
Goethite and Hematite Nanoparticles Show Promising Anti-Toxoplasma Properties

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Parasites
2.2.2. Cytotoxicity of Metal NPs in Mammalian Cells
2.2.3. In Vitro Growth Inhibition Assessment by Use of Luciferase Reporter Assays
2.2.4. Tryptophan-Coated Iron Oxide Nanoparticles
3. Results
3.1. Optimal Nanoparticle Concentration Assessment
3.2. Concentration-Dependent Growth Inhibition by Iron Oxide Nanoparticles
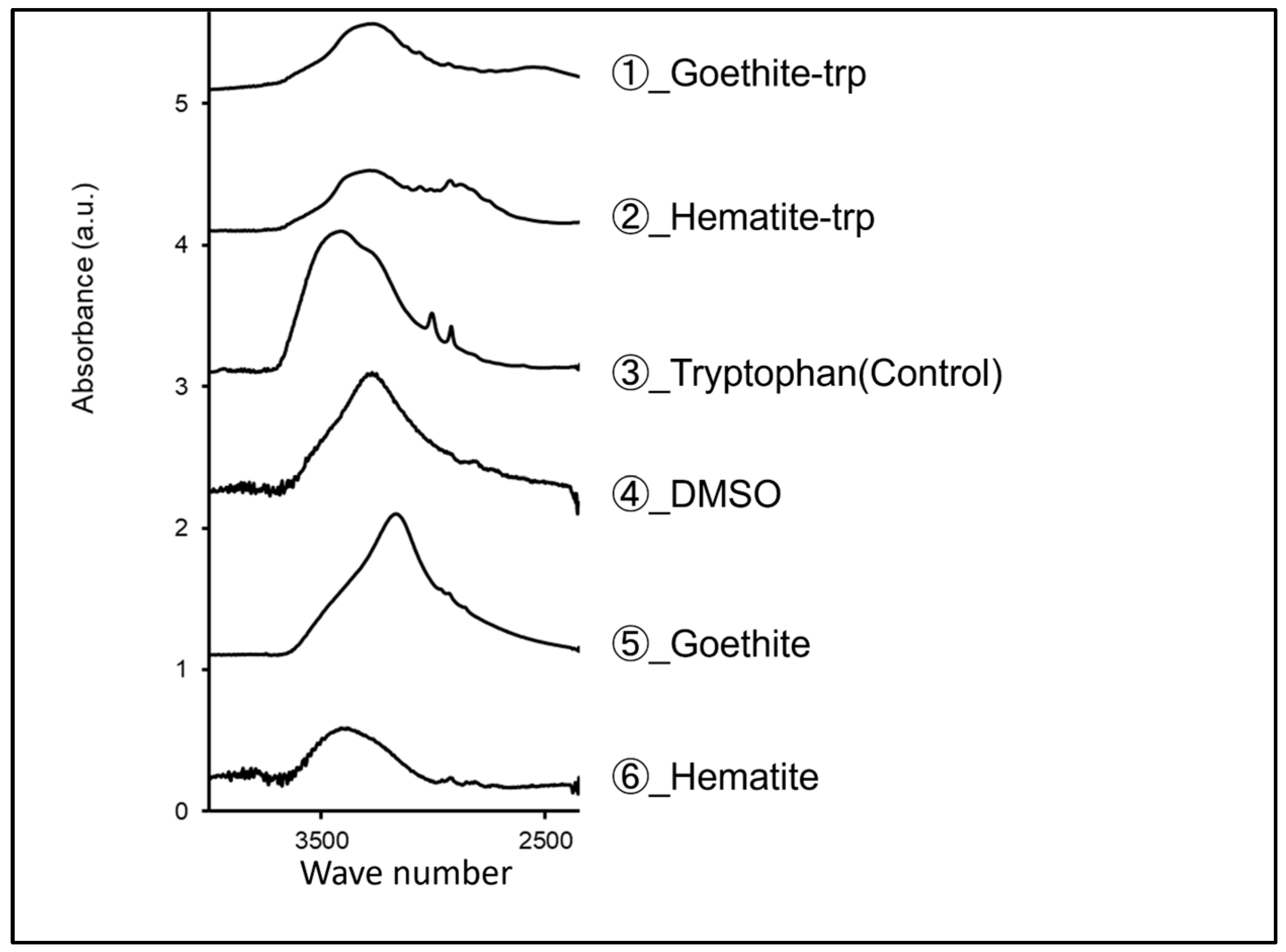
3.3. Surface Modification of Iron Oxide Nanoparticles with Tryptophan
3.4. Host Cytotoxicity Testing of Tryptophan-Coated Iron Oxide Nanoparticles
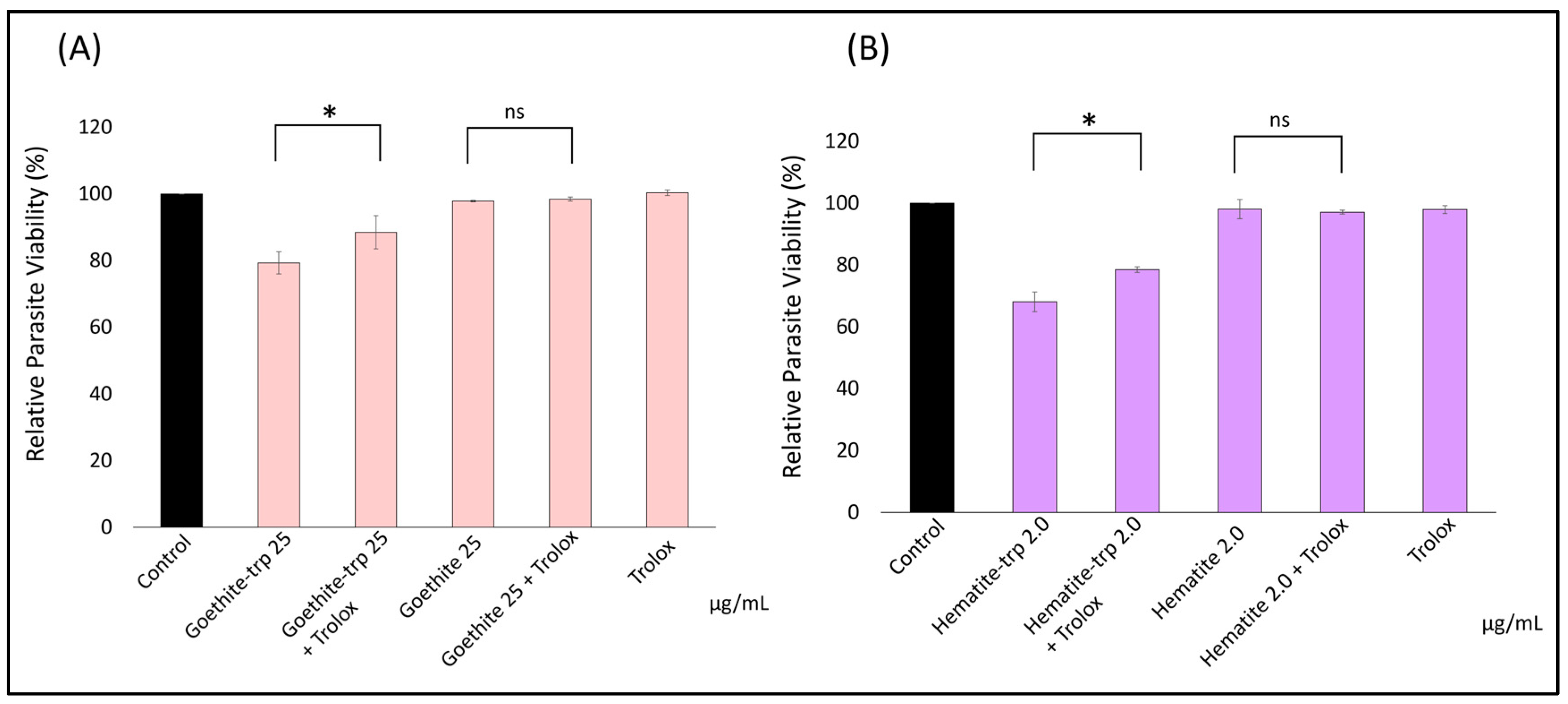
3.5. Tryptophan-Coated Iron Oxide Nanoparticles Increase the Growth Inhibitory Effect of the Nanoparticles on Toxoplasma
3.6. The Growth Inhibitory Effect of Iron Oxide Nanoparticles May Be Due to the Generation of Reactive Oxygen Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.-N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization Estimates of the Global and Regional Disease Burden of 11 Foodborne Parasitic Diseases, 2010: A Data Synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef]
- Black, M.W.; Boothroyd, J.C. Lytic cycle of Toxoplasma gondii. Microbiol. Mol. Biol. Rev. 2000, 64, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Beck, H.P.; Blake, D.; Dard, M.L.; Felger, I.; Pedraza-Diaz, S.; Regidor-Cerrillo, J.; Gomez-Bautista, M.; Ortega-Mora, L.M.; Putignani, L.; Shiels, B.; et al. Molecular approaches to diversity of populations of apicomplexan parasites. Int. J. Parasitol. 2009, 39, 175–189. [Google Scholar] [CrossRef]
- Zhang, Y.; Lai, B.S.; Juhas, M. Toxoplasma gondii secretory proteins and their role in invasion and pathogenesis. Microbiol. Res. 2019, 227, 126293. [Google Scholar] [CrossRef]
