
Unveiling the Potential of Extracellular Vesicles as Biomarkers and Therapeutic Nanotools for Gastrointestinal Diseases
 , , , , , and
, , , , , and 
Abstract
:1. Introduction
1.1. Prokaryotic Cell-Derived EVs
1.2. Eukaryotic Cell-Derived EVs
Mammalian Cells
1.3. Plant-Derived EVs
1.4. EVs for Therapeutic or Diagnostic Purposes
2. EVs as Mediators in Gastroenteric Pathogenesis and Biomarkers of GIDs
2.1. Aim of the Review
2.2. EVs in Liver Pathogenesis
2.3. EVs in Pancreas Pathogenesis
2.4. EVs in Gastric Pathogenesis
2.5. EVs in Intestinal Pathogenesis
3. Functional Gastrointestinal Diseases and EV-Based Therapy
3.1. EVs in Liver Disease Therapy
3.2. EVs in Gastritis and Esophagitis Therapy
3.3. EVs in Colitis Therapy
3.4. EVs in Pancreatitis Therapy

{kind=link}
{kind=link}
{kind=link}
| Organ | Pathology | Biotechnology EV-Based Preparation | Targets and/or Pathways | Bibliography |
|---|---|---|---|---|
| Liver | Fatty liver and inflammation | EVs derived from intestinal microbiota: reduction in adipose dysfunction and inflammation | TLR2 ↑ TLR4 ↓ | [89] |
| MASLD/ MASH | EVs derived from Huc-MSCs (combined with Nilotinib): Reduction in liver fibrosis, liver steatosis and inflammation; liver reparation and regeneration | FTO ↓ TGF-β1 ↓ ERK1/2 and Bcl-2 ↑ IKKB/NFkB/casp-9/-3 ↓ NLRP3 ↓ | [90,91,94] | |
| MASLD | Enrichment of oleic acid in exosomes: reduction in liver fibrosis in MASLD | elongase-6/RIP-1 ↑ | [44] | |
| Stomach | Esophagitis reflux | EVs released by BMSC:
| H3K27me3 | [99] |
| Gastritis | Exosomes derived from vaccine milk functionalized by PEG: stable new oral delivery system of si-RNA | ONCOGENES ↓ PD1/PD-L1 inhibitory axis ↓ | [100,102] | |
| Esophagitis fistula | Allogenic EVs from adipose tissue-derived stromal cells, co-administered with a thermo-responsive gel: new strategy in the therapy of esophagitis fistula | Epigenetic regulation | [103] | |
| Colon | Colitis | EVs released by ginger: improvement of colitis in a mice model | IL-22 ↑ IL-18 ↑ IL-10 ↑ | [106,107,108] |
| EVs derived from cow’s milk enriched with paclitaxel: improvement of pharmacokinetics and reduction in drug toxicity | MAPK ↑ | [112] | ||
| Pancreas | Pancreatitis | EVs released by MSCs:
| VEGF ↑ IL-6 ↓ TNF-α ↓ BAX ↓ BCL-2 ↑ NF-kB in Nucleoprotein ↓ NF-kB in plasmaprotein ↑ NLRP3 ↓ | [116,117,120,122,123] |
4. Structural Gastroenteric Disease and EV-Based Therapy
4.1. Inflammatory Bowel Diseases
4.2. Gastrointestinal Cancers
4.2.1. EVs in HCC therapy
4.2.2. EVs in Pancreatic Cancer Therapy
4.2.3. EVs in GC Therapy
4.2.4. EVs in CRC Therapy
5. Clinical Trials
6. Conclusions and Perspective
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Lee, K.S.; Kim, E.S. Explainable Artificial Intelligence in the Early Diagnosis of Gastrointestinal Disease. Diagnostics 2022, 12, 2740. [Google Scholar] [CrossRef] [PubMed]
- Milivojevic, V.; Milosavljevic, T. Burden of Gastroduodenal Diseases from the Global Perspective. Curr. Treat Options Gastroenterol. 2020, 18, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Mignini, I.; Piccirilli, G.; Termite, F.; Paratore, M.; Esposto, G.; Laterza, L.; Scaldaferri, F.; Ainora, M.E.; Gasbarrini, A.; Zocco, M.A. Extracellular Vesicles: Novel Potential Therapeutic Agents in Inflammatory Bowel Diseases. Cells 2024, 13, 90. [Google Scholar] [CrossRef]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef]
- Liao, W.; Du, Y.; Zhang, C.; Pan, F.; Yao, Y.; Zhang, T.; Peng, Q. Exosomes: The next Generation of Endogenous Nanomaterials for Advanced Drug Delivery and Therapy. Acta Biomater 2019, 86, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.Q.; Chng, W.H.; Liang, J.; Yeo, H.Q.; Lee, C.K.; Belaid, M.; Tollemeto, M.; Wacker, M.G.; Czarny, B.; Pastorin, G. Plant-Derived Extracellular Vesicles: Recent Advancements and Current Challenges on Their Use for Biomedical Applications. J. Extracell. Vesicles 2022, 11, e12283. [Google Scholar] [CrossRef]
- Diaz-Garrido, N.; Cordero, C.; Olivo-Martinez, Y.; Badia, J.; Baldomà, L. Cell-to-Cell Communication by Host-Released Extracellular Vesicles in the Gut: Implications in Health and Disease. Int. J. Mol. Sci. 2021, 22, 2213. [Google Scholar] [CrossRef]
- Fang, Y.; Wang, Z.; Liu, X.; Tyler, B.M. Biogenesis and Biological Functions of Extracellular Vesicles in Cellular and Organismal Communication with Microbes. Front. Microbiol. 2022, 13, 817844. [Google Scholar] [CrossRef] [PubMed]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; Degeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-Syntenin-ALIX Regulates the Biogenesis of Exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, J.; Kadungure, T.; Beyene, J.; Zhang, H.; Lu, Q. ARMMs as a Versatile Platform for Intracellular Delivery of Macromolecules. Nat. Commun. 2018, 9, 960. [Google Scholar] [CrossRef]
- Fordjour, F.K.; Guo, C.; Ai, Y.; Daaboul, G.G.; Gould, S.J. A Shared, Stochastic Pathway Mediates Exosome Protein Budding along Plasma and Endosome Membranes. J. Biol. Chem. 2022, 298, 102394. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, M.; Névo, N.; Jouve, M.; Valenzuela, J.I.; Maurin, M.; Verweij, F.J.; Palmulli, R.; Lankar, D.; Dingli, F.; Loew, D.; et al. Specificities of Exosome versus Small Ectosome Secretion Revealed by Live Intracellular Tracking of CD63 and CD9. Nat. Commun. 2021, 12, 4389. [Google Scholar] [CrossRef]
- Matsui, T.; Osaki, F.; Hiragi, S.; Sakamaki, Y.; Fukuda, M. ALIX and Ceramide Differentially Control Polarized Small Extracellular Vesicle Release from Epithelial Cells. EMBO Rep. 2021, 22, e51475. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Zhang, Q.; Franklin, J.L.; Coffey, R.J. Extracellular Vesicles and Nanoparticles: Emerging Complexities. Trends Cell Biol. 2023, 33, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Bawazir, M.; Dhall, A.; Kim, H.E.; He, L.; Heo, J.; Hwang, G. Implication of Surface Properties, Bacterial Motility, and Hydrodynamic Conditions on Bacterial Surface Sensing and Their Initial Adhesion. Front. Bioeng. Biotechnol. 2021, 9, 643722. [Google Scholar] [CrossRef] [PubMed]
- De la Canal, L.; Pinedo, M. Extracellular Vesicles: A Missing Component in Plant Cell Wall Remodeling. J. Exp. Bot. 2018, 69, 4655–4658. [Google Scholar] [CrossRef]
- Scavo, M.P.; Rizzi, F.; Depalo, N.; Armentano, R.; Coletta, S.; Serino, G.; Fanizza, E.; Pesole, P.L.; Cervellera, A.; Carella, N.; et al. Exosome Released FZD10 Increases Ki-67 Expression via Phospho-ERK1/2 in Colorectal and Gastric Cancer. Front. Oncol. 2021, 11, 730093. [Google Scholar] [CrossRef]
- Scavo, M.P.; Rizzi, F.; Depalo, N.; Fanizza, E.; Ingrosso, C.; Curri, M.L.; Giannelli, G. A Possible Role of Fzd10 Delivering Exosomes Derived from Colon Cancers Cell Lines in Inducing Activation of Epithelial–Mesenchymal Transition in Normal Colon Epithelial Cell Line. Int. J. Mol. Sci. 2020, 21, 6705. [Google Scholar] [CrossRef]
