Below-Ground Interspecific Competition of Apple (Malus pumila M.)–Soybean (Glycine max L. Merr.) Intercropping Systems Based on Niche Overlap on the Loess Plateau of China

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
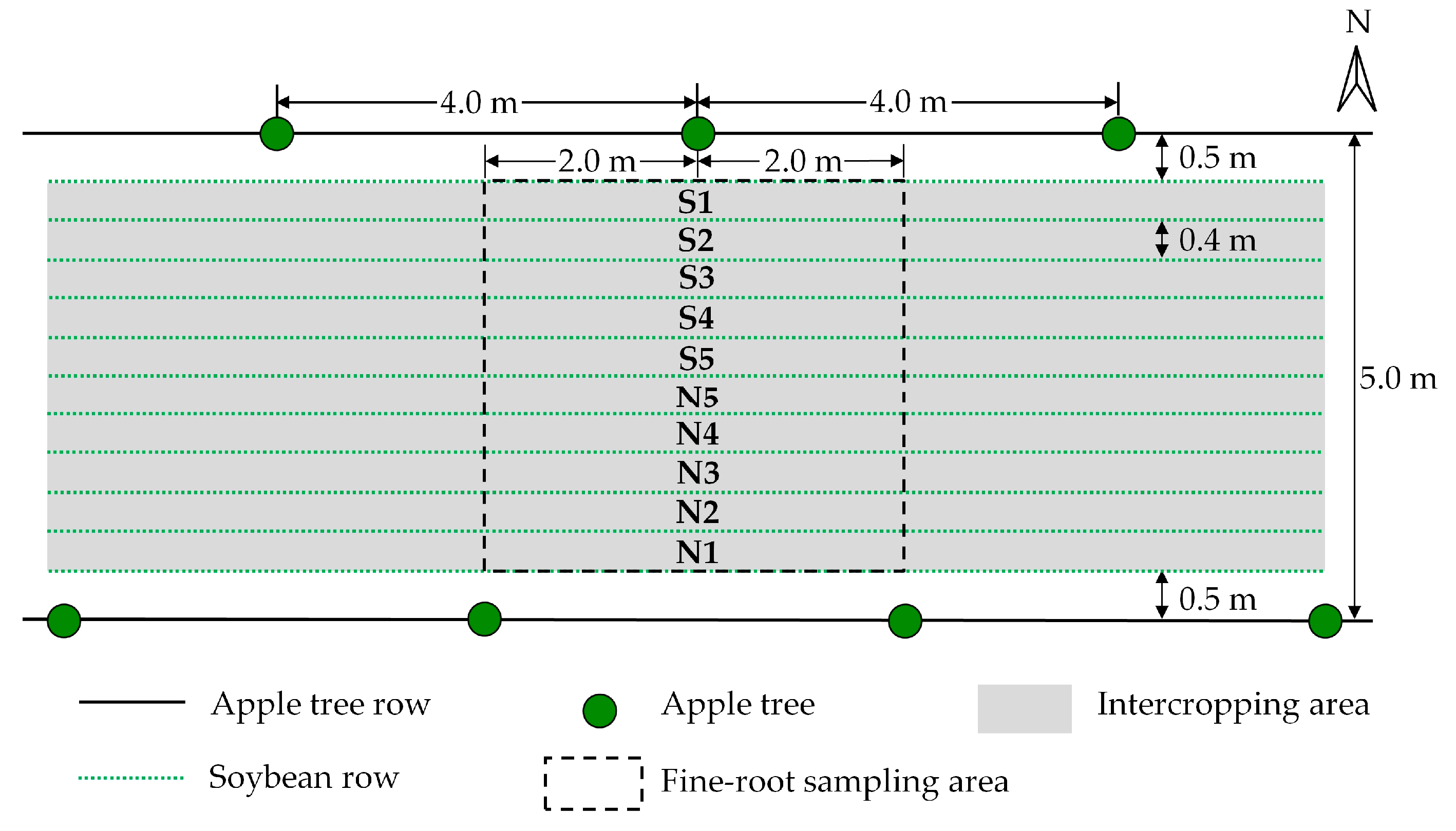
2.2. Materials and Experimental Design
2.3. Fine-Root Biomass Density
2.4. Fine-Root Niche Differentiation
2.5. Below-Ground Interspecific Competition Intensity Index
2.6. Statistical Analyses
3. Results
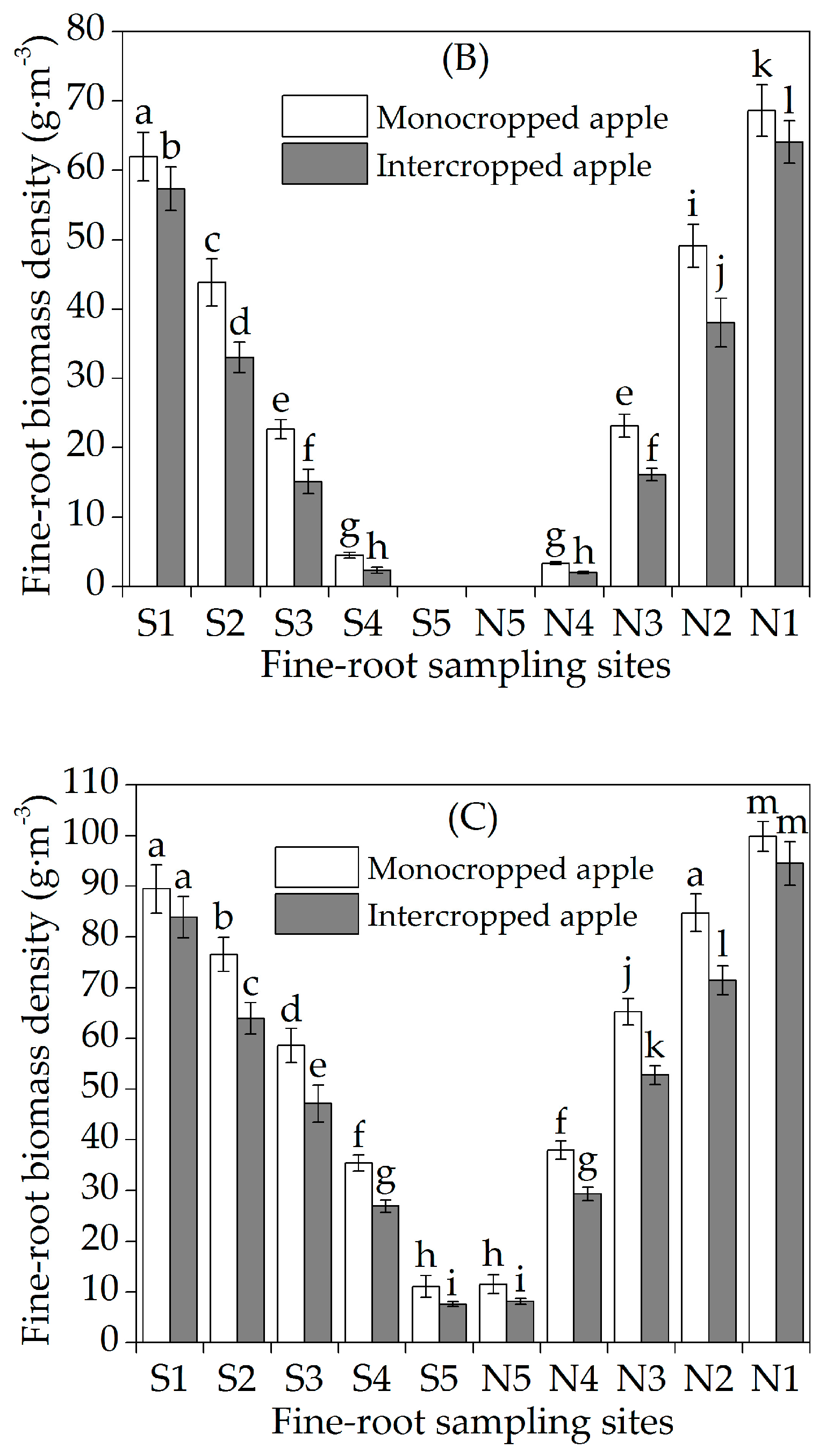
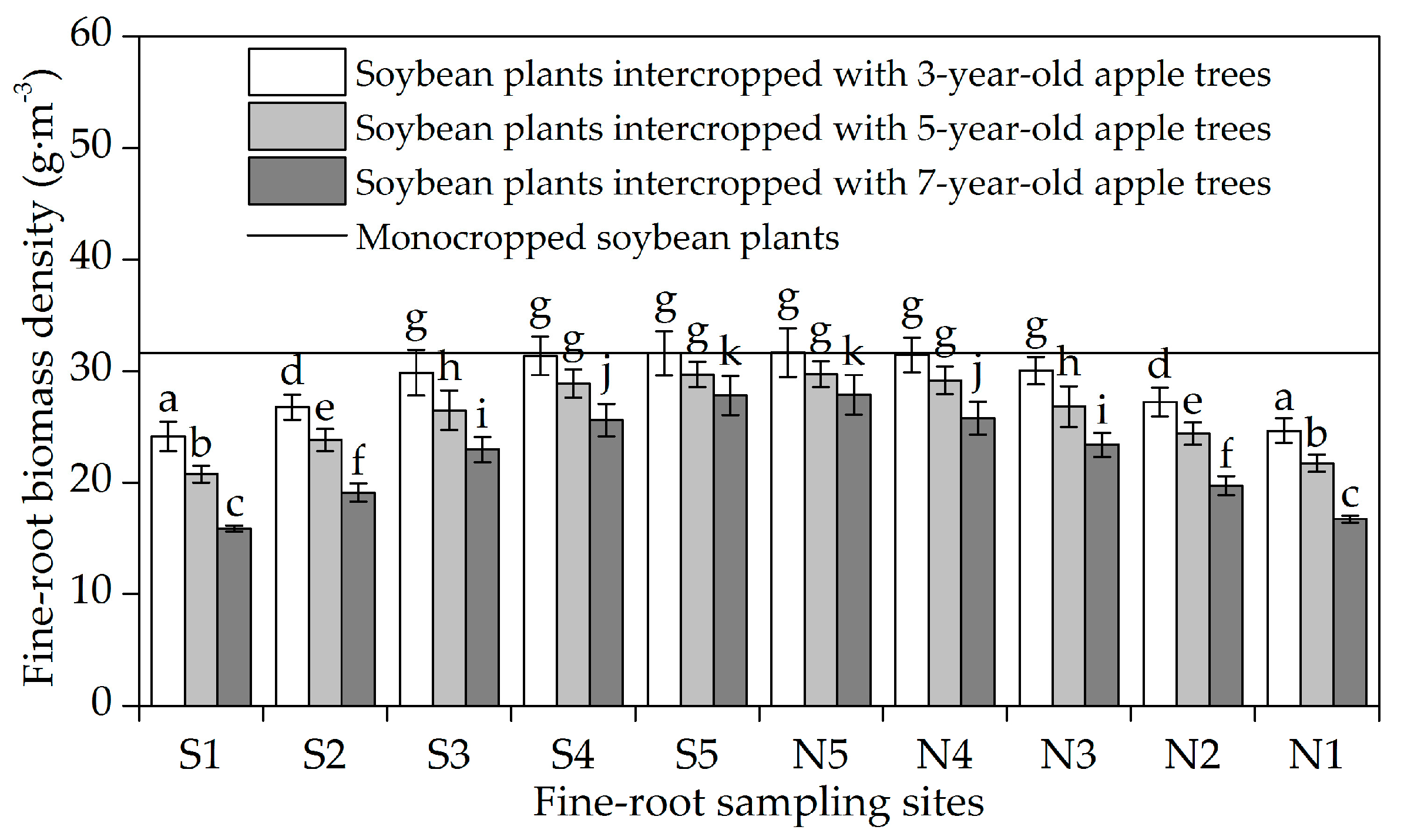
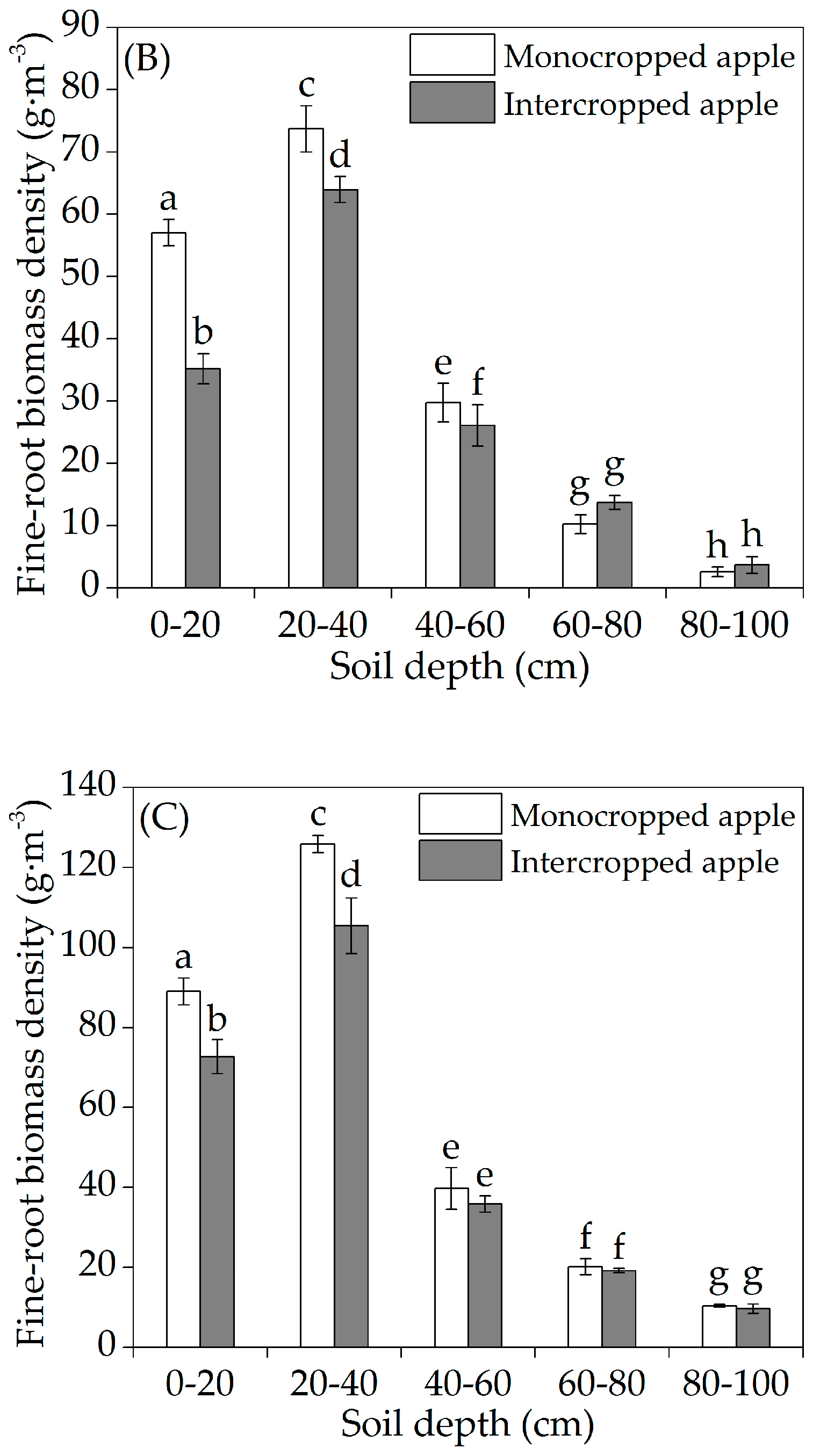
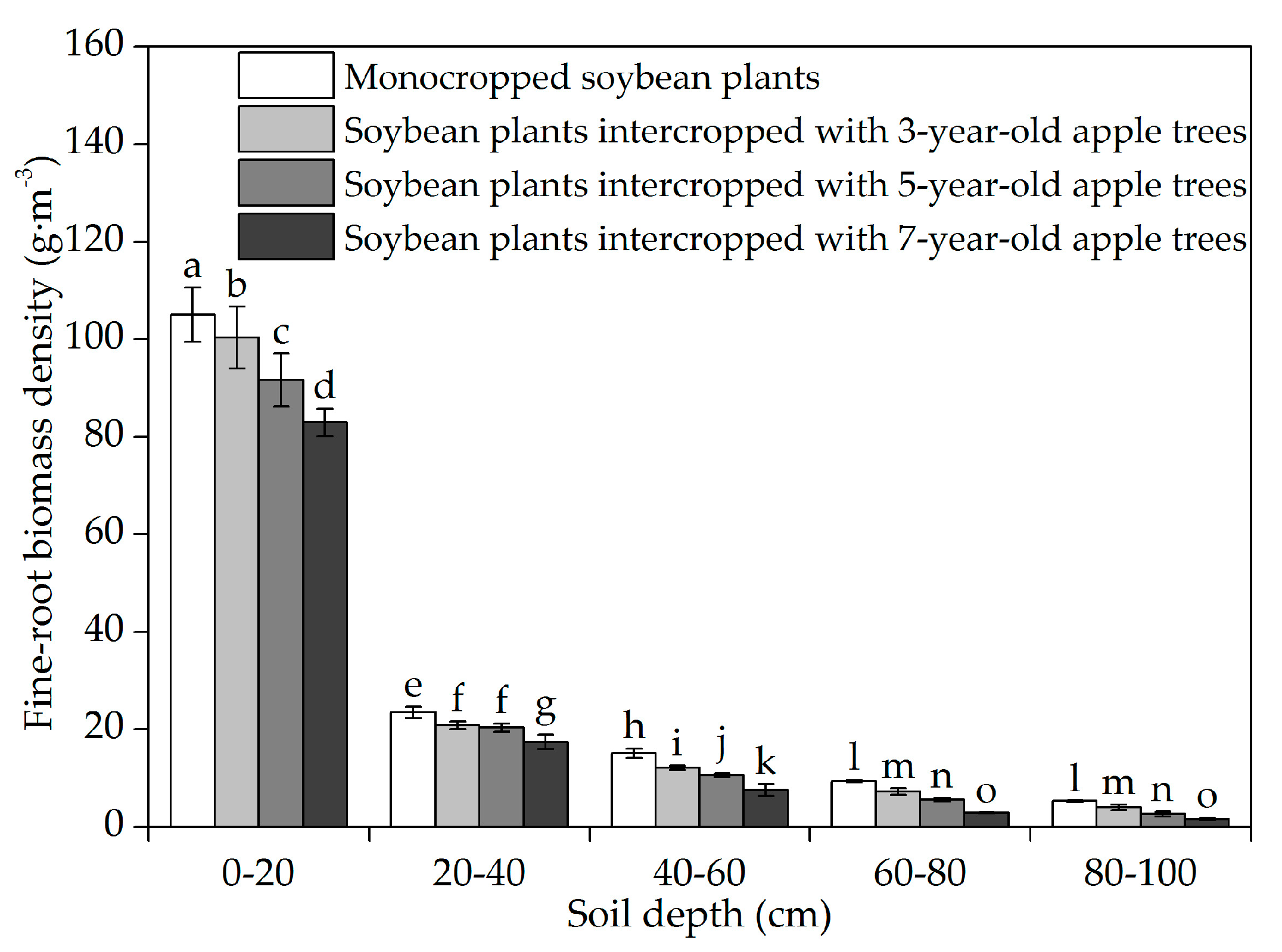
3.1. Horizontal Fine-Root Distribution
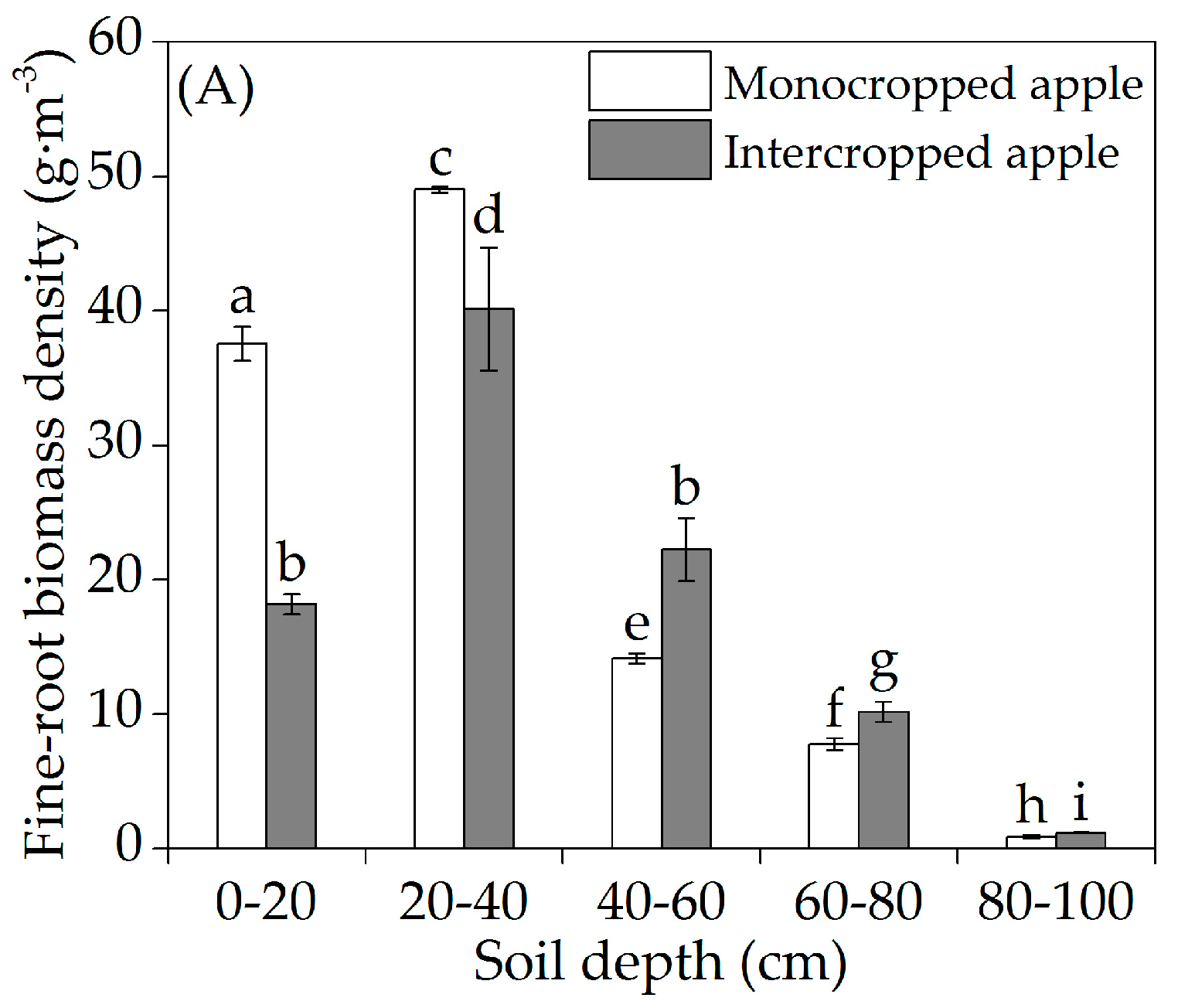
3.2. Vertical Fine-Root Distribution
3.3. Fine-Root Niche Differentiation
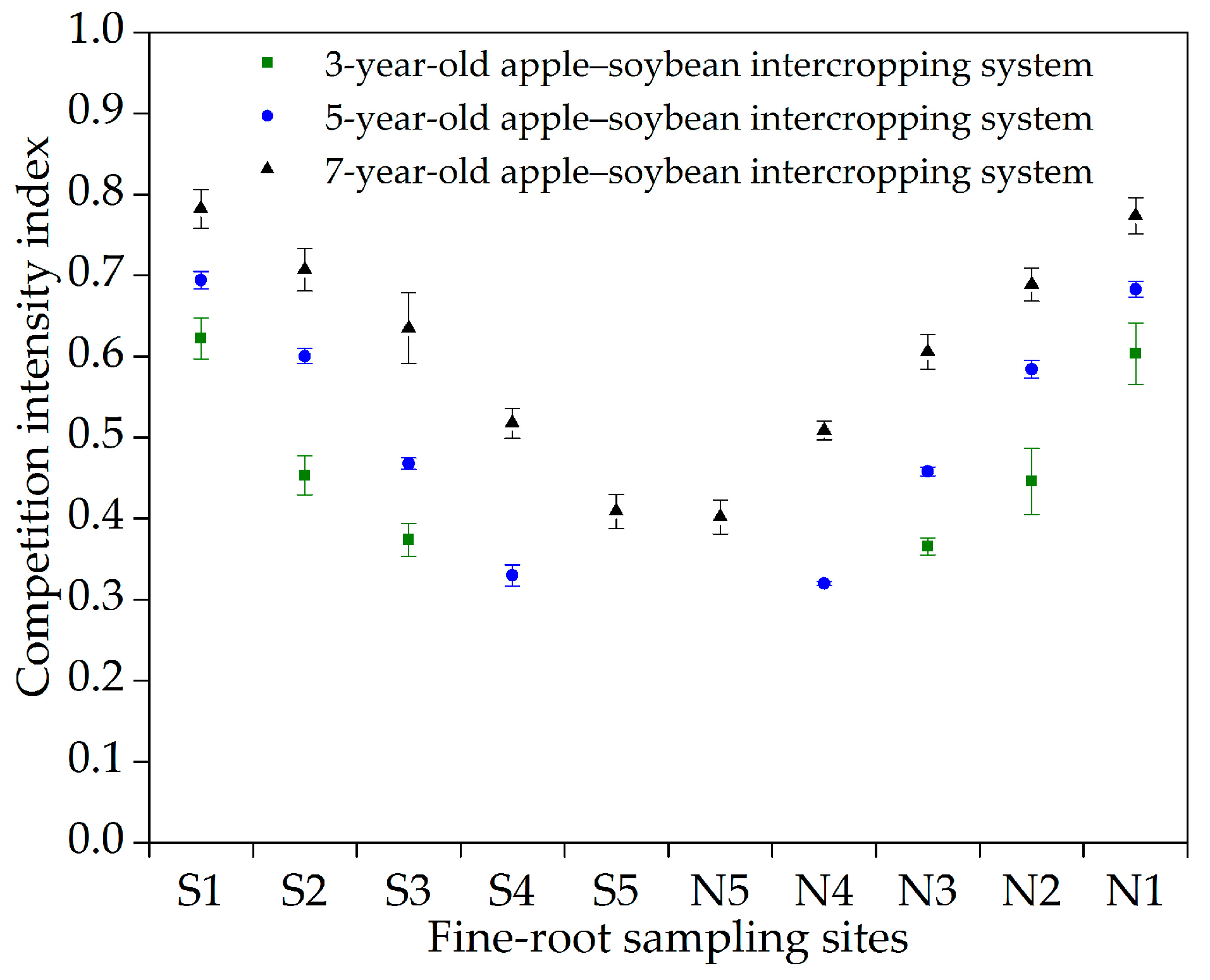
3.4. Below-Ground Interspecific Competition Intensity
4. Discussion
4.1. Horizontal Fine-Root Distribution
4.2. Vertical Fine-Root Distribution
4.3. Fine-Root Niche Differentiation
4.4. Below-Ground Interspecific Competition Intensity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- George, S.J.; Kumar, B.M.; Wahid, P.A.; Kamalam, N.V. Root competition for phosphorus between the tree and herbaceous components of silvopastoral systems in Kerala, India. Plant Soil 1996, 179, 189–196. [Google Scholar] [CrossRef]
