Characterization of Ovine A3Z1 Restriction Properties against Small Ruminant Lentiviruses (SRLVs)

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples and Cells
2.2. RNA Extraction and cDNA Synthesis
2.3. Viral Stocks
2.4. Amplification of Complete A3Z1/A3A and Sequence Analysis
2.5. Quantification of Small Ruminant APOBEC3 Expression
2.6. In Vitro A3Z1/A3Z1Tr Expression
2.7. Viral Infection
2.8. Immunoblot and Immunoprecipitation
2.9. Lentiviral Incorporation of A3Z1
2.10. Hypermutation
3. Results
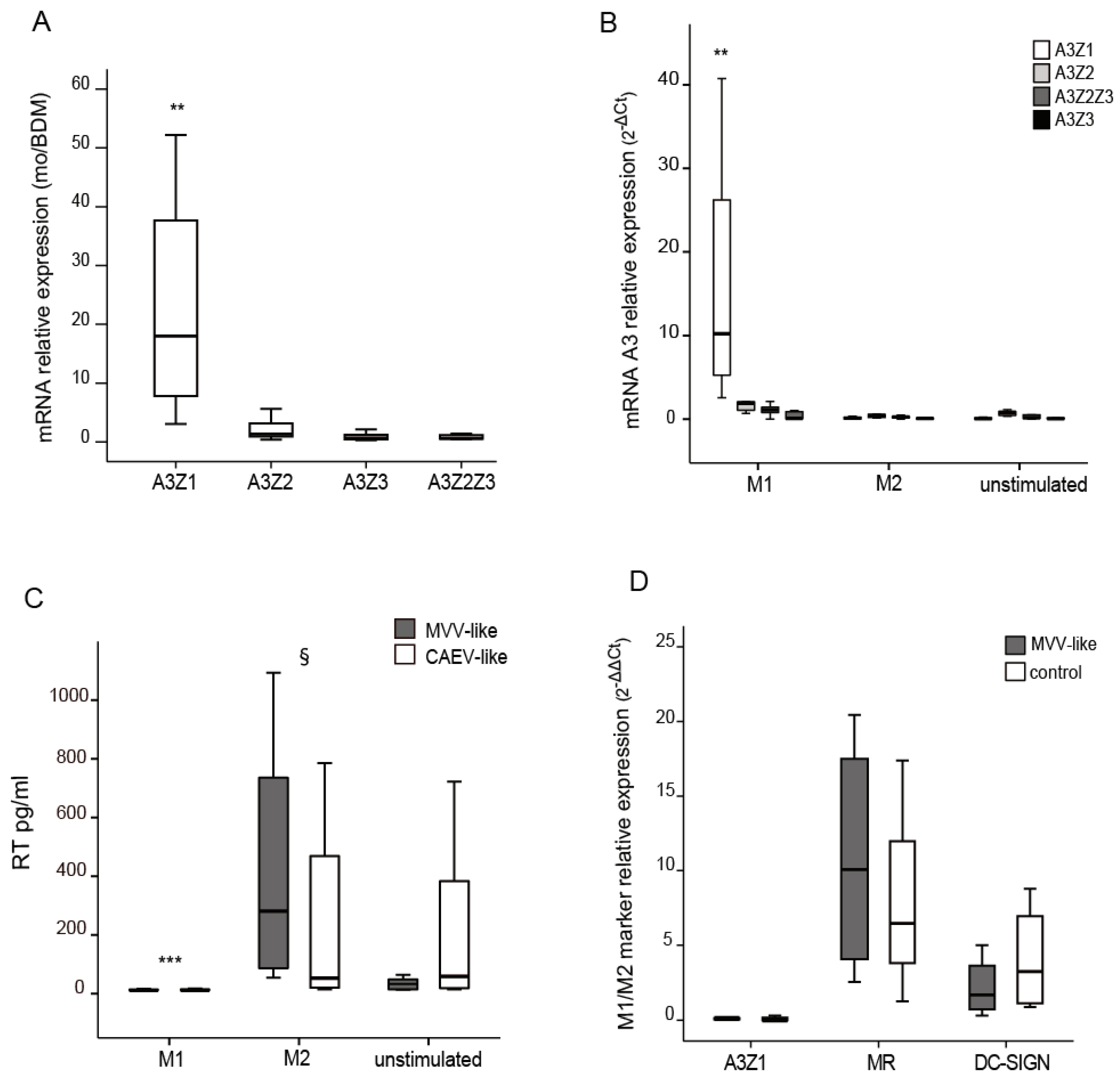
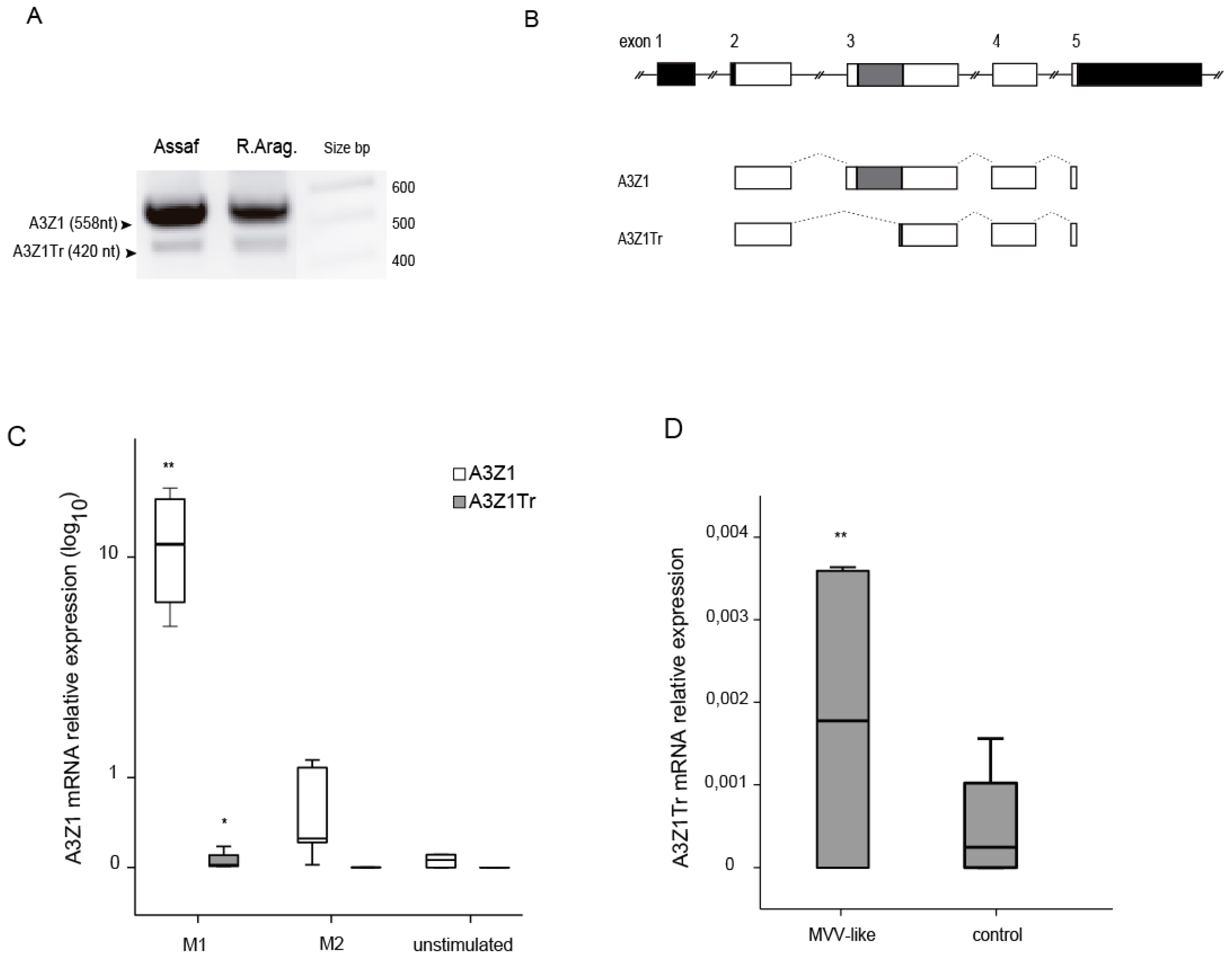
3.1. APOBEC3 Expression in Monocytes and Blood Monocyte-Derived Macrophages (BDM)
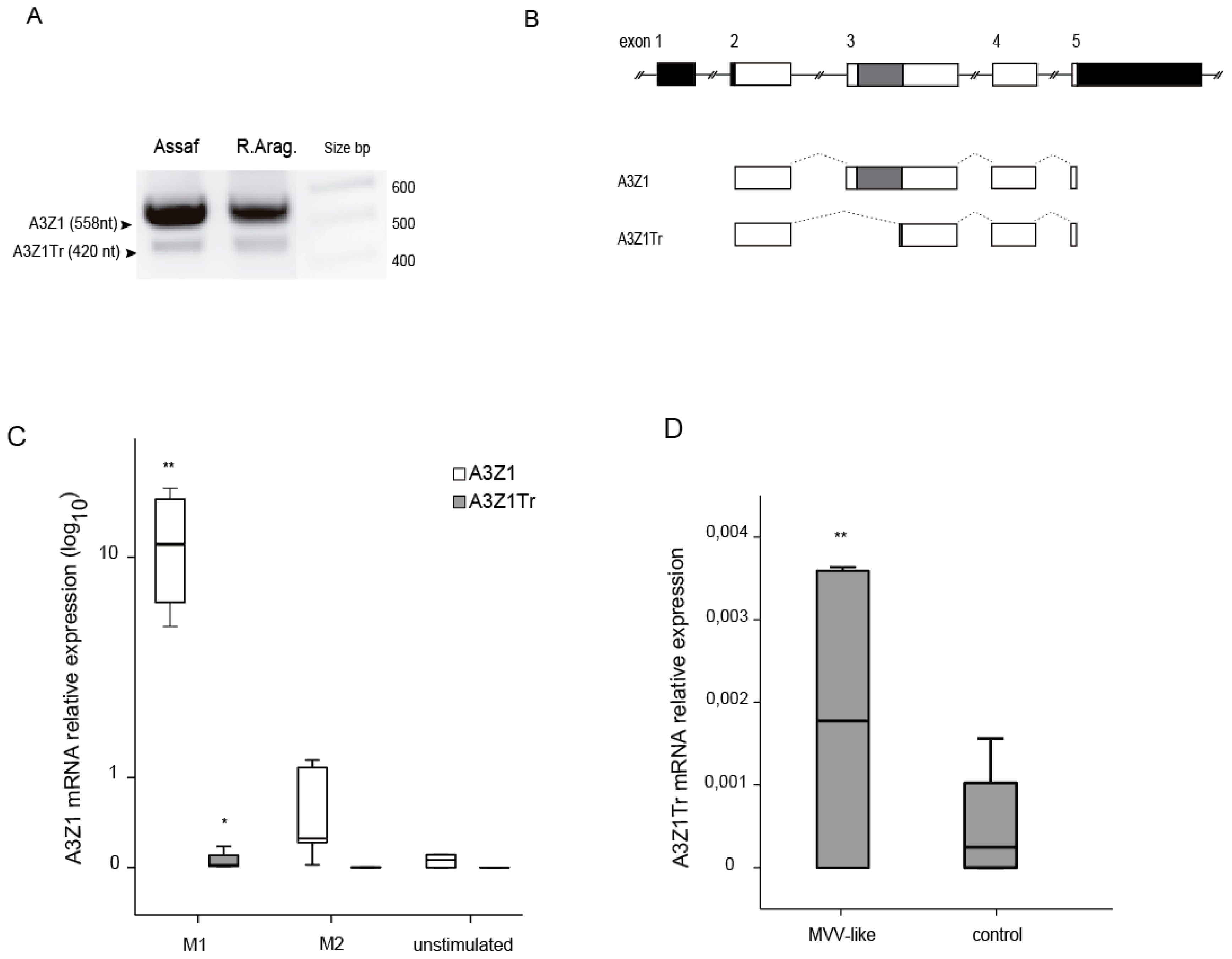
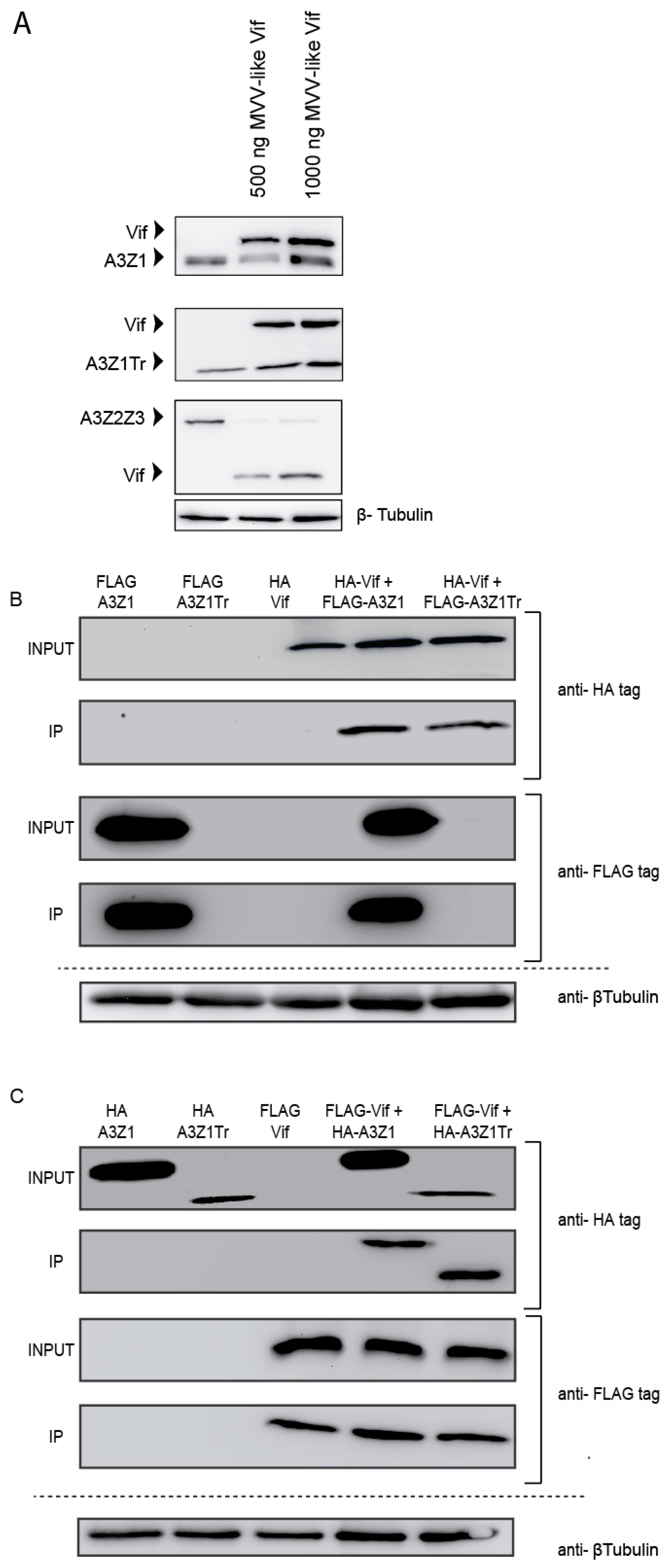
3.2. A3Z1-A3Z1Tr Interaction
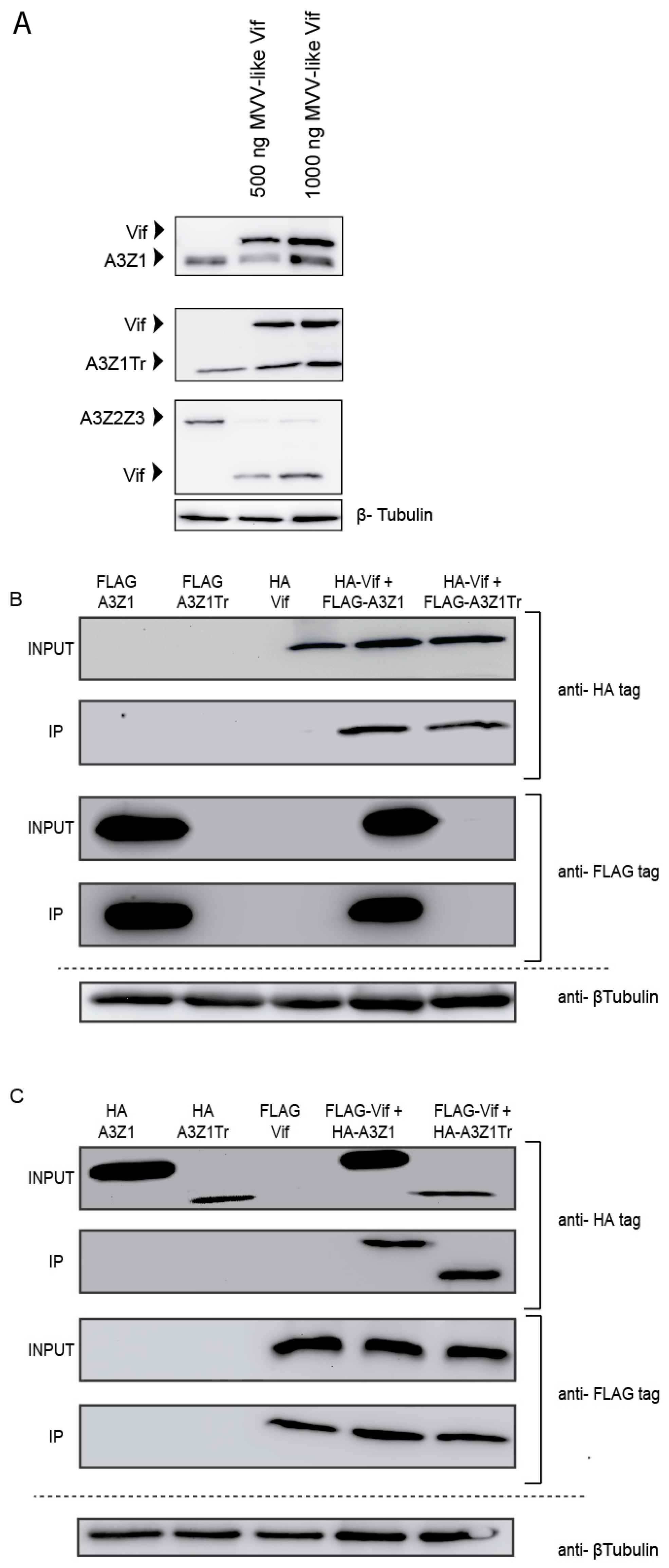
3.3. Incorporation into Viral Particles
3.4. Vif Degradation
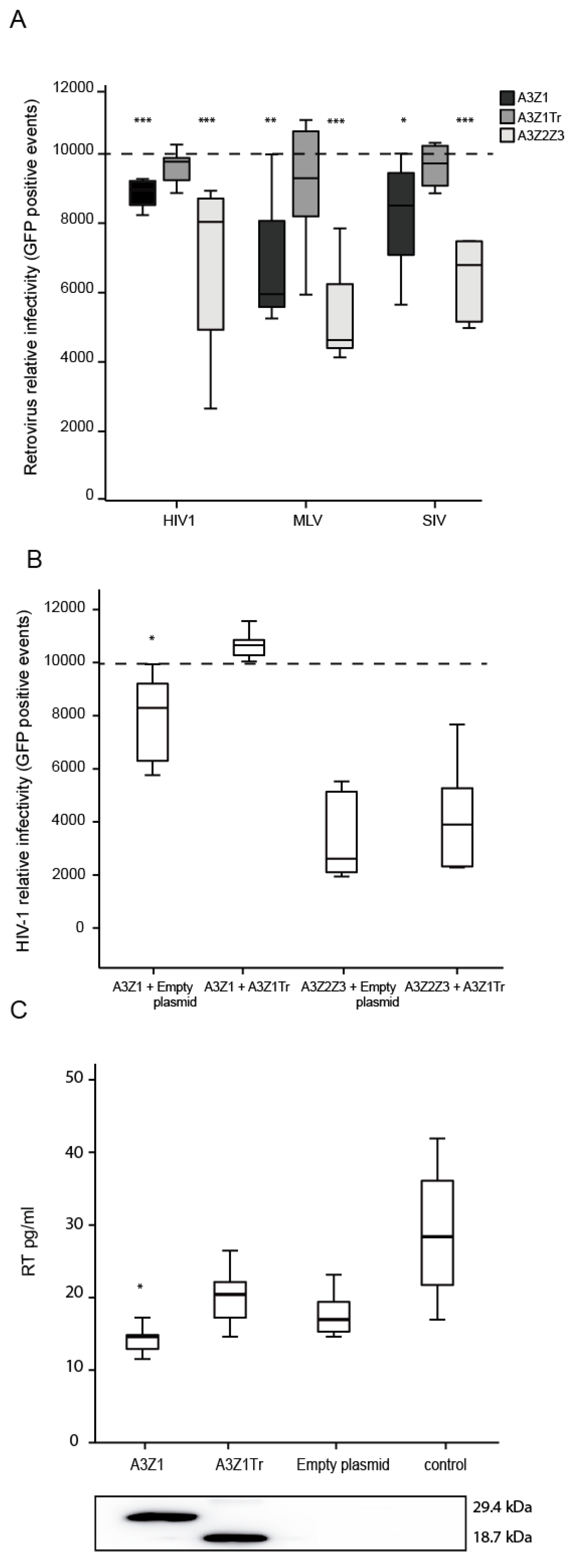
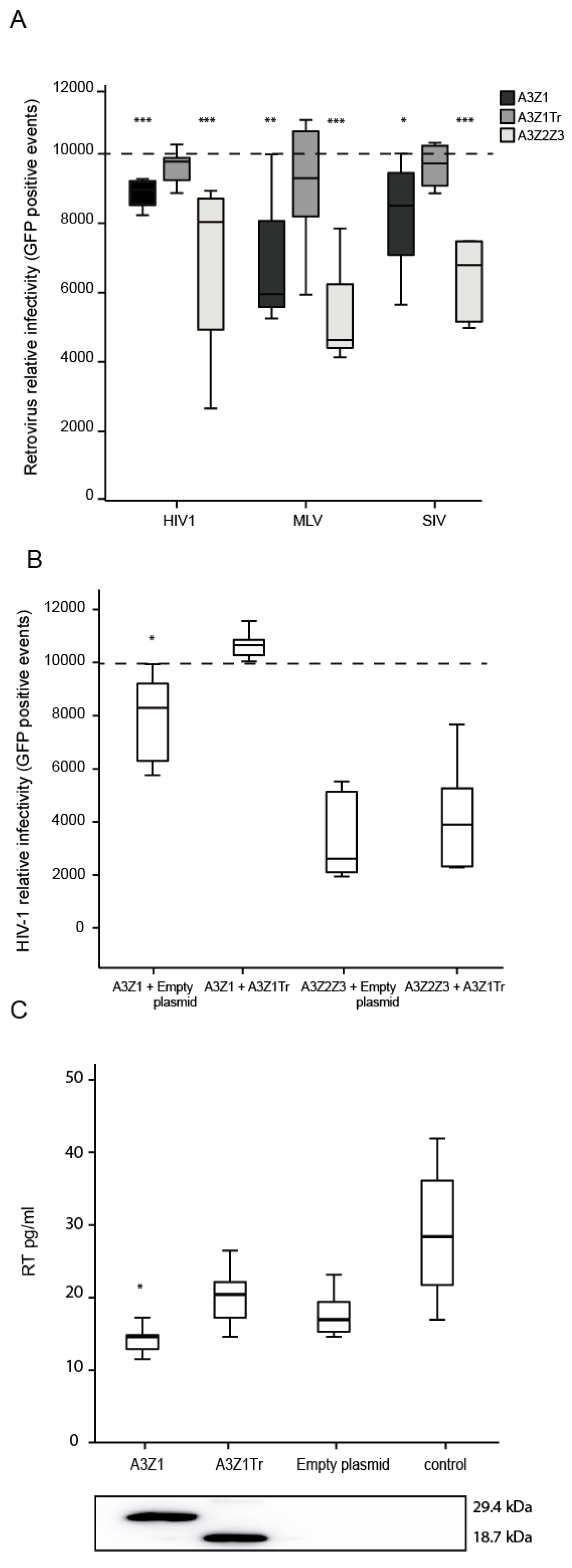
3.5. APOBEC3Z1 Restriction of Retrovirus Infection
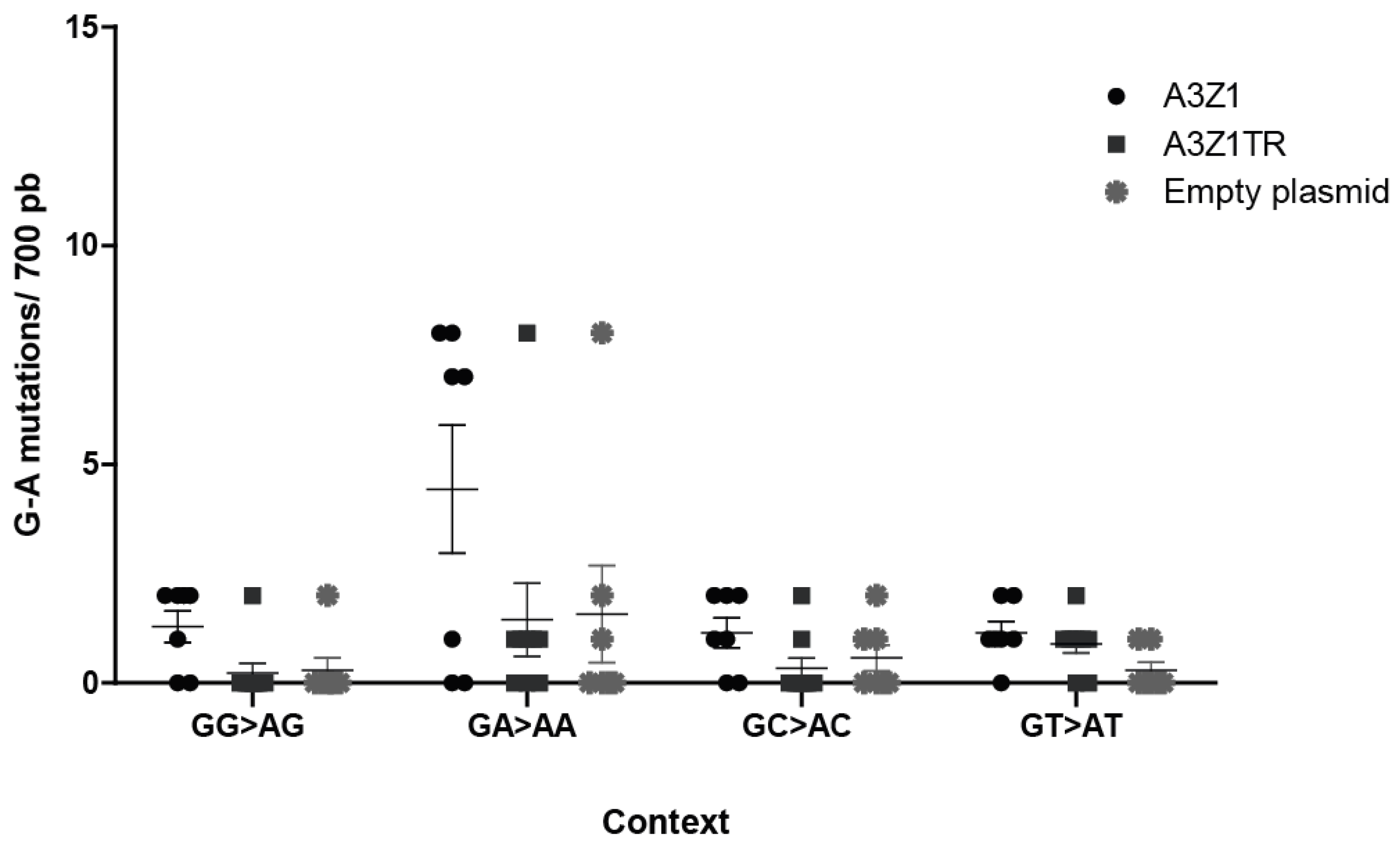
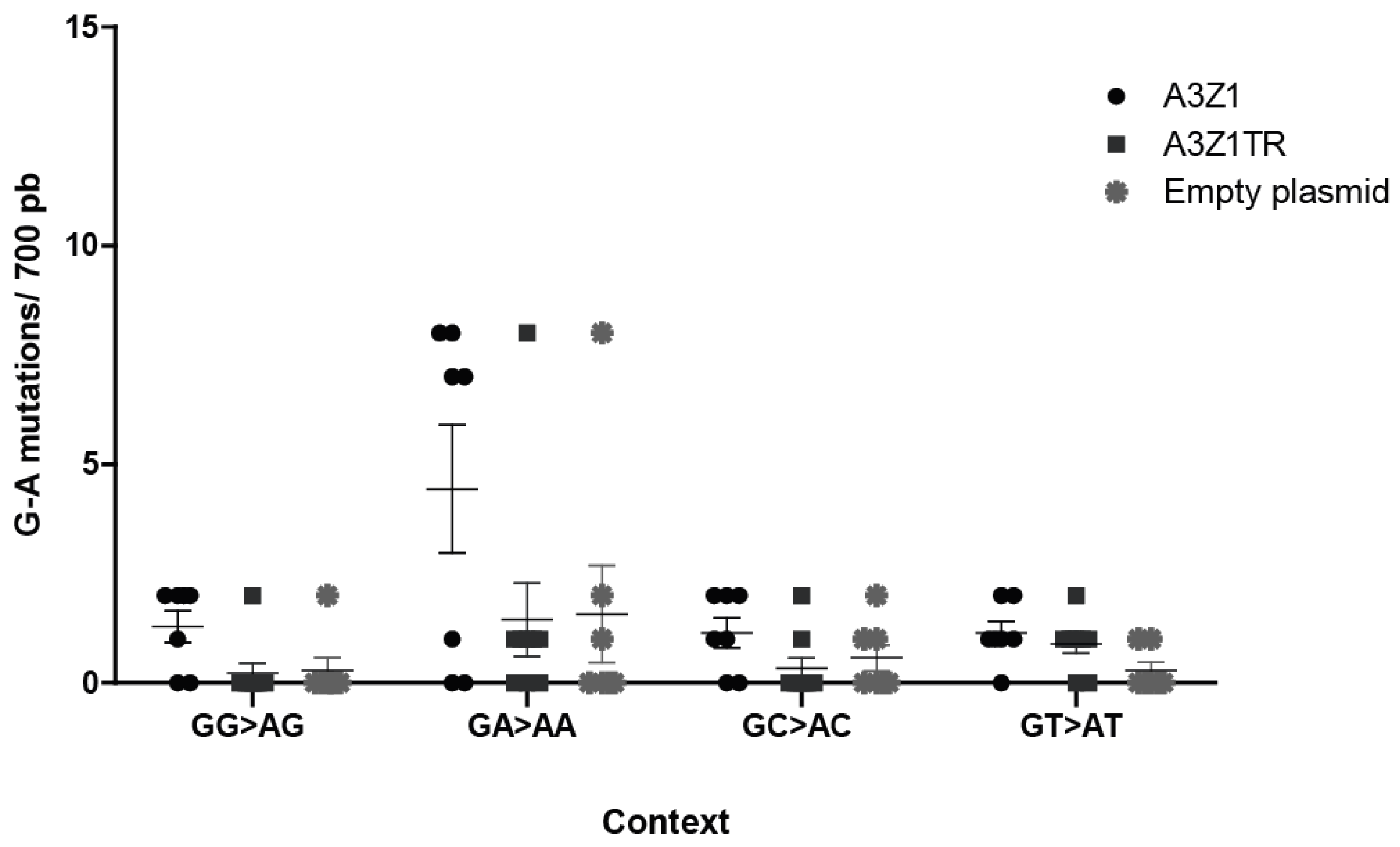
3.6. A3Z1 DNA Hypermutation Activity
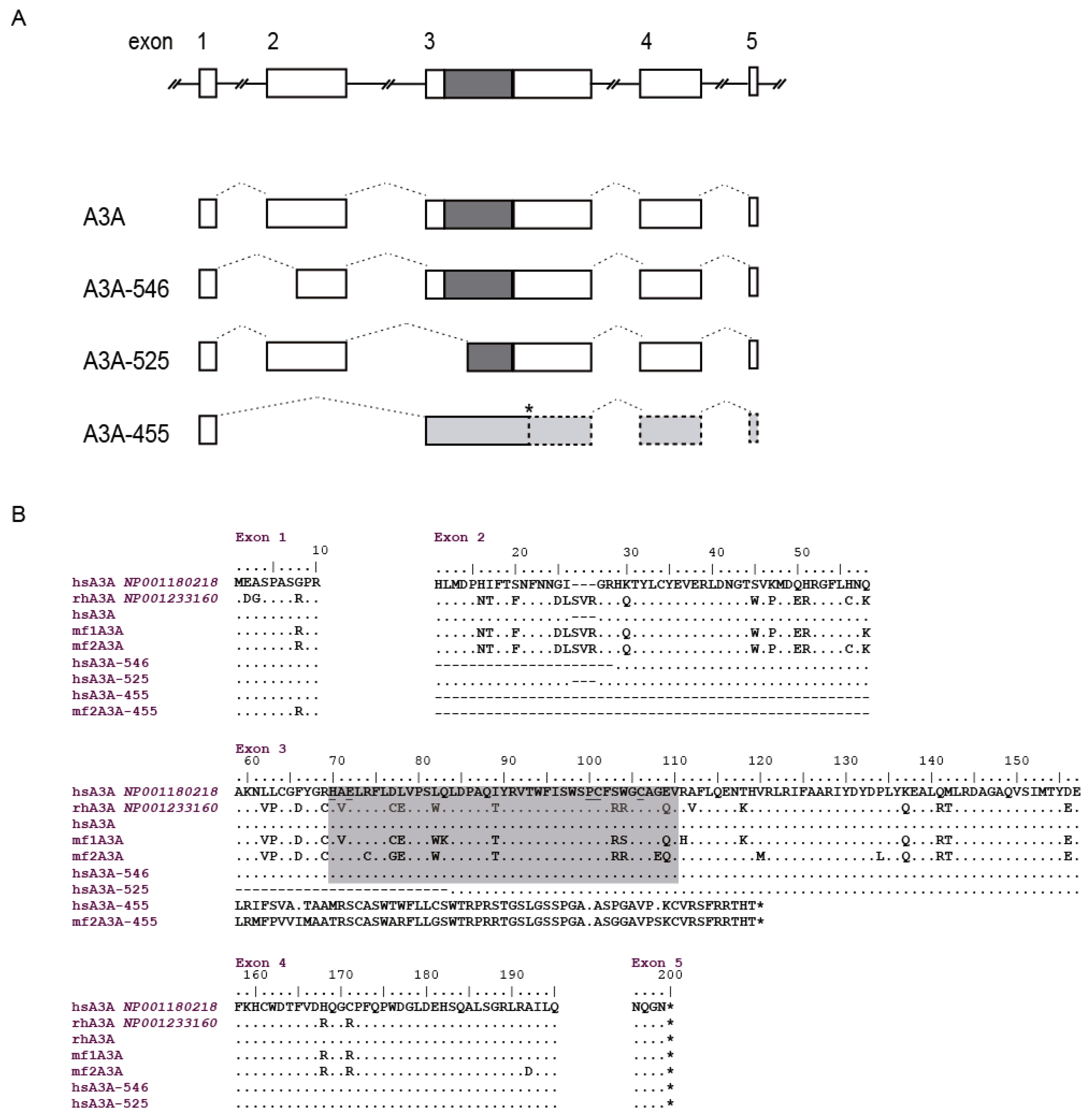
3.7. A3Z1/A3A Sequence Analysis
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harris, R.S.; Dudley, J.P. APOBECs and virus restriction. Virology 2015, 479–480, 131–145. [Google Scholar] [CrossRef] [PubMed]
- LaRue, R.S.; Andresdottir, V.; Blanchard, Y.; Conticello, S.G.; Derse, D.; Emerman, M.; Greene, W.C.; Jonsson, S.R.; Landau, N.R.; Lochelt, M.; et al. Guidelines for naming nonprimate APOBEC3 genes and proteins. J. Virol. 2009, 83, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.N.; Holmes, R.K.; Malim, M.H. Antiviral potency of APOBEC proteins does not correlate with cytidine deamination. J. Virol. 2006, 80, 8450–8458. [Google Scholar] [CrossRef] [PubMed]