- Scavo, M.P.; Cigliano, A.; Depalo, N.; Fanizza, E.; Bianco, M.G.; Denora, N.; Laquintana, V.; Curri, M.L.; Lorusso, D.; Lotesoriere, C.; et al. Erratum: Frizzled-10 Extracellular Vesicles Plasma Concentration Is Associated with Tumoral Progression in Patients with Colorectal and Gastric Cancer. J Oncol 2020, 2020, 2715968. [Google Scholar] [CrossRef]
- Zeng, Y.; Qiu, Y.; Jiang, W.; Shen, J.; Yao, X.; He, X.; Li, L.; Fu, B.; Liu, X. Biological Features of Extracellular Vesicles and Challenges. Front. Cell Dev. Biol. 2022, 10, 816698. [Google Scholar] [CrossRef] [PubMed]
- Aljabali, A.A.; Obeid, M.A.; Bashatwah, R.M.; Serrano-Aroca, Á.; Mishra, V.; Mishra, Y.; El-Tanani, M.; Hromić-Jahjefendić, A.; Kapoor, D.N.; Goyal, R.; et al. Nanomaterials and Their Impact on the Immune System. Int. J. Mol. Sci. 2023, 24, 2008. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Guan, Y.; Xie, A.; Yan, Z.; Gao, S.; Li, W.; Rao, L.; Chen, X.; Chen, T. Extracellular Vesicles: A Rising Star for Therapeutics and Drug Delivery. J. Nanobiotechnol. 2023, 21, 1–51. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Jin, X.; Chen, S.; Yang, N.; Feng, G. Plant-Derived Extracellular Vesicles—A Novel Clinical Anti-Inflammatory Drug Carrier Worthy of Investigation. Biomed. Pharmacother. 2023, 169, 115904. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Hao, H.; Liu, Q.; Li, J.; Yao, Y.; Liu, Y.; Zhang, T.; Zhang, Z.; Yi, H. Effect of Extracelluar Vesicles Derived from Akkermansia Muciniphila on Intestinal Barrier in Colitis Mice. Nutrients 2023, 15, 4722. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Zhong, Y.; Xie, J.; Wang, Z.; Zhang, W.; Pi, Y.; Zhang, W.; Liu, L.; Luo, J.; Xu, W. Bacteroides Acidifaciens and Its Derived Extracellular Vesicles Improve DSS-Induced Colitis. Front. Microbiol. 2023, 14, 1304232. [Google Scholar] [CrossRef] [PubMed]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Bovine Milk-Derived Exosomes for Drug Delivery. Cancer Lett. 2016, 371, 48–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, D.; Ye, Z.; Xu, J. Engineering Extracellular Vesicles as Delivery Systems in Therapeutic Applications. Adv. Sci. 2023, 10, e2300552. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wu, D.; Ma, X.; Wang, J.; Hou, W.; Zhang, W. Exosomes as Drug Carriers for Cancer Therapy and Challenges Regarding Exosome Uptake. Biomed. Pharmacother. 2020, 128, 110237. [Google Scholar] [CrossRef]
- Zheng, Y.; Hasan, A.; Nejadi Babadaei, M.M.; Behzadi, E.; Nouri, M.; Sharifi, M.; Falahati, M. Exosomes: Multiple-Targeted Multifunctional Biological Nanoparticles in the Diagnosis, Drug Delivery, and Imaging of Cancer Cells. Biomed. Pharmacother. 2020, 129, 110442. [Google Scholar] [CrossRef]
- Longatti, A.; Schindler, C.; Collinson, A.; Jenkinson, L.; Matthews, C.; Fitzpatrick, L.; Blundy, M.; Minter, R.; Vaughan, T.; Shaw, M.; et al. High Affinity Single-Chain Variable Fragments Are Specific and Versatile Targeting Motifs for Extracellular Vesicles. Nanoscale 2018, 10, 14230–14244. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, M.; Sung, J.; Wang, L.; Jung, Y.; Merlin, D. Autologous Exosome Transfer: A New Personalised Treatment Concept to Prevent Colitis in a Murine Model. J. Crohns Colitis 2020, 14, 841–855. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, M.; Villa, A.; Crescenti, D.; Marzagalli, M.; Kuryk, L.; Limonta, P.; Mazzaferro, V.; Ciana, P. Heterologous and Cross-Species Tropism of Cancer-Derived Extracellular Vesicles. Theranostics 2019, 9, 5681–5693. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Jordan, V.; Blenkiron, C.; Chamley, L.W. Biodistribution of Extracellular Vesicles Following Administration into Animals: A Systematic Review. J. Extracell. Vesicles 2021, 10, e12085. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular Vesicles as a Next-Generation Drug Delivery Platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Dai, N.; Sheng, K.; Lu, H.; Wang, J.; Chen, L.; Wang, Y. Gut Bacterial Extracellular Vesicles: Important Players in Regulating Intestinal Microenvironment. Gut Microbes 2022, 14, 2134689. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Chen, P.; Xi, Y.; Sheng, J. From Trash to Treasure: The Role of Bacterial Extracellular Vesicles in Gut Health and Disease. Front. Immunol. 2023, 14, 1274295. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Huang, Z.; Yao, J.; Jin, Y. Extracellular Vesicles-Mediated Interaction within Intestinal Microenvironment in Inflammatory Bowel Disease. J. Adv. Res. 2022, 37, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Corrado, C.; Barreca, M.M.; Zichittella, C.; Alessandro, R.; Conigliaro, A. Molecular Mediators of Rna Loading into Extracellular Vesicles. Cells 2021, 10, 3355. [Google Scholar] [CrossRef]
- Schirizzi, A.; Contino, M.; Carrieri, L.; Riganti, C.; De Leonardis, G.; Scavo, M.P.; Perrone, M.G.; Miciaccia, M.; Kopecka, J.; Refolo, M.G.; et al. The Multiple Combination of Paclitaxel, Ramucirumab and Elacridar Reverses the Paclitaxel-Mediated Resistance in Gastric Cancer Cell Lines. Front. Oncol. 2023, 13, 1129832. [Google Scholar] [CrossRef]
- Wu, L.; Xue, M.; Lai, S.; Chen, J.; Lin, Y.; Ding, N.; Zhong, J.; Chen, S.; Wang, L. Hypoxia Derived Exosomes Promote the Proliferation and Metastasis of Colorectal Cancer through the Regulation of HIF-1α/MiR-4299/ZBTB4. Life Sci. 2023, 329, 121872. [Google Scholar] [CrossRef] [PubMed]
- Scavo, M.P.; Depalo, N.; Rizzi, F.; Carrieri, L.; Serino, G.; Franco, I.; Bonfiglio, C.; Pesole, P.L.; Cozzolongo, R.; Gianuzzi, V.; et al. Exosomal FZD-7 Expression Is Modulated by Different Lifestyle Interventions in Patients with NAFLD. Nutrients 2022, 14, 1133. [Google Scholar] [CrossRef] [PubMed]
- Negro, R.; Mastrogiacomo, R.; Carrieri, L.; Rizzi, F.; Arrè, V.; Minervini, G.; Fanizza, E.; Bianco, G.; Panniello, A.; Striccoli, M.; et al. Encapsulation of MCC950 in Liposomes Decorated with Anti-Frizzled 1 Improves Drug Bioavailability and Effectiveness in Fatty Liver Disease. ACS Appl. Mater. Interfaces 2023, 15, 33322–33334. [Google Scholar] [CrossRef] [PubMed]
- Scavo, M.P.; Negro, R.; Arrè, V.; Depalo, N.; Carrieri, L.; Rizzi, F.; Mastrogiacomo, R.; Serino, G.; Notarnicola, M.; De Nunzio, V.; et al. The Oleic/Palmitic Acid Imbalance in Exosomes Isolated from NAFLD Patients Induces Necroptosis of Liver Cells via the Elongase-6/RIP-1 Pathway. Cell Death Dis. 2023, 14, 635. [Google Scholar] [CrossRef] [PubMed]
- Scavo, M.P.; Lisco, G.; Depalo, N.; Rizzi, F.; Volpe, S.; Arrè, V.; Carrieri, L.; Notarnicola, M.; De Nunzio, V.; Curri, M.L.; et al. Semaglutide Modulates Extracellular Matrix Production of LX-2 Cells via Exosomes and Improves Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). Int. J. Mol. Sci. 2024, 25, 1493. [Google Scholar] [CrossRef] [PubMed]
- Akuta, N.; Kawamura, Y.; Watanabe, C.; Nishimura, A.; Okubo, M.; Mori, Y.; Fujiyama, S.; Sezaki, H.; Hosaka, T.; Kobayashi, M.; et al. Impact of Sodium Glucose Cotransporter 2 Inhibitor on Histological Features and Glucose Metabolism of Non-Alcoholic Fatty Liver Disease Complicated by Diabetes Mellitus. Hepatol. Res. 2019, 49, 531–539. [Google Scholar] [CrossRef]