- Cannell, M.G.R.; Van Noordwijk, M.; Ong, C.K. The central agroforestry hypothesis: The tree must acquire resources that the crop would not otherwise acquire. Agofor. Syst. 1996, 34, 27–31. [Google Scholar] [CrossRef]
- Zamora, D.S.; Jose, S.; Nair, P.K.R. Morphological plasticity of cotton roots in response to interspecific competition with pecan in an alley cropping system in the southern United States. Agrofor. Syst. 2007, 69, 107–116. [Google Scholar] [CrossRef]
- Ong, C.K.; Corlett, J.E.; Singh, R.P. Above and below ground interaction in agroforestry systems. For. Ecol. Manag. 1991, 45, 45–57. [Google Scholar] [CrossRef]
- Anderson, L.S.; Sinclair, F.L. Ecological interactions in agroforestry systems. Agrofor. Abstr. 1993, 6, 57–91. [Google Scholar]
- Huxley, P.A.; Pinney, A.; Akunda, E.; Muraya, P. A tree/crop interface orientation experiment with a Grevillea robusta, hedgerow and maize. Agofor. Syst. 1994, 26, 23–45. [Google Scholar] [CrossRef]
- Gao, L.; Xu, H.; Bi, H.; Xi, W.; Bao, B.; Wang, X.; Bi, C.; Chang, Y. Intercropping competition between apple trees and crops in agroforestry systems on the Loess Plateau of China. PLoS ONE 2013, 8, e70739. [Google Scholar] [CrossRef] [PubMed]
- Mutua, J.N. Tree Root Architecture and Competition for Water in an Agroforestry System in Semi Arid Kenya. Ph.D. Thesis, University of Nairobi, Nairobi, Kenya, September 1998. [Google Scholar]
- Kumar, S.S.; Kumar, B.M.; Wahid, P.A.; Kamalam, N.V.; Fisher, R.F. Root competition for phosphorus between coconut, multipurpose trees and kacholam (Kaempferia galanga L.) in Kerala, India. Agrofor. Syst. 1999, 46, 131–146. [Google Scholar] [CrossRef]
- Smith, D.M.; Jackson, N.A.; Roberts, J.M.; Ong, C.K. Root distributions in a Grevillea robusta-maize agroforestry system in semi-arid Kenya. Plant Soil 1999, 211, 191–205. [Google Scholar] [CrossRef]
- Livesley, S.J.; Gregory, P.J.; Buresh, R.J. Competition in tree row agroforestry systems. 1. Distribution and dynamics of fine roots length and biomass. Plant Soil 2000, 227, 149–161. [Google Scholar] [CrossRef]
- Xu, H.; Bi, H.; Xi, W.; Powell, R.L.; Gao, L.; Yun, L. Root distribution variation of crops under walnut-based intercropping systems in the Loess Plateau of China. Pak. J. Agric. Sci. 2014, 51, 773–778. [Google Scholar]
