
First Molecular Detection and Characterization of Fowl Aviadenovirus Serotype 11 from Broiler Chickens in Chile
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
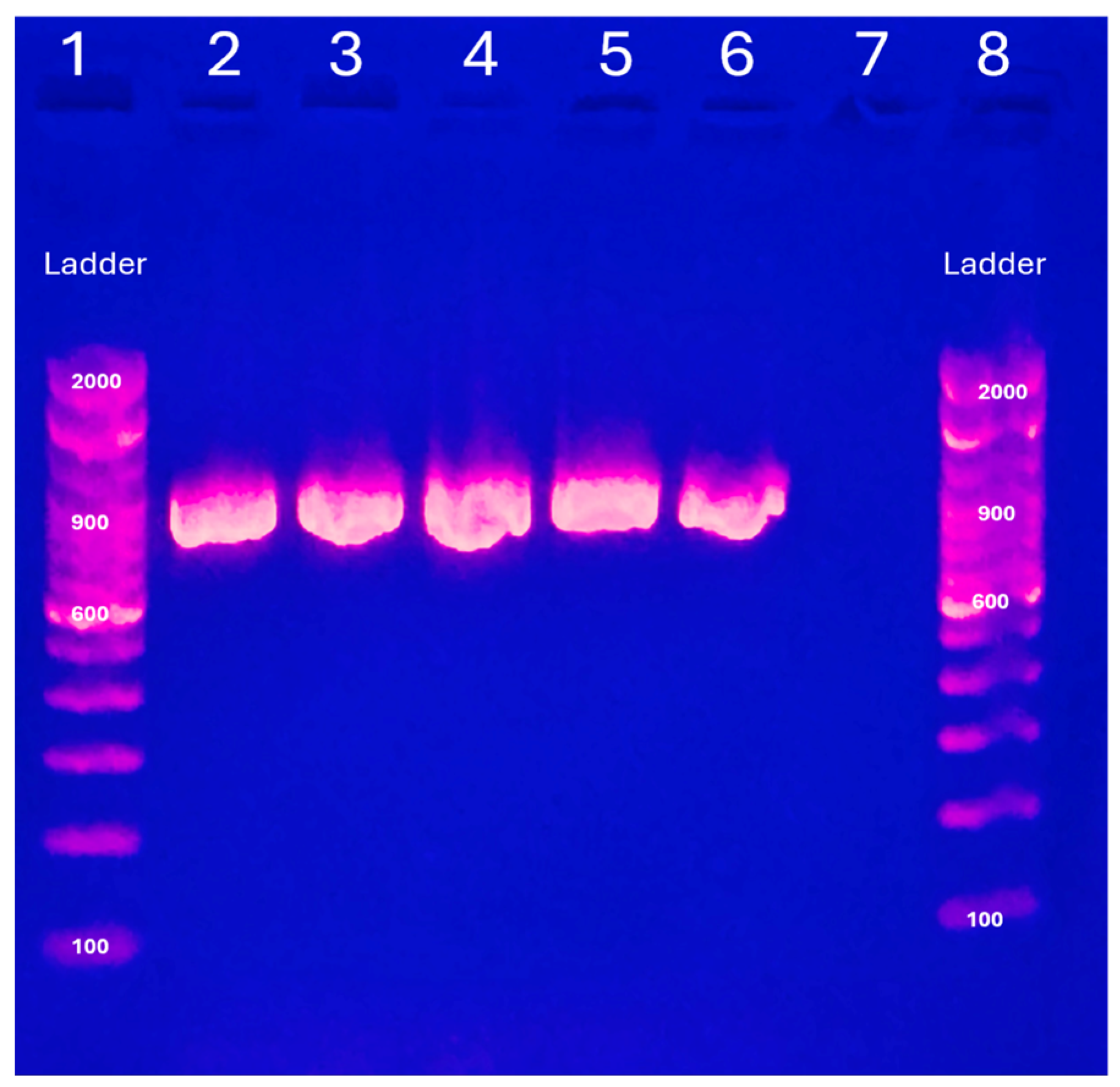
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benkő, M.; Aoki, K.; Arnberg, N.; Davison, A.J.; Echavarría, M.; Hess, M.; Jones, M.S.; Kaján, G.L.; Kajon, A.E.; Mittal, S.K.; et al. ICTV Report Consortium. ICTV Virus Taxonomy Profile: Adenoviridae 2022. J. Gen. Virol. 2022, 103, 001721. [Google Scholar] [CrossRef] [PubMed]
- Hess, M. Detection and differentiation of avian adenoviruses: A review. Avian Pathol. 2000, 29, 195–206. [Google Scholar] [CrossRef]
- Schachner, A.; Marek, A.; Grafl, B.; Hess, M. Detailed molecular analyses of the hexon loop-1 and fibers of fowl aviadenoviruses reveal new insights into the antigenic relationship and confirm that specific genotypes are involved in field outbreaks of inclusion body hepatitis. Vet. Microbiol. 2016, 186, 13–20. [Google Scholar] [CrossRef]
- Marek, A.; Günes, A.; Schulz, E.; Hess, M. Classification of fowl adenoviruses by use of phylogenetic analysis and high-resolution melting-curve analysis of the hexon L1 gene region. J. Virol. Methods 2010, 170, 147–154. [Google Scholar] [CrossRef] [PubMed]
- El-Shall, N.A.; El-Hamid, H.S.A.; Elkady, M.F.; Ellakany, H.F.; Elbestawy, A.R.; Gado, A.R.; Geneedy, A.M.; Hasan, M.E.; Jaremko, M.; Selim, S.; et al. Epidemiology, pathology, prevention, and control strategies of inclusion body hepatitis and hepatitis-hydropericardium syndrome in poultry: A comprehensive review. Front. Vet. Sci. 2022, 9, 963199. [Google Scholar] [CrossRef]
- Chavan, V.G.; Awandkar, S.P.; Kulkarni, M.B.; Chavhan, S.G.; Kulkarni, R.C.; Agnihotri, A.A. Molecular phylodynamics of fowl adenovirus serotype 11 and 8b from inclusion body hepatitis outbreaks. Virus Genes 2023, 59, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, P.; Brown, T.; Villegas, P.; McMurray, B.; Page, R.K.; Rowland, G.N.; Thayer, S.G. Adenovirus-induced inclusion body hepatitis in four-day-old broiler breeders. Avian Dis. 1997, 41, 472–474. [Google Scholar] [CrossRef]
- Helmboldt, C.F.; Frazier, M.N. Avian hepatic inclusion bodies of unknown significance. Avian Dis. 1963, 7, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, J.K.; Eckroade, R.J.; Klopp, S.; Krauss, W.C. Characterization of several viruses isolated from chickens with inclusion body hepatitis and aplastic anemia. Avian Dis. 1974, 18, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Fadly, A.M.; Winterfield, R.W.; Olander, H.J. Role of the bursa of Fabricius in the pathogenicity of inclusion body hepatitis and infectious bursal disease viruses. Avian Dis. 1976, 20, 467–477. [Google Scholar] [CrossRef]
