Protoparvovirus Knocking at the Nuclear Door


1
Department of Biological and Environmental Science and Nanoscience Center, University of Jyvaskyla, FI-40500 Jyvaskyla, Finland
2
Laboratoire de Microbiologie Fondamentale et Pathogénicité, University of Bordeaux, UMR 5234, F-33076 Bordeaux, France
3
Centre national de la recherche scientifique (CNRS), Microbiologie Fondamentale et Pathogénicité, UMR 5234, F-33076 Bordeaux, France
4
Centre Hospitalier Universitaire de Bordeaux, Service de Virologie, F-33076 Bordeaux, France
*
Author to whom correspondence should be addressed.
Viruses 2017, 9(10), 286; https://doi.org/10.3390/v9100286
Submission received: 5 September 2017
/
Revised: 28 September 2017
/
Accepted: 29 September 2017
/
Published: 2 October 2017
(This article belongs to the Special Issue Protoparvoviruses: Friends or Foes?)
{kind=link}
Abstract
:Protoparvoviruses target the nucleus due to their dependence on the cellular reproduction machinery during the replication and expression of their single-stranded DNA genome. In recent years, our understanding of the multistep process of the capsid nuclear import has improved, and led to the discovery of unique viral nuclear entry strategies. Preceded by endosomal transport, endosomal escape and microtubule-mediated movement to the vicinity of the nuclear envelope, the protoparvoviruses interact with the nuclear pore complexes. The capsids are transported actively across the nuclear pore complexes using nuclear import receptors. The nuclear import is sometimes accompanied by structural changes in the nuclear envelope, and is completed by intranuclear disassembly of capsids and chromatinization of the viral genome. This review discusses the nuclear import strategies of protoparvoviruses and describes its dynamics comprising active and passive movement, and directed and diffusive motion of capsids in the molecularly crowded environment of the cell.
1. Introduction
DNA viruses (with exception of poxviridae family), and some RNA viruses, such as retroviruses, orthomyxoviruses and bornaviruses, have to enter the cell nucleus due to their need of the nuclear DNA replication and transcription machinery. Their genomes have to be transported from the cell periphery into the nucleus facilitated by multiple coordinated interactions between the virus (or a subviral structure) and host proteins. Further, the viral genome has to be released at a defined site in order to allow replication. Growing evidence shows that, for most viruses, uncoating occurs at the cytoplasmic side of the nuclear envelope (NE).
The genus protoparvovirus in the subfamily Parvovirinae includes naked single-stranded DNA viruses with a wide host spectrum from carnivores and primates to rodents [1]. Among carnivores they infect various species including dogs (canine parvovirus (CPV)), minks (mink enteritis virus (MEV)) and cats (feline parvovirus (FPV)). Primate protoparvoviruses infect humans (bufavirus, tusavirus, cutavirus), rodent protoparvoviruses infect mice (minute virus of mice (MVM)) and rats (parvovirus H-1 (PV H-1), rat parvovirus 1 (RPV1)), and ungulate protoparvoviruses infect swines (porcine parvovirus (PPV)). While some protoparvoviruses cause diseases such as MVM, CPV and FPV, others are not associated with any illness, e.g., bufavirus and PV H-1. Research on PVs is not only driven by disease prevention and cure but also by their use in parvovirus-based virotherapy. Some protoparvoviruses like PV H-1 possess natural oncotropism and their cytotoxicity is used in experimental cancer therapy [2,3].
Members of protoparvoviruses have a small, icosahedral and a non-enveloped capsid of 18–26 nm in diameter. The capsid encloses a single-stranded, linear, non-segmented and often negative-sense DNA genome of 4–6 kb in length with duplex hairpin telomeres at both termini [4,5]. Although rare, certain protoparvovirus species (e.g., LuIII) also encapsidate positive strands [6]. While the genome has two genes transcribed from two open reading frames (ORFs) [7,8], alternative splicing and proteolytic cleavage account for synthesis of 4 to 5 proteins. The left-hand ORF encodes two non-structural (NS1-2) proteins, both having multiple functions in the viral life cycle. NS1 regulates viral gene expression [9], and induces cell cycle arrest and apoptosis [10,11]. NS1 is also involved in the induction of single-stranded DNA breaks, blockage of cellular DNA replication [12] and alteration of cytoskeletal and the nuclear structures [13,14]. In virions, the protein is covalently bound to the 5’ end of the viral DNA [15]. Exposed on the virus surface, it becomes removed during cell entry upon new infection [16]. NS2 plays an indirect role in parvovirus infection facilitating viral replication and capsid assembly. NS2 interactions are not well understood but it has a critical role in active nuclear export of mature particles via binding to the nuclear export factor Crm1 and modulation of viral host range [17,18,19,20].
The right-hand ORF encodes three structural viral proteins (VPs 1–3). The capsid has a T = 1 symmetry formed from 60 copies of the VPs, which are splice variants of the VP1 gene [21]. The largest protein, VP1 constitutes ~10% of the capsid proteins. The major capsid protein is VP2, which is involved in the nuclear assembly of capsids [22,23]. The VP1 and VP2 have a common C-terminal sequence and the complete sequence of VP2 (64 to 66 kD) is contained in VP1 (83 to 86 kD) [5,24]. In addition, the VP1 has a unique N-terminal domain (VP1u). VP3 is the smallest capsid protein formed by a 15 to 20 aa cleavage from the N-terminus of VP2. It is only found in mature DNA-containing capsids, where its ratio with VP2 defines the virus species and maturity [25,26]. Because of their high immunogenicity, the VPs are the main antigens of these viruses [27,28].
Parvoviruses exploit multiple cellular processes including endocytic pathways, and active nuclear transport during the initial phase of infection. Capsids enter the cell via receptor-mediated endocytosis for genome transport toward the nucleus. Transport to the nuclear periphery is followed by endosomal release of the capsids into the cytoplasm [29,30,31,32,33,34]. Nuclear import of the genome is dependent on capsid interactions with the nuclear transport machinery, including members of the karyopherin β superfamily (importins) and nuclear pore complexes (NPCs). Nuclear import of capsids and their disassembly are followed by genome replication and viral assembly, which require the import of structural and nonstructural viral proteins.
2. Cytoplasmic Transport
At early stages of the endocytic pathway, vesicles from the plasma membrane are rapidly targeted to the early endosomes. Maturation of early-to-late endosomes with an acidic pH of 4.5–5.5 is followed by the formation of enlarged multivesicular structures [35]. The fusion of an endosome with a lysosome generates a transient hybrid organelle, the endolysosome [36]. The endolysosomes are converted to classical dense lysosomes, which constitute a storage organelle for lysosomal hydrolases used for degradation of macromolecules [37]. Evidently, viruses using this entry route had to develop a strategy for endosomal escape prior to lysosomal degradation, with the exception of reoviruses, which require lysosomal proteases for their partial uncoating [38].
After the stepwise endosomal uptake, CPV and other parvoviruses, including adeno-associated virus 2 (AAV2), a member of the genus dependoparvovirus, are present in small vesicles scattered around the cytoplasm, and in large vesicles accumulated in the perinuclear area [30,39,40,41,42,43]. This is in line with earlier observations of size-distribution of cargo-filled endosomes and their perinuclear accumulation: small endosomes with little cargo are found in the cell periphery, whereas large endosomes rich in cargo are located close to the nucleus [44,45,46]. The endosome-associated movement of viral capsids fluctuates from fast to slow. The fast movement could correspond to capsids located in small endosomes actively moving along microtubules (MTs) [43]. The slow movement corresponds to capsids accumulated in endosomes at the perinuclear area, where they move very slowly or are nearly immobile (velocity < 1 µm/s) [43,45,46,47,48,49,50,51]. In parvovirus entry, the endosomal transportation is followed by capsid release into cytoplasm, which appears to be inefficient. This is induced by the low endosomal pH leading to activation of a phospholipase A2 (PLA2) activity located in the N terminus of VP1 [52].