- Zhang, W.; Ahanbieke, P.; Wang, B.J.; Xu, W.L.; Li, L.H.; Christie, P.; Li, L. Root distribution and interactions in jujube tree/wheat agroforestry system. Agrofor. Syst. 2013, 87, 929–939. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, B.J.; Gan, Y.W.; Duan, Z.P.; Hao, X.D.; Xu, W.L.; Lv, X.; Li, L.H. Competitive interaction in a jujube tree/wheat agroforestry system in northwest China’s Xinjiang Province. Agrofor. Syst. 2016, 91, 881–893. [Google Scholar] [CrossRef]
- Mulia, R.; Dupraz, C. Unusual fine root distributions of two deciduous tree species in southern France: What consequences for modelling of tree root dynamics? Plant Soil 2006, 281, 71–85. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Cahill, J.F.; McNickle, G.G.; Haag, J.J.; Lamb, E.G.; Nyanumba, S.M.; Clair, C.C.S. Plants integrate information about nutrients and neighbors. Science 2010, 328, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Schroth, G. Tree root characteristics as criteria for species selection and systems design in agroforestry. Agrofor. Syst. 1995, 30, 125–143. [Google Scholar] [CrossRef]
- Caldwell, M.M.; Richards, J.H. Competing root systems: Morphology and models of absorption. In On the Economy of Plant Form and Function; Givnish, T.J., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 251–273. [Google Scholar]
- McIntyre, B.D.; Riha, S.J.; Ong, C.K. Light interception and evapotranspiration in hedgerow agroforestry systems. Agric. For. Meteorol. 1996, 81, 31–40. [Google Scholar] [CrossRef]
- May, R.; McLean, A.R. Interspecific competition and multispecies coexistence. In Theoretical Ecology: Principles and Applications, 3rd ed.; McLean, A.R., Ed.; Oxford University Press: New York, NY, USA, 2007; pp. 84–97. [Google Scholar]
- Zhu, Q.K.; Zhu, J.Z. Sustainable Management Technology for Conversion of Cropland to Forest in Loess Area; Chinese Forestry Press: Beijing, China, 2003; pp. 179–186. [Google Scholar]
- Xu, H.; Bi, H.; Gao, L.; Yun, L.; Chang, Y.; Xi, W.; Liao, W.; Bao, B. Distribution and morphological variation of fine root in a walnut-soybean intercropping system in the Loess plateau of China. Int. J. Agric. Biol. 2013, 15, 998–1002. [Google Scholar]
- Pianka, E.R. The structure of lizard communities. Annu. Rev. Ecol. Syst. 1973, 4, 53–74. [Google Scholar] [CrossRef]