- Thietart, S.; Rautou, P.E. Extracellular Vesicles as Biomarkers in Liver Diseases: A Clinician’s Point of View. J. Hepatol. 2020, 73, 1507–1525. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, J.; Verhulst, S.; Mannaerts, I.; Sowa, J.P.; Best, J.; Canbay, A.; Reynaert, H.; van Grunsven, L.A. A PDGFRβ-Based Score Predicts Significant Liver Fibrosis in Patients with Chronic Alcohol Abuse, NAFLD and Viral Liver Disease. EBioMedicine 2019, 43, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Payancé, A.; Silva-Junior, G.; Bissonnette, J.; Tanguy, M.; Pasquet, B.; Levi, C.; Roux, O.; Nekachtali, O.; Baiges, A.; Hernández-Gea, V.; et al. Hepatocyte Microvesicle Levels Improve Prediction of Mortality in Patients with Cirrhosis. Hepatology 2018, 68, 1508–1518. [Google Scholar] [CrossRef]
- Nimitrungtawee, N.; Inmutto, N.; Chattipakorn, S.C.; Chattipakorn, N. Extracellular Vesicles as a New Hope for Diagnosis and Therapeutic Intervention for Hepatocellular Carcinoma. Cancer Med. 2021, 10, 8253–8271. [Google Scholar] [CrossRef]
- Pu, C.; Huang, H.; Wang, Z.; Zou, W.; Lv, Y.; Zhou, Z.; Zhang, Q.; Qiao, L.; Wu, F.; Shao, S. Extracellular Vesicle-Associated Mir-21 and Mir-144 Are Markedly Elevated in Serum of Patients with Hepatocellular Carcinoma. Front. Physiol. 2018, 9, 930. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Yang, J.; Chen, M.; Cui, L.; Wang, T.; Gao, W.; Tian, J.; Wei, R. MicroRNA-21 as a Diagnostic Marker for Hepatocellular Carcinoma: A Systematic Review and Meta-Analysis. Pak. J. Med. Sci. 2019, 35, 1466–1471. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Ye, Y.; Bu, D.; Zhao, G.; Song, T.; Liu, P.; Yu, W.; Wang, H.; Li, H.; Ren, X.; et al. Imbalance of TGF-Β1/BMP-7 Pathways Induced by M2-Polarized Macrophages Promotes Hepatocellular Carcinoma Aggressiveness. Mol. Ther. 2021, 29, 2067–2087. [Google Scholar] [CrossRef] [PubMed]
- LeBleu, V.S.; Kalluri, R. Exosomes as a Multicomponent Biomarker Platform in Cancer. Trends Cancer 2020, 6, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Alesanco, A.; Marcuello, M.; Pastor-Jiménez, M.; López-Puerto, L.; Bonjoch, L.; Gironella, M.; Carrascal, M.; Abian, J.; de-Madaria, E.; Closa, D. Acute Pancreatitis Promotes the Generation of Two Different Exosome Populations. Sci. Rep. 2019, 9, 19887. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.; Liu, F.; Fan, J.; Sun, D.; Liu, C.; Lyon, C.J.; Bernard, D.W.; Li, Y.; Yokoi, K.; Katz, M.H.; et al. Nanoplasmonic Quantification of Tumour-Derived Extracellular Vesicles in Plasma Microsamples for Diagnosis and Treatment Monitoring. Nat. Biomed. Eng. 2017, 1, 0021. [Google Scholar] [CrossRef] [PubMed]
- Nesteruk, K.; Levink, I.J.M.; de Vries, E.; Visser, I.J.; Peppelenbosch, M.P.; Cahen, D.L.; Fuhler, G.M.; Bruno, M.J. Extracellular Vesicle-Derived MicroRNAs in Pancreatic Juice as Biomarkers for Detection of Pancreatic Ductal Adenocarcinoma. Pancreatology 2022, 22, 626–635. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.S.; Ciprani, D.; O’Shea, A.; Liss, A.S.; Yang, R.; Fletcher-Mercaldo, S.; Mino-Kenudson, M.; Fernández-del Castillo, C.; Weissleder, R. Extracellular Vesicle Analysis Allows for Identification of Invasive IPMN. Gastroenterology 2021, 160, 1345–1358.e11. [Google Scholar] [CrossRef] [PubMed]
- Papadakos, S.P.; Dedes, N.; Gkolemi, N.; Machairas, N.; Theocharis, S. The EPH/Ephrin System in Pancreatic Ductal Adenocarcinoma (PDAC): From Pathogenesis to Treatment. Int. J. Mol. Sci. 2023, 24, 3015. [Google Scholar] [CrossRef]
- Manne, A.; Esnakula, A.; Abushahin, L.; Tsung, A. Understanding the Clinical Impact of Muc5ac Expression on Pancreatic Ductal Adenocarcinoma. Cancers 2021, 13, 3059. [Google Scholar] [CrossRef]
- Manne, A.; Kasi, A.; Esnakula, A.K.; Paluri, R.K. Predictive Value of MUC5AC Signature in Pancreatic Ductal Adenocarcinoma: A Hypothesis Based on Preclinical Evidence. Int. J. Mol. Sci. 2023, 24, 8087. [Google Scholar] [CrossRef] [PubMed]
- Petrik, J.; Lauks, S.; Garlisi, B.; Lawler, J. Thrombospondins in the Tumor Microenvironment. Semin. Cell Dev. Biol. 2024, 155, 3–11. [Google Scholar] [CrossRef]
- Mo, W.Y.; Cao, S.Q. MiR-29a-3p: A Potential Biomarker and Therapeutic Target in Colorectal Cancer. Clin. Transl. Oncol. 2023, 25, 563–577. [Google Scholar] [CrossRef]
- Saberi, S.; Esmaeili, M.; Saghiri, R.; Shekari, F.; Mohammadi, M. Assessment of the Mixed Origin of the Gastric Epithelial Extracellular Vesicles in Acellular Transfer of Helicobacter Pylori Toxins and a Systematic Review. Microb. Pathog. 2023, 177, 106024. [Google Scholar] [CrossRef] [PubMed]
- González, M.F.; Burgos-Ravanal, R.; Shao, B.; Heinecke, J.; Valenzuela-Valderrama, M.; Corvalán, A.H.; Quest, A.F.G. Extracellular Vesicles from Gastric Epithelial GES-1 Cells Infected with Helicobacter Pylori Promote Changes in Recipient Cells Associated with Malignancy. Front. Oncol. 2022, 12, 962920. [Google Scholar] [CrossRef]
- Sugano, K. Screening of Gastric Cancer in Asia. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, P.W.; Yang, W.Q.; Mi, H.; Pan, J.L.; Huang, Y.C.; Hou, Z.K.; Hou, Q.K.; Luo, Q.; Liu, F. Bin Identification of Non-Invasive Biomarkers for Chronic Atrophic Gastritis from Serum Exosomal MicroRNAs. BMC Cancer 2019, 19, 129. [Google Scholar] [CrossRef]
- Jiang, C.; Zhang, J.; Wang, W.; Shan, Z.; Sun, F.; Tan, Y.; Tong, Y.; Qiu, Y. Extracellular Vesicles in Gastric Cancer: Role of Exosomal LncRNA and MicroRNA as Diagnostic and Therapeutic Targets. Front. Physiol. 2023, 14, 1158839. [Google Scholar] [CrossRef]
- Hur, J.Y.; Lee, K.Y. Characteristics and Clinical Application of Extracellular Vesicle-derived DNA. Cancers 2021, 13, 3827. [Google Scholar] [CrossRef]
- Lin, L.Y.; Yang, L.; Zeng, Q.; Wang, L.; Chen, M.L.; Zhao, Z.H.; Ye, G.D.; Luo, Q.C.; Lv, P.Y.; Guo, Q.W.; et al. Tumor-Originated Exosomal LncUEGC1 as a Circulating Biomarker for Early-Stage Gastric Cancer. Mol. Cancer 2018, 17, 84. [Google Scholar] [CrossRef]
- Zhao, R.; Zhang, Y.; Zhang, X.; Yang, Y.; Zheng, X.; Li, X.; Liu, Y.; Zhang, Y. Exosomal Long Noncoding RNA HOTTIP as Potential Novel Diagnostic and Prognostic Biomarker Test for Gastric Cancer. Mol. Cancer 2018, 17, 68. [Google Scholar] [CrossRef]
- Wang, X.; Zhou, G.; Zhou, W.; Wang, X.; Wang, X.; Miao, C. Exosomes as a New Delivery Vehicle in Inflammatory Bowel Disease. Pharmaceutics 2021, 13, 1644. [Google Scholar] [CrossRef]
- Liu, R.; Tang, A.; Wang, X.; Chen, X.; Zhao, L.; Xiao, Z.; Shen, S. Inhibition of LncRNA NEAT1 Suppresses the Inflammatory Response in IBD by Modulating the Intestinal Epithelial Barrier and by Exosome-Mediated Polarization of Macrophages. Int. J. Mol. Med. 2018, 42, 2903–2913. [Google Scholar] [CrossRef] [PubMed]
