Regeneration Ecology of the Rare Plant Species Verbascum dingleri: Implications for Species Conservation

 , , , and
, , , and
Abstract
:1. Introduction
- (i)
- Determining the environmental drivers forcing species distribution;
- (ii)
- Studying species fruit and seed diversity;
- (iii)
- Investigating seed germination behavior in laboratory (controlled environment) and ambient conditions (nursery), including the effect of fruit characteristics on seed germination;
- (iv)
- Developing a scientific approach for the species and its habitat conservation by producing seedlings from seeds in a nursery.
2. Materials and Methods
2.1. Species Description
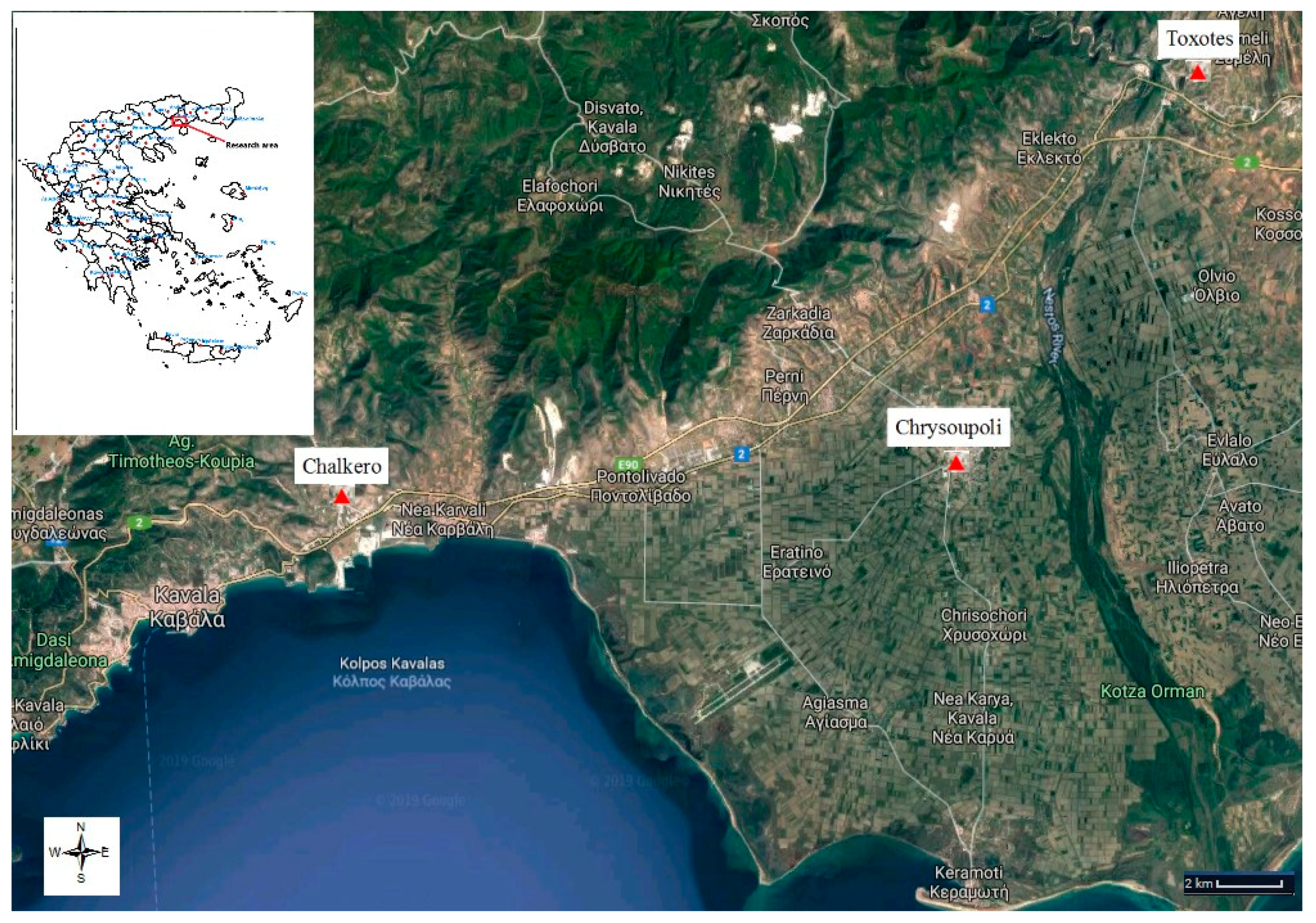
2.2. Species Distribution
2.3. Estimation of Environmental Drivers Limiting the Species Distribution
2.4. Community Characteristics and Estimation of Possible Biotic Interactions
2.5. Fruit and Seed Collection and Laboratory Analysis
2.6. Seed Germination under Controlled and Ambient Environmental Conditions
2.7. Seed Germination and Seedling Emergence at Nursery Conditions
2.8. Statistical Analysis
3. Results
3.1. Species Geographical Distribution
3.2. Community Characteristics
3.3. Fruit and Seed Diversity
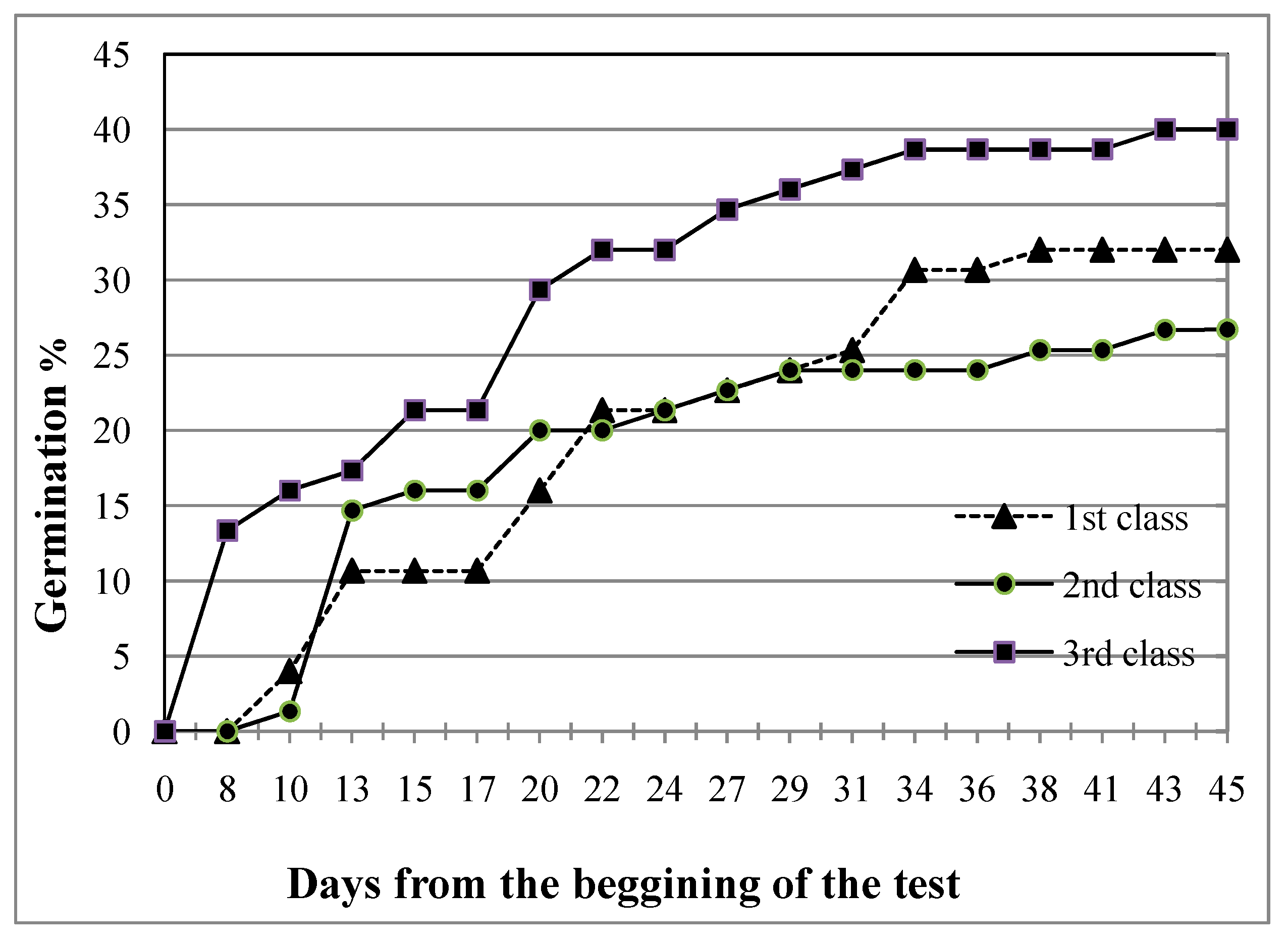
3.4. Seed Germination Behavior
4. Discussion