- Yang, X.W.; Ma, J.S. A review on some terms related to niche and their measurements. Chin. J. Ecol. 1992, 11, 44–49. [Google Scholar]
- Liu, W.; Cao, W. Niche characteristics of main plant species in spruce-fir forests in Changbai Mountains. Chin. J. Ecol. 2011, 30, 1766–1774. [Google Scholar]
- Schroth, G. A review of belowground interactions in agroforestry, focussing on mechanisms and management options. Agrofor. Syst. 1999, 43, 5–34. [Google Scholar] [CrossRef]
- Eastham, J.; Rose, C.W. Tree/pasture interactions at a range of tree densities in an agroforestry experiment. 1. Rooting patterns. Aust. J. Agric. Res. 1990, 41, 683–695. [Google Scholar] [CrossRef]
- Jackson, R.B.; Canadell, J.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. A global analysis of root distribution for terrestrial biomass. Oecologia 1996, 108, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Yun, L.; Bi, H.; Ren, Y.; Wu, J.; Chen, P.P.; Ma, W.J. Research on soil moisture relations among types of agroforestry system in the Loess Region. Bull. Soil Water Conserv. 2008, 28, 110–114. [Google Scholar]
- Kasperbauer, M.J.; Busscher, W.J. Genotypic differences in cotton root penetration of a compacted sub-soil layer. Crop Sci. 1991, 31, 1376–1378. [Google Scholar] [CrossRef]
