Potential Indicators of Soil Health Degradation in Different Land Use-Based Ecosystems in the Shiwaliks of Northwestern India

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site Description, Soil Sampling, and Basic Properties
2.2. Enzyme Assay
2.3. Culturable Soil Microbial Count
2.4. Statistical Analysis
3. Results
3.1. Soil Physio-Chemical Properties and Microfloral Composition as Potential Indicators of Soil Health
3.2. Soil Bacterial Composition in Different Land Use Systems and Soil Depths
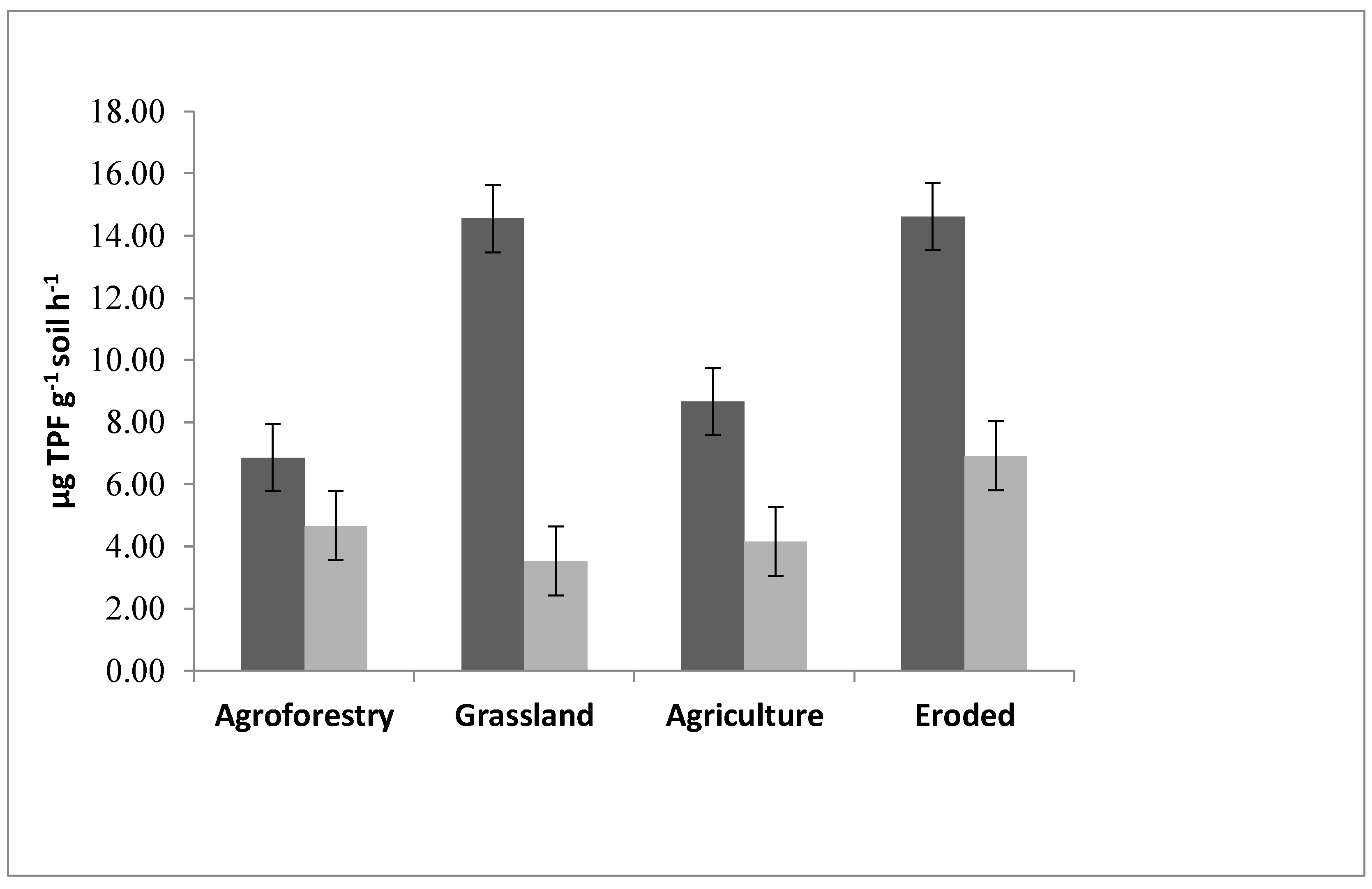
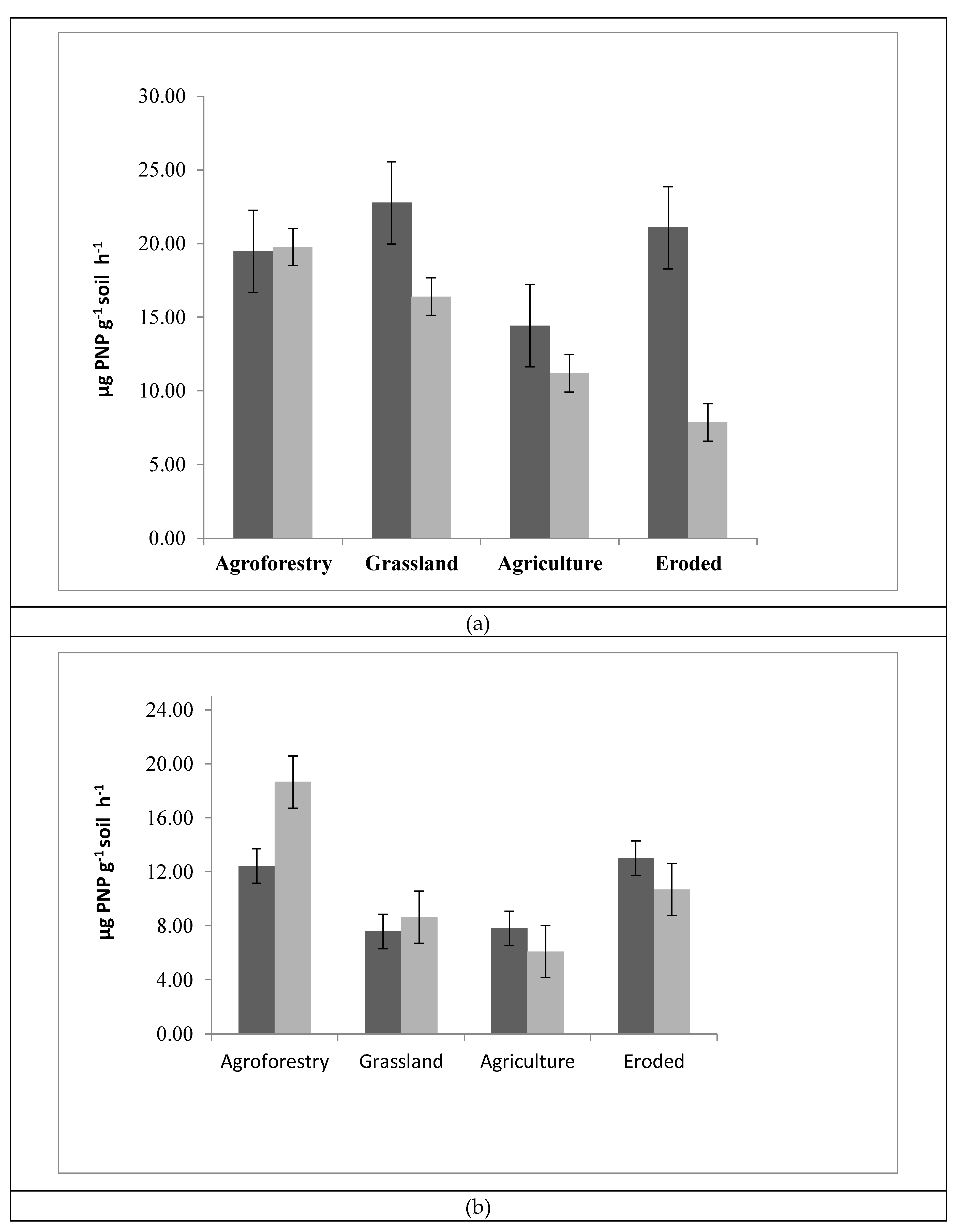
3.3. Enzyme Activities
3.4. Relationships between Soil Properties and Culturable Microbial Viable Cell Counts
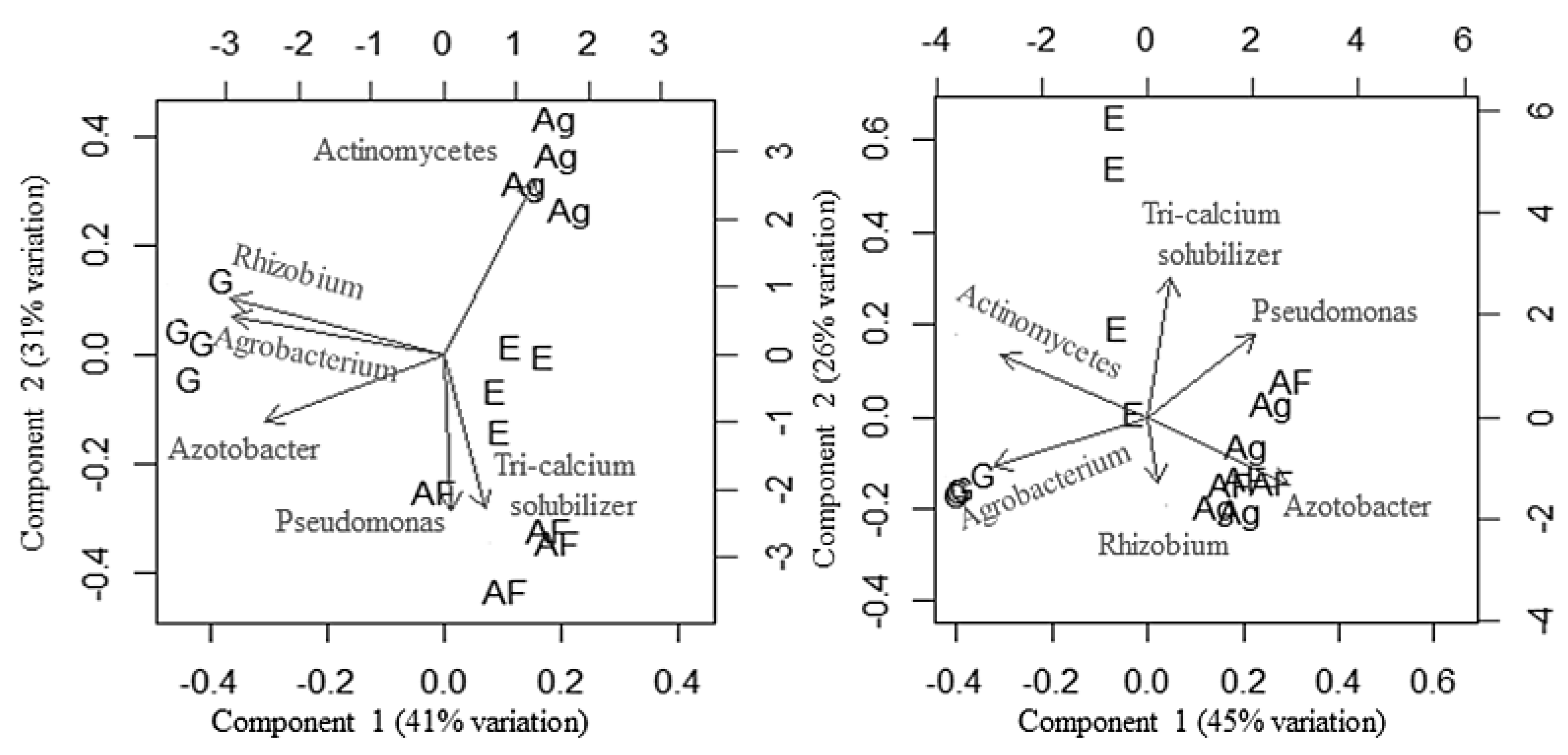
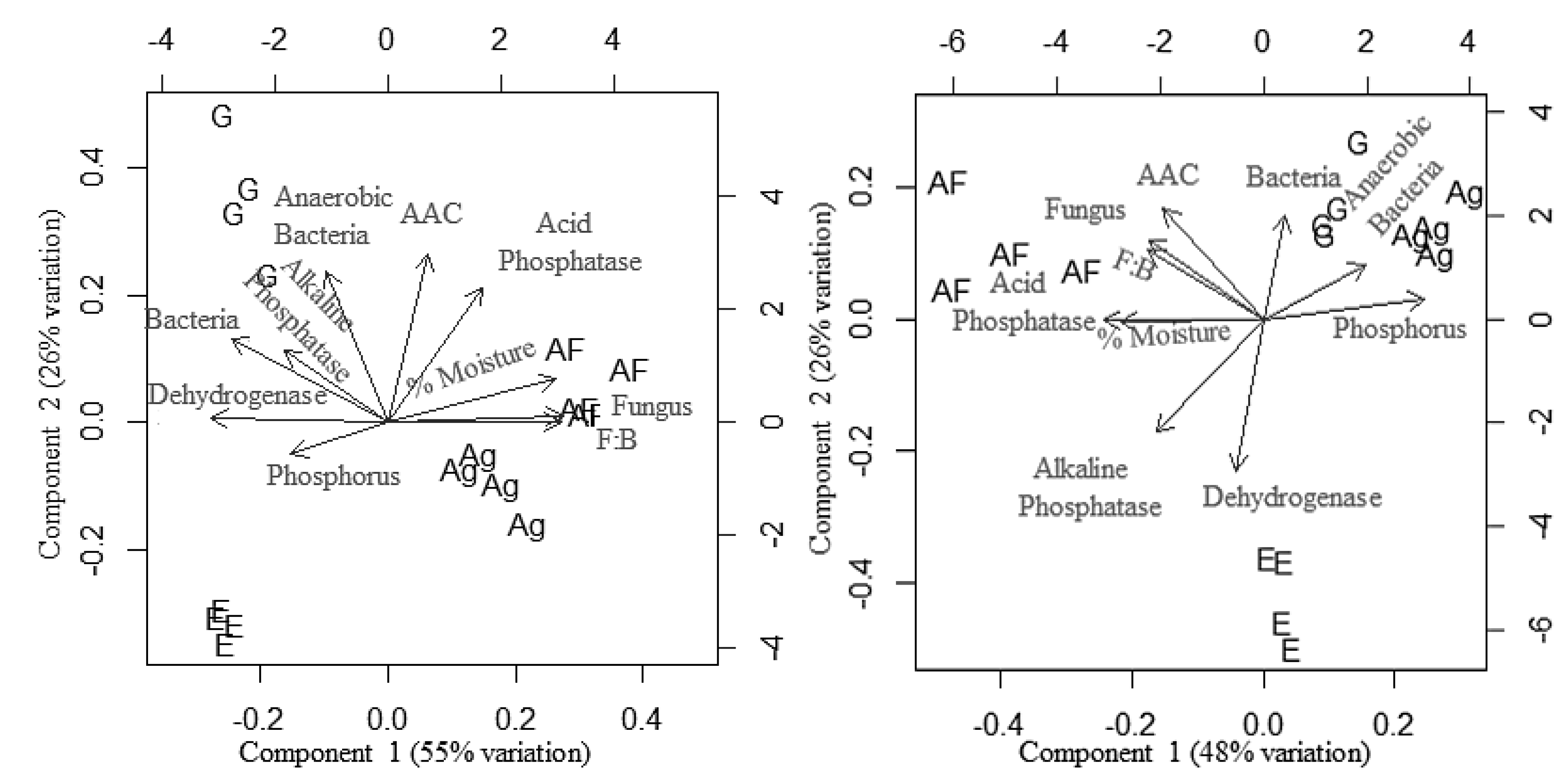
3.5. Effect of Land Use Systems and Soil Properties on Soil Microfloral Communities
4. Discussion
4.1. Soil Enzymatic Activity as an Indicator of Soil Health and Land Use System
4.2. Culturable Soil Microfloral Communities as Indicators of Soil Health and Land Use Change
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lal, R. Soil carbon dynamics in cropland and rangeland. Environ. Pollut. 2002, 116, 353–362. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of soil properties and microbial communities to agriculture: Implications for primary productivity and soil health indicators. Front. Plant Sci. 2016, 7, 990. [Google Scholar] [CrossRef] [PubMed]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- Doran, J.W.; Parkin, T.B. Defining and assessing soil quality. In Defining Soil Quality for a Sustainable Environment; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; American Society of Agronomy: Madison, WI, USA, 1994; pp. 3–21. [Google Scholar]
- Wall, D.H.; Virginia, R.A. Controls on soil biodiversity: Lessons from extreme environments. Appl. Soil Ecol. 1999, 13, 127–150. [Google Scholar] [CrossRef]