- Austeng, M.E.; Eskild, A.; Jacobsen, M.; Jenum, P.A.; Whitelaw, A.; Engdahl, B. Maternal infection with Toxoplasma gondii in pregnancy and the risk of hearing loss in the offspring. Int. J. Audiol. 2010, 49, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Garnaud, C.; Fricker-Hidalgo, H.; Evengard, B.; Alvarez-Martinez, M.J.; Petersen, E.; Kortbeek, L.M.; Robert-Gangneux, F.; Villena, I.; Costache, C.; Paul, M.; et al. Toxoplasma gondii-specific IgG avidity testing in pregnant women. Clin. Microbiol. Infect. 2020, 26, 1155–1160. [Google Scholar] [CrossRef] [PubMed]
- Dunn, D.; Wallon, M.; Peyron, F.; Petersen, E.; Peckham, C.; Gilbert, R. Mother-to-child transmission of toxoplasmosis: Risk estimates for clinical counselling. Lancet 1999, 353, 1829–1833. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.; Alvarez-Barrientos, A.; Sánchez, A.; Fernández, M.; Roncero, C.; Benito, M.; Fabregat, I. Reactive oxygen species (ROS) mediates the mitochondrial-dependent apoptosis induced by transforming growth factor ß in fetal hepatocytes. FASEB J. 2001, 15, 741–751. [Google Scholar] [CrossRef]
- Attias, M.; Teixeira, D.E.; Benchimol, M.; Vommaro, R.C.; Crepaldi, P.H.; De Souza, W. The life-cycle of Toxoplasma gondii reviewed using animations. Parasites Vectors 2020, 13, 588. [Google Scholar] [CrossRef] [PubMed]
- Curtis, A.; Wilkinson, C. Nantotechniques and approaches in biotechnology. Trends Biotechnol. 2001, 19, 97–101. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, Y.; Song, L.; Cui, X.; Zhou, J.; Jin, G.; Boccaccini, A.R.; Virtanen, S. Iron oxide nanoparticle-based nanocomposites in biomedical application. Trends Biotechnol. 2023, 41, 1471–1487. [Google Scholar] [CrossRef]
- Kamau, E.T.; Srinivasan, A.R.; Brown, M.J.; Fair, M.G.; Caraher, E.J.; Boyle, J.P. A focused small-molecule screen identifies 14 compounds with distinct effects on Toxoplasma gondii. Antimicrob. Agents Chemother. 2012, 56, 5581–5590. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Sulaiman, F.A. Evaluation of metal nanoparticles for drug delivery systems. J. Biomed. Res. 2015, 29, 145–149. [Google Scholar] [CrossRef] [PubMed]