The high viscosity and molecular crowding restricts viral capsid motility in the cytoplasm requiring active and directed transport to the nucleus. Active transport can be facilitated by actin polymerization as observed for intracellular vaccinia- and baculoviruses [53,54]. However, nearly all viruses with a nuclear phase use cytoplasmic dynein motor complex for active transport along MT to reach the perinuclear space [55,56]. Dynein is composed of 12 to 14 different chains, and moves cargos at a velocity of ≈2–3.5 μm/s [57]. Cargo-dynein interactions are based on various binding partners including the dynein–dynactin complex [58]. Dynein-mediated transport is also used by protoparvovirus capsids to reach the NE [43,59,60,61,62,63,64,65,66,67,68], which is consistent with the velocity of 3.5 and ~2.0 μm/s observed for AAV2 and AAV9 [41,69,70].
3. Molecular Mechanisms of Nuclear Import
Nuclear viruses infecting non-dividing, e.g., resting cells must have developed strategies to pass the NE. The majority of them use NPC-mediated transport. An alternative pathway was described for the simian virus 40 (SV40), which not only traverses nuclear pores, but is also able to disrupt the inner nuclear membrane after internalization into the ER lumen [71].
NPCs are macromolecular structures composed of approximately 30 different protein species collectively called nucleoporins (Nups) [72,73]. Filaments extending in the cytoplasm are composed of, e.g., Nup358 (RanBP2) and Nup 214, whereas others such as Nup62 are part of NE-embedded scaffold rings, which form the hydrophobic transport channel [74]. On the nuclear side of NPC, eight filaments extrude the central part constituting the nuclear basket formed of Nups Tpr and Nup153. Nup153 may be flexible as its localization on the cytoplasmic side also has been reported [75]. The channel through NPC has a functional limit diameter of ~39 nm [76]. It is filled with a mesh of hydrophobic phenylalanine-glycine (FG) repeat-containing Nup domains [77,78,79] restricting size- and charge-dependent passive diffusion of molecules smaller than ~20–40 kDa (diameter < 4–5 nm) [80,81,82].
Active nuclear import of larger molecules of <25 megadaltons (diameter < 40 nm) is facilitated by nuclear import receptors (importins or karyopherins; Kaps) [83]. A total of nine different receptors have been described, differing in their cargo specificity. They mediate approximately 1000 translocations per NPC per second [84,85]. Given that there are 400 to 18,500 NPCs per nucleus [86], the transport receptors allow millions of exchange reactions per second per cell. The prerequisite for nuclear localization of molecular cargo is the NLSs. The classical NLSs (cNLSs) are formed of mono- or bipartite clusters of typically four to six basic amino acids. The prototype NLS found in SV40 T-antigen (SV40TAg) has the amino acid sequence PKKKRKV [87]. In the classical import pathway, the cNLS is bound by importin α (Kapα) adaptor protein, which binds to importin β (Kapβ1), and the formed trimeric importin α-importin β-NLS complex can enter the nucleus. Moreover, importin β can directly interact with cargos comprising an importin-β-binding domain [88].
Interaction of the import receptor with the NLS on the cargo can be regulated by post-translational modifications of the NLS or nearby residues. Alternatively, the NLS can be exposed or hidden, either based on structural changes of the protein with NLS (e.g., phosphorylation of SV40Tag) [89] or by protein–protein interactions (e.g., NFkB-IkB) [90]. Attachment of the cargo modifies the structure of import receptor, which allows its interaction with Nups and passage through the nuclear pore. The import reaction is terminated in the nuclear basket where the cargo-import receptor-complex binds to Nup153 [91]. This leads to dissociation of import receptor from its cargo, a process induced by GTP-bound form of Ras-related nuclear protein (Ran) [92,93]. While the cargo diffuses deeper into the nucleus, the import receptor-RanGTP-complex is exported through the NPC [88].
3.1. Nuclear Entry of Viral DNA Genomes
The nuclear import of most viruses and their genomes is restricted by the size of the virus capsid. Most viruses exceed the maximal size of the nuclear pore thus requiring their disassembly on the cytoplasmic side of NPC. This is well documented for adenoviral capsids, which disassemble to pentons and hexons after their docking to Nup214 and binding to kinesin 1 without need of nuclear import receptors [94,95]. It is assumed that the pulling forces by kinesin 1 on capsid bound to NPCs disintegrate them and lead to viral genome release [96,97]. The genome, in complex with the viral proteins VII, X and TP (terminal protein), then interacts indirectly with the nuclear import receptors. While, the identity of these import receptors has remained unclear, importin β, importin-7 and transportin-1 have been suggested together with adapter molecules hsp70 and/or histone H1 [95,98,99].
In contrast, herpes simplex virus capsids with a diameter of 125 nm associate with the NPCs either by direct binding of the inner tegument protein pUL25 to Nup214 or to importin β [100,101,102]. NPC-interaction acts as a triggering element for herpesviral DNA release, which is poorly understood but may involve proteolytic cleavage of UL36 [103]. During capsid assembly the HSV genome is packed in an energy-consuming process leading to a quasi-crystalline density [104]. Opening of the capsid at a site opposing the NPC causes an injection of DNA through the nuclear pore. After the repulsion comes to an end, the part of the genome which remains outside the nucleus and/or in the capsid is pulled into the nucleus by transcription of the nuclear genome end encoding for the immediate-early genes [105].
Very few viral capsids have diameters below the nuclear transport limit; amongst them parvoviral and circoviral capsids (15–30 nm), and that of hepatitis B virus (HBV; 36 nm). Nuclear import of circovirus genomes is practically uninvestigated, yet it was shown that the only capsid protein Cap comprises the NLS [106]. HBV capsids are composed of a single protein species, which contains the NLS [107] and also an importin-β-binding domain [108]. Accordingly, HBV capsids are imported into the nuclear basket using importin α and β, but in contrast to classical cargos the capsid shows high affinity for Nup153 so that the capsid becomes arrested inside the nuclear basket [109]. Genome release of HBV is poorly understood; however, it does require capsid disassembly to core protein dimers, as with adenoviruses.
3.2. Nuclear Import of Protoparvoviral Capsids
Acidification of parvoviral capsids upon endosomal entry leads to exposure of the N terminal part of VP1, hidden in the virion, and absent from VP2 (VP1 unique; VP1u). Aside of the PLA2 domain, described before, CPV VP1 comprises one stretch of basic amino acids (4PAKRARRGYK13) similar to classical NLSs (cNLSs) [110]. In addition, four stretches have been found within MVM VP1 [60]. This sequence was shown to mediate nuclear import of bovine serum albumin, and mutational analysis exhibited its importance in early CPV infection [111,112]. The closest relative of CPV, the feline panleukopeniavirus (FPV), as well as mink enterititis virus (MEV) and blue fox parvovirus Tai’an have similar sequences (4PAKRARRGLV13) in their VP1u proteins. Such cNLSs were further identified on Ungulate parvovirus 1 species porcine parvovirus (PPV) VP1u and MVM VP1-N. In contrast, MVM, an example of the Rodent protoparvovirus 1 species, have, in addition with two cNLSs on VP1 and VP2 proteins, a non-classical NLS (ncNLS) nuclear localization motif (NLM) in their C-terminal region of VP1/2. These NLMs are conserved amongst protoparvoviruses [22,60]. Other protoparvoviruses such as hamster parvovirus H1 and LuIII (Rodent protoparvovirus 1), and rat parvovirus 1 (Rodent protoparvovirus 2) have predicted NLS motifs (4PAKRAKRGWV13). The sequence analysis showed that such putative cNLS sequences are also found in VP1/VP2 of human protoparvoviruses, bufa-, tusa- and cutaviruses [113].