- Bhowmik, A.; Fortuna, A.M.; Cihacek, L.; Bary, A.; Cogger, C.G. Use of biological indicators of soil health to estimate reactive nitrogen dynamics in long-term organic vegetable and pasture systems. Soil Biol. Biochem. 2016, 103, 308–319. [Google Scholar] [CrossRef] [Green Version]
- Bhowmik, A.; Fortuna, A.M.; Cihacek, L.; Bary, A.; Carr, P.M.; Cogger, C.G. Potential carbon sequestration and nitrogen cycling in long term organic management systems. Renew. Agric. Food Syst. 2017, 32, 498–510. [Google Scholar] [CrossRef]
- Bhowmik, A.; Fortuna, A.M.; Cihacek, L.; Rahman, S.; Borhan, M.S.; Carr, P. Use of Laboratory Incubation Techniques to Estimate Green House Gas footprints from Conventional and No-Tillage Organic Agroecosystems. Soil Biol. Biochem. 2017, 112, 204–215. [Google Scholar] [CrossRef]
- Saha, D.; Kukal, S.S.; Sharma, S. Landuse impacts on SOC fractions and aggregate stability in typic ustochrepts of Northwest India. Plant Soil 2011, 339, 457–470. [Google Scholar] [CrossRef]
- Brevik, E.C.; Sauer, T.J. The past, present, and future of soils and human health studies. Soil 2015, 1, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Grego, S.; Ceccanti, B. Ecological significance of the biological activity in soils. In Soil Biochemistry; Bollag, J.M., Stotzky, G., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1990; pp. 293–355. [Google Scholar]
- Guo, L.B.; Gifford, R.M. Soil carbon stocks and land use change: A meta-analysis. Glob. Chang. Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis; Page, A.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1994; pp. 775–834. [Google Scholar]
- Acosta-Martinez, V.; Zobeck, T.M.; Allen, V. Soil microbial, chemical and physical properties in continuous cotton and integrated crop-livestock systems. Soil Sci. Soc. Am. J. 2004, 68, 1875–1884. [Google Scholar] [CrossRef]
- Kandeler, E.; Kampichler, C.; Horak, O. Influence of heavy metals on the functional diversity of soil microbial communities. Biol. Fert. Soils 1996, 23, 299–306. [Google Scholar] [CrossRef]