- Heydari, R.; Fayazzadeh, S.; Shahrokh, S.; Shekari, F.; Farsad, F.; Meyfour, A. Plasma Extracellular Vesicle LncRNA H19 as a Potential Diagnostic Biomarker for Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2023, izad219. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.J.; Li, W.W.; Wang, X.L.; Ma, A.H.; Qin, Y.H. Efficacy of Extracellular Vesicles as a Cell-Free Therapy in Colitis: A Systematic Review and Meta-Analysis of Animal Studies. Front. Pharmacol. 2023, 14, 1260134. [Google Scholar] [CrossRef] [PubMed]
- Masi, L.; Capobianco, I.; Magrì, C.; Marafini, I.; Petito, V.; Scaldaferri, F. MicroRNAs as Innovative Biomarkers for Inflammatory Bowel Disease and Prediction of Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 7991. [Google Scholar] [CrossRef]
- Shen, Q.; Huang, Z.; Ma, L.; Yao, J.; Luo, T.; Zhao, Y.; Xiao, Y.; Jin, Y. Extracellular Vesicle MiRNAs Promote the Intestinal Microenvironment by Interacting with Microbes in Colitis. Gut Microbes 2022, 14, 2128604. [Google Scholar] [CrossRef]
- Scavo, M.P.; Depalo, N.; Tutino, V.; De Nunzio, V.; Ingrosso, C.; Rizzi, F.; Notarnicola, M.; Curri, M.L.; Giannelli, G. Exosomes for Diagnosis and Therapy in Gastrointestinal Cancers. Int. J. Mol. Sci. 2020, 21, 367. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.B.; Yan, C.; Mu, L.; Mi, Y.L.; Zhao, H.; Hu, H.; Li, X.L.; Tao, D.D.; Wu, Y.Q.; Gong, J.P.; et al. Exosomal Wnt-Induced Dedifferentiation of Colorectal Cancer Cells Contributes to Chemotherapy Resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef]
- Sun, B.; Li, Y.; Zhou, Y.; Ng, T.K.; Zhao, C.; Gan, Q.; Gu, X.; Xiang, J. Circulating Exosomal CPNE3 as a Diagnostic and Prognostic Biomarker for Colorectal Cancer. J. Cell Physiol. 2019, 234, 1416–1425. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Yang, X.; Jiang, Y.; Li, A.; Cong, J.; Li, Y.; Xie, Q.; Xu, C.; Liu, D. Identification of Faecal Extracellular Vesicles as Novel Biomarkers for the Non-Invasive Diagnosis and Prognosis of Colorectal Cancer. J. Extracell. Vesicles 2023, 12, e12300. [Google Scholar] [CrossRef]
- Rong, W.; Shao, S.; Pu, Y.; Ji, Q.; Zhu, H. Circulating Extracellular Vesicle-Derived MARCKSL1 Is a Potential Diagnostic Non-Invasive Biomarker in Metastatic Colorectal Cancer Patients. Sci. Rep. 2023, 13, 9957. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Suzuki, R.; Takagi, T.; Sugimoto, M.; Ohira, H. Circulating Extracellular Vesicle-encapsulated MicroRNA as Screening Biomarkers for Intraductal Papillary Mucinous Neoplasm. Oncol. Lett. 2020, 20, 315. [Google Scholar] [CrossRef]
- Leoni, G.; Neumann, P.A.; Kamaly, N.; Quiros, M.; Nishio, H.; Jones, H.R.; Sumagin, R.; Hilgarth, R.S.; Alam, A.; Fredman, G.; et al. Annexin A1’containing Extracellular Vesicles and Polymeric Nanoparticles Promote Epithelial Wound Repair. J. Clin. Investig. 2015, 125, 1215–1227. [Google Scholar] [CrossRef] [PubMed]
- Fikree, A.; Byrne, P. Management of Functional Gastrointestinal Disorders. Clin. Med. J. R. Coll. Physicians Lond. 2021, 21, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Huang, X.; Xiu, H.; Sun, Y.; Chen, J.; Cheng, G.; Song, Z.; Peng, Y.; Shen, Y.; Wang, J.; et al. Extracellular Vesicles: Natural Liver-Accumulating Drug Delivery Vehicles for the Treatment of Liver Diseases. J. Extracell. Vesicles 2020, 10, e12030. [Google Scholar] [CrossRef]
- Grigoryeva, E.S.; Tashireva, L.A.; Savelieva, O.E.; Zavyalova, M.V.; Popova, N.O.; Kuznetsov, G.A.; Andryuhova, E.S.; Perelmuter, V.M. The Association of Integrins Β3, Β4, and AVβ5 on Exosomes, CTCs and Tumor Cells with Localization of Distant Metastasis in Breast Cancer Patients. Int. J. Mol. Sci. 2023, 24, 2929. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Sun, S.; Chen, Z.; Xiang, S.; Ding, Z.; Huang, Z.; Zhang, B. Understanding the Versatile Roles and Applications of EpCAM in Cancers: From Bench to Bedside. Exp. Hematol. Oncol. 2022, 11, 97. [Google Scholar] [CrossRef]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S.; et al. Akkermansia Muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front. Microbiol. 2019, 10, 2155. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, Q.; Guo, M.; Fang, H.; Li, T.; Zhu, Y.; Jiang, H.; Xiao, P.; Hu, M. Mesenchymal Stem Cell–Derived Exosomes in Various Chronic Liver Diseases: Hype or Hope? J. Inflamm. Res. 2024, 17, 171–189. [Google Scholar] [CrossRef]
- Azparren-Angulo, M.; Royo, F.; Gonzalez, E.; Liebana, M.; Brotons, B.; Berganza, J.; Goñi-de-Cerio, F.; Manicardi, N.; Abad-Jordà, L.; Gracia-Sancho, J.; et al. Extracellular Vesicles in Hepatology: Physiological Role, Involvement in Pathogenesis, and Therapeutic Opportunities. Pharmacol. Ther. 2021, 218, 107683. [Google Scholar] [CrossRef]
- Shao, M.; Xu, Q.; Wu, Z.; Chen, Y.; Shu, Y.; Cao, X.; Chen, M.; Zhang, B.; Zhou, Y.; Yao, R.; et al. Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells Ameliorate IL-6-Induced Acute Liver Injury through MiR-455-3p. Stem. Cell Res. Ther. 2020, 11, 37. [Google Scholar] [CrossRef]
- Shiha, G.; Nabil, A.; Lotfy, A.; Soliman, R.; Hassan, A.A.; Ali, I.S.; Gad, D.F.; Zahran, F. Antifibrotic Effect of Combination of Nilotinib and Stem Cell-Conditioned Media on CCl4-Induced Liver Fibrosis. Stem. Cells Int. 2020, 2020, 6574010. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, L.; Hu, H.; Wang, H.; Wang, X.; Jiang, J.; Ma, Y.; Yang, J.; Hou, Y.; Xie, D.; et al. Pretreatment of Exosomes Derived from HUCMSCs with TNF-α Ameliorates Acute Liver Failure by Inhibiting the Activation of NLRP3 in Macrophage. Life Sci. 2020, 246, 117401. [Google Scholar] [CrossRef]
- Moulin, C.; Crupi, M.J.F.; Ilkow, C.S.; Bell, J.C.; Boulton, S. Extracellular Vesicles and Viruses: Two Intertwined Entities. Int. J. Mol. Sci. 2023, 24, 1036. [Google Scholar] [CrossRef]
- De, A.; Bhagat, N.; Mehta, M.; Taneja, S.; Duseja, A. Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD) Definition Is Better than MAFLD Criteria for Lean Patients with NAFLD. J. Hepatol. 2023, 80, e61–e62. [Google Scholar] [CrossRef]
- Long, M.T.; Noureddin, M.; Lim, J.K. AGA Clinical Practice Update: Diagnosis and Management of Nonalcoholic Fatty Liver Disease in Lean Individuals: Expert Review. Gastroenterology 2022, 163, 764–774.e1. [Google Scholar] [CrossRef]
- Zeng, X.; Zhu, M.; Liu, X.; Chen, X.; Yuan, Y.; Li, L.; Liu, J.; Lu, Y.; Cheng, J.; Chen, Y. Oleic Acid Ameliorates Palmitic Acid Induced Hepatocellular Lipotoxicity by Inhibition of ER Stress and Pyroptosis. Nutr. Metab. 2020, 17, 11. [Google Scholar] [CrossRef]
- Chen, Q.; Duan, X.; Xu, M.; Fan, H.; Dong, Y.; Wu, H.; Zhang, M.; Liu, Y.; Nan, Z.; Deng, S.; et al. BMSC-EVs Regulate Th17 Cell Differentiation in UC via H3K27me3. Mol. Immunol. 2020, 118, 191–200. [Google Scholar] [CrossRef]
- Warren, M.R.; Zhang, C.; Vedadghavami, A.; Bokvist, K.; Dhal, P.K.; Bajpayee, A.G. Milk Exosomes with Enhanced Mucus Penetrability for Oral Delivery of SiRNA. Biomater. Sci. 2021, 9, 4260–4277. [Google Scholar] [CrossRef] [PubMed]