Parvoviral capsids can be observed within the nucleus shortly after infection [62,114]. The observation of the conserved NLS combined with the attachment of at least importin β to cytosolic CPV capsids suggest that the capsids follow the classical nuclear import pathway. In fact, capsids have been observed on the nuclear side of the NPC [43]. Observations using the AAV2 and the protoparvovirus PV H-1 showed that their attachment to NE causes its local degradation [107]. This nuclear envelope breakdown (NEBD) is similar to mitotic NEBD, both of them need PKCα, cyclin-dependent kinases and Ca2+. The NEBD that is facilitated by host caspases and mitotic enzymes in PV H-1 and MVM infected cells [115,116,117]. Phospholipase A2 activity on MVM capsids has not been reported to be involved in causing NE disruption. Instead, the virus utilizes cellular caspases, especially caspase-3, a protease involved in NE breakdown during apoptosis, to facilitate nuclear membrane disruptions [117]. Interestingly, piercing or bending of the NE by MT during mitosis was shown to trigger NEBD. The capsid attachment to Nup153 suggests that capsids are able to enter the nuclear basket allowing for their access and permeabilization of the NE. This process is independent on PLA2, at least for AAV2 [118]. Of note, Ca2+ release was a prerequisite for parvovirus-mediated activation of PKC, as of cyclin-dependent kinases, yet called for parvovirus interaction with the NPC (e.g., Nup358, Nup62 and Nup153). Finally, microinjection of MVM capsids into the cytoplasm of Xenopus laevis oocytes resulted in the damage of nuclear envelope allowing NPC-independent entry of capsids [116]. Notwithstanding the apparent contradiction (Figure 1), the observations in concert suggest that protoparvoviruses first attach to the NPC directly or via importin β (and potentially importin α), and then penetrate the nuclear pore. The nuclear import through the NPC is able to trigger an extensive modification of nuclear envelope leading to NEBD facilitating the passage of capsid into the nucleus, however, this does not rule out the possibility that a portion of the capsids might be actively transported into the nucleus by importins through the intact NPCs and the nuclear envelope. At the moment, part of underlying mechanisms and specific interactions leading to parvovirus nuclear entry are still unknown and additional studies are required for better understanding these processes.
3.3. Nuclear Entry of Capsid Subunits
Although the importance of the conventional NLS on parvoviral VP1 is substantiated by its phylogenetic conservation, its activities during the viral life cycle are not fully understood. It is known to be involved in NPC interaction during viral entry and in nuclear import of newly synthesized viral capsid proteins during the S-phase of the cell cycle (Figure 1). Nuclear translocation of the newly formed VP2 protein, without cNLS, is driven by a structured nuclear localization motif (NLM) [119]. The studies with MVM have demonstrated that VP1 and VP2 form a trimer prior to nuclear entry. It seems that the NLMs dominate over the NLSs of VP1, driving VP2 and VP1/VP2 subunits (2VP2/1VP1, and 3VP2) into the nucleus. The trimers have the capacity to form capsids in the cytosol when subjected to cell contact-triggered density-arrest signals. This has been shown in MVM studies, where the cell cycle arrest in G0, G1 or G1/S transition inhibits nuclear transport of the VPs [119]. It is also known that the NLM is exposed on VP1/VP2 trimers but not on assembled capsids [22,60]. VP1/VP2 heterotrimers undergo modifications that are required for their nuclear import in a cell cycle-dependent manner. Searching for the molecular mechanism revealed that VP2 phosphorylation by the protein kinase Raf-1 is required [120]. Accordingly, VP1/VP2 heterotrimers expressed in insect cells, which are devoid of Raf-1, failed to enter the nucleus and Raf-1 expression correlated with cell permissiveness to MVM infection [120].
4. End of Import-Capsid Disassembly in the Nucleus
Viral gene expression in the nucleus requires uncoating and genome delivery from a protecting protein shell. As protoparvovirus capsids enter the nucleus interior, they must maintain their genome in an encapsidated state until they reach in the nucleoplasm. How and where the capsids uncoat for gene expression not until nuclear entry is a long-standing question remaining to be answered. Earlier studies have shown that in infectious parvovirus particles, 20–30 nucleotides of the 5’ end are exposed on the capsid surface [5,16,121,122]. However, the roles of these 5’ ends or of the NLS in nuclear release of viral DNA are unknown. Although the molecular mechanisms of capsid disassembly are yet unknown, recent studies of intranuclear CPV capsids dynamics have indicated that slow moving nucleoplasmic CPV capsids are accompanied by the rapidly moving capsid-derived components, either remnants thereof or dissociated constituent proteins. This shows that capsids are disassembled after their entry into the nucleoplasm [43]. Within the nucleus, once the genome becomes released, transcription and replication are initiated via nuclear host proteins. Recent studies have shown that epigenetic modification of the parvoviral genome, specifically the activation of promoters, plays a critical role in progression of the parvoviral life cycle [123,124,125,126].
5. Concluding Remarks
The nuclear pore complex constitutes a gate for the entry of viral DNA genome into the nucleus. Viruses have found multiple strategies to allow the passage of their genome. They include disassembly either in the cytosol or at the NPC followed by nuclear import of a genomic complex through the NPC. The genome of some viruses can be injected into the nucleus from a capsid docked at the pore complex. Parvoviruses with capsid diameters falling below the exclusion limit fit in through the NPC. The import is facilitated by NLSs and interactions with nuclear import machinery. Subsequently, these interactions can promote viral capsid entry by inducing NE disintegration. The elucidation of the diverse strategies of parvoviral nuclear import will no doubt shed new light on the complexity of this process and lead to improved means of cancer treatment and gene therapy.
Acknowledgments
We are grateful to Klaus Hedman for comments on the manuscript. The research was financed by the Jane and Aatos Erkko Foundation (Vihinen-Ranta), the Finnish cultural foundation (Mäntylä), and the Academy of Finland under the award number 138388 (Vihinen-Ranta). We apologize to all our colleagues whose work could not be cited because of length restrictions.
