
Decoding Post-Viral Fatigue: The Basal Ganglia’s Complex Role in Long-COVID


Abstract
1. Introduction
2. Basal Ganglia Dysfunction in Long-COVID Fatigue: Evidence from Neuroimaging
2.1. Evidence of Metabolic Dysfunction
2.2. Functional Connectivity Disruptions
2.3. Mismatch in Structural Architecture
3. Theories on Causative Mechanisms
3.1. Inflammation-Induced Dopamine Signaling Dysfunction
3.2. Disruption of Cortical-Striatal Motivational Pathways
3.3. Loss of Excitatory Basal Ganglia Input to Arousal Centers
4. Fatigue-Related Symptoms and Basal Ganglia Dysfunction
5. Therapeutic Opportunities
6. The Challenges of Long-COVID Fatigue Research
7. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- World Health Organization. A Clinical Case Definition of Post-COVID-19 Condition by a Delphi Consensus. World Health Organization. 2021. Available online: https://www.who.int/publications/i/item/WHO-2019-nCoV-Post_COVID-19_condition-Clinical_case_definition-2021.1 (accessed on 6 October 2021).
- Sudre, C.H.; Murray, B.; Varsavsky, T.; Graham, M.S.; Penfold, R.S.; Bowyer, R.C.; Pujol, J.C.; Klaser, K.; Antonelli, M.; Canas, L.S.; et al. Attributes and predictors of long-COVID. Nat. Med. 2021, 27, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Taquet, M.; Geddes, J.R.; Husain, M.; Luciano, S.; Harrison, P.J. 6-month neurological and psychiatric outcomes in 236,379 survivors of COVID-19: A retrospective cohort study using electronic health records. Lancet Psychiatry 2021, 8, 416–427. [Google Scholar] [CrossRef] [PubMed]
- Tenforde, M.W.; Kim, S.S.; Lindsell, C.J.; Rose, E.B.; Shapiro, N.I.; Files, D.C.; Gibbs, K.W.; Erickson, H.L.; Steingrub, J.S.; Smithline, H.A.; et al. Symptom duration and risk factors for delayed return to usual health among outpatients with COVID-19 in a multistate health care systems network—United States, March–June 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, O.; Chan, M.; Charron, C.; Chakravarty, M.M. Basal Ganglia Dysfunction in Long-COVID: A Cross-Sectional Association Study. Lancet 2021, 6, 100134. [Google Scholar]
- Graham, E.L.; Clark, J.R.; Orban, Z.S.; Lim, P.H.; Szymanski, A.L.; Taylor, C.; DiBiase, R.M.; Tong Jia, D.; Balabanov, R.; Koralnik, I.J.; et al. Persistent neurologic symptoms and cognitive dysfunction in non-hospitalized COVID-19 “long haulers”. Ann. Clin. Transl. Neurol. 2021, 8, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Paprocka, J.; Machnikowska-Sokołowska, M.; Gruszczyńska, K.; Emich-Widera, E. Neuroimaging of Basal Ganglia in Neurometabolic Diseases in Children. Brain Sci. 2020, 10, 849. [Google Scholar] [CrossRef]