- Narvaiza, I.; Linfesty, D.C.; Greener, B.N.; Hakata, Y.; Pintel, D.J.; Logue, E.; Landau, N.R.; Weitzman, M.D. Deaminase-independent inhibition of parvoviruses by the APOBEC3A cytidine deaminase. PLoS Pathog. 2009, 5, e1000439. [Google Scholar] [CrossRef] [PubMed]
- Stavrou, S.; Crawford, D.; Blouch, K.; Browne, E.P.; Kohli, R.M.; Ross, S.R. Different modes of retrovirus restriction by human APOBEC3A and APOBEC3G in vivo. PLoS Pathog. 2014, 10, e1004145. [Google Scholar] [CrossRef] [PubMed]
- Hultquist, J.F.; Lengyel, J.A.; Refsland, E.W.; LaRue, R.S.; Lackey, L.; Brown, W.L.; Harris, R.S. Human and rhesus APOBEC3D, APOBEC3F, APOBEC3G, and APOBEC3H demonstrate a conserved capacity to restrict Vif-deficient HIV-1. J. Virol. 2011, 85, 11220–11234. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Greenwell-Wild, T.; Nares, S.; Jin, W.; Lei, K.J.; Rangel, Z.G.; Munson, P.J.; Wahl, S.M. Myeloid differentiation and susceptibility to HIV-1 are linked to APOBEC3 expression. Blood 2007, 110, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Crespo, H.; Bertolotti, L.; Juganaru, M.; Glaria, I.; Andres, D.D.; Amorena, B.; Rosati, S.; Reina, R.; de Andres, D. Small ruminant macrophage polarization may play a pivotal role on lentiviral infection. Vet. Res. 2013, 44, 83. [Google Scholar] [CrossRef] [PubMed]
- Cassetta, L.; Kajaste-Rudnitski, A.; Coradin, T.; Saba, E.; Della Chiara, G.; Barbagallo, M.; Graziano, F.; Alfano, M.; Cassol, E.; Vicenzi, E.; et al. M1 polarization of human monocyte-derived macrophages restricts pre and postintegration steps of HIV-1 replication. Aids 2013, 27, 1847–1856. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Vuillermoz, S.; Mornex, J.F.; Greenland, T. Genomic heterogeneity in the pol region of ovine lentiviruses obtained from bronchoalveolar cells of infected sheep from France. J. Gen. Virol. 1995, 76, 1533–1537. [Google Scholar] [CrossRef] [PubMed]