4.1. Distribution and Habitat of V. dingleri
4.2. Species Regeneration Ecology
4.3. Species Conservation
Author Contributions
Conflicts of Interest
References
- Primack, Ρ.Β. Essentials of Conservation Biology, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2006; p. 580. [Google Scholar]
- Jusaitis, M.; Polomka, L.; Sorensen, B. Habitat specificity, seed germination and experimental translocation of the endangered herb Brachycome muelleri (Asteraceae). Biol. Conserv. 2004, 116, 251–266. [Google Scholar] [CrossRef]
- Shen, S.K.; Wang, Y.H.; Wang, B.Y.; Ma, H.Y.; Shen, G.Z.; Ham, Z.W. Distribution, stand characteristics and habitat of a critically endangered plant Euryodendron excelsum HT Chang (Theaceae): Implications for conservation. Plant Species Biol. 2009, 24, 133–138. [Google Scholar] [CrossRef]
- Shen, S.K.; Wang, Y.H.; Ma, H.Y. Seed germination requirements and responses to desiccation and storage of Apterosperma oblata (Theaceae), an endangered tree from south-eastern China: Implications for restoration. Plant Species Biol. 2010, 25, 158–163. [Google Scholar] [CrossRef]
- Navarro, L.; Guitián, J. Seed germination and seedling survival of two threatened endemic species of the northwest Iberian Peninsula. Biol. Conserv. 2003, 109, 313–320. [Google Scholar] [CrossRef]