- Huang, Y.; Tan, C.; Wu, J.; Chen, M.; Wang, Z.; Luo, L.; Zhou, X.; Liu, X.; Huang, X.; Yuan, S.; et al. Impact of coronavirus disease 2019 on pulmonary function in early convalescence phase. Respir. Res. 2021, 22, 163. [Google Scholar]
- Ferrucci, R.; Cuffaro, L.; Capozza, A.; Rosci, C.; Maiorana, N.; Groppo, E.; Reitano, M.R.; Poletti, B.; Ticozzi, N.; Tagliabue, L.; et al. Brain positron emission tomography (PET) and cognitive abnormalities one year after COVID-19. J. Neurol. 2023, 270, 1823–1834. [Google Scholar] [CrossRef]
- Douaud, G.; Lee, S.; Alfaro-Almagro, F.; Arthofer, C.; Wang, C.; McCarthy, P.; Lange, F.; Andersson, J.; Griffanti, L.; Smith, S.M.; et al. Brain imaging before and after COVID-19 in UK Biobank. Nature 2022, 604, 697–707. [Google Scholar] [CrossRef]
- Hannanu, F.F.; Clément, J.; Morelot-Panzini, C. Brain metabolic impairment in patients with persisting symptoms following SARS-CoV-2 infection: A [18F]FDG-PET study. Eur. J. Nucl. Med. Mol. Imaging 2022, 49, 588–600. [Google Scholar]
- Lu, Y.; Li, X.; Geng, D.; Mei, N.; Wu, P.Y.; Huang, C.C.; Jia, T.; Zhao, Y.; Wang, D.; Xiao, A.; et al. Cerebral micro-structural changes in COVID-19 patients—An MRI-based 3-month follow-up study. eClinicalMedicine 2020, 25, 100484. [Google Scholar] [CrossRef] [PubMed]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Meziani, F.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2021, 382, 2268–2270. [Google Scholar] [CrossRef]
- Kandemirli, S.G.; Dogan, L.; Sarikaya, Z.T.; Kara, S.; Akinci, C.; Kaya, D.; Kaya, Y.; Yildirim, D.; Tuzuner, F.; Yildirim, M.S.; et al. Brain MRI findings in patients in the intensive care unit with COVID-19 infection. Radiology 2020, 297, E232–E235. [Google Scholar]
- Varatharaj, A.; Thomas, N.; Ellul, M.A.; Davies NW, S.; Pollak, T.A.; Tenorio, E.L.; Sultan, M.; Easton, A.; Breen, G.; Zandi, M.; et al. Neurological and neuropsychiatric complications of COVID-19 in 153 patients: A UK-wide surveillance study. Lancet Psychiatry 2020, 7, 875–882. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ling, Q.; Manyande, A.; Wu, D.; Xiang, B. Brain Imaging Changes in Patients Recovered From COVID-19: A Narrative Review. Front. Neurosci. 2022, 16, 855868. [Google Scholar] [CrossRef]
- Luo, Y.; Gander, P.; Wang, Y.; Ponto, L.; Rudroff, T. Long-COVID-19 patients exhibited decreased globus pallidus activity independent of fatigue status—An exploratory FDG-PET study. In Proceedings of the Society for Neuroscience Annual Meeting, Washington, DC, USA, 11–15 November 2023. [Google Scholar]
- Hampshire, A.; Trender, W.; Chamberlain, S.R.; Jolly, A.E.; Grant, J.E.; Patrick, F.; Mazibuko, N.; Williams, S.C.; Barnby, J.M.; Hellyer, P.; et al. Cognitive deficits in people who have recovered from COVID-19. eClinicalMedicine 2021, 39, 101044. [Google Scholar] [CrossRef] [PubMed]