- Gomis, S.; Goodhope, A.R.; Ojkic, A.D.; Willson, P. Inclusion body hepatitis as a primary disease in broilers in Saskatchewan, Canada. Avian Dis. 2006, 50, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Abghour, S.; Zro, K.; Mouahid, M.; Tahiri, F.; Tarta, M.; Berrada, J.; Kichou, F. Isolation and characterization of fowl aviadenovirus serotype 11 from chickens with inclusion body hepatitis in Morocco. PLoS ONE 2019, 14, e0227004. [Google Scholar] [CrossRef] [PubMed]
- Mase, M.; Hiramatsu, K.; Nishijima, N.; Iguchi, H.; Honda, S.; Hanyu, S.; Iseki, H.; Watanabe, S. Fowl Adenoviruses Type 8b Isolated from Chickens with Inclusion Body Hepatitis in Japan. Avian Dis. 2020, 64, 330–334. [Google Scholar] [CrossRef]
- Bertran, K.; Blanco, A.; Antilles, N.; Nofrarías, M.; Valle, R.M.; Cobos, À.; Ramis, A.; Biarnés, M.; Majó, N. A 10-Year Retrospective Study of Inclusion Body Hepatitis in Meat-Type Chickens in Spain (2011–2021). Viruses 2021, 13, 2170. [Google Scholar] [CrossRef]
- Chitradevi, S.; Sukumar, K.; Suresh, P.; Balasubramaniam, G.A.; Kannan, D. Molecular typing and pathogenicity assessment of fowl adenovirus associated with inclusion body hepatitis in chicken from India. Trop. Anim. Health Prod. 2021, 53, 412. [Google Scholar] [CrossRef] [PubMed]
- Niczyporuk, J.S.; Kozdrun, W.; Czekaj, H.; Piekarska, K.; Stys-Fijol, N. Characterisation of adenovirus strains represented species B and E isolated from broiler chicken flocks in eastern Poland. Heliyon 2021, 7, e06225. [Google Scholar] [CrossRef] [PubMed]
- Tsiouris, V.; Mantzios, T.; Kiskinis, K.; Guérin, J.L.; Croville, G.; Brellou, G.D.; Apostolopoulou, E.P.; Petridou, E.J.; Georgopoulou, I. First Detection and Identification of FAdV-8b as the Causative Agent of an Outbreak of Inclusion Body Hepatitis in a Commercial Broiler Farm in Greece. Vet. Sci. 2022, 9, 160. [Google Scholar] [CrossRef]
- Alzuheir, I.M.; Jalboush, N.H.; Fayyad, A.F.; Daibes, R.A. Phylogenetic analysis and pathological characterization of fowl adenovirus isolated during inclusion body hepatitis outbreak in Tubas, Palestine. Vet. Res. Forum 2021, 12, 511–514. [Google Scholar] [CrossRef]