- Leroux, C.; Chastang, J.; Greenland, T.; Mornex, J.F. Genomic heterogeneity of small ruminant lentiviruses: Existence of heterogeneous populations in sheep and of the same lentiviral genotypes in sheep and goats. Arch. Virol. 1997, 142, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Narayan, O.; Kennedy-Stoskopf, S.; Sheffer, D.; Griffin, D.E.; Clements, J.E. Activation of caprine arthritis-encephalitis virus expression during maturation of monocytes to macrophages. Infect. Immun. 1983, 41, 67–73. [Google Scholar] [PubMed]
- Pepin, M.; Vitu, C.; Russo, P.; Mornex, J.F.; Peterhans, E. Maedi-visna virus infection in sheep: A review. Vet. Res. 1998, 29, 341–367. [Google Scholar] [PubMed]
- Liu, B.; Sarkis, P.T.; Luo, K.; Yu, Y.; Yu, X.F. Regulation of APOBEC3F and human immunodeficiency virus type 1 Vif by Vif-Cul5-ElonB/C E3 ubiquitin ligase. J. Virol. 2005, 79, 9579–9587. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Rose, K.M.; Kozak, S.L.; Kabat, D. HIV-1 Vif protein binds the editing enzyme APOBEC3G and induces its degradation. Nat. Med. 2003, 9, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Sheehy, A.M.; Gaddis, N.C.; Malim, M.H. The antiretroviral enzyme APOBEC3G is degraded by the proteasome in response to HIV-1 Vif. Nat. Med. 2003, 9, 1404–1407. [Google Scholar] [CrossRef] [PubMed]
- Larue, R.S.; Lengyel, J.; Jonsson, S.R.; Andresdottir, V.; Harris, R.S. Lentiviral Vif degrades the APOBEC3Z3/APOBEC3H protein of its mammalian host and is capable of cross-species activity. J. Virol. 2010, 84, 8193–8201. [Google Scholar] [CrossRef] [PubMed]