- Meinhardt, J.; Radke, J.; Dittmayer, C.; Franz, J.; Thomas, C.; Mothes, R.; Laue, M.; Schneider, J.; Brünink, S.; Greuel, S.; et al. Olfactory transmucosal SARS-CoV-2 invasion as a port of central nervous system entry in individuals with COVID-19. Nat. Neurosci. 2021, 24, 168–175. [Google Scholar] [CrossRef]
- Zhou, F.; Tao, M.; Shang, L.; Liu, Y.; Pan, G.; Jin, Y.; Wang, L.; Hu, S.; Li, J.; Zhang, M.; et al. Assessment of sequelae of COVID-19 nearly 1 year after diagnosis. Front. Med. 2021, 8, 717194. [Google Scholar] [CrossRef]
- Hafiz, R.; Gandhi, T.K.; Mishra, S.; Prasad, A.; Mahajan, V.; Di, X.; Natelson, B.H.; Biswal, B.B. Higher limbic and basal ganglia volumes in surviving COVID-negative patients and the relations to fatigue. Neuroimage Rep. 2022, 2, 100095. [Google Scholar] [CrossRef]
- Heine, J.; Schwichtenberg, K.; Hartung, T.J.; Rekers, S.; Chien, C.; Boesl, F.; Rust, R.; Hohenfeld, C.; Bungenberg, J.; Finke, C.; et al. Structural brain changes in patients with long-COVID fatigue: A prospective observational study. eClinicalMedicine 2023, 58, 101874. [Google Scholar] [CrossRef]
- Deters, J.R.; Fietsam, A.C.; Gander, P.E.; Boles Ponto, L.L.; Rudroff, T. Effect of Long-COVID-19 on Brain Volume and Glucose Metabolism: Influence of Time Since Infection and Fatigue Status. Brain Sci. 2023, 13, 675. [Google Scholar] [CrossRef]
- Trifilieff, P.; Feng, B.; Urizar, E.; Winiger, V.; Ward, R.D.; Taylor, K.M.; Martinez, D.; Moore, H.; Balsam, P.D.; Simpson, E.H.; et al. Increasing dopamine D2 receptor expression in the adult nucleus accumbens enhances motivation. Mol. Psychiatry 2013, 18, 1025–1033. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M.; Farrar, A.; Mingote, S.M. Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology 2007, 191, 461–482. [Google Scholar] [CrossRef] [PubMed]
- Berke, J.D. What does dopamine mean? Nat. Neurosci. 2018, 21, 787–793. [Google Scholar] [CrossRef]
- Yamamoto, B.K.; Zhu, W. The effects of methamphetamine on the production of free radicals and oxidative stress. J. Pharmacol. Exp. Ther. 1998, 287, 107–114. [Google Scholar]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036–1046. [Google Scholar] [CrossRef]
- Yohn, S.E.; Thompson, C.; Randall, P.A.; Lee, C.A.; Müller, C.E.; Baqi, Y.; Correa, M.; Salamone, J.D. The VMAT-2 inhibitor tetrabenazine alters effort-related decision making as measured by the T-maze barrier choice task: Reversal with the adenosine A2A antagonist MSX-3 and the catecholamine uptake blocker bupropion. Psychopharmacology 2015, 232, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Kobayashi, S.; Numata, T.; Inagaki, N. Cytokine alteration of dopamine transporter: A possible causative factor in COVID-19 related fatigue. Med. Hypotheses 2021, 153, 110734, Epub ahead of print. [Google Scholar]