- Bandick, A.K.; Dick, R.P. Field management effects on soil enzyme activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Trevors, J.T. Dehydrogenase activity in soil: A comparison between the INT and TTC assay. Soil Biol. Biochem. 1984, 16, 673–674. [Google Scholar] [CrossRef]
- Ndiaye, E.L.; Sandeno, J.M.; McGrath, D.; Dick, R.P. Integrative biological indicators for detecting change in soil quality. Am. J. Altern. Agric. 2000, 15, 26–36. [Google Scholar] [CrossRef]
- Moore, J.M.; Klose, S.; Tabatabai, M.A. Soil microbial biomass carbon and nitrogen as affected by cropping systems. Biol. Fert. Soils 2000, 31, 200–210. [Google Scholar] [CrossRef]
- Kukal, S.S.; Sur, H.S.; Gill, S.S. Factors responsible for soil erosion hazard in submontane Punjab. Soil Use Manag. 1991, 7, 38–44. [Google Scholar] [CrossRef]
- Sitaula, B.K.; Bajracharya, R.M.; Singh, B.R.; Solberg, B. Factors affecting organic carbon dynamics in soils of nepal/himalayan region-a review and analysis. Nutr. Cycl. Agroecosyst. 2004, 70, 215–229. [Google Scholar] [CrossRef]
- Gupta, N.; Kukal, S.S.; Bawa, S.S.; Dhaliwal, G.S. Soil organic carbon and aggregation under poplar based agroforestry system in relation to tree age and soil type. Agrofor. Syst. 2009, 76, 27–35. [Google Scholar] [CrossRef]
- De, M.; Saha, D.; Chakraborty, S. Soil structure and strength characteristics in relation to slope segments in a degraded Typic Ustochrepts of Northwest India. Soil. Horizons. 2014, 55, 1. [Google Scholar] [CrossRef]
- Saha, D.; Kukal, S.S. Soil structural stability and water retention characteristics under different land uses of degraded lower Himalayas of North-West India. Land Degrad. Dev. 2015, 26, 263–271. [Google Scholar] [CrossRef]
- Blake, G.R.; Hartge, K.H. Bulk density. In Methods of Soil Analysis, Part I; Klute, A., Ed.; ASA Monograph No. 9: Madison, WI, USA, 1986; pp. 363–376. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Frank, S.; Watanabe, F.S. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA: Washington, DC, USA, 1954; p. 18. [Google Scholar]