- Van Noordwijk, M.; Purnomosidhi, P. Root architecture in relation to tree-soil-crop interactions and shoot pruning in agroforestry. Agrofor. Syst. 1995, 30, 161–173. [Google Scholar] [CrossRef]
- Bolte, A.; Villanueva, I. Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur. J. For. Res. 2006, 125, 15–26. [Google Scholar] [CrossRef]
- Ong, C.K.; Black, C.R.; Marshall, F.M.; Corlett, J.E. Principles of resource capture and utilization of light and water. In Tree-Crop Interactions: A Physiological Approach; Ong, C.K., Huxley, P., Eds.; CAB International: Wallingford, UK, 1996; pp. 73–158. [Google Scholar]
- Wanvestraut, R.H.; Jose, S.; Nair, P.K.R.; Brecke, B.J. Competition for water in a pecan (Carya illinoensis K. Koch)-cotton (Gossypium hirsutum L.) alley cropping system in the southern United States. Agrofor. Syst. 2004, 60, 167–179. [Google Scholar] [CrossRef]
- Harpole, W. Neutral theory of species diversity. Nat. Educ. Knowl. 2012, 1, 31–36. [Google Scholar]
- Jose, S.; Gillespie, A.R.; Pallardy, S.G. Interspecific interactions in temperate agroforestry. Agrofor. Syst. 2004, 61, 237–255. [Google Scholar] [CrossRef]