- Hauser, T.U.; Fiore, V.G.; Moutoussis, M.; Dolan, R.J. Computational Psychiatry of ADHD: Neural Gain Impairments across Marrian Levels of Analysis. Trends Neurosci. 2016, 39, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Haber, S.N.; Behrens, T.E.J. The neural network underlying incentive-based learning: Implications for interpreting circuit disruptions in psychiatric disorders. Neuron 2014, 83, 1019–1039. [Google Scholar] [CrossRef]
- Le Heron, C.; Apps, M.A.J.; Husain, M. The anatomy of apathy: A neurocognitive framework for amotivation in psychopathology. Neurosci. Biobehav. Rev. 2020, 118, 550–578. [Google Scholar]
- Da Cunha, C.; Gomez-A, A.; Blaha, C.D. The role of the basal ganglia in motivated behavior. Rev. Neurosci. 2012, 23, 747–767. [Google Scholar] [CrossRef] [PubMed]
- Do, J.P.; Guo, D.; Ming, G.L.; Song, H. BDNF upregulation rescues NMDA-induced alterations in synaptic plasticity in young and aged hippocampal dentate gyrus granule neurons. Learn. Mem. 2012, 19, 89–96. [Google Scholar]
- Guo, Z.V.; Inagaki, H.K.; Daie, K.; Druckmann, S.; Gerfen, C.R.; Svoboda, K. Maintenance of persistent activity in a frontal thalamocortical loop. Nature 2015, 526, 181–186. [Google Scholar] [CrossRef]
- Qiu, M.H.; Vetrivelan, R.; Fuller, P.M.; Lu, J. Basal ganglia control of sleep-wake behavior and cortical activation. Eur. J. Neurosci. 2010, 31, 499–507. [Google Scholar] [CrossRef]
- Lazarus, M.; Chen, J.F.; Urade, Y.; Huang, Z.-L. Role of the basal ganglia in the control of sleep and wakefulness. Curr. Opin. Neurobiol. 2013, 23, 780–785. [Google Scholar] [CrossRef]
- Paterson, R.W.; Brown, R.L.; Benjamin, L.; Nortley, R.; Wiethoff, S.; Bharucha, T.; Jayaseelan, D.L.; Kumar, G.; Raftopoulos, R.E.; Zambreanu, L.; et al. The emerging spectrum of COVID-19 neurology: Clinical, radiological and laboratory findings. Brain 2022, 143, 3104–3120. [Google Scholar] [CrossRef]
- Rudroff, T.; Fietsam, A.C.; Deters, J.R.; Bryant, A.D.; Kamholz, J. Long-COVID-19 Fatigue: Potential Contributing Factors. Brain Sci. 2020, 10, 1012. [Google Scholar] [CrossRef]
- Havervall, S.; Rosell, A.; Phillipson, M.; Mangsbo, S.M.; Nilsson, P.; Hober, S.; Thålin, C. Symptoms and Functional Impairment Assessed 8 Months After Mild COVID-19 Among Health Care Workers. JAMA 2021, 325, 2015–2016. [Google Scholar] [CrossRef] [PubMed]