- Yoder, R.E. A direct method of aggregate size analysis of soils and a study of the physical nature of erosion losses. J. Am. Soc. Agron. 1936, 28, 337–351. [Google Scholar] [CrossRef]
- Kemper, W.D.; Rosenau, R.C. Aggregate stability and size distribution. In Methods of Soil Analysis, Part 1; Klute, A., Ed.; American Society of Agronomy: Madison, WI, USA, 1986; pp. 425–442. [Google Scholar]
- Youker, R.E.; McGuinness, J.L. A short method of obtaining mean weight diameter values of aggregate analyses of soils. Soil Sci. 1956, 83, 291–294. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analysis, Part 2: Chemical and Microbiological Properties; Page, A.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 903–948. [Google Scholar]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press Ltd.: London, UK, 1998. [Google Scholar]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and regression by randomForest. R News 2002, 2, 18–22. [Google Scholar]
- Bastida, F.; Moreno, J.L.; Hernández, T.; García, C. Microbiological degradation index of soils in a semiarid climate. Soil Biol. Biochem. 2006, 38, 3463–3473. [Google Scholar] [CrossRef]
- Melero, S.; Vanderlinden, K.; Ruiz, J.C.; Madejon, E. Long-term effect on soil biochemical status of a vertisol under conservation tillage system in semi-arid Mediterranean conditions. Eur. J. Soil Biol. 2008, 44, 437–442. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, W.; Burger, M.; Yang, L.; Gong, P.; Wu, Z. Changes in soil carbon and enzyme activity as a result of different long-term fertilization regimes in a greenhouse field. PLoS ONE 2015, 10, e0118371. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Juma, N.G.; Tabatabai, M.A. Phosphatase activity in corn and soybean roots: Conditions for assay and effects of metals. Plant Soil 1988, 107, 39–47. [Google Scholar] [CrossRef]