- Cerabolini, B.; De Andreis, R.; Ceriani, R.M.; Pierce, S.; Raimondi, B. Seed germination and conservation of endangered species from the Italian Alps: Physoplexis comosa and Primula glaucescens. Biol.Conserv. 2004, 117, 351–356. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; 260p. [Google Scholar]
- Butola, J.S.; Badola, H.K. Effect of pre-sowing treatment on seed germination and seedling vigour in Angelica glauca, a threatened medicinal herb. Curr. Sci. 2004, 87, 796–799. [Google Scholar]
- Herranz, J.M.; Ferrandis, P.; Martínez-Duro, E. Seed germination ecology of the threatened endemic Iberian Delphinium fissumsub sp. sordidum (Ranunculaceae). Plant Ecol. 2010, 211, 89–106. [Google Scholar] [CrossRef]
- Mattfeld, J. Eine neue Koenigskerze aus Thracien, Verbascum dingleri. Bull. Soc. Bot. Bulg. 1926, 1, 101–103. [Google Scholar]
- Global Biodiversity Information Facility (GBIF). Available online: https://www.gbif.org/ (accessed on 31 March 2019).
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tsanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Englera; Botanic Garden and Botanical Museum: Berlin, Germany; Hellenic Botanical Society: Athens, Greece, 2013. [Google Scholar]
- Flora of Greece Web. 2019. Available online: http://portal.cybertaxonomy.org/flora-greece/intro (accessed on 31 March 2019).