- Groff, D.; Sun, A.; Ssentongo, A.E.; Ba, D.M.; Parsons, N.; Poudel, G.R.; Lekoubou, A.; Oh, J.S.; Ericson, J.E.; Ssentongo, P.; et al. Short-term and Long-term Rates of Postacute Sequelae of SARS-CoV-2 Infection: A Systematic Review. JAMA Netw. Open 2021, 4, e2128568. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; Assaf, G.S.; McCorkell, L.; Wei, H.; Low, R.J.; Re’em, Y.; Redfield, S.; Austin, J.P.; Akrami, A. Characterizing long-COVID in an international cohort: 7 months of symptoms and their impact. eClinicalMedicine 2021, 38, 101019. [Google Scholar] [CrossRef]
- Behnood, S.A.; Shafran, R.; Bennett, S.D.; Zhang, A.X.D.; O’Mahoney, L.L.; Stephenson, T.J.; Ladhani, S.N.; De Stavola, B.L.; Viner, R.M.; Swann, O.V. Persistent symptoms following SARS-CoV-2 infection amongst children and young people: A meta-analysis of controlled and uncontrolled studies. J. Infect. 2022, 84, 158–170. [Google Scholar] [CrossRef]
- Lopez-Leon, S.; Wegman-Ostrosky, T.; Perelman, C.; Sepulveda, R.; Rebolledo, P.A.; Cuapio, A.; Villapol, S. More than 50 Long-term effects of COVID-19: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 16144. [Google Scholar] [CrossRef] [PubMed]
- Natelson, B.H. Post-infectious fatigue. In Brain and Behavior: Plasticity, Compensation and Recovery; Conti, F., Priori, A., Grassi, C., Eds.; Springer: Cham, Switzerland, 2006; Volume 1226. [Google Scholar]
- Sigfrid, L.; Drake, T.M.; Pauley, E.; Jesudason, E.C.; Olliaro, P.; Lim, W.S.; Gillesen, A.; Berry, C.; Lowe, D.J.; McPeake, J.; et al. Long-COVID in adults discharged from UK hospitals after COVID-19: A prospective, multicenter cohort study using the ISARIC WHO Clinical Characterization Protocol. Lancet Reg. Health Eur. 2021, 8, 100186. [Google Scholar] [CrossRef]
- Gemelli Against COVID-19 Post-Acute Care Study Group. Long-COVID-19 global health strategies: The need for an interdisciplinary approach. Aging Clin. Exp. Res. 2020, 32, 1613–1620. [Google Scholar] [CrossRef]
- Brosschot, J.F.; Gerin, W.; Thayer, J.F. The perseverative cognition hypothesis: A review of worry, prolonged stress-related physiological activation, and health. J. Psychosom. Res. 2006, 60, 113–124. [Google Scholar] [CrossRef]
- Thomas, F. Stigma, fatigue and social breakdown: Exploring the impacts of HIV/AIDS on patient and carer well-being in the Caprivi Region, Namibia. Soc. Sci. Med. 2006, 63, 3174–3187. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.J.; Webb, T.L.; Rowse, G. Does improving sleep lead to better mental health? A protocol for a meta-analytic review of randomized controlled trials. BMJ Open 2017, 7, e016873. [Google Scholar] [CrossRef]
- Moturu, S.T.; Khayal, I.; Aharony, N.; Pan, J.; Pentland, A. Using social sensing to understand the links between sleep, mood, and stress. In Proceedings of the 2011 Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Boston, MA, USA, 9–11 October 2011. [Google Scholar]