- Kameli, N.; Dragojlovic-kerkache, A.; Savelkoul, P.; Stassen, F.R. Plant-derived Extracellular Vesicles: Current Findings, Challenges, and Future Applications. Membranes 2021, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Li, W.; Shao, L.; Zhou, A.; Zhao, M.; Li, P.; Zhang, Z.; Wu, J. Both Extracellular Vesicles from Helicobacter Pylori-Infected Cells and Helicobacter Pylori Outer Membrane Vesicles Are Involved in Gastric/Extragastric Diseases. Eur. J. Med. Res. 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.K.A.; Perretta, S.; Perrod, G.; Pidial, L.; Lindner, V.; Carn, F.; Lemieux, S.; Alloyeau, D.; Boucenna, I.; Menasché, P.; et al. Thermoresponsive Gel Embedded with Adipose Stem-Cell-Derived Extracellular Vesicles Promotes Esophageal Fistula Healing in a Thermo-Actuated Delivery Strategy. ACS Nano 2018, 12, 9800–9814. [Google Scholar] [CrossRef]
- He, Z.; Li, W.; Zheng, T.; Liu, D.; Zhao, S. Human Umbilical Cord Mesenchymal Stem Cells-Derived Exosomes Deliver MicroRNA-375 to Downregulate ENAH and Thus Retard Esophageal Squamous Cell Carcinoma Progression. J. Exp. Clin. Cancer Res. 2020, 39, 140. [Google Scholar] [CrossRef]
- Vijay, A.; Valdes, A.M. Role of the Gut Microbiome in Chronic Diseases: A Narrative Review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef] [PubMed]
- Borghi, M.; Pariano, M.; Solito, V.; Puccetti, M.; Bellet, M.M.; Stincardini, C.; Renga, G.; Vacca, C.; Sellitto, F.; Mosci, P.; et al. Targeting the Aryl Hydrocarbon Receptor with Indole-3-Aldehyde Protects From Vulvovaginal Candidiasis via the IL-22-IL-18 Cross-Talk. Front. Immunol. 2019, 10, 2364. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.N.; Swimm, A.; Sonowal, R.; Bretin, A.; Gewirtz, A.T.; Jones, R.M.; Kalman, D. Indoles from the Commensal Microbiota Act via the AHR and IL-10 to Tune the Cellular Composition of the Colonic Epithelium during Aging. Proc. Natl. Acad. Sci. USA 2020, 117, 21519–21526. [Google Scholar] [CrossRef]
- Sun, B.; Sawant, H.; Borthakur, A.; Bihl, J.C. Emerging Therapeutic Role of Gut Microbial Extracellular Vesicles in Neurological Disorders. Front. Neurosci. 2023, 17, 1241418. [Google Scholar] [CrossRef]
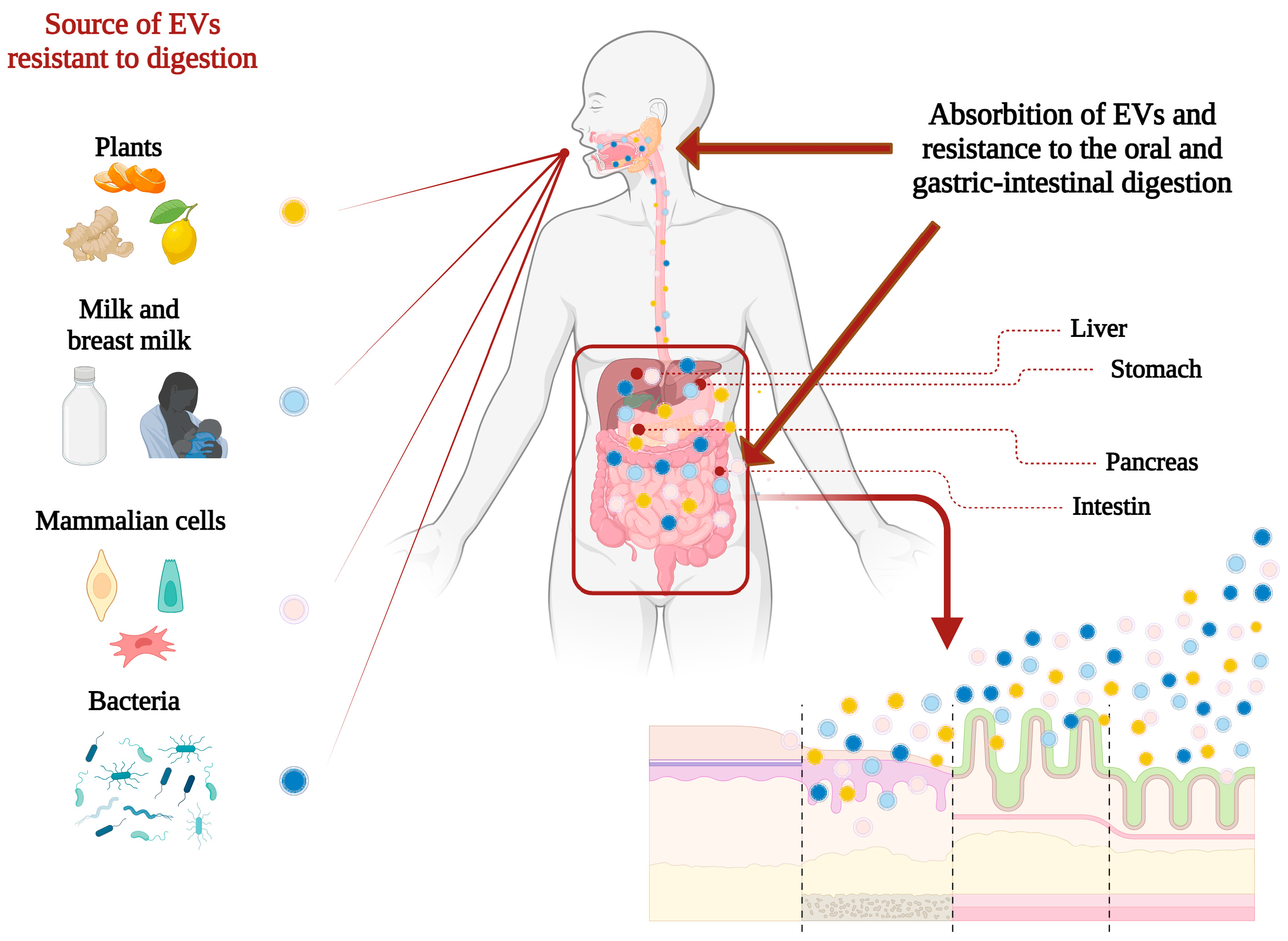
- Donoso-Meneses, D.; Figueroa-Valdés, A.I.; Khoury, M.; Alcayaga-Miranda, F. Oral Administration as a Potential Alternative for the Delivery of Small Extracellular Vesicles. Pharmaceutics 2023, 15, 716. [Google Scholar] [CrossRef]
- Tulkens, J.; Vergauwen, G.; Van Deun, J.; Geeurickx, E.; Dhondt, B.; Lippens, L.; De Scheerder, M.A.; Miinalainen, I.; Rappu, P.; De Geest, B.G.; et al. Increased Levels of Systemic LPS-Positive Bacterial Extracellular Vesicles in Patients with Intestinal Barrier Dysfunction. Gut 2020, 69, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Mondal, J.; Pillarisetti, S.; Junnuthula, V.; Surwase, S.S.; Hwang, S.R.; Park, I.K.; Lee, Y.K. Extracellular Vesicles and Exosome-like Nanovesicles as Pioneering Oral Drug Delivery Systems. Front. Bioeng Biotechnol. 2023, 11, 1307878. [Google Scholar] [CrossRef]
- Kandimalla, R.; Aqil, F.; Alhakeem, S.S.; Jeyabalan, J.; Tyagi, N.; Agrawal, A.; Yan, J.; Spencer, W.; Bondada, S.; Gupta, R.C. Targeted Oral Delivery of Paclitaxel Using Colostrum-Derived Exosomes. Cancers 2021, 13, 3700. [Google Scholar] [CrossRef] [PubMed]
- Barone, A.; D’avanzo, N.; Cristiano, M.C.; Paolino, D.; Fresta, M. Macrophage-Derived Extracellular Vesicles: A Promising Tool for Personalized Cancer Therapy. Biomedicines 2022, 10, 1252. [Google Scholar] [CrossRef]
- Oz, T.; Kaushik, A.; Kujawska, M. Neural Stem Cells for Parkinson’s Disease Management: Challenges, Nanobased Support, and Prospects. World J. Stem. Cells 2023, 15, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem. Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.R.; Teixeira, G.Q.; Santos, S.G.; Barbosa, M.A.; Almeida-Porada, G.; Gonçalves, R.M. Mesenchymal Stromal Cell Secretome: Influencing Therapeutic Potential by Cellular Pre-Conditioning. Front. Immunol. 2018, 9, 2837. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jankovic, M.G.; Fellabaum, C.; Volarevic, A.; Djonov, V.; Arsenijevic, A.; Volarevic, V. Molecular Mechanisms Responsible for Anti-Inflammatory and Immunosuppressive Effects of Mesenchymal Stem Cell-Derived Factors. In Advances in Experimental Medicine and Biology; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; Volume 1084. [Google Scholar]
- Han, L.; Zhao, Z.; Chen, X.; Yang, K.; Tan, Z.; Huang, Z.; Zhou, L.; Dai, R. Human Umbilical Cord Mesenchymal Stem Cells-Derived Exosomes for Treating Traumatic Pancreatitis in Rats. Stem. Cell Res. Ther. 2022, 13, 221. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, H.; Zhangdi, H.; Xu, R.; Zhang, X.; Liu, J.; Hu, Y.; Ning, D.; Jin, S. Hair Follicle-MSC-Derived Small Extracellular Vesicles as a Novel Remedy for Acute Pancreatitis. J. Control. Release 2022, 352, 1104–1115. [Google Scholar] [CrossRef]
- Wang, N.; Ma, J.; Ren, Y.; Xiang, S.H.; Jia, R.R. Secreted Klotho from Exosomes Alleviates Inflammatioand Apoptosis in Acute Pancreatitis. Am. J. Transl. Res. 2019, 11, 3375–3383. [Google Scholar]
- Jia, Y.C.; Ding, Y.X.; Mei, W.T.; Wang, Y.T.; Zheng, Z.; Qu, Y.X.; Liang, K.; Li, J.; Cao, F.; Li, F. Extracellular Vesicles and Pancreatitis: Mechanisms, Status and Perspectives. Int. J. Biol. Sci. 2021, 17, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.; Kong, F.; Wu, D. Prospect of Mesenchymal Stem-Cell-Conditioned Medium in the Treatment of Acute Pancreatitis: A Systematic Review. Biomedicines 2023, 11, 2343. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The Global Burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.T.; Wang, B.; Lv, L.L.; Liu, B.C. Extracellular Vesicle-Based Nanotherapeutics: Emerging Frontiers in Anti-Inflammatory Therapy. Theranostics 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Fan, H.; Shou, Z.; Xu, M.; Chen, Q.; Ai, C.; Dong, Y.; Liu, Y.; Nan, Z.; Wang, Y.; et al. Extracellular Vesicles Containing MiR-146a Attenuate Experimental Colitis by Targeting TRAF6 and IRAK1. Int. Immunopharmacol. 2019, 68, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yu, M.; Chen, F.; Wang, L.; Ye, C.; Chen, Q.; Zhu, Q.; Xie, D.; Shao, M.; Yang, L. A Novel Delivery Nanobiotechnology: Engineered MiR-181b Exosomes Improved Osteointegration by Regulating Macrophage Polarization. J. Nanobiotechnol. 2021, 19, 269. [Google Scholar] [CrossRef]
- Ma, L.; Shen, Q.; Lyu, W.; Lv, L.; Wang, W.; Yu, M.; Yang, H.; Tao, S.; Xiao, Y. Clostridium Butyricum and Its Derived Extracellular Vesicles Modulate Gut Homeostasis and Ameliorate Acute Experimental Colitis. Microbiol. Spectr. 2022, 10, e0136822. [Google Scholar] [CrossRef]
- Seay, T.W.; Suo, Z. Roles of Extracellular Vesicles on the Progression and Metastasis of Hepatocellular Carcinoma. Cells 2023, 12, 1879. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, S.; Rendón-Huerta, E.P.; Ortiz-Navarrete, V.; Montaño, L.F. CD38 and Regulation of the Immune Response Cells in Cancer. J. Oncol. 2021, 2021, 6630295. [Google Scholar] [CrossRef]
- Lam, J.H.; Ng, H.H.M.; Lim, C.J.; Sim, X.N.; Malavasi, F.; Li, H.; Loh, J.J.H.; Sabai, K.; Kim, J.K.; Ong, C.C.H.; et al. Expression of CD38 on Macrophages Predicts Improved Prognosis in Hepatocellular Carcinoma. Front. Immunol. 2019, 10, 2093. [Google Scholar] [CrossRef]