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis. Mineralogical, Organic and Inorganic Methods; Springer: Berlin/Heidelberg, Germany, 2007; p. 993. [Google Scholar]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils; Pearson Education Limitted: Harlow, UK, 2017; p. 1083. [Google Scholar]
- Barkman, J.J.; Doing, H.; Segal, S. Kritische Bemerkungen und Vorschläge zur quantitativen Vegetationsanalyse. Acta Bot. Neerl. 1964, 13, 394–419. [Google Scholar] [CrossRef]
- Chytrý, M.; Otýpková, Z. Plot sizes used for phytosociological sampling of European vegetation. J. Veg. Sci. 2003, 14, 563–570. [Google Scholar] [CrossRef]
- Graniszewska, M.; Muranyi, R.; Prokopiv, A. Methods of germination and cryogenic storage of rare species seeds from the Ukrainian Carpathians. Visnyk L’viv Univ. Ser. Biol. 2004, 36, 153–159. [Google Scholar]
- ISTA (International Seed Testing Association). International rules for seed testing. Seed Sci. Technol. 1999, 13, 229–513. [Google Scholar]
- Senel, E.; Ozdener, Y.; Incedere, D. Effect of temperature, light, seed weight and GA3 on the germination of Verbascum bithynicum, Verbascum wiedemannianum and Salvia dicroantha. Pak. J. Biol. Sci. 2007, 10, 1118–1121. [Google Scholar] [PubMed]
- Seipel, T.; Alexander, J.M.; Daehler, C.C.; Rew, L.J.; Edwards, P.J.; Dar, P.A.; McDougall, K.; Naylor, B.; Parks, C.; Pollnac, F.W.; et al. Performance of the herb Verbascum thapsus along environmental gradients in its native and non-native ranges. J. Biogeogr. 2015, 42, 132–143. [Google Scholar] [CrossRef]
- Ganatsas, P.; Tsakaldimi, M.; Zachariadis, G. Effect of air traffic pollution on seed quality characteristics of Pinus brutia. Environ. Exp. Bot. 2011, 74, 157–161. [Google Scholar] [CrossRef]
- Ganatsas, P.; Tsakaldimi, M.; Thanos, C. Seed and cone diversity and seed germination of Pinus pinea in Strofylia Site of the Natura 2000 Network. Biodivers. Conserv. 2008, 17, 2427–2439. [Google Scholar] [CrossRef]
- Martínez-Díaz, E.; Martínez-Sánchez, J.J.; Conesa, E.; Franco, J.A.; Vicente, M.J. Germination and morpho-phenological traits of Allium melananthum, a rare species from south-eastern Spain. Flora 2018, 249, 16–23. [Google Scholar] [CrossRef]
- Hellenic National Meteorological Service. Climatic Data of the Greek Network, Period 1930–1975; Hellenic National Meteorological Service: Athens, Greece, 1978. (In Greek) [Google Scholar]
- Casavecchia, S.; Biscotti, N.; Pesaresi, S.; Edoardo Biondi, E. The Paliurus spina-christi dominated vegetation in Europe, 2015. Biologia 2015, 70, 879–892. [Google Scholar] [CrossRef]
- USDA Forest Service. An Interim Guide to the Conservation and Management of Pacific Yew; USDA Forest Service: Portland, OR, USA, 1992. [Google Scholar]
- Piovesan, G.; Saba, E.P.; Biondi, F.; Alessandrini, A.; Di Filippo, A.; Schirone, B. Population ecology of yew (Taxus baccata L.) in the Central Apennines: Spatial patterns and their relevance for conservation strategies. Plant Ecol. 2009, 205, 23–46. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005; 302p. [Google Scholar]