- Islam, M.N.; Rahman, M.M.; Rahman, M.K.; Alam, J. First Evidence of Fowl Adenovirus Induced Inclusion Body Hepatitis in Chicken in Bangladesh. Can. J. Infect. Dis. Med. Microbiol. 2023, 2023, 7253433. [Google Scholar] [CrossRef]
- Joubert, H.W.; Aitchison, H.; Maartens, L.H.; Venter, E.H. Molecular differentiation and pathogenicity of Aviadenoviruses isolated during an outbreak of inclusion body hepatitis in South Africa. J. S. Afr. Vet. Assoc. 2014, 85, 1058. [Google Scholar] [CrossRef]
- Mendelson, C.; Nothelfer, H.B.; Monreal, G. Identification and characterization of an avian adenovirus isolated from a ‘spiking mortality syndrome’ field outbreak in broilers on the Delmarva Peninsula, USA. Avian Pathol. 1995, 24, 693–706. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Lara, R.; Calderón, K.; Chumbe, A.; Montesinos, R.; Montalván, Á.; González, A.E.; Icochea, E.; Fernández-Díaz, M. Complete Genome Sequence of Fowl Aviadenovirus Serotype 8b Isolated in South America. Genome Announc. 2016, 4, e01174-16. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, D.; Nuñez, L.F.N.; Santander Parra, S.H.; Astolfi-Ferreira, C.S.; Piantino Ferreira, A.J. Molecular characterization of fowl adenovirus group I in commercial broiler chickens in Brazil. Virus Dis. 2018, 29, 83–88. [Google Scholar] [CrossRef]
- Santander-Parra, S.H.; Caza, M.; Nuñez, L. Detection, Quantification and Molecular Characterization of Fowl Adenoviruses Circulating in Ecuadorian Chicken Flocks during 2019–2021. Vet. Sci. 2023, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Meulemans, G.; Boschmans, M.; Berg, T.P.; Decaesstecker, M. Polymerase chain reaction combined with restriction enzyme analysis for detection and differentiation of fowl adenoviruses. Avian Pathol. 2001, 30, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Raue, R.; Hess, M. Hexon based PCRs combined with restriction enzyme analysis for rapid detection and differentiation of fowl adenoviruses and egg drop syndrome virus. J. Virol. Methods 1998, 73, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; Prusas, C.; Raue, R.; Cerda, L.; Geisse, C.; González, C.; Hess, M. Characterization of fowl adenoviruses from outbreaks of inclusion body hepatitis/hydropericardium syndrome in Chile. Avian Dis. 1999, 43, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, K.; Suryanarayana, V.V.; Raghavan, R. Detection of fowl adenovirus associated with hydropericardium hepatitis syndrome by a polymerase chain reaction. Vet. Res. Commun. 2022, 26, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Hayashima, A.; Tanaka, T.; Ninomiya, N.; Tonogawa, T.; Nakazato, S.; Mase, M. Virucidal effect of commercial disinfectants on fowl adenovirus serotype 1 strains causing chicken gizzard erosion in Japan. J. Appl. Poult. Res. 2020, 29, 383–390. [Google Scholar] [CrossRef]