- MubarakAli, D.; Thajuddin, N.; Jeganathan, K.; Gunasekaran, M. Plant extract mediated synthesis of silver and gold nanoparticles and its antibacterial activity against clinically isolated pathogens. Colloids Surf. B Biointerfaces 2011, 85, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Rios Coronado, P.E.; Longstaff, X.R.R.; Tarashansky, A.J.; Wang, B. Nanomedicine Approaches Against Parasitic Worm Infections. Adv. Healthc. Mater. 2018, 7, e1701494. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Molefe, N.I.; Awakan, O.J.; Nwonuma, C.O.; Alejolowo, O.O.; Olaolu, T.; Maimako, R.F.; Suganuma, K.; Han, Y.; Kato, K. Metal nanoparticles restrict the growth of protozoan parasites. Artif. Cells Nanomed. Biotechnol. 2018, 46, S86–S94. [Google Scholar] [CrossRef] [PubMed]
- Arisawa, M. Development of Metal Nanoparticle Catalysis toward Drug Discovery. Chem. Pharm. Bull. 2019, 67, 733–771. [Google Scholar] [CrossRef]
- Chen, X.; Ding, L.; Huang, X.; Xiong, Y. Tailoring noble metal nanoparticle designs to enable sensitive lateral flow immunoassay. Theranostics 2022, 12, 574–602. [Google Scholar] [CrossRef] [PubMed]
- Butkus, M.A.; Labare, M.P.; Starke, J.A.; Moon, K.; Talbot, M. Use of aqueous silver to enhance inactivation of coliphage MS-2 by UV disinfection. Appl. Environ. Microbiol. 2004, 70, 2848–2853. [Google Scholar] [CrossRef] [PubMed]
- Debbage, P. Targeted drugs and nanomedicine: Present and future. Curr. Pharm. Des. 2009, 15, 153–172. [Google Scholar] [CrossRef]
- Allahverdiyev, A.M.; Abamor, E.S.; Bagirova, M.; Ustundag, C.B.; Kaya, C.; Kaya, F.; Rafailovich, M. Antileishmanial effect of silver nanoparticles and their enhanced antiparasitic activity under ultraviolet light. Int. J. Nanomed. 2011, 6, 2705–2714. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.H.; Soliman, M.I.; Azzam, A.M.; Mostafa, A.B. Antiparasitic activity of silver and copper oxide nanoparticles against Entamoeba histolytica and Cryptosporidium parvum cysts. J. Egypt. Soc. Parasitol. 2015, 45, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Alphandery, E. Biodistribution and targeting properties of iron oxide nanoparticles for treatments of cancer and iron anemia disease. Nanotoxicology 2019, 13, 573–596. [Google Scholar] [CrossRef]