- Núñez, M.R.; Calvo, L. Effect of high temperatures on seed germination of Pinus sylvestris and Pinus halepensis. For. Ecol. Manag. 2000, 131, 183–190. [Google Scholar] [CrossRef]
- Schütz, W.; Rave, G. The effect of cold stratification and light on the seed germination of temperate sedges (Carex) from various habitats and implications for regenerative strategies. Plant Ecol. 1999, 144, 215–230. [Google Scholar] [CrossRef]
- Catara, S.; Cristaudo, A.; Gualtieri, A.; Galesi, R.; Impelluso, C.; Onofri, A. Threshold temperatures for seed germination in nine species of Verbascum (Scrophulariaceae). Seed Sci. Res. 2016, 26, 30–46. [Google Scholar] [CrossRef]
- Papastergios, G.; Filippidis, A.; Christofides, G.; Kassoli-Fournaraki, A.; Fernández–Turiel, J.L.; Georgakopoulos, A.; Gimeno, D. Trace elements in uncultivated surface soils in the Kavala area, northeastern Greece. In Proceedings of the 11th International Congress, Athens, Greece, 24–26 May 2007. [Google Scholar]
- Papastergios, G. Environmental Geochemical Study of Soils and Sediments in Coastal Areas, East of Kavala (Macedonia, Greece) and Production of Geochemical Maps via the Use of GIS. Ph.D. Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2008; 224p. (In Greek, with English summary). [Google Scholar]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C. Ecology of the endangered species Solidago shortii. VI. Effects of habitat type, leaf litter, and soil type on seed germination. J. Torrey Bot. Soc. 1999, 126, 117–123. [Google Scholar] [CrossRef]
- Cirak, C. Seed germination protocols for ex situ conservation of some Hypericum species from Turkey. Am. J. Plant Phys. 2007, 2, 287–294. [Google Scholar]
- Shen, S.; Wang, Y.; Zhang, A.; Fuqin, W.; Jiang, L.; Jiang, L. Conservation and reintroduction of a critically endangered plant Euryodendron excelsum. Oryx 2013, 47, 13–18. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil Sample | Depth Cm | Rock Appearance (%) | Soil Texture | pH | Organic Matter (%) | C (%) | Total N (%) | C/N | P ppm | K ppm |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 13 | 85 | SL | 7.76 | 6.28 | 3.64 | 0.315 | 11.6 | 4.08 | 165 |
| 2 | 18 | 85 | SL | 7.83 | 6.86 | 3.98 | 0.356 | 11.2 | 9.09 | 320 |
| 3 | 14 | 80 | SL | 7.78 | 5.93 | 3.44 | 0.450 | 7.70 | 13.75 | 602 |
| 4 | 19 | 85 | L | 7.84 | 9.63 | 5.58 | 0.559 | 10.0 | 7.29 | 259 |
| Mean ± std error | 16 ± 1.47 | 83.8 ± 1.25 | SL | 7.80 ± 0.02 | 7.18 ± 0.84 | 4.16 ± 0.49 | 0.42 ± 0.054 | 10.1 ± 0.88 | 8.55 ± 2.02 | 336.5 ± 94.07 |
| Species | Family | Dominance | ||||
|---|---|---|---|---|---|---|
| Plot 1 | Plot 2 | Plot 3 | Plot 4 | Plot 5 | ||
| Euphorbia dendroides | Euphorbiaceae | 2a | 2m | 2m | 2b | 2a |
| Paliurus spina-christi | Rhamnaceae | 2a | 3 | 2a | 2a | 2b |
| Helianthemum sp. | Cistaceae | + | + | r | + | |
| Avena barbata | Poaceae | 1 | 1 | + | 2m | 1 |
| Crupina vulgaris | Asteraceae | R | + | + | ||
| Allium sphaerocephalon | Amaryllidaceae | 1 | 1 | + | + | 1 |
| Planta gobellardii | Plantaginaceae | 2m | + | 1 | 1 | 2m |
| V. dingleri | Scrophulariaceae | 1 | + | 1 | 1 | 1 |