- Joey Lee, J.Y.; Yeong, J.; Nadia Lee, L.W.J.; Loo, L.-H.; Dong, J. 627 ImmunoAtlas: An Online Public Portal for Sharing, Visualizing, and Referencing Multiplex Immunohistochemistry/Immunofluorescence (MIHC/IF) Images and Results for Immuno-Oncology. J. Immunother. Cancer 2021, 9, A657. [Google Scholar] [CrossRef]
- Ke, M.; Zhang, Z.; Xu, B.; Zhao, S.; Ding, Y.; Wu, X.; Wu, R.; Lv, Y.; Dong, J. Baicalein and Baicalin Promote Antitumor Immunity by Suppressing PD-L1 Expression in Hepatocellular Carcinoma Cells. Int. Immunopharmacol. 2019, 75, 105824. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ke, H. Overcoming the Resistance of Hepatocellular Carcinoma to PD-1/PD-L1 Inhibitor and the Resultant Immunosuppression by CD38 SiRNA-Loaded Extracellular Vesicles. Oncoimmunology 2023, 12, 2152635. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Jaffar Ali, D.; Qi, Y.; Li, Y.; Sun, B.; Liu, R.; Sun, B.; Xiao, Z. Engineered Extracellular Vesicles Mediated CRISPR-Induced Deficiency of IQGAP1/FOXM1 Reverses Sorafenib Resistance in HCC by Suppressing Cancer Stem Cells. J. Nanobiotechnol. 2023, 21, 154. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.J.; Zheng, B.; Wang, H.Y.; Chen, L. New Knowledge of the Mechanisms of Sorafenib Resistance in Liver Cancer. Acta Pharmacol. Sin. 2017, 38, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Duda, D.G. Overcoming Sorafenib Treatment-Resistance in Hepatocellular Carcinoma: A Future Perspective at a Time of Rapidly Changing Treatment Paradigms. EBioMedicine 2020, 52, 102644. [Google Scholar] [CrossRef] [PubMed]
- Zou, G.-M. Liver Cancer Stem Cells as an Important Target in Liver Cancer Therapies. Anticancer. Agents. Med. Chem. 2012, 10, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Nio, K.; Yamashita, T.; Kaneko, S. The Evolving Concept of Liver Cancer Stem Cells. Mol. Cancer 2017, 16, 4. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, Y.; Sun, R.; Liu, Y.; Wang, Y.; Luo, H.; Shi, B.; Jiang, H.; Li, Z. Chimeric Anti-GPC3 SFv-CD3ε Receptor-Modified T Cells with IL7 Co-Expression for the Treatment of Solid Tumors. Mol. Ther. Oncolytics 2022, 25, 160–173. [Google Scholar] [CrossRef]
- Shih, T.C.; Wang, L.; Wang, H.C.; Wan, Y.J.Y. Glypican-3: A Molecular Marker for the Detection and Treatment of Hepatocellular Carcinoma. Liver Res. 2020, 4, 168–172. [Google Scholar] [CrossRef]
- Partyka, O.; Pajewska, M.; Kwaśniewska, D.; Czerw, A.; Deptała, A.; Budzik, M.; Cipora, E.; Gąska, I.; Gazdowicz, L.; Mielnik, A.; et al. Overview of Pancreatic Cancer Epidemiology in Europe and Recommendations for Screening in High-Risk Populations. Cancers 2023, 15, 3634. [Google Scholar] [CrossRef] [PubMed]
- Neoptolemos, J.P.; Kleeff, J.; Michl, P.; Costello, E.; Greenhalf, W.; Palmer, D.H. Therapeutic Developments in Pancreatic Cancer: Current and Future Perspectives. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Xia, P.; Xu, X.; Shen, J.; Ding, Y.; Jiang, Y.; Wang, H.; Xie, X.; Zhang, X.; Li, W.; et al. Targeted Delivery of Organic Small-Molecule Photothermal Materials with Engineered Extracellular Vesicles for Imaging-Guided Tumor Photothermal Therapy. J. Nanobiotechnol. 2023, 21, 442. [Google Scholar] [CrossRef]
- Ding, Y.; Cao, F.; Sun, H.; Wang, Y.; Liu, S.; Wu, Y.; Cui, Q.; Mei, W.T.; Li, F. Exosomes Derived from Human Umbilical Cord Mesenchymal Stromal Cells Deliver Exogenous MiR-145-5p to Inhibit Pancreatic Ductal Adenocarcinoma Progression. Cancer Lett. 2019, 442, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Ji, J.; Chen, X.; Xu, W.; Chen, H.; Zhu, S.; Wu, J.; Wu, Y.; Sun, Y.; Sai, W.; et al. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Carrying Hsa-MiRNA-128-3p Suppress Pancreatic Ductal Cell Carcinoma by Inhibiting Galectin-3. Clin. Transl. Oncol. 2022, 24, 517–531. [Google Scholar] [CrossRef]
- Draguet, F.; Dubois, N.; Bouland, C.; Pieters, K.; Bron, D.; Meuleman, N.; Stamatopoulos, B.; Lagneaux, L. Extracellular Vesicles Derived from Human Umbilical Cord Mesenchymal Stromal Cells as an Efficient Nanocarrier to Deliver SiRNA or Drug to Pancreatic Cancer Cells. Cancers 2023, 15, 2901. [Google Scholar] [CrossRef]
- Araujo-Abad, S.; Manresa-Manresa, A.; Rodríguez-Cañas, E.; Fuentes-Baile, M.; García-Morales, P.; Mallavia, R.; Saceda, M.; de Juan Romero, C. New Therapy for Pancreatic Cancer Based on Extracellular Vesicles. Biomed. Pharmacother. 2023, 162, 114657. [Google Scholar] [CrossRef]
- Sun, H.; Bhandari, K.; Burrola, S.; Wu, J.; Ding, W.Q. Pancreatic Ductal Cell-Derived Extracellular Vesicles Are Effective Drug Carriers to Enhance Paclitaxel’s Efficacy in Pancreatic Cancer Cells through Clathrin-Mediated Endocytosis. Int. J. Mol. Sci. 2022, 2, 4773. [Google Scholar] [CrossRef]
- Marshall, J.F. Targeting CDH17 in Cancer: When Blocking the Ligand Beats Blocking the Receptor? Clin. Cancer Res. 2018, 24, 253–255. [Google Scholar] [CrossRef]
- Xia, P.; Yuan, H.; Tian, M.; Zhong, T.; Hou, R.; Xu, X.; Ma, J.; Wang, H.; Li, Z.; Huang, D.; et al. Surface-Engineered Extracellular Vesicles with CDH17 Nanobodies to Efficiently Deliver Imaging Probes and Chemo-Photothermal Drugs for Gastric Cancer Theragnostic. Adv. Funct. Mater. 2023, 33, 2209393. [Google Scholar] [CrossRef]
- Liu, M.; Li, L.; Jin, D.; Liu, Y. Nanobody—A Versatile Tool for Cancer Diagnosis and Therapeutics. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2021, 13, e1697. [Google Scholar] [CrossRef]
- Li, D.H.; Smith, B.D. Deuterated Indocyanine Green (ICG) with Extended Aqueous Storage Shelf-Life: Chemical and Clinical Implications. Chem. Eur. J. 2021, 27, 14535–14542. [Google Scholar] [CrossRef] [PubMed]
- You, Q.; Wang, F.; Du, R.; Pi, J.; Wang, H.; Huo, Y.; Liu, J.; Wang, C.; Yu, J.; Yang, Y.; et al. M6A Reader YTHDF1-Targeting Engineered Small Extracellular Vesicles for Gastric Cancer Therapy via Epigenetic and Immune Regulation. Adv. Mater. 2023, 35, e2204910. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Yun, J.; Tang, W.; Familiari, G.; Relucenti, M.; Wu, J.; Li, X.; Chen, H.; Chen, R. Therapeutic M6A Eraser ALKBH5 MRNA-Loaded Exosome-Liposome Hybrid Nanoparticles Inhibit Progression of Colorectal Cancer in Preclinical Tumor Models. ACS Nano 2023, 17, 11838–11854. [Google Scholar] [CrossRef] [PubMed]
- Tran, P.H.L.; Wang, T.; Yin, W.; Tran, T.T.D.; Nguyen, T.N.G.; Lee, B.J.; Duan, W. Aspirin-Loaded Nanoexosomes as Cancer Therapeutics. Int. J. Pharm. 2019, 572, 118786. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.D.; Kim, H.Y.; Choi, Y.H.; Park, J.O.; Choi, E. Tumor-Derived Extracellular Vesicles for the Active Targeting and Effective Treatment of Colorectal Tumors in Vivo. Drug Deliv. 2022, 29, 2105444. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, M.; Baghaei, K.; Hajivalili, M.; Zali, M.R.; Ebtekar, M.; Amani, D. The Anti-Tumor Effects of CT-26 Derived Exosomes Enriched by MicroRNA-34a on Murine Model of Colorectal Cancer. Life Sci. 2022, 290, 120234. [Google Scholar] [CrossRef]