- LaRue, R.S.; Jonsson, S.R.; Silverstein, K.A.; Lajoie, M.; Bertrand, D.; El-Mabrouk, N.; Hotzel, I.; Andresdottir, V.; Smith, T.P.; Harris, R.S. The artiodactyl APOBEC3 innate immune repertoire shows evidence for a multi-functional domain organization that existed in the ancestor of placental mammals. BMC Mol. Biol. 2008, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, S.R.; Hache, G.; Stenglein, M.D.; Fahrenkrug, S.C.; Andresdottir, V.; Harris, R.S. Evolutionarily conserved and non-conserved retrovirus restriction activities of artiodactyl APOBEC3F proteins. Nucleic Acids Res. 2006, 34, 5683–5694. [Google Scholar] [CrossRef] [PubMed]
- Sargan, D.R.; Bennet, I.D.; Cousens, C.; Roy, D.J.; Blacklaws, B.A.; Dalziel, R.G.; Watt, N.J.; McConnell, I. Nucleotide sequence of EV1, a British isolate of maedi-visna virus. J. Gen. Virol. 1991, 72, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
- Glaria, I.; Reina, R.; Crespo, H.; de Andres, X.; Ramirez, H.; Biescas, E.; Perez, M.M.; Badiola, J.; Lujan, L.; Amorena, B.; et al. Phylogenetic analysis of SRLV sequences from an arthritic sheep outbreak demonstrates the introduction of CAEV-like viruses among Spanish sheep. Vet. Microbiol. 2009, 138, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer 3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Saltarelli, M.; Querat, G.; Konings, D.A.; Vigne, R.; Clements, J.E. Nucleotide sequence and transcriptional analysis of molecular clones of CAEV which generate infectious virus. Virology 1990, 179, 347–364. [Google Scholar] [CrossRef]
- Hotzel, I.; Cheevers, W.P. Caprine arthritis-encephalitis virus envelope surface glycoprotein regions interacting with the transmembrane glycoprotein: Structural and functional parallels with human immunodeficiency virus type 1 GP120. J. Virol. 2003, 77, 11578–11587. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, R.S.; Lovsin, N.; Tanuri, A.; Peterlin, B.M. Vpr.A3A chimera inhibits HIV replication. J. Biol. Chem. 2008, 283, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.R.; Stanley, D.J.; Hultquist, J.F.; Johnson, J.R.; Mietrach, N.; Binning, J.M.; Jonsson, S.R.; Barelier, S.; Newton, B.W.; Johnson, T.L.; et al. Lineage-specific viral hijacking of non-canonical E3 ubiquitin ligase cofactors in the evolution of Vif anti-APOBEC3 activity. Cell Rep. 2015, 11, 1236–1250. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Zhu, D.; Wang, C.; Su, C.; Ma, J.; Ma, J.; Wang, X. Core-binding factor subunit beta is not required for non-primate lentiviral Vif-mediated APOBEC3 degradation. J. Virol. 2014, 88, 12112–12122. [Google Scholar] [CrossRef] [PubMed]
- Berger, G.; Durand, S.; Fargier, G.; Nguyen, X.N.; Cordeil, S.; Bouaziz, S.; Muriaux, D.; Darlix, J.L.; Cimarelli, A. APOBEC3A is a specific inhibitor of the early phases of HIV-1 infection in myeloid cells. PLoS Pathog. 2011, 7, e1002221. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Terzian, C.; Peeters, M.; Wain-Hobson, S.; Vartanian, J.P. Evolution of the primate APOBEC3A cytidine deaminase gene and identification of related coding regions. PLoS ONE 2012, 7, e30036. [Google Scholar] [CrossRef] [PubMed]
- Coffin, J.M.; Hughes, S.H.; Varmus, H.E. The Interactions of Retroviruses and their Hosts. In Retroviruses; Coffin, J.M., Hughes, S.H., Varmus, H.E., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1997. [Google Scholar]
- Blacklaws, B.A. Small ruminant lentiviruses: Immunopathogenesis of visna-maedi and caprine arthritis and encephalitis virus. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 259–269. [Google Scholar] [CrossRef] [PubMed]
- Morin, T.; Grezel, D.; Bouzar, B.; Villet, S. Experimental model of lentivirus reactivation for virological and immunological studies in caprine arthritis-encephalitis virus (CAEV) infected goats. Bull. Vet. Inst. Pulawy 2003, 47, 275–286. [Google Scholar]