- Fishbain, D.A.; Cole, B.; Cutler, R.B.; Lewis, J.; Rosomoff, H.L.; Fosomoff, R.S. Is pain fatiguing? A structured evidence-based review. Pain Med. 2003, 4, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kadetoff, D.; Kosek, E. The effects of static muscular contraction on blood pressure, heart rate, pain ratings and pressure pain thresholds in healthy individuals and patients with fibromyalgia. Eur. J. Pain 2007, 11, 39–47. [Google Scholar] [CrossRef]
- Bower, J.E. Cancer-related fatigue--mechanisms, risk factors, and treatments. Nat. Rev. Clin. Oncol. 2014, 11, 597–609. [Google Scholar] [CrossRef]
- Manjaly, Z.M.; Harrison, N.A.; Critchley, H.D.; Do, C.T.; Stefanics, G.; Wenderoth, N.; Lutterotti, A.; Müller, A.; Stephan, K.E. Pathophysiological and cognitive mechanisms of fatigue in multiple sclerosis. J. Neurol. Neurosurg. Psychiatry 2019, 90, 642–651. [Google Scholar] [CrossRef]
- Skrzypczak-Wiercioch, A.; Sałat, K. Lipopolysaccharide-Induced Model of Neuroinflammation: Mechanisms of Action, Research Application and Future Directions for Its Use. Molecules 2022, 27, 5481. [Google Scholar] [CrossRef]
- Ledford, H. Long COVID trial provides tentative hope for treatment. Nature 2024, 601, 356–357. [Google Scholar]
- Sanghavi, D.; Bansal, P.; Kaur, I.P.; Mughal, M.S.; Keshavamurthy, C.; Cusick, A.; Schram, J.; Yarrarapu, S.N.S.; Giri, A.R.; Kaur, N.; et al. Impact of colchicine on mortality and morbidity in COVID-19: A systematic review. Ann. Med. 2022, 54, 775–789. [Google Scholar] [CrossRef]
- Saucier, J.; Comeau, D.; Robichaud, G.A.; Chamard-Witkowski, L. Reactive gliosis and neuroinflammation: Prime suspects in the pathophysiology of post-acute neuroCOVID-19 syndrome. Front. Neurol. 2023, 14, 1221266. [Google Scholar] [CrossRef]
- Salamone, J.D.; Correa, M.; Farrar, A.M.; Nunes, E.J.; Pardo, M. Dopamine, behavioral economics, and effort. Front. Behav. Neurosci. 2009, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Treadway, M.T.; Bossaller, N.A.; Shelton, R.C.; Zald, D.H. Effort-based decision-making in major depressive disorder: A translational model of motivational anhedonia. J. Abnorm. Psychol. 2012, 121, 553–558. [Google Scholar] [CrossRef]
- Downs, B.W.; Blum, K.; Bagchi, D.; Kushner, S.; Bagchi, M.; Galvin, J.M.; Lewis, M.; Siwicki, D.; Brewer, R.; Boyett, B.; et al. Molecular neurobiological and systemic health benefits of achieving dopamine homeostasis in the face of a catastrophic pandemic (COVID- 19): A mechanistic exploration. J. Syst. Integr. Neurosci. 2020, 7. [Google Scholar] [CrossRef]