- Di Gioia, S.; Hossain, M.N.; Conese, M. Biological Properties and Therapeutic Effects of Plant-Derived Nanovesicles. Open Med. 2020, 15, 1096–1122. [Google Scholar] [CrossRef]
- Babaker, M.A.; Aljoud, F.A.; Alkhilaiwi, F.; Algarni, A.; Ahmed, A.; Khan, M.I.; Saadeldin, I.M.; Alzahrani, F.A. The Therapeutic Potential of Milk Extracellular Vesicles on Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 6812. [Google Scholar] [CrossRef]
- Xiong, L.; Wei, Y.; Jia, Q.; Chen, J.; Chen, T.; Yuan, J.; Pi, C.; Liu, H.; Tang, J.; Yin, S.; et al. The Application of Extracellular Vesicles in Colorectal Cancer Metastasis and Drug Resistance: Recent Advances and Trends. J. Nanobiotechnol. 2023, 21, 143. [Google Scholar] [CrossRef]
- Liang, G.; Zhu, Y.; Ali, D.J.; Tian, T.; Xu, H.; Si, K.; Sun, B.; Chen, B.; Xiao, Z. Engineered Exosomes for Targeted Co-Delivery of MiR-21 Inhibitor and Chemotherapeutics to Reverse Drug Resistance in Colon Cancer. J. Nanobiotechnol. 2020, 18, 10. [Google Scholar] [CrossRef]
- Bagheri, E.; Abnous, K.; Farzad, S.A.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Targeted Doxorubicin-Loaded Mesenchymal Stem Cells-Derived Exosomes as a Versatile Platform for Fighting against Colorectal Cancer. Life Sci. 2020, 261, 118369. [Google Scholar] [CrossRef]
- Kwon, S.H.; Al Faruque, H.; Kee, H.; Kim, E.; Park, S. Exosome-Based Hybrid Nanostructures for Enhanced Tumor Targeting and Hyperthermia Therapy. Colloids Surf. B Biointerfaces 2021, 205, 111915. [Google Scholar] [CrossRef]
- Su, R.; Dong, L.; Li, C.; Nachtergaele, S.; Wunderlich, M.; Qing, Y.; Deng, X.; Wang, Y.; Weng, X.; Hu, C.; et al. R-2HG Exhibits Anti-Tumor Activity by Targeting FTO/M6A/MYC/CEBPA Signaling. Cell 2018, 172, 90–105.e23. [Google Scholar] [CrossRef]
- Fiorucci, S.; Baldoni, M.; Ricci, P.; Zampella, A.; Distrutti, E.; Biagioli, M. Bile Acid-Activated Receptors and the Regulation of Macrophages Function in Metabolic Disorders. Curr. Opin. Pharmacol. 2020, 53, 45–54. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, L.; Li, C.; Yu, Y.; Yi, Y.; Wang, J.; Chen, D. Exosome-Induced Regulation in Inflammatory Bowel Disease. Front. Immunol. 2019, 10, 1464. [Google Scholar] [CrossRef] [PubMed]



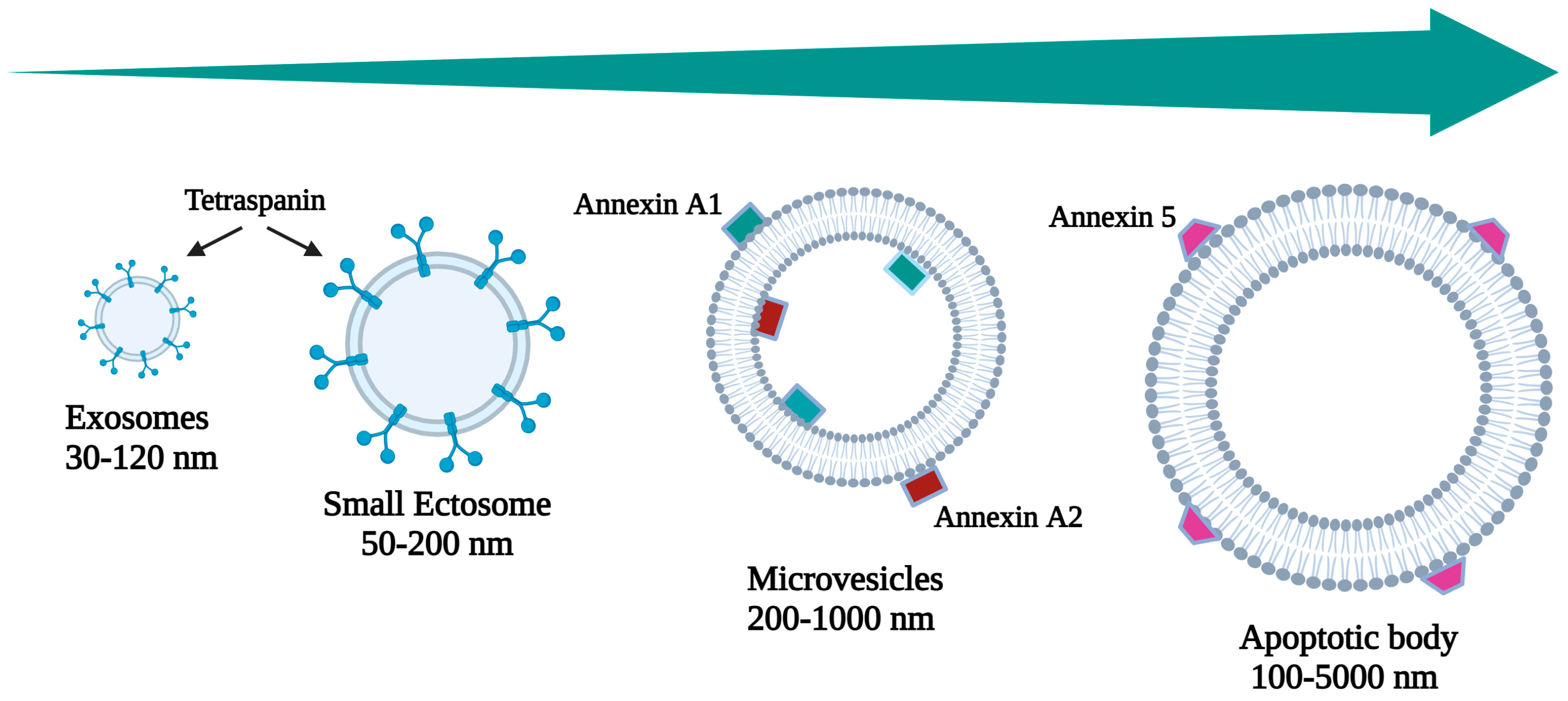
| Name | Category | Size (nm) | Origin | Markers | Bibliography |
|---|---|---|---|---|---|
| Exosomes | Small EVs | 30–120 | Multivesicular endosomes and amphisomes | CD63 Syntenin 1 CD81 | [9,10] |
| Small ectosomes | Small EVs | 50–200 | Ectosome | TSG101 CD9 ARDC1 CD 147 | [11,12,13] |
| Microvesicles | Small to large | 200–1000 | Ectosome | Annexin A1 Annexin A2 α-Actin 4 | [14] |
| Apoptotic Body | Small to large | 100–5000 | Apoptosis | Annexin V | [15] |
| Organ | Pathology | EVs Markers | Targets and/or Pathway | Bibliography |
|---|---|---|---|---|
| Pancreas | Pancreatitis | miR-155 | SOCS1 regulation | [55,83] |
| PDAC | miR-192-5p | Epigenetic regulation | [57] | |
| miR-21 | MMP2, MMP9, and VEGF regulation | |||
| miR-25 | Cell proliferation promotion | |||
| miR-16 | Post-transcriptional expression of Bcl-2 regulation | |||
| PC | MUC2, MUC4, MUC5AC, MUC6, MUC13 | Elevated pathway regulation determining the severity of PC | [58,59,60,61,62] | |
| EGFR, GPC1, WNT-2, EpCAM, MUC1 | Aberrant protein | |||
| Das-1 | 3′-sulfated Lewis A/C recognition | |||
| STMN1 | Cell proliferation inhibition | |||
| TSP1 | Participation in the differentiation of Th17 cells | |||
| TSP2 | Osteoblasts regulation | |||
| ZEB1 | EMT induction | |||
| EphA2 | Cell migration | |||
| S100A4 | Src and FAK activation | |||
| PSCA | Thymic lymphocyte differentiation, maturation, and activation | |||
| HOOK1 | Re-arrangement of the cytoskeleton | |||
| PTPN6 | SP1/MAPK signaling pathway inhibition | |||
| FBN1 | Immune cell infiltration in tumors increases | |||
| Stomach and Esofagous | GERD and Gastritis | miR-29a-3p | EMT, ZEB 1 and ZEB2 regulation | [63] |
| Atrofic Gastritis | miR-122-5p | Targeting CTDNEP1/LPIN1 | [67] | |
| GC | BARHL2 | Cell differentiation | [69] | |
| LncUEGC1 | Unknown | [70] | ||
| LncRNA HOTTIP | Proliferation promotion and apoptosis inhibition | [71] | ||
| FZD10 | Increasing of Ki-67 expression via Phospho-ERK1/2 | [20] | ||
| Colon | IBD | Annexin A1 | Inflammation reduction | [75,84] |
| miR-200b-3p NEAT lncRNA | Protein expression regulation | [76,77] | ||
| PSMA7 | Role in proteosomal activity | [73] | ||
| lncRNA H19 | Unknown | [72] | ||
| CRC | FZD10 | Increasing of Ki-67 expression via Phos-pho-ERK1/2 | [20] | |
| CD147 | Glycolipid metabolism reprogrammation | [18] | ||
| A33 | Cell surface targeting for antibody-based therapy | [81] | ||
| MARCKSL1 | Role in the Immune System | [82] | ||
| Wnt | Role in via β-catenin/SOX2 | [79] | ||
| CPNE3 | Promotion of cell proliferation via the PI3K/AKT pathway | [80] |
| Organ | Pathology | EV-Based Preparations and Applications | References |
|---|---|---|---|
| liver | HCC | EVs released by BM-MSCs as a delivery system to convey siCD38 to HCC cells: potential limitation of HCC growth and spread | [134] |
| Engineered EVs mediated CRISPR-Cas9 to reverse the therapy resistance of sorafenib: the suppression of HCC cancer stem cells HLC9-EVs engineered with HN3: increasing in specific homing | [135,141,142] | ||
| pancreas | PDAC PC | Delivery system based on CR-modified EVs for PTT | [145] |
| EVs derived from HUCMSCs loaded with miR-145-5p: apoptosis and cancer growth inhibition | [146] | ||
| EVs derived from HUCMSCs loaded with hsa-miRNA-128–3p: apoptosis and cancer growth inhibition | [147] | ||
| EVs derived from HUCMSCs loaded with KRAS G12D targeting siRNA: a reduction in the proliferation, migration, and viability in PANC-1 cells | [148] | ||
| HUCMSC-EVs with DOXO: the rapid uptake in PANC-1 cells induced apoptotic cell death more efficiently than free DOXO | |||
| RWP-1-derived EVs loaded with temozolomide and EPZ015666: a greater antiproliferative action of EVs encapsulating on the PC cell line EVs derived from the hTERT-HPNE cell line and incubated with PTX: PDAC cell death via clathrin-mediated endocytosis | [149,150] | ||
| stomach | GC | EVs from HEK-293 cells targeting CDH17-positive GC cancers by the engineered nanobodies into EVs with genetic engineering techniques | [151,152,153,154] |
| Engineered small EVs with high CD47 expression and c(RGDyC) modification to deliver short RNAi against YTHDF1: a novel therapeutic strategy for GC via epigenetic and immune regulation | [155,166] | ||
| colon | IBD | EVs released by MSCs and loaded with miR-146: a new strategy in the treatment of IBD in mouse models | [126] |
| EVs and bacteria-derived membrane vesicles (OMVs) derived from normal feces: restore the intestinal barrier in IBD subjects | [38,127,128,167] | ||
| Exo containing PSMA7 derived from saliva: an important protein biomarker for IBD | [168] | ||
| CRC | Folic acid-modified exo–liposome hybrid nanoparticles loaded with ALKBH5 mRNA: a novel therapeutic strategy for CRC | [156] | |