Author Contributions
Elina Mäntylä, Michael Kann and Maija Vihinen-Ranta wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J. The family Parvoviridae. Arch. Virol. 2014, 159, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Angelova, A.L.; Geletneky, K.; Nüesch, J.P.; Rommelaere, J. Tumor selectivity of oncolytic parvoviruses: From in vitro and animal models to cancer patients. Front. Bioeng. Biotechnol. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Marchini, A.; Bonifati, S.; Scott, E.M.; Angelova, A.L.; Rommelaere, J. Oncolytic parvoviruses: From basic virology to clinical applications. Virol. J. 2015, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Reed, A.P.; Jones, E.V.; Miller, T.J. Nucleotide sequence and genome organization of canine parvovirus. J. Virol. 1988, 62, 266–276. [Google Scholar] [PubMed]
- Xie, Q.; Chapman, M.S. Canine parvovirus capsid structure, analyzed at 2.9 Å resolution. J. Mol. Biol. 1996, 264, 497–520. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.; Tattersal, P. Genome packaging sense is controlled by the efficiency of the nick site in the right-end replication origin of parvoviruses minute virus of mice and LuIII. J. Virol. 2014, 79, 2287–2300. [Google Scholar] [CrossRef] [PubMed]
- Hanson, N.D.; Rhode, S.L. Parvovirus NS1 stimulates P4 expression by interaction with the terminal repeats and through DNA amplification. J. Virol. 1991, 65, 4325–4333. [Google Scholar] [PubMed]
- Tullis, G.; Schoborg, R.V.; Pintel, D.J. Characterization of the temporal accumulation of minute virus of mice replicative intermediates. J. Gen. Virol. 1994, 75, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Gottlieb, R.L.; Tattersall, P. Replication initiator protein NS1 of the parvovirus minute virus of mice binds to modular divergent sites distributed throughout duplex viral DNA. J. Virol. 2007, 81, 13015–13027. [Google Scholar] [CrossRef] [PubMed]
- Moffatt, S.; Yaegashi, N.; Tada, K.; Tanaka, N.; Sugamura, K. Human parvovirus B19 nonstructural (NS1) protein induces apoptosis in erythroid lineage cells. J. Virol. 1998, 72, 3018–3028. [Google Scholar] [PubMed]
- Morita, E.; Nakashima, A.; Asao, H.; Sato, H.; Sugamura, K. Human parvovirus B19 nonstructural protein (NS1) induces cell cycle arrest at G(1) phase. J. Virol. 2003, 77, 2915–2921. [Google Scholar] [CrossRef] [PubMed]
- Op de Beeck, O.D.; Caillet-Fauquet, P. The NS1 protein of the autonomous parvovirus minute virus of mice blocks cellular DNA replication: A consequence of lesions to the chromatin? J. Virol. 1997, 71, 5323–5329. [Google Scholar] [PubMed]
- Nüesch, J.P.F.; Lachmann, S.; Rommelaere, J. Selective alterations of the host cell architecture upon infection with parvovirus minute virus of mice. Virology 2005, 331, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Ihalainen, T.O.; Niskanen, E.A.; Jylhävä, J.; Paloheimo, O.; Dross, N.; Smolander, H.; Langowski, J.; Timonen, J.; Vihinen-Ranta, M. Parvovirus induced alterations in nuclear architecture and dynamics. PLoS ONE 2009, 4, e5948. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; Tattersall, P. High-mobility group 1/2 proteins are essential for initiating rolling-circle-type DNA replication at a parvovirus fairpin origin. J. Virol. 1998, 72, 8477–8484. [Google Scholar] [PubMed]
- Cotmore, S.F.; Tattersall, P. A genome-linked copy of the NS-1 polypeptide is located on the outside of infectious parvovirus particles. J. Virol. 1989, 63, 3902–3911. [Google Scholar] [PubMed]
- Bodendorf, U.; Cziepluch, C.; Jauniaux, J.; Rommelaere, J.; Salomé, N. Nuclear export factor CRM1 interacts with nonstructural proteins NS2 from parvovirus minute virus of mice. J. Virol. 1999, 73, 7769–7779. [Google Scholar] [PubMed]
- Miller, C.L.; Pintel, D.J. Interaction between parvovirus NS2 protein and nuclear export factor Crm1 is important for viral egress from the nucleus of murine cells. J. Virol. 2002, 76, 3257–3266. [Google Scholar] [CrossRef] [PubMed]
- López-Bueno, A.; Valle, N.; Gallego, J.M.; Pérez, J.; Almendral, J.M. Enhanced cytoplasmic sequestration of the nuclear export receptor CRM1 by NS2 mutations developed in the host regulates parvovirus fitness. J. Virol. 2004, 78, 10674–10684. [Google Scholar] [CrossRef] [PubMed]
- Engelsma, D.; Valle, N.; Fish, A.; Salomé, N.; Almendral, J.M.; Fornerod, M. A supraphysiological nuclear export signal is required for parvovirus nuclear export. Mol. Biol. Cell 2008, 19, 2544–2552. [Google Scholar] [CrossRef] [PubMed]
- Agbandje-McKenna, M.; Chapman, M.S. Correlating structure with function in the viral capsid. In Parvoviruses; Kerr, J., Cotmore, S.F., Bloom, M.E., Linden, R.M., Parrish, C.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 125–139. ISBN 9780340811986. [Google Scholar]
- Lombardo, E.; Ramírez, J.C.; Agbandje McKenna, M.; Almendral, J.M. A β-stranded motif drives capsid protein oligomers of the parvovirus minute virus of mice into the nucleus for viral assembly. J. Virol. 2000, 74, 3804–3814. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Martínez, C.; Grueso, E.; Carroll, M.; Rommelaere, J.; Almendral, J.M. Essential role of the unordered VP2 n-terminal domain of the parvovirus MVM capsid in nuclear assembly and endosomal enlargement of the virion fivefold channel for cell entry. Virology 2012, 432, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Tattersall, P.; Cawte, P.J.; Shatkin, A.J.; Ward, D.C. Three structural polypeptides coded for by minite virus of mice, a parvovirus. J. Virol. 1976, 20, 273–289. [Google Scholar] [PubMed]
- Tsao, J.; Chapman, M.S.; Agbandje, M.; Keller, W.; Simth, K.; Wu, H.; Luo, M.; Smith, T.J.; Rossmann, M.G.; Compans, R.W. The three-dimensional structure of canine parvovirus and its functional implications. Science 1991, 251, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.S.; Agbandje-McKenna, M. Atomic structure of viral particles. In Parvoviruses; Kerr, J., Cotmore, S.F., Bloom, M.E., Linden, R.M., Parrish, C.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 107–123. ISBN 9780340811986. [Google Scholar]
- Tullis, G.E.; Burger, L.R.; Pintel, D.J. The minor capsid protein VP1 of the autonomous parvovirus minute virus of mice is dispensable for encapsidation of progeny single-stranded DNA but is required for infectivity. J. Virol. 1993, 67, 131–141. [Google Scholar] [PubMed]