- Yang, J.; Hu, C.C.; Hu, H.; Yu, R.D.; Xia, Z.; Ye, X.Z.; Zhu, J. QTL Network: Mapping and visualizing genetic architecture of complex traits in experimental populations. Bioinformatics 2008, 24, 721–723. [Google Scholar] [CrossRef] [PubMed]
- Speir, T.W.; Ross, D.J. Soil phosphatase and sulphatase. In Soil Enzymes; Burns, R.G., Ed.; Academic Press: London, UK, 2006; pp. 197–250. [Google Scholar]
- Kumar, A.; Rakshit, R.; Bhowmik, A.; Mandal, N.; Das, A.; Adhikary, S. Nanoparticle-Induced Changes in Resistance and Resilience of Sensitive Microbial Indicators towards Heat Stress in Soil. Sustainability 2019, 11, 862. [Google Scholar] [CrossRef]
- Li, D.; Zhu, H.; Liu, K.; Liu, X.; Leggewie, G.; Udvardi, M.; Wang, D. Purple acid phosphatases of Arabidopsis thaliana: Comparative analysis and differential regulation by phosphate deprivation. J. Biol. Chem. 2002, 277, 27772–27781. [Google Scholar] [CrossRef] [PubMed]
- Kai, M.; Takazumi, K.; Adachi, H.; Wasaki, J.; Shinano, T.; Osaki, M. Cloning and characterization of four phosphate transporter cDNAs in tobacco. Plant Sci. 2002, 163, 837–846. [Google Scholar] [CrossRef]
- Grierson, P.F.; Adams, M.A. Plant species affect acid phosphatase, ergosterol and microbial P in a Jarrah (Eucalyptus marginata n ex SM) forest in south-western Australia. Soil Biol. Biochem. 2000, 32, 1817–1827. [Google Scholar] [CrossRef]
- Arutyunyan, E.A.; Galstyan, A.S.H. Determination of the activity of alkaline and acid phosphatase in soils. Agrochimija 1975, 5, 128–133. [Google Scholar]
- Six, J.; Frey, S.D.; Thiet, R.K.; Batten, K.M. Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Sci. Soc. Am. J. 2006, 70, 555–569. [Google Scholar] [CrossRef]
- Malik, A.A.; Chowdhury, S.; Schlager, V.; Oliver, A.; Puissant, J.; Vazquez, P.G.M.; Jehmilch, N.; Bergen, M.V.; Griffiths, R.I.; Gleixner, G. Soil fungal:bacterial rations are linked to altered carbon cycling. Front. Microbiol. 2016, 7, 1247. [Google Scholar] [CrossRef]