- Jauregui, P.; Crespo, H.; Glaria, I.; Lujan, L.; Contreras, A.; Rosati, S.; de Andres, D.; Amorena, B.; Towers, G.J.; Reina, R. Ovine TRIM5α can restrict visna/maedi virus. J. Virol. 2012, 86, 9504–9509. [Google Scholar] [CrossRef] [PubMed]
- Gendelman, H.E.; Narayan, O.; Kennedy-Stoskopf, S.; Kennedy, P.G.; Ghotbi, Z.; Clements, J.E.; Stanley, J.; Pezeshkpour, G. Tropism of sheep lentiviruses for monocytes: Susceptibility to infection and virus gene expression increase during maturation of monocytes to macrophages. J. Virol. 1986, 58, 67–74. [Google Scholar] [PubMed]
- Trapp, S.; Derby, N.R.; Singer, R.; Shaw, A.; Williams, V.G.; Turville, S.G.; Bess, J.W., Jr.; Lifson, J.D.; Robbiani, M. Double-stranded RNA analog poly(I:C) inhibits human immunodeficiency virus amplification in dendritic cells via type I interferon-mediated activation of APOBEC3G. J. Virol. 2009, 83, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Malim, M.H. APOBEC proteins and intrinsic resistance to HIV-1 infection. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2009, 364, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Stenglein, M.D.; Burns, M.B.; Li, M.; Lengyel, J.; Harris, R.S. APOBEC3 proteins mediate the clearance of foreign DNA from human cells. Nat. Struct. Mol. Biol. 2010, 17, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Lassen, K.G.; Wissing, S.; Lobritz, M.A.; Santiago, M.; Greene, W.C. Identification of two APOBEC3F splice variants displaying HIV-1 antiviral activity and contrasting sensitivity to Vif. J. Biol. Chem. 2010, 285, 29326–29335. [Google Scholar] [CrossRef] [PubMed]