- Nelson, C.D.S.; Palermo, L.S.; Hafenstein, S.L.; Parrish, C.R. Different mechanisms of antibody-mediated neutralization of parvoviruses revealed using the Fab fragments of monoclonal antibodies. Virology 2007, 361, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Kalela, A.; Mäkinen, P.; Kakkola, L.; Marjomäki, V.; Vuento, M. Intracellular route of canine parvovirus entry. J. Virol. 1998, 72, 802–806. [Google Scholar] [PubMed]
- Parker, J.S.L.; Parrish, C.R. Cellular uptake and infection by canine parvovirus involves rapid dynamin-regulated clathrin-mediated endocytosis, followed by slower intracellular trafficking. J. Virol. 2000, 74, 1919–1930. [Google Scholar] [CrossRef] [PubMed]
- Suikkanen, S.; Sääjärvi, K.; Hirsimäki, J.; Välilehto, O.; Reunanen, H.; Vihinen-Ranta, M.; Vuento, M. Role of recycling endosomes and lysosomes in dynein-dependent entry of canine parvovirus. J. Virol. 2002, 76, 4401–4411. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Suikkanen, S.; Parrish, C.R. Pathways of cell infection by parvoviruses and adeno-associated viruses. J. Virol. 2004, 78, 6709–6714. [Google Scholar] [CrossRef] [PubMed]
- Cureton, D.K.; Harbison, C.E.; Cocucci, E.; Parrish, C.R.; Kirchhausen, T. Limited transferrin receptor clustering allows rapid diffusion of canine parvovirus into clathrin endocytic structures. J. Virol. 2012, 86, 5330–5340. [Google Scholar] [CrossRef] [PubMed]
- Garcin, P.O.; Panté, N. The minute virus of mice exploits different endocytic pathways for cellular uptake. Virology 2015, 482, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Sorkin, A.; von Zastrow, M. Signal transduction and endocytosis: Close encounters of many kinds. Nat. Rev. Mol. Cell Biol. 2002, 3, 600–614. [Google Scholar] [CrossRef] [PubMed]
- Bird, P.I.; Trapani, J.A.; Villadangos, J.A. Endolysosomal proteases and their inhibitors in immunity. Nat. Rev. Immunol. 2009, 9, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Luzio, J.P.; Pryor, P.R.; Bright, N.A. Lysosomes: Fusion and function. Nat. Rev. Mol. Cell Biol. 2007, 8, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Golden, J.W.; Linke, J.; Schmechel, S.; Thoemke, K.; Schiff, L.A. Addition of exogenous protease facilitates reovirus infection in many restrictive cells. J. Virol. 2002, 76, 7430–7443. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Yuan, W.; Parrish, C.R. Cytoplasmic trafficking of the canine parvovirus capsid and its role in infection and nuclear transport. J. Virol. 2000, 74, 4853–4859. [Google Scholar] [CrossRef] [PubMed]
- Lux, K.; Goerlitz, N.; Schlemminger, S.; Perabo, L.; Goldnau, D.; Endell, J.; Leike, K.; Kofler, D.M.; Finke, S.; Hallek, M.; et al. Green fluorescent protein-tagged adeno-associated virus particles allow the study of cytosolic and nuclear trafficking. J. Virol. 2005, 79, 11776–11787. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Samulski, R.J. Cytoplasmic trafficking, endosomal escape, and perinuclear accumulation of adeno-associated virus type 2 particles are facilitated by microtubule network. J. Virol. 2012, 86, 10462–10473. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, S.C.; Samulski, R.J. Recombinant adeno-associated virus utilizes host cell nuclear import machinery to enter the nucleus. J. Virol. 2014, 88, 4132–4144. [Google Scholar] [CrossRef] [PubMed]
- Mäntylä, E.; Chacko, J.V.; Aho, V.; Parrish, C.R.; Shahin, V.; Kann, M.; Digman, M.A.; Gratton, E.; Vihinen-Ranta, M. Viral highway to nucleus exposed by image correlation analyses. Sci. Rep. 2017. under review. [Google Scholar]
- Rink, J.; Ghigo, E.; Kalaidzidis, Y.; Zerial, M. Rab conversion as a mechanism of progression from early to late endosomes. Cell 2005, 122, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Huotari, J.; Helenius, A. Endosome maturation. EMBO J. 2011, 30, 3481–3500. [Google Scholar] [CrossRef] [PubMed]
- Foret, L.; Dawson, J.; Villaseñor, R.; Collinet, C.; Deutsch, A.; Brusch, L.; Zerial, M.; Kalaidzidis, Y.; Jülicher, F. A General theoretical framework to infer endosomal network dynamics from quantitative image analysis. Curr. Biol. 2012, 22, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Matteoni, R.; Kreis, T.E. Translocation and clustering of endosomes and lysosomes depends on microtubules. J. Cell Biol. 1987, 105, 1253–1265. [Google Scholar] [CrossRef] [PubMed]
- Pangarkar, C.; Dinh, A.; Mitragotri, S. Endocytic pathway rapidly delivers internalized molecules to lysosomes: An analysis of vesicle trafficking, clustering and mass transfer. J. Controll. Release 2012, 162, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; DiBenedetto, J.R.; West, M.; Rowland, A.A.; Voeltz, G.K. Endoplasmic reticulum–endosome contact increases as endosomes traffic and mature. Mol. Biol. Cell 2013, 24, 1030–1040. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, D.; Cyphersmith, A.; Zapata, J.A.; Kim, Y.J.; Payne, C.K. Lysosome transport as a function of lysosome diameter. PLoS ONE 2014, 9, e86847. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.P.; Du, W.; Ji, Q.; Xue, B.; Jiang, D.; Zhu, Y.; Ren, H.; Zhang, C.; Lou, J.; Yu, L.; et al. Vesicle size regulates nanotube formation in the cell. Sci. Rep. 2016, 6, 24002. [Google Scholar] [CrossRef] [PubMed]
- Zádori, Z.; Szelei, J.; Lacoste, M.; Li, Y.; Gariépy, S.; Raymond, P.; Allaire, M.; Nabi, I.; Tijssen, P. A Viral phospholipase A2 is required for parvovirus infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef]
- Cudmore, S.; Cossart, P.; Griffiths, G.; Way, M. Actin-based motility of vaccinia virus. Nature 1995, 378, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, T.; Volkman, L.E.; Welch, M.D. Actin-based motility drives baculovirus transit to the nucleus and cell surface. J. Cell Biol. 2010, 190, 187. [Google Scholar] [CrossRef] [PubMed]
- Sodeik, B.; Ebersold, M.W.; Helenius, A. Microtubule-mediated transport of incoming herpes simplex virus 1 capsids to the nucleus. J. Cell Biol. 1997, 136, 1007–1021. [Google Scholar] [CrossRef] [PubMed]
- Brandenburg, B.; Zhuang, X. Virus trafficking-learning from single-virus tracking. Nat. Rev. Microbiol. 2007, 5, 197–208. [Google Scholar] [CrossRef] [PubMed]