- Steer-Cope, P.A.; Sandy, J.R.; O’Rourke, D.; Scott, P.C.; Browning, G.F.; Noormohammadi, A.H. Vaccination with FAdV-8a induces protection against inclusion body hepatitis caused by homologous and heterologous strains. Avian Pathol. 2019, 48, 396–405. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′ to 3′) | Position 1 | PCR Product (bp) | Reference |
|---|---|---|---|---|
| Hexon A | CAARTTCAGRCAGACGGT | 144–161 | 897 | [25] |
| Hexon B | TAGTGATGMCGSGACATCAT | 1041–1021 | ||
| H1 | TGGGACATGGGGGCGACCTA | 296–314 | 1219 | [26] |
| H2 | AAGGGATTGACGTTGTCCA | 1514–1496 |
| CL | FAdV-A | FAdV-D | FAdV-C | FAdV-E | FAdV-B | |
|---|---|---|---|---|---|---|
| CL | 0.612 | 0.071 | 0.687 | 0.416 | 0.563 | |
| FAdV-A | 0.392 | 0.611 | 0.531 | 0.611 | 0.529 | |
| FAdV-D | 0.076 | 0.384 | 0.689 | 0.396 | 0.532 | |
| FAdV-C | 0.392 | 0.328 | 0.396 | 0.617 | 0.647 | |
| FAdV-E | 0.294 | 0.354 | 0.286 | 0.356 | 0.446 | |
| FAdV-B | 0.340 | 0.348 | 0.329 | 0.370 | 0.299 |
| N° | FAdV Isolate/GenBank Accession No | FAdV Species | FAdV Serotype | Positions of Nucleotide Sequence Hexon Gene | |||
|---|---|---|---|---|---|---|---|
| 223 | 273 | 450 | 462 | ||||
| 1 | KX247375 | D | 11 | A | A | T | A |
| 2 | 15927/CHILE/2023 | D | 11 | G | G | C | G |
| 3 | 15929/CHILE/2023 | D | 11 | G | G | C | G |
| 4 | 15950/CHILE/2023 | D | 11 | G | G | C | G |
| 5 | 15996/CHILE/2023 | D | 11 | G | G | C | G |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cádiz, L.; Guzmán, M.; Navarrete, F.; Torres, P.; Hidalgo, H. First Molecular Detection and Characterization of Fowl Aviadenovirus Serotype 11 from Broiler Chickens in Chile. Microbiol. Res. 2024, 15, 626-633. https://doi.org/10.3390/microbiolres15020041
Cádiz L, Guzmán M, Navarrete F, Torres P, Hidalgo H. First Molecular Detection and Characterization of Fowl Aviadenovirus Serotype 11 from Broiler Chickens in Chile. Microbiology Research. 2024; 15(2):626-633. https://doi.org/10.3390/microbiolres15020041
Chicago/Turabian StyleCádiz, Leandro, Miguel Guzmán, Fernando Navarrete, Paulina Torres, and Hector Hidalgo. 2024. "First Molecular Detection and Characterization of Fowl Aviadenovirus Serotype 11 from Broiler Chickens in Chile" Microbiology Research 15, no. 2: 626-633. https://doi.org/10.3390/microbiolres15020041