- Harari, A.; Ooms, M.; Mulder, L.C.; Simon, V. Polymorphisms and splice variants influence the antiretroviral activity of human APOBEC3H. J. Virol. 2009, 83, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Weiss, A. Negative regulation of CD45 by differential homodimerization of the alternatively spliced isoforms. Nat. Immunol. 2002, 3, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Burns, K.; Janssens, S.; Brissoni, B.; Olivos, N.; Beyaert, R.; Tschopp, J. Inhibition of interleukin 1 receptor/Toll-like receptor signaling through the alternatively spliced, short form of MyD88 is due to its failure to recruit IRAK-4. J. Exp. Med. 2003, 197, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Henriet, S.; Mercenne, G.; Bernacchi, S.; Paillart, J.C.; Marquet, R. Tumultuous relationship between the human immunodeficiency virus type 1 viral infectivity factor (Vif) and the human APOBEC-3G and APOBEC-3F restriction factors. Microbiol. Mol. Biol. Rev. 2009, 73, 211–232. [Google Scholar] [CrossRef] [PubMed]
- Sheehy, A.M.; Gaddis, N.C.; Choi, J.D.; Malim, M.H. Isolation of a human gene that inhibits HIV-1 infection and is suppressed by the viral Vif protein. Nature 2002, 418, 646–650. [Google Scholar] [CrossRef] [PubMed]
- Thielen, B.K.; McNevin, J.P.; McElrath, M.J.; Hunt, B.V.; Klein, K.C.; Lingappa, J.R. Innate immune signaling induces high levels of TC-specific deaminase activity in primary monocyte-derived cells through expression of APOBEC3A isoforms. J. Biol. Chem. 2010, 285, 27753–27766. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Localization | Amplicon Size (nt) | Reference |
|---|---|---|---|---|
| A3Z1-Out-Fw | AGGACTCGGAGCCAGGGACGA | Z1 3′-UTR | 621 | This work |
| A3Z1-Out-Rv | TCCTGCCATCTTAGAGAGGCTG | Z1 5′-UTR | ||
| A3A-Hs-Fw | ATGGAAGCCAGCCCAGCATC | A3A met | 600 | This work |
| A3A-Hs-Rv | TCAGTTTCCCTGATTCTGGAG | A3A stop | ||
| qA3Z1-Fw | TCCGTTCTTGGAATCTGGAC | Z1 CDS | 151 | [8] |
| qA3Z1-Rv | GTATAGATGCGGGAGGCAAA | Z1 CDS | ||
| qA3Z2-Fw | TTGAACCACCCTGTCTTTCC | Z2 3′-UTR | 188 | This work |