- Waring, B.G.; Averill, C.; Hawkes, C.V. Differences in fungal and bacterial physiology alter soil carbon and nitrogen cycling: Insights from meta-analysis and theoretical models. Ecol. Lett. 2013, 16, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon use efficiency of microbial communities: Stoichiometry, methodology and modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.; Kandji, S.T. Carbon sequestration in tropical agroforestry systems. Agric. Ecosyst. Environ. 2003, 99, 15–27. [Google Scholar] [CrossRef]
- Montagnini, F.; Nair, P.K.R. Carbon sequestration: An underexploited environmental benefit of agroforestry systems. Agrofor. Syst. 2004, 61, 281–295. [Google Scholar]
- Joshi, S.R.; Saikia, P.; Koijam, K. Characterization of microbial indicators to assess the health of degraded soil in Cherrapunjee, India-highest rainfall area of the world. Int. J. Biotechnol. Biochem. 2009, 5, 379–391. [Google Scholar]
- Elliot, L.F.; Lynch, J.M. The international workshop on establishment of microbial inocula in soils: Cooperative research project on biological resource management of the Organization for Economic Cooperation and Development (OECD). Am. J. Altern. Agric. 1995, 10, 50–73. [Google Scholar] [CrossRef]
- Ghorbani-Nasrabadi, R.; Greiner, R.; Alikhani, H.; Hamedi, J.; Yakhchali, B. Distribution of Actinobacteria in different soil ecosystems and effect of media composition on extracellular phosphatase activity. J. Soil Sci. Plant Nutr. 2013, 13, 223–236. [Google Scholar]
- Moreno-De Las Heras, M. Development of soil physical structure and biological functionality in mining spoils affected by soil erosion in a Mediterranean-Continental environment. Geoderma 2009, 149, 249–256. [Google Scholar] [CrossRef]
- Richardson, A.E. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants. Funct. Plant Biol. 2001, 28, 897–906. [Google Scholar] [CrossRef]