- King, S.J.; Schroer, T.A. Dynactin increases the processivity of the cytoplasmic dynein motor. Nat. Cell Biol. 2000, 2, 20–24. [Google Scholar] [PubMed]
- Roberts, A.J.; Kon, T.; Knight, P.J.; Sutoh, K.; Burgess, S.A. Functions and mechanics of dynein motor proteins. Nat. Rev. Mol. Cell Biol. 2013, 14, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.S.; Wilcher, R.; Samulski, R.J. Infectious entry pathway of adeno-associated virus and adeno-associated virus vectors. J. Virol. 2000, 74, 2777–2785. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, E.; Ramírez, J.C.; Garcia, J.; Almendral, J.M. Complementary roles of multiple nuclear targeting signals in the capsid proteins of the parvovirus minute virus of mice during assembly and onset of infection. J. Virol. 2002, 76, 7049–7059. [Google Scholar] [CrossRef] [PubMed]
- Kelkar, S.; de, B.P.; Gao, G.; Wilson, J.M.; Crystal, R.G.; Leopold, P.L. A Common mechanism for cytoplasmic dynein-dependent microtubule binding shared among adeno-associated virus and adenovirus serotypes. J. Virol. 2006, 80, 7781–7785. [Google Scholar] [CrossRef] [PubMed]
- Suikkanen, S.; Aaltonen, T.; Nevalainen, M.; Välilehto, O.; Lindholm, L.; Vuento, M.; Vihinen-Ranta, M. Exploitation of microtubule cytoskeleton and dynein during parvoviral traffic toward the nucleus. J. Virol. 2003, 77, 10270–10279. [Google Scholar] [CrossRef] [PubMed]
- Mani, B.; Baltzer, C.; Valle, N.; Almendral, J.M.; Kempf, C.; Ros, C. Low pH-dependent endosomal processing of the incoming parvovirus minute virus of mice virion leads to externalization of the VP1 N-terminal sequence (N-VP1), N-VP2 cleavage, and uncoating of the full-length genome. J. Virol. 2006, 80, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, F.; Bleker, S.; Leuchs, B.; Fischer, R.; Kleinschmidt, J. Adeno-associated virus type 2 capsids with externalized VP1/VP2 trafficking domains are generated prior to passage through the cytoplasm and are maintained until uncoating occurs in the nucleus. J. Virol. 2006, 80, 11040–11054. [Google Scholar] [CrossRef] [PubMed]
- Bantel-Schaal, U.; Braspenning-Wesch, I.; Kartenbeck, J. Adeno-associated virus type 5 exploits two different entry pathways in human embryo fibroblasts. J. Gen. Virol. 2009, 90, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Grieger, J.C.; Snowdy, S.; Samulski, R.J. Separate basic region motifs within the adeno-associated virus capsid proteins are essential for infectivity and assembly. J. Virol. 2006, 80, 5199–5210. [Google Scholar] [CrossRef] [PubMed]
- Harbison, C.E.; Lyi, S.M.; Weichert, W.S.; Parrish, C.R. Early steps in cell infection by parvoviruses: Host-specific differences in cell receptor binding but similar endosomal trafficking. J. Virol. 2009, 83, 10504–10514. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kim, Y.J.; Ji, M.; Fang, J.; Siriwon, N.; Zhang, L.I.; Wang, P. Enhancing gene delivery of adeno-associated viruses by cell-permeable peptides. Mol. Ther. Methods Clin. Dev. 2014, 1, 12. [Google Scholar] [CrossRef] [PubMed]
- Kural, C.; Kim, H.; Syed, S.; Goshima, G.; Gelfand, V.I.; Selvin, P.R. Kinesin and dynein move a peroxisome in vivo: A tug-of-war or coordinated movement? Science 2005, 308, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Castle, M.J.; Perlson, E.; Holzbaur, E.L.F.; Wolfe, J.H. Long-distance axonal transport of AAV9 is driven by dynein and kinesin-2 and is trafficked in a highly motile Rab7-positive compartment. Mol. Ther. 2014, 22, 554–566. [Google Scholar] [CrossRef] [PubMed]
- Kartenbeck, J.; Stukenbrok, H.; Helenius, A. Endocytosis of simian virus 40 into the endoplasmic reticulum. J. Cell Biol. 1989, 109, 2721–2729. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Goryaynov, A.; Yang, W. Super-resolution 3D tomography of interactions and competition in the nuclear pore complex. Nat. Struct. Mol. Biol. 2016, 23, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Beck, M.; Hurt, E. The nuclear pore complex: Understanding its function through structural insight. Nat. Rev. Mol. Cell Biol. 2017, 18, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Alber, F.; Dokudovskaya, S.; Veenhoff, L.M.; Zhang, W.; Kipper, J.; Devos, D.; Suprapto, A.; Karni-Schmidt, O.; Williams, R.; Chait, B.T.; et al. The molecular architecture of the nuclear pore complex. Nature 2007, 450, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Cardarelli, F.; Lanzano, L.; Gratton, E. Capturing directed molecular motion in the nuclear pore complex of live cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9863–9868. [Google Scholar] [CrossRef] [PubMed]
- Panté, N.; Kann, M. Nuclear pore complex is able to transport macromolecules with diameters of ~39 nm. Mol. Biol. Cell 2002, 13, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Strawn, L.A.; Shen, T.; Shulga, N.; Goldfarb, D.S.; Wente, S.R. Minimal nuclear pore complexes define FG repeat domains essential for transport. Nat. Cell Biol. 2004, 6, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.; Görlich, D. A saturated FG-repeat hydrogel can reproduce the permeability properties of nuclear pore complexes. Cell 2007, 130, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.S.; Belmont, B.J.; Sante, J.M.; Rexach, M.F. Natively unfolded nucleoporins gate protein diffusion across the nuclear pore complex. Cell 2007, 129, 83–96. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, A.; van der Giessen, E.; Onck, P.R. Energetics of transport through the nuclear pore complex. PLoS ONE 2016, 11, e0148876. [Google Scholar] [CrossRef] [PubMed]
- Wente, S.R.; Rout, M.P. The nuclear pore complex and nuclear transport. Cold Spring Harb. Perspect. Biol. 2010, 2, a000562. [Google Scholar] [CrossRef] [PubMed]
- Popken, P.; Ghavami, A.; Onck, P.R.; Poolman, B.; Veenhoff, L.M. Size-dependent leak of soluble and membrane proteins through the yeast nuclear pore complex. Mol. Biol. Cell 2015, 26, 1386–1394. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M. Molecular mechanism of the nuclear protein import cycle. Nat. Rev. Mol. Cell Biol. 2007, 8, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Gelles, J.; Musser, S.M. Imaging of single-molecule translocation through nuclear pore complexes. Proc. Natl. Acad. Sci. USA 2004, 101, 12887–12892. [Google Scholar] [CrossRef] [PubMed]