- Phillips-Howard, P.A.; West, L.J. Serious adverse drug reactions to pyrimethamine-sulphadoxine, pyrimethamine-dapsone and to amodiaquine in Britain. J. R. Soc. Med. 1990, 83, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Whiteley, C.G. Interaction of nanoparticles with arginine kinase from Trypanosoma brucei: Kinetic and mechanistic evaluation. Int. J. Biol. Macromol. 2013, 62, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, O.S.; Ishii, K.; Kato, K. L-tryptophan-titanium oxide nanoparticles showed selective anti-Toxoplasma gondii activity and improved host biocompatibility. Biomed. Pharmacother. 2023, 162, 114597. [Google Scholar] [CrossRef] [PubMed]
- Blader, I.J.; Koshy, A.A. Toxoplasma gondii development of its replicative niche: In its host cell and beyond. Eukaryot. Cell 2014, 13, 965–976. [Google Scholar] [CrossRef]
- Prescott, S.; Mutka, T.; Baumgartel, K.; Yoo, J.Y.; Morgan, H.; Postolache, T.T.; Seyfang, A.; Gostner, J.M.; Fuchs, D.; Kim, K.; et al. Tryptophan metabolism and immune alterations in pregnant Hispanic women with chronic Toxoplasma gondii infection. Am. J. Reprod. Immunol. 2023, 90, e13768. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, T.; Sharma, S.; Kumar, M.; Hussain, M.A.; Chauhan, N.; Kalia, I.; Sahu, A.K.; Rana, V.S.; Bharti, R.; Haldar, A.K.; et al. Tryptophan-Kynurenine Pathway Attenuates β-Catenin-Dependent pro-Parasitic Role of STING-TICAM2-IRF3-IDO1 Signalosome in Toxoplasma gondii Infection. Cell Death Dis. 2019, 10, 161. [Google Scholar] [CrossRef]
- Suryawanshi, R.; Patil, C.; Borase, H.; Salunke, I.; Elisabeth, M.; Philippe, G.; Patil, S. In vitro antiparasitic activity of microbial pigments and their combination with phytosynthesized metal nanoparticles. Parasitol. Int. 2015, 64, 353–356. [Google Scholar] [CrossRef]
