 47, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, C.W. Sequencing and mutational analysis of the non-coding regions of influenza A virus. Vet. Microbiol. 2009, 135, 239–247. [Google Scholar] [CrossRef] [PubMed]




© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, G.; He, D.; Wang, Z.; Ou, S.; Yuan, R.; Li, S. Cloning the Horse RNA Polymerase I Promoter and Its Application to Studying Influenza Virus Polymerase Activity. Viruses 2016, 8, 119. https://doi.org/10.3390/v8060119
Lu G, He D, Wang Z, Ou S, Yuan R, Li S. Cloning the Horse RNA Polymerase I Promoter and Its Application to Studying Influenza Virus Polymerase Activity. Viruses. 2016; 8(6):119. https://doi.org/10.3390/v8060119
Chicago/Turabian StyleLu, Gang, Dong He, Zengchao Wang, Shudan Ou, Rong Yuan, and Shoujun Li. 2016. "Cloning the Horse RNA Polymerase I Promoter and Its Application to Studying Influenza Virus Polymerase Activity" Viruses 8, no. 6: 119. https://doi.org/10.3390/v8060119