- Zhang, J.X.; Zhang, J.J. Case Report of Improvement in Long-COVID Symptoms in an Air Force Medic Treated With Transcranial Magnetic Stimulation Using Electro-Magnetic Brain Pulse Technique. Mil. Med. 2023, 188, e3711–e3715. [Google Scholar] [CrossRef]
- Linnhoff, S.; Koehler, L.; Haghikia, A.; Zaehle, T. The therapeutic potential of non-invasive brain stimulation for the treatment of Long-COVID-related cognitive fatigue. Front. Immunol. 2023, 13, 935614. [Google Scholar] [CrossRef]
- Noda, Y.; Sato, A.; Shichi, M.; Sato, A.; Fujii, K.; Iwasa, M.; Nagano, Y.; Kitahata, R.; Osawa, R. Real world research on transcranial magnetic stimulation treatment strategies for neuropsychiatric symptoms with long-COVID in Japan. Asian J. Psychiatr. 2023, 81, 103438. [Google Scholar] [CrossRef] [PubMed]
- Roet, M.; Boonstra, J.; Sahin, E.; Mulders, A.E.P.; Leentjens, A.F.G.; Jahanshahi, A. Deep Brain Stimulation for Treatment-Resistant Depression: Towards a More Personalized Treatment Approach. J. Clin. Med. 2020, 9, 2729. [Google Scholar] [CrossRef]
- Chase, A. Therapeutic neuromodulation for post-viral fatigue syndromes. Brain Stimul. 2023, 16, 25–31. [Google Scholar]
- Rudroff, T. The challenges of long-COVID-19 fatigue research. Front. Psychol. 2023, 14, 1120928. [Google Scholar] [CrossRef] [PubMed]
- Fietsam, A.C.; Bryant, A.D.; Rudroff, T. Fatigue and perceived fatigability, not objective fatigability, are prevalent in people with long-COVID-19. Exp. Brain Res. 2023, 241, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Rudroff, T. Revealing the Complexity of Fatigue: Review of Persistent Challenges and Promises of Artificial Intelligence. Brain Sciences. Brain Sci. 2024, 14, 186. [Google Scholar] [CrossRef] [PubMed]
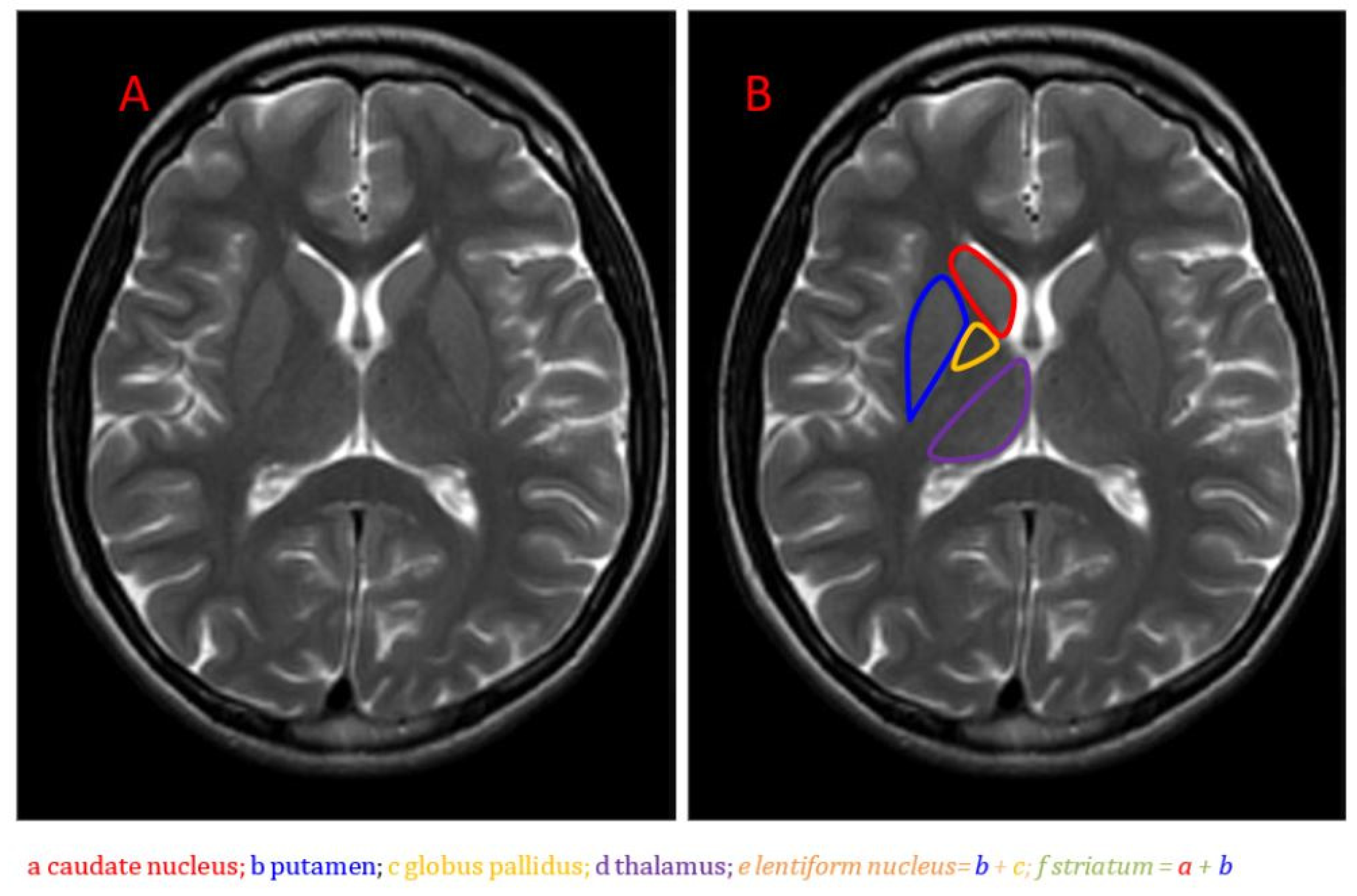

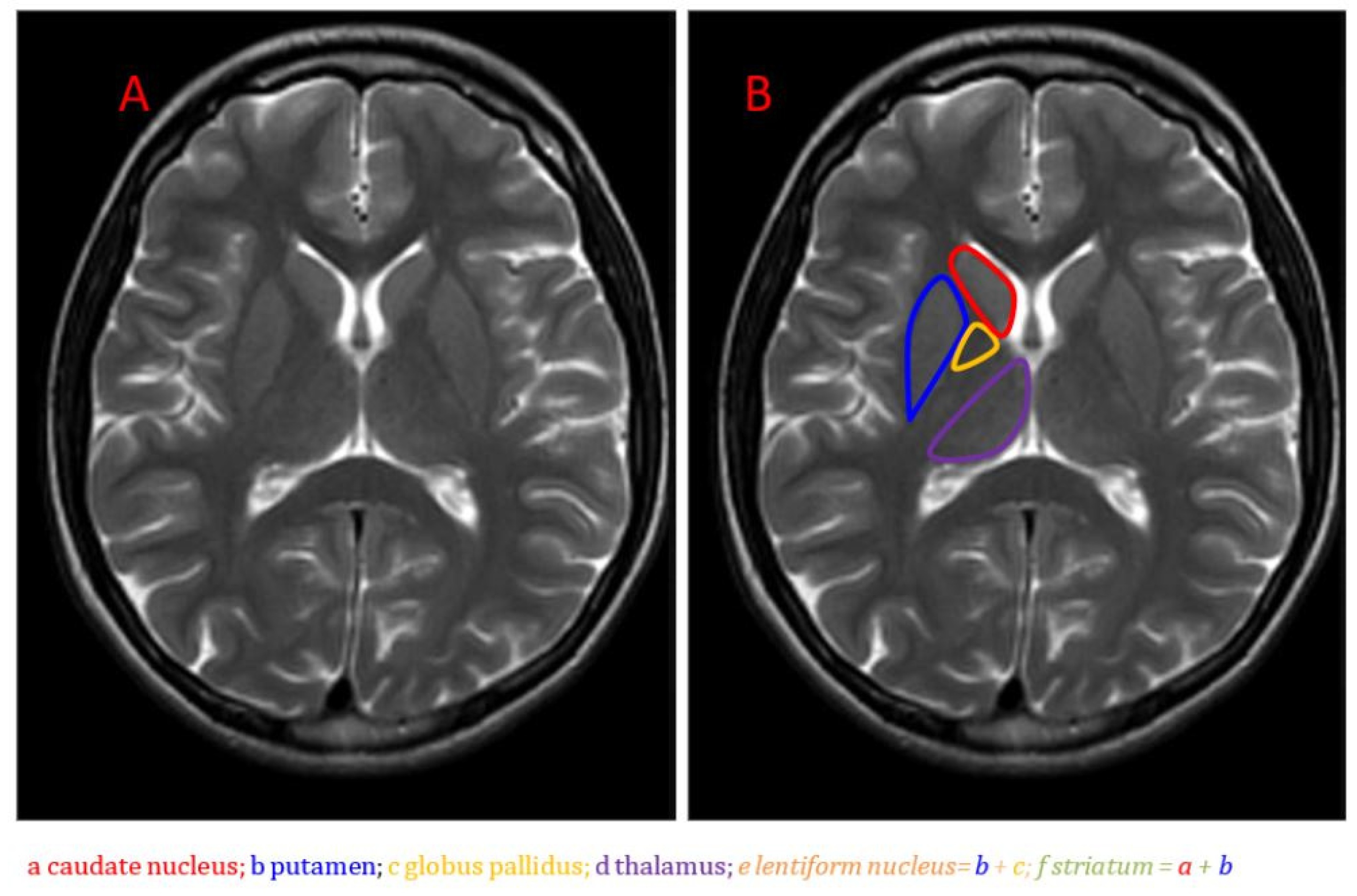{kind=link}
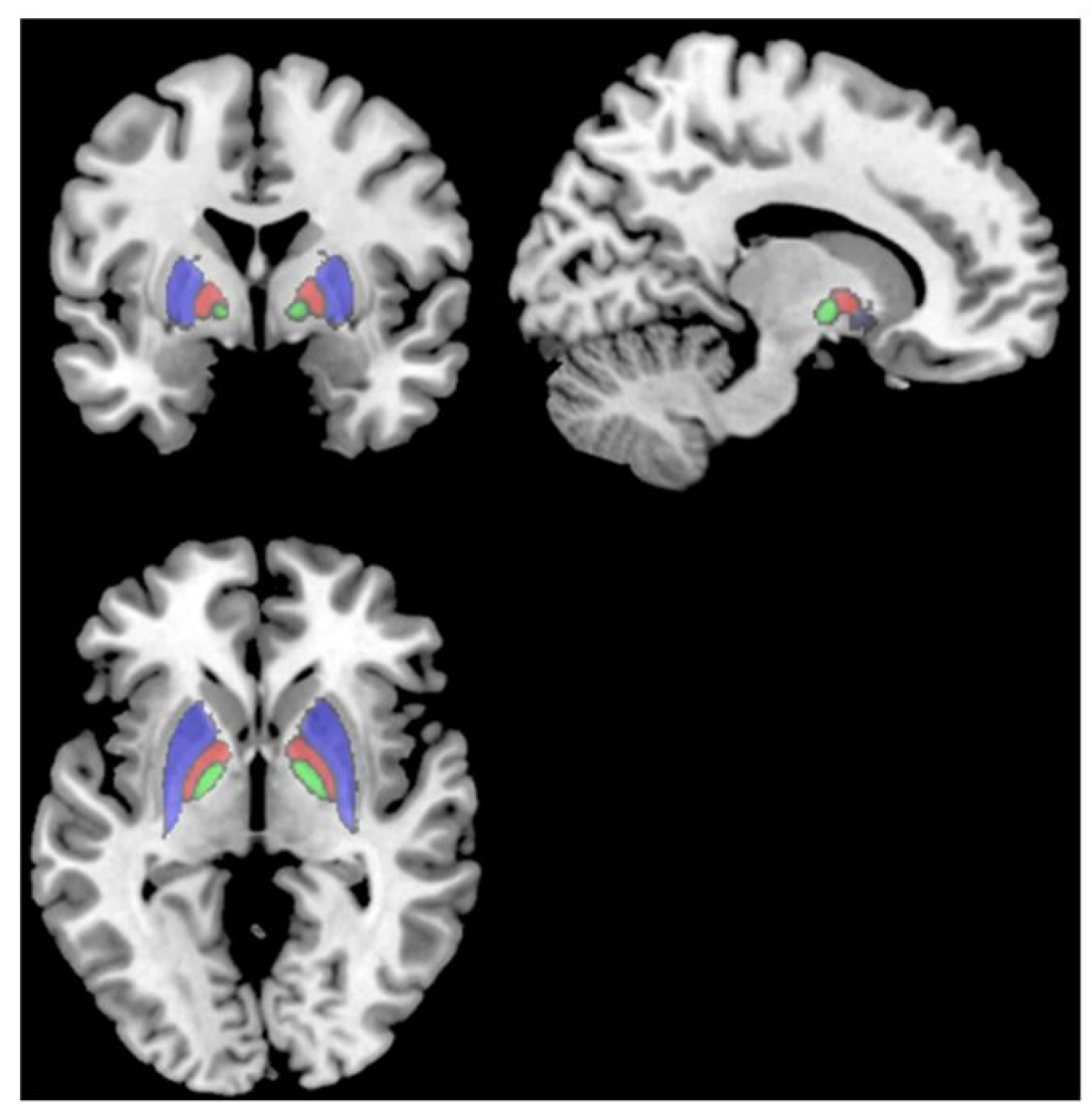
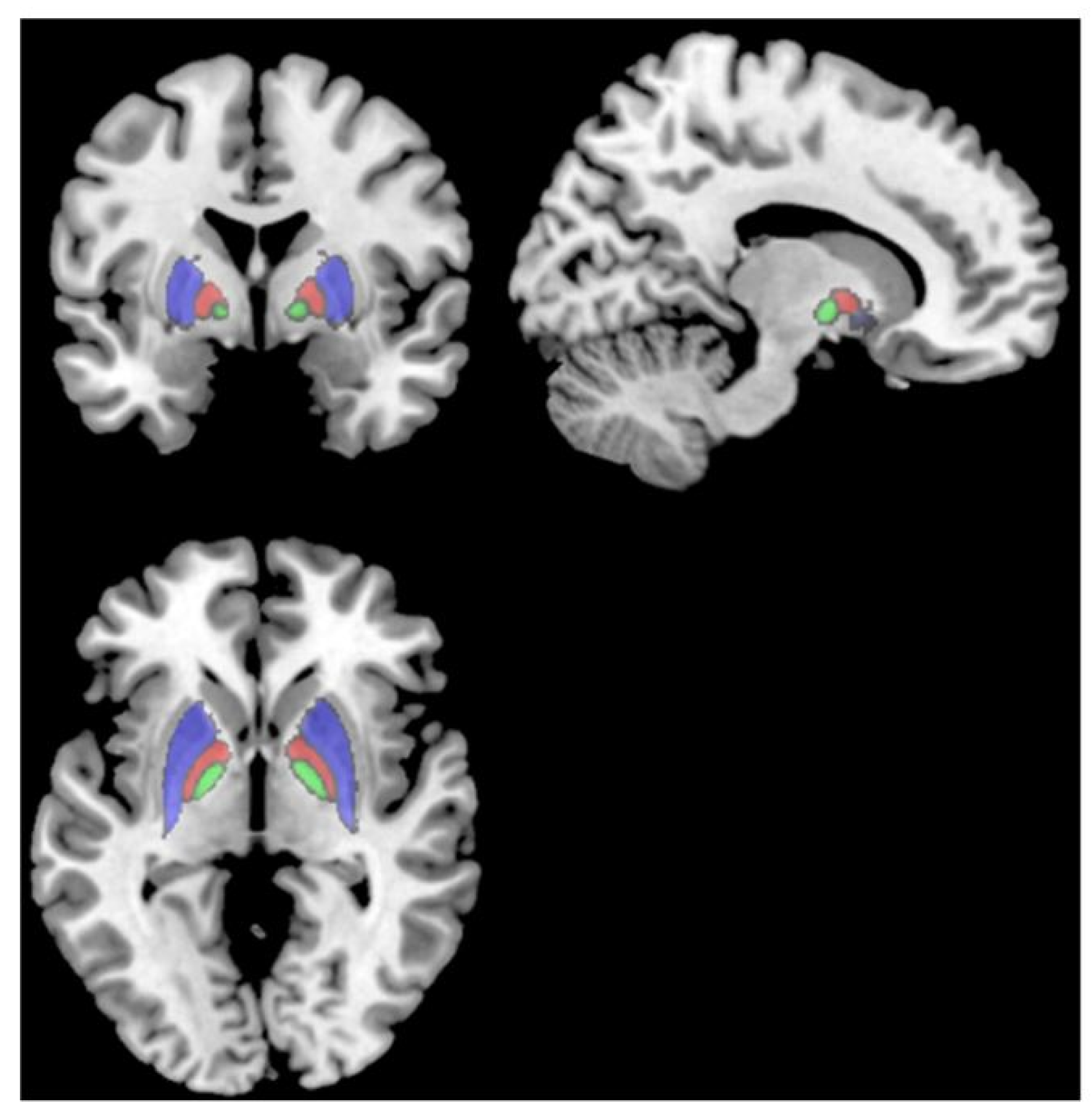{kind=link}
| Reference | Subjects | Disease Duration | Comorbidities | Hospitalization | Observed Brain Changes |
|---|---|---|---|---|---|
| Douaud et al., [10] | 600+ SARS-CoV-2 infected | Not specified | Cognitive impairment | Not specified | Connectivity weakening between basal ganglia, thalami, and sensory regions |
| Lu et al., [12] | Long-COVID patients vs. controls | 10 months | Hypertension, memory loss, headache, tremor, impaired mobility, myalgia | Hospitalized | Striatal neuroinflammation compromised neuronal integrity in caudate and putamen |
| Kandemirli et al., [14] | ICU COVID-19 patients | Not specified | Hypertension, diabetes mellitus, cerebrovascular accident, chronic kidney disease, coronary artery disease | In ICU | 56% had basal ganglia hyperintensities, reduced NAA/Cr and NAA/Cho ratios |