| qA3Z2-Rv | CAGGCTTCAGGGTTGTTGTT | Z2 3′-UTR | ||
| qA3Z3-Fw | GGGCGAGGAGATTGTGTTT | Z3 CDS | 182 | This work |
| qA3Z3-Rv | AAGCAGCCTTTGTCAAGCAT | Z3 CDS | ||
| qA3Z2-Z3-Fw | CAGGCCTTGGAAGAAACTGA | Z2 CDS | 151 | This work |
| qA3Z2-Z3-Rv | CCTCCGGTAGTAAGGTGGTG | Z3 CDS | ||
| qA3Z1Tr-Fw3 | GCTTTGTGCGCAACAAGAAA | Z1 CDS | 85 | This work |
| MR qPCR-Fw | TGGCAAATCCAGTTGTTAAGATGTT | MR CDS | ||
| MR qPCR-Rv | AGAATGTTGAATACTGTGGCGAGTT | MR CDS | 91 | [8] |
| DC-SIGN-Fw | GGTTCCGGAGTCTGACTGAAGTT | DC CDS | ||
| DC-SIGN-Rv | GGTCAGGCGCTGTAGGATCTC | DC CDS | 73 | [8] |
| β-actin-Fw | CTCACGGAGCGTGGCTACA | Actin CDS | ||
| β-actin-Rv | GCCATCTCCTGCTCGAAGTC | Actin CDS | 88 | [8] |
| A3Z1-EcoRI-Fw | TTTGAATTCGCCACCATGGATGAAAACACCTTCACTG | Z1 met | 588 | This work |
| A3Z1-XhoI-Rv | TTTCTCGAGCTACTATCAGTTTTGCTGAGCCCTGA | Z1 stop | ||
| LTR-Fw | TGACACAGCAAATGTAACCGCAA | LTR | 5126 | This work |
| SP33-Rv | CTTCCCCTTCCCAGAGTACCTGAG | POL |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Pablo-Maiso, L.; Glaria, I.; Crespo, H.; Nistal-Villán, E.; Andrésdóttir, V.; De Andrés, D.; Amorena, B.; Reina, R. Characterization of Ovine A3Z1 Restriction Properties against Small Ruminant Lentiviruses (SRLVs). Viruses 2017, 9, 345. https://doi.org/10.3390/v9110345
De Pablo-Maiso L, Glaria I, Crespo H, Nistal-Villán E, Andrésdóttir V, De Andrés D, Amorena B, Reina R. Characterization of Ovine A3Z1 Restriction Properties against Small Ruminant Lentiviruses (SRLVs). Viruses. 2017; 9(11):345. https://doi.org/10.3390/v9110345
Chicago/Turabian StyleDe Pablo-Maiso, Lorena, Idoia Glaria, Helena Crespo, Estanislao Nistal-Villán, Valgerdur Andrésdóttir, Damián De Andrés, Beatriz Amorena, and Ramsés Reina. 2017. "Characterization of Ovine A3Z1 Restriction Properties against Small Ruminant Lentiviruses (SRLVs)" Viruses 9, no. 11: 345. https://doi.org/10.3390/v9110345