- Ribbeck, K.; Görlich, D. Kinetic analysis of translocation through nuclear pore complexes. EMBO J. 2001, 20, 1320–1330. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Segura, L.M.; Lafarga, M.; Berciano, M.T.; Hernandez, P.; Andres, M.A. Distribution of nuclear pores and chromatin organization in neurons and glial cells of the rat cerebellar cortex. J. Comp. Neurol. 1989, 290, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Goldfarb, D.S.; Gariepy, J.; Schoolnik, G.; Kornberg, R.D. Synthetic peptides as nuclear localization signals. Nature 1986, 322, 641–644. [Google Scholar] [CrossRef] [PubMed]
- Ström, A.; Weiss, K. Importin-β-like nuclear transport receptors. Genome Biol. 2001, 2, reviews3008.1–reviews3008.9. [Google Scholar]
- Jans, D.A.; Ackermann, M.J.; Bischoff, J.R.; Beach, D.H.; Peters, R. p34cdc2-mediated phosphorylation at T124 inhibits nuclear import of SV-40 T antigen proteins. J. Cell Biol. 1991, 115, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Brown, K.; Siebenlist, U. Activation of NF-κB requires proteolysis of the inhibitor I κB-α: Signal-induced phosphorylation of I κB-α alone does not release active NF-κB. Proc. Natl. Acad. Sci. USA 1995, 92, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Christie, M.; Chang, C.; Róna, G.; Smith, K.M.; Stewart, A.G.; Takeda, A.A.S.; Fontes, M.R.M.; Stewart, M.; Vértessy, B.G.; Forwood, J.K.; et al. Structural biology and regulation of protein import into the nucleus. J. Mol. Biol. 2016, 428, 2060–2090. [Google Scholar] [CrossRef] [PubMed]
- Rexach, M.; Blobel, G. Protein import into nuclei: Association and dissociation reactions involving transport substrate, transport factors, and nucleoporins. Cell 1995, 83, 683–692. [Google Scholar] [CrossRef]
- Stewart, M.; Rhodes, D. Switching affinities in nuclear trafficking. Nat. Struct. Mol. Biol. 1999, 6, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Rux, J.J.; Burnett, R.M. Type-specific epitope locations revealed by X-ray crystallographic study of adenovirus type 5 hexon. Mol. Ther. 2000, 1, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Trotman, L.C.; Mosberger, N.; Fornerod, M.; Stidwill, R.P.; Greber, U.F. Import of adenovirus DNA involves the nuclear pore complex receptor CAN/Nup214 and histone H1. Nat. Cell Biol. 2001, 3, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Greber, U.F.; Willetts, M.; Webster, P.; Helenius, A. Stepwise dismantling of adenovirus 2 during entry into cells. Cell 1993, 75, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Strunze, S.; Engelke, M.F.; Wang, I.H.; Puntener, D.; Boucke, K.; Schleich, S.; Way, M.; Schoenenberger, P.; Burckhardt, C.J.; Greber, U.F. Kinesin-1-mediated capsid disassembly and disruption of the nuclear pore complex promote virus infection. Cell Host Microbe 2011, 10, 210–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saphire, A.C.S.; Guan, T.; Schirmer, E.C.; Nemerow, G.R.; Gerace, L. Nuclear import of adenovirus DNA in vitro involves the nuclear protein import pathway and HSC70. J. Biol. Chem. 2000, 275, 4298–4304. [Google Scholar] [CrossRef] [PubMed]
- Hindley, C.E.; Lawrence, F.J.; Matthews, D.A. A role for transportin in the nuclear import of adenovirus core proteins and DNA. Traffic 2007, 8, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Ojala, P.M.; Sodeik, B.; Ebersold, M.W.; Kutay, U.; Helenius, A. Herpes simplex virus type 1 entry into host cells: Reconstitution of capsid binding and uncoating at the nuclear pore complex in vitro. Mol. Cell. Biol. 2000, 20, 4922–4931. [Google Scholar] [CrossRef] [PubMed]
- Copeland, A.M.; Newcomb, W.W.; Brown, J.C. Herpes simplex virus replication: Roles of viral proteins and nucleoporins in capsid-nucleus attachment. J. Virol. 2009, 83, 1660–1668. [Google Scholar] [CrossRef] [PubMed]
- Pasdeloup, D.; Blondel, D.; Isidro, A.L.; Rixon, F.J. Herpesvirus capsid association with the nuclear pore complex and viral DNA release involve the nucleoporin CAN/Nup214 and the capsid protein pUL25. J. Virol. 2009, 83, 6610–6623. [Google Scholar] [CrossRef] [PubMed]
- Rout, M.P.; Aitchison, J.D.; Suprapto, A.; Hjertaas, K.; Zhao, Y.; Chait, B.T. The yeast nuclear pore complex. J. Cell Biol. 2000, 148, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Booy, F.P.; Newcomb, W.W.; Trus, B.L.; Brown, J.C.; Baker, T.S.; Steven, A.C. Liquid-crystalline, phage-like packing of encapsidated DNA in herpes simplex virus. Cell 1991, 64, 1007–1015. [Google Scholar] [CrossRef]
- Newcomb, W.W.; Cockrell, S.K.; Homa, F.L.; Brown, J.C. Polarized DNA ejection from the herpesvirus capsid. J. Mol. Biol. 2009, 392, 885–894. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Tikoo, S.K.; Babiuk, L.A. Nuclear localization of the ORF2 protein encoded by porcine circovirus type 2. Virology 2001, 285, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Kann, M.; Sodeik, B.; Vlachou, A.; Gerlich, W.H.; Helenius, A. Phosphorylation-dependent binding of hepatitis B virus core particles to the nuclear pore complex. J. Cell Biol. 1999, 145, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, J.C.; Pierson, E.E.; Keifer, D.Z.; Delaleau, M.; Gallucci, L.; Cazenave, C.; Kann, M.; Jarrold, M.F.; Zlotnick, A. Importin β can bind hepatitis B virus core protein and empty core-like particles and induce structural changes. PLoS Pathog. 2016, 12, e1005802. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, A.; Schwarz, A.; Foss, M.; Zhou, L.; Rabe, B.; Hoellenriegel, J.; Stoeber, M.; Panté, N.; Kann, M. Nucleoporin 153 arrests the nuclear import of hepatitis B virus capsids in the nuclear basket. PLoS Pathog. 2010, 6, e1000741. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chen, S.; Wang, M.; Cheng, A. The role of nuclear localization signal in parvovirus life cycle. Virol. J. 2017, 14, 80. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Kakkola, L.; Kalela, A.; Vilja, P.; Vuento, M. Characterization of a nuclear localization signal of canine parvovirus capsid proteins. Eur. J. Biochem. 1997, 250, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Vihinen-Ranta, M.; Wang, D.; Weichert, W.S.; Parrish, C.R. The VP1 N-terminal sequence of canine parvovirus affects nuclear transport of capsids and efficient cell infection. J. Virol. 2002, 76, 1884–1891. [Google Scholar] [CrossRef] [PubMed]