| Asparagus acutifolius | Asparagaceae | 2m | 1 | + | + | 1 |
| Melica ciliata | Poaceae | 1 | + | 1 | 1 | 1 |
| Capsella bursa-pastoris | Brassicaceae | + | 1 | + | + | + |
| Oleaeuropea ssp. europaea | Oleaceae | 2b | 2a | 2b | 2b | 2a |
| Quercus coccifera | Fagaceae | 2b | 2a | 2b | 2a | 2b |
| Dracunculus vulgaris | Araceae | R | r | |||
| Hordeum sp. | Poaceae | 1 | + | 1 | + | |
| Bromustectorum | Poaceae | + | + | + | + | + |
| Vulpia sp. | Poaceae | 1 | 2m | + | 1 | |
| Fruit Traits | Seed Traits | ||||||
|---|---|---|---|---|---|---|---|
| Fruit Class | Mean Fruit Diameter/mm | Number of Mature Seeds | Number of Immature Seeds | Seed Length/Mm | Seed Fresh Weight/gr | Seed Dry Weight/gr | Seed Moisture Content (%) of Dry Mass |
| 1st | 3.66 ± 0.02a | 79.05 ± 4.50a | 0.42 ± 0.23c | 0.644 ± 0.018 ns | 5.0 × 10−4 | 3.9 × 10−4 | 22.9 |
| 2nd | 3.26 ± 0.03b | 57.04 ± 3.51b | 3.92 ± 1.48b | 0.626 ± 0.020 ns | 4.8 × 10−4 | 3.6 × 10−4 | 27.8 |
| 3rd | 2.61 ± 0.08c | 29.77 ± 3.88c | 5.61 ± 1.43a | 0.627 ± 0.033 ns | 4.6 × 10−4 | 3.4 × 10−4 | 29.4 |
| Mean | 3.24 ± 0.06 | 58.18 ± 3.37 | 3.04 ± 0.86 | 0.633 ± 0.014 | 4.8 × 10−4 | 3.6 × 10−4 | 26.7 |
| Correlations | ||||||
|---|---|---|---|---|---|---|
| Fruit Diameter | Fruit Class | N of Normal Seeds | N of Abnormal Seeds | Germination Final % | ||
| Fruit diameter | Pearson Correlation | 1.0 | −0.928 ** | 0.544 | 0.0 b | −0.364 |
| Sig. (2-tailed) | 0.000 | 0.130 | 0.335 | |||
| N | 64 | 64 | 64 | 12 | ||
| Fruit class | Pearson Correlation | −0.532 | 0.0 b | 0.105 | ||
| Sig. (2-tailed) | 0.141 | . | 0.787 | |||
| N | 64 | 64 | 12 | |||
| N of normal seeds | Pearson Correlation | 0.0 b | 0.051 | |||
| Sig. (2-tailed) | . | 0.896 | ||||
| N | 64 | 12 | ||||
| Seed Germination | |||
|---|---|---|---|
| In Laboratory Conditions | At the Nursery (Ambient Conditions) | ||
| Fruit Class | Final Germination (%) | Mean Germination Time (MGT) | Fully Developed Seedlings (%) |
| 1st | 32.0 | 24.7 | 22.0 |
| 2nd | 26.7 | 18.8 | 14.5 |
| 3rd | 40.0 | 17.8 | 9.5 |
| Significance | ns: p > 0.05 | ns: p > 0.05 | |
| Mean | 32.9 | 15.3 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganatsas, P.; Tsakaldimi, M.; Damianidis, C.; Stefanaki, A.; Kalapothareas, T.; Karydopoulos, T.; Papapavlou, K. Regeneration Ecology of the Rare Plant Species Verbascum dingleri: Implications for Species Conservation. Sustainability 2019, 11, 3305. https://doi.org/10.3390/su11123305
Ganatsas P, Tsakaldimi M, Damianidis C, Stefanaki A, Kalapothareas T, Karydopoulos T, Papapavlou K. Regeneration Ecology of the Rare Plant Species Verbascum dingleri: Implications for Species Conservation. Sustainability. 2019; 11(12):3305. https://doi.org/10.3390/su11123305
Chicago/Turabian StyleGanatsas, Petros, Marianthi Tsakaldimi, Christos Damianidis, Anastasia Stefanaki, Theodoros Kalapothareas, Theodoros Karydopoulos, and Kelly Papapavlou. 2019. "Regeneration Ecology of the Rare Plant Species Verbascum dingleri: Implications for Species Conservation" Sustainability 11, no. 12: 3305. https://doi.org/10.3390/su11123305