- Köberl, M.; Dita, M.; Martinuz, A.; Staver, C.; Berg, G. Agroforestry leads to shifts within the gammaproteobacterial microbiome of banana plants cultivated in Central America. Front. Microbiol. 2015, 6, 91. [Google Scholar]
- Radhapriya, P.; Ramachandran, A.; Anandham, R.; Mahalingam, S. Pseudomonas aeruginosa RRALC3 enhances the biomass, nutrient and carbon contents of Pongami pinnata seedlings in degraded forest soil. PLoS ONE 2015, 10, e0139881. [Google Scholar] [CrossRef] [PubMed]
- Berthrong, S.T.; Yeager, C.M.; Gallegos-Graves, L.; Steven, B.; Eichorst, S.A.; Jackson, R.B.; Kuske, C.R. Nitrogen Fertilization Has a Stronger Effect on Soil Nitrogen-Fixing Bacterial Communities than Elevated Atmospheric CO2. Appl. Environ. Microbiol. 2014, 80, 3103–3112. [Google Scholar] [CrossRef] [PubMed]
- Oelze, J. Respiratory protection of nitrogenase in Azotobacter species: Is a widely held hypothesis unequivocally supported by experimental evidence? FEMS Micorbiol. Rev. 2000, 24, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Kibunja, C.N.; Mwaura, F.B.; Mugendi, D.N. Long-term land management effects on soil properties and microbial populations in a maize-bean rotation at Kabete, Kenya. Afr. J. Agric. Res. 2010, 5, 108–113. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agroforestry | Grassland | Agriculture | Eroded | |||||
|---|---|---|---|---|---|---|---|---|
| 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm | 0–15 cm | 15–30 cm | |
| pH | 6.61 ± 0.01 | 6.90 ± 0.00 | 7.51 ± 0.00 | 7.61 ± 0.00 | 7.62 ± 0.01 | 7.59 ± 0.01 | 7.50 ± 0.02 | 7.60 ± 0.01 |
| EC (dS/m) | 0.58 ± 0.03 | 0.56 ± 0.04 | 0.60 ± 0.01 | 0.56 ± 0.02 | 0.60 ± 0.02 | 0.58 ± 0.01 | 0.64 ± 0.00 | 0.60 ± 0.00 |
| BD (g cc−1) | 1.51 ± 0.02 | 1.48 ± 0.01 | 1.43 ± 0.02 | 1.45 ± 0.01 | 1.45 ± 0.03 | 1.46 ± 0.01 | 1.30 ± 0.08 | 1.28 ± 0.06 |
| AAC g kg−1 | 0.77 ± 0.04 | 0.84 ± 0.01 | 0.99 ± 0.03 | 0.45 ± 0.06 | 0.73 ± 0.05 | 0.58 ± 0.03 | 0.36 ± 0.01 | 0.25 ± 0.02 |
| Textural class | SL | SL | SL | SL | SL | SL | SL | SL |