- Hanski, L. Some comments on the measurement of niche metrics. Ecology 1978, 59, 168–174. [Google Scholar] [CrossRef]
- Abrams, P. Some comments on measuring niche overlap. Ecology 1980, 61, 44–49. [Google Scholar] [CrossRef]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Y.; Bi, H.; Xu, H.; Duan, H.; Peng, R.; Wang, J. Below-Ground Interspecific Competition of Apple (Malus pumila M.)–Soybean (Glycine max L. Merr.) Intercropping Systems Based on Niche Overlap on the Loess Plateau of China. Sustainability 2018, 10, 3022. https://doi.org/10.3390/su10093022
Sun Y, Bi H, Xu H, Duan H, Peng R, Wang J. Below-Ground Interspecific Competition of Apple (Malus pumila M.)–Soybean (Glycine max L. Merr.) Intercropping Systems Based on Niche Overlap on the Loess Plateau of China. Sustainability. 2018; 10(9):3022. https://doi.org/10.3390/su10093022
Chicago/Turabian StyleSun, Yubo, Huaxing Bi, Huasen Xu, Hangqi Duan, Ruidong Peng, and Jingjing Wang. 2018. "Below-Ground Interspecific Competition of Apple (Malus pumila M.)–Soybean (Glycine max L. Merr.) Intercropping Systems Based on Niche Overlap on the Loess Plateau of China" Sustainability 10, no. 9: 3022. https://doi.org/10.3390/su10093022