- Mäntylä, E.; Vihinen-Ranta, M. Analysis of human parvovirus capsid protein NLSs. unpublished.
- Seisenberger, G.; Ried, M.U.; Endress, T.; Büning, H.; Hallek, M.; Bräuchle, C. Real-time single-molecule imaging of the infection pathway of an adeno-associated virus. Science 2001, 294, 1929–1932. [Google Scholar] [CrossRef] [PubMed]
- Porwal, M.; Cohen, S.; Snoussi, K.; Popa-Wagner, R.; Anderson, F.; Dugot-Senant, N.; Wodrich, H.; Dinsart, C.; Kleinschmidt, J.A.; Panté, N.; et al. Parvoviruses cause nuclear envelope breakdown by activating key enzymes of mitosis. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Panté, N. Pushing the envelope: Microinjection of Minute virus of mice into Xenopus oocytes causes damage to the nuclear envelope. J. Gen. Virol. 2005, 86, 3243–3252. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Marr, A.K.; Garcin, P.; Panté, N. Nuclear envelope disruption involving host caspases plays a role in the parvovirus replication cycle. J. Virol. 2011, 85, 4863–4874. [Google Scholar] [CrossRef] [PubMed]
- Popa-Wagner, R.; Porwal, M.; Kann, M.; Reuss, M.; Weimer, M.; Florin, L.; Kleinschmidt, J.A. Impact of VP1-specific protein sequence motifs on adeno-associated virus type 2 Intracellular trafficking and nuclear entry. J. Virol. 2012, 86, 9163–9174. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ranedo, J.; Hernando, E.; Riolobos, L.; Domínguez, C.; Kann, M.; Almendral, J.M. The mammalian cell cycle regulates parvovirus nuclear capsid assembly. PLoS Pathog. 2015, 11, e1004920105. [Google Scholar] [CrossRef] [PubMed]
- Riolobos, L.; Valle, N.; Hernando, E.; Maroto, B.; Kann, M.; Almendral, J.M. Viral oncolysis that targets Raf-1 signaling control of nuclear transport. J. Virol. 2010, 84, 2090–2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Bu, W.; Bhatia, S.; Hare, J.; Somasundaram, T.; Azzi, A. The atomic structure of adeno-associated virus (AAV-2), a vector for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 10405–10410. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.S.; Ponnazhagan, S.; Srivastava, A. Rescue and replication of adeno-associated virus type 2 as well as vector DNA sequences from recombinant plasmids containing deletions in the viral inverted terminal repeats: Selective encapsidation of viral genomes in progeny virions. J. Virol. 1996, 70, 1668–1677. [Google Scholar] [PubMed]
- Ben-Asher, E.; Bratosin, S.; Aloni, Y. Intracellular DNA of the parvovirus minute virus of mice is organized in a minichromosome structure. J. Virol. 1982, 41, 1044–1054. [Google Scholar] [PubMed]
- Marcus-Sekura, C.; Carter, B.J. Chromatin-like structure of adeno-associated virus DNA in infected cells. J. Virol. 1983, 48, 79–87. [Google Scholar] [PubMed]
- Penaud-Budloo, M.; Le Guiner, C.; Nowrouzi, A.; Toromanoff, A.; Chérel, Y.; Chenuaud, P.; Schmidt, M.; von Kalle, C.; Rolling, F.; Moullier, P.; et al. Adeno-associated virus vector genomes persist as episomal chromatin in primate muscle. J. Virol. 2008, 82, 7875–7885. [Google Scholar] [CrossRef] [PubMed]
- Mäntylä, E.; Salokas, K.; Oittinen, M.; Aho, V.; Mäntysaari, P.; Palmujoki, L.; Kalliolinna, O.; Ihalainen, T.O.; Niskanen, E.A.; Timonen, J.; et al. Promoter-targeted histone acetylation of chromatinized parvoviral genome is essential for the progress of infection. J. Virol. 2016, 90, 4059–4066. [Google Scholar] [CrossRef] [PubMed]
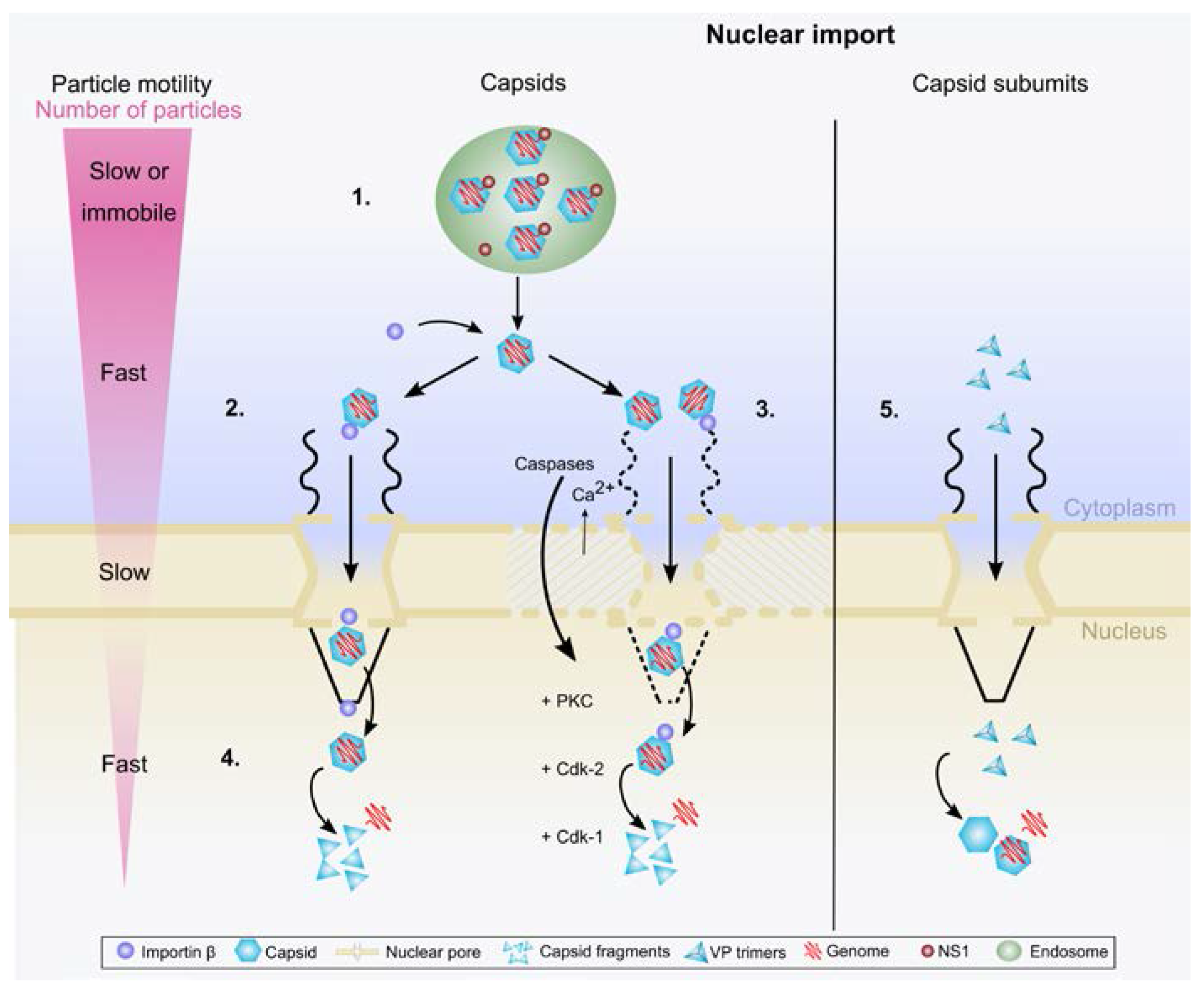
Figure 1.
Overview on the parvoviral nuclear import. (1) Viruses enter the cell by endocytosis. Inside the endosome viral NLS is exposed and the NS1 protein is detached from the capsid. Endosomal escape is followed by cytoplasmic interaction of capsid with importin β. In the cytoplasm viruses exploit dynein- and microtubule-mediated transport towards the nucleus; (2) after reaching the nuclear envelope, viruses are transported through the nuclear pore complex; (3) the direct or importin-mediated indirect interaction with the nuclear pores may also induce nuclear envelope breakdown allowing virus entry into the nucleoplasm. Host cell caspases, nuclear inflow of Ca2+, and activation of mitotic enzymes including Cdk-1/2 and PKC are involved in this process; (4) nuclear import is followed by viral capsid disassembly and release of their genome in the nucleoplasm; (5) in the late stage of infection, the newly synthesized viral capsid proteins are imported into the nucleus as phosphorylated trimeric assembly intermediates.
Figure 1.
Overview on the parvoviral nuclear import. (1) Viruses enter the cell by endocytosis. Inside the endosome viral NLS is exposed and the NS1 protein is detached from the capsid. Endosomal escape is followed by cytoplasmic interaction of capsid with importin β. In the cytoplasm viruses exploit dynein- and microtubule-mediated transport towards the nucleus; (2) after reaching the nuclear envelope, viruses are transported through the nuclear pore complex; (3) the direct or importin-mediated indirect interaction with the nuclear pores may also induce nuclear envelope breakdown allowing virus entry into the nucleoplasm. Host cell caspases, nuclear inflow of Ca2+, and activation of mitotic enzymes including Cdk-1/2 and PKC are involved in this process; (4) nuclear import is followed by viral capsid disassembly and release of their genome in the nucleoplasm; (5) in the late stage of infection, the newly synthesized viral capsid proteins are imported into the nucleus as phosphorylated trimeric assembly intermediates.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mäntylä, E.; Kann, M.; Vihinen-Ranta, M. Protoparvovirus Knocking at the Nuclear Door. Viruses 2017, 9, 286. https://doi.org/10.3390/v9100286
AMA Style
Mäntylä E, Kann M, Vihinen-Ranta M. Protoparvovirus Knocking at the Nuclear Door. Viruses. 2017; 9(10):286. https://doi.org/10.3390/v9100286
Chicago/Turabian StyleMäntylä, Elina, Michael Kann, and Maija Vihinen-Ranta. 2017. "Protoparvovirus Knocking at the Nuclear Door" Viruses 9, no. 10: 286. https://doi.org/10.3390/v9100286
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.