- Adeyemi, O.S.; Murata, Y.; Sugi, T.; Han, Y.; Kato, K. Exploring Amino Acid-Capped Nanoparticles for Selective Anti-Parasitic Action and Improved Host Biocompatibility. J. Biomed. Nanotechnol. 2018, 14, 847–867. [Google Scholar] [CrossRef]
- Amaldoss, M.J.N.; Pandzic, E.; Koshy, P.; Kumar, N.; Sorrell, C.C.; Unnikrishnan, A. Detection and quantification of nanoparticle-induced intracellular ROS in live cells by laser scanning confocal microscopy. Methods 2022, 207, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Ikemura, K.; Inoue, K.; Mizutani, H.; Oka, H.; Iwamoto, T.; Okuda, M. An antioxidant Trolox restores decreased oral absorption of cyclosporine A after liver ischemia-reperfusion through distinct mechanisms between CYP3A and P-glycoprotein in the small intestine. Eur. J. Pharmacol. 2012, 690, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Schafer, Z.T.; Grassian, A.R.; Song, L.; Jiang, Z.; Gerhart-Hines, Z.; Irie, H.Y.; Gao, S.; Puigserver, P.; Brugge, J.S. Antioxidant and oncogene rescue of metabolic defects caused by loss of matrix attachment. Nature 2009, 461, 109–113. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishii, K.; Akahoshi, E.; Adeyemi, O.S.; Bando, H.; Fukuda, Y.; Ogawa, T.; Kato, K. Goethite and Hematite Nanoparticles Show Promising Anti-Toxoplasma Properties. Pharmaceutics 2024, 16, 413. https://doi.org/10.3390/pharmaceutics16030413
Ishii K, Akahoshi E, Adeyemi OS, Bando H, Fukuda Y, Ogawa T, Kato K. Goethite and Hematite Nanoparticles Show Promising Anti-Toxoplasma Properties. Pharmaceutics. 2024; 16(3):413. https://doi.org/10.3390/pharmaceutics16030413
Chicago/Turabian StyleIshii, Kosei, Eiji Akahoshi, Oluyomi Stephen Adeyemi, Hironori Bando, Yasuhiro Fukuda, Tomoyuki Ogawa, and Kentaro Kato. 2024. "Goethite and Hematite Nanoparticles Show Promising Anti-Toxoplasma Properties" Pharmaceutics 16, no. 3: 413. https://doi.org/10.3390/pharmaceutics16030413