| EVs derived from CT26 and loaded with miR-34a mimics: a new strategy for CRC gene therapy in mouse model | [159] | ||
| EVs derived from milk: a reduction in primary tumor burden in mouse models of CRC and breast cancer | [161,162] | ||
| Exosomes secreted from HEK293T transfected with Her2-mCherry plasmid and loaded with both 5-FU and miR-21i: reversing the drug resistance in CRC cells | [164,165] | ||
| Exosomes secreted from MSCs modified with 5TR1 or with magnetic nanoparticles, folic acid, and DOXO: the inhibition of CRC growth in mouse models | [151,152,153,154] |
| Study Title | Condition or Disease | EVs for Biomarker Detection | EVs for Therapy | ClinicalTrials.gov Identifier |
|---|---|---|---|---|
| Portal Hypertension in Non-alcoholic Fatty Liver Disease: Association with Cardiovascular Risk and Identification of non-invasive biomarkers | NAFLD | Identification of non-invasive biomarkers, including EVs, and of the presence and severity of portal hypertension by liquid biopsy | NCT04191044 | |
| Liver Health and Metabolic Func-tion in People with Obesity | NAFLD | Amount, content, and function of EVs from adipose, liver, and blood tissue collected before and after ~20% weight loss and at the time of surgery | NCT03701828 | |
| Role of Immune System in Obesity-related Inflammation and Cardiometabolic Risk | NAFLD MASLD | Investigation of the signaling between cells and organs examined by isolating the exosomes from the blood, subcutaneous and omental adipose tissue of patients scheduled for gallbladder, inguinal hernia, hysterectomy, or myomectomy surgery | NCT01104220 | |
| Muscles in Liver Diseases | Liver Diseases | Identification of circulating mediators, including EVs, that could be responsible for the complications of liver disease: EVs released by the muscle and acting on different organs | NCT04758793 | |
| Safety of Injection of Placental MSC-Derived Exosomes for Treatment of Resistant Perianal Fistula in Crohn’s Patients | Perianal Fistula Crohn’s Disease | Identification of inflammatory markers in exosomes through laboratory workup, including CRP, IL-6, TNF-a, calprotectin | Evaluation of the safety and clinical efficacy of injected exosomes | NCT05499156 |
| Study of ExoFlo for the Treatment of Perianal Fistulas | Perianal Fistula Crohn’s Disease | Ex Vivo Culture-expanded Adult Allogeneic Bone Marrow MSC-derived EVs for the Treatment of Perianal Fistulizing Crohn’s Disease Local injection of normal saline | NCT05836883 | |
| Study of ExoFlo for the Treatment of Medically refractory Crohn’s Disease | Crohn’s Disease | Intravenous ex vivo culture-expanded adult allogeneic bone marrow MSC-EVs in subjects with medically refractory Crohn’s disease who do not respond to monoclonal antibody-based therapy: evaluation of feasibility and efficacy | NCT05130983 | |
| Study to Evaluate the Epidemiology and the Characteristics “Omics” in Patients Recently Diagnosed of Inflammatory Bowel Disease in Spain (IBDomics) | Inflammatory Bowel Diseases | Characterization of the composition of serum EVs in newly diagnosed IBD patients, aiming to identify the molecular and cellular pathways involved in IBD development and pathogenesis correlation of the serum proteomic profile and the density and composition of serum EVs with an IBD phenotype | NCT03689257 | |
| Study of ExoFlo for the Treatment of Medically Refractory Ulcerative Colitis | Ulcerative Colitis Inflammatory Bowel Diseases | Intravenous ex vivo culture-expanded adult allogeneic bone marrow MSC-EVs in subjects with medically refractory Crohn’s disease who do not respond to monoclonal antibody-based therapy: evaluation of feasibility, safety, and efficacy | NCT05176366 | |
| Pancreatic Cancer Initial Detection Via Liquid Biopsy (PANCAID) | Pancreatic Cancer Chronic | Detection of pancreatic cancer in bio-banked samples, including EVs, of patients with histologically confirmed pancreatic lesions via liquid biopsy | NCT06283576 | |
| Impact of Graft Reconditioning with Hypothermic Machine Perfusion on HCC Recurrence After Liver Transplantation | Hepatocellular Carcinoma | Identification of microRNAs from liquid-biopsy-derived EVs, as tools of prognostic information on IRI favoring hepatocellular carcinoma recurrence | NCT06236568 | |
| Metronomic Capecitabine, Oxaliplatin and UGT1A1 Genotype-directed Irinotecan in Metastatic Pancreatic Cancer Patients | Metastatic Pancreatic Cancer | Identification of exosomal proteins secreted by EVs from plasma using mass spectrometry, at different times: at pre-dose, the end of oxaliplatin infusion, and the end of irinotecan infusion | NCT05929885 | |
| A Prospective Feasibility Study Evaluating EVs Obtained by Liquid Biopsy for Neoadjuvant Treatment Response Assessment in Rectal Cancer | Rectal Cancer | Tumor-EV detection and quantification for the response assessment and follow-up of patients with adenocarcinoma of the rectum | NCT04852653 | |
| Contents of Circulating Extracellular Vesicles: Biomarkers in Colorectal Cancer Patients | Colorectal Cancer | Exploitation via liquid biopsy of circulating tumor-exosomes containing markers, namely specific miRNAs, useful as biomarkers of the early prognosis of patients with colon cancer | NCT04523389 | |
| Exosome-based Liquid Biopsies for Upper Gastrointestinal Cancers Diagnosis | Gastric Cancer Esophagus Cancer | Identification and quantification via proteomic analysis of specific proteins as biomarkers for upper gastrointestinal tumors Multicentre and retrospective study | NCT06278064 | |
| Prospectively Predict the Efficacy of Treatment of Gastrointestinal Tumors Based on Peripheral Multi-omics Liquid Biopsy | Advanced Gastric Adenocarcinoma | Validation of 4 plasma EV-derived proteins and their combination as a signature score for robustly predicting immunotherapeutic outcomes Prospective study | NCT04993378 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrè, V.; Mastrogiacomo, R.; Balestra, F.; Serino, G.; Viti, F.; Rizzi, F.; Curri, M.L.; Giannelli, G.; Depalo, N.; Scavo, M.P. Unveiling the Potential of Extracellular Vesicles as Biomarkers and Therapeutic Nanotools for Gastrointestinal Diseases. Pharmaceutics 2024, 16, 567. https://doi.org/10.3390/pharmaceutics16040567
Arrè V, Mastrogiacomo R, Balestra F, Serino G, Viti F, Rizzi F, Curri ML, Giannelli G, Depalo N, Scavo MP. Unveiling the Potential of Extracellular Vesicles as Biomarkers and Therapeutic Nanotools for Gastrointestinal Diseases. Pharmaceutics. 2024; 16(4):567. https://doi.org/10.3390/pharmaceutics16040567
Chicago/Turabian StyleArrè, Valentina, Rita Mastrogiacomo, Francesco Balestra, Grazia Serino, Federica Viti, Federica Rizzi, Maria Lucia Curri, Gianluigi Giannelli, Nicoletta Depalo, and Maria Principia Scavo. 2024. "Unveiling the Potential of Extracellular Vesicles as Biomarkers and Therapeutic Nanotools for Gastrointestinal Diseases" Pharmaceutics 16, no. 4: 567. https://doi.org/10.3390/pharmaceutics16040567