| FC % | 23.4 ± 1.2 | 23.8 ± 1.4 | 19.5 ± 2.1 | 23.5 ± 1.09 | 10.8 ± 1.7 | 11.1 ± 2.1 | 17.5 ± 2.3 | 21.4 ± 2.0 |
| PWP % | 4.60 ± 0.10 | 3.80 ± 0.10 | 4.00 ± 0.30 | 4.9 ± 0.1 | 1.60 ± 0.10 | 2.20 ± 0.20 | 2.90 ± 0.20 | 6.80 ± 0.10 |
| NH4-N (kg/ha) | 11.3 ± 0.2 | 20.4 ± 0.5 | 12.6 ± 0.1 | 5.0 ± 0.7 | 10.1 ± 0.3 | 22.3 ± 1.2 | 10.4 ± 0.3 | 12.4 ± 0.9 |
| NO3-N (kg/ha) | 10.1 ± 0.1 | 8.70 ± 1.10 | 10.4 ± 0.2 | 4.8 ± 1.2 | 7.50 ± 0.20 | 9.90 ± 0.80 | 7.4 ± 0.3 | 12.1 ± 0.6 |
| Available P (kg/ha) | 16.7 ± 0.4 | 14.5 ± 1.1 | 16.5 ± 0.5 | 22.1 ± 0.8 | 13.0 ± 0.9 | 22.7 ± 0.6 | 17.8 ± 1.2 | 20.3 ± 0.5 |
| Macroaggregate > 2 mm (%) | 76.2 ± 1.2 | 83.4 ± 1.5 | 95.6 ± 3.4 | 52.4 ± 2.2 | 50.5 ± 4.2 | 37.8 ± 2.4 | 39.3 ± 9.9 | 27.5 ± 7.8 |
| Microaggregates < 2 mm (%) | 23.6 ± 3.2 | 16.5 ± 0.9 | 4.40 ± 1.00 | 47.6 ± 3.6 | 50.1 ± 11.2 | 62.0 ± 10.6 | 60.3 ± 11.7 | 71.8 ± 14.6 |
| MWD (mm) | 3.97 ± 0.90 | 4.27 ± 1.10 | 4.80 ± 0.8 | 3.20 ± 0.4 | 2.85 ± 1.00 | 2.41 ± 0.50 | 2.37 ± 1.3 | 1.38 ± 0.80 |
| 0–15 cm Soil Depth | ||||
|---|---|---|---|---|
| Land Use | Bacteria (× 106) | Fungus (× 103) | Actinobacteria (× 104) | Anaerobic (× 104) |
| Agroforestry | 5.00 | 10.0 | 0.30 | 2.00 |
| Grassland | 18.0 | 2.00 | 3.00 | 10.6 |
| Agriculture | 8.00 | 7.00 | 70.00 | 3.90 |
| Eroded | 12.0 | 2.00 | 0.60 | 0.80 |
| LSD (0.05) | 2.60 | 1.60 | 9.49 | 3.22 |
| 15–30 cm Soil Depth | ||||
| Agroforestry | 3.00 | 1.00 | 0.10 | 0.90 |
| Grassland | 3.00 | 1.00 | 1.20 | 1.90 |
| Agriculture | 3.00 | - | 0.20 | 20.6 |
| Eroded | 2.00 | - | 1.40 | 1.40 |
| LSD (0.05) | NS | 0.59 | 0.53 | 5.83 |
| 0–15 cm Soil Depth | |||||
|---|---|---|---|---|---|
| Land Use | Tri calcium Solubilizers (× 103) | Fluorescent Pseudomonas (× 104) | Azotobacter (× 104) | Rhizobium (× 104) | Agrobacterium (× 104) |
| Agroforestry | 10.75 | 40.0 | 9.25 | 1.15 | 3.15 |
| Grassland | 5.50 | 13.5 | 11.5 | 12.3 | 32.8 |
| Agriculture | 1.75 | 7.00 | 5.75 | 3.50 | 7.00 |
| Eroded | 14.3 | 1.00 | 4.50 | 3.05 | 14.0 |
| LSD (0.05) | 5