| Helms et al., [13] | 58 Severe COVID-19 patients | Not specified | Confusion, cognitive dysfunction | Hospitalized | Basal ganglia hyperintensities (MRI) |
| Hampshire et al., [18] | Recovered COVID-19 patients | Not specified | Anxiety, depression, lung conditions, psychiatric conditions | Hospitalized | Cognitive deficits linked to disrupted basal ganglia communication |
| Meinhardt et al., [19] | 33 deceased COVID-19 patients | Not specified | Not specified | Not specified | Olfactory SARS-CoV-2 invasion as path to central nervous system |
| Zhou et al., [20] | Recovered COVID-19 patients | Nearly 1 year | Diabetes, hypertension, hyperlipidemia | Hospitalized 17.5–41.5 days | Reduced white matter integrity in nigrostriatal pathways |
| Hafiz et al., [21] | 46 COVID-19 vs. 30 controls | 2 weeks post-discharge | Fatigue | Hospitalized | Higher gray matter volume in limbic regions and basal ganglia; correlation with fatigue |
| Heine et al., [22] | 50 long-COVID vs. 47 controls | Median 7.5 months | Anxiety, depression, sleep problems in long-COVID | 13% Hospitalized | Thalamus and basal ganglia volume loss, surface deformations, altered diffusion; correlated with fatigue |
| Deters et al., [23] | 33 mild COVID-19 patients | <6 months (n = 18) vs. >6 months (n = 15) | Persistent fatigue in some >6 month patients | Not hospitalized | Smaller putamen, pallidum, thalamus volumes in >6 month group, especially fatigued; frontal hypometabolism |
| Luo et al., [17] | 32 post-COVID-19 patients (16 fatigued, 16 non-fatigued) | 6.9 ± 4.8 months (FT), 8.5 ± 5.7 months (NF) | Not specified | Not hospitalized | Decreased globus pallidus activity in both fatigued and non-fatigued groups compared to healthy controls; non-fatigued group showed greater hypometabolism; right hemisphere more affected in both groups |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudroff, T. Decoding Post-Viral Fatigue: The Basal Ganglia’s Complex Role in Long-COVID. Neurol. Int. 2024, 16, 380-393. https://doi.org/10.3390/neurolint16020028
Rudroff T. Decoding Post-Viral Fatigue: The Basal Ganglia’s Complex Role in Long-COVID. Neurology International. 2024; 16(2):380-393. https://doi.org/10.3390/neurolint16020028
Chicago/Turabian StyleRudroff, Thorsten. 2024. "Decoding Post-Viral Fatigue: The Basal Ganglia’s Complex Role in Long-COVID" Neurology International 16, no. 2: 380-393. https://doi.org/10.3390/neurolint16020028
APA StyleRudroff, T. (2024). Decoding Post-Viral Fatigue: The Basal Ganglia’s Complex Role in Long-COVID. Neurology International, 16(2), 380-393. https://doi.org/10.3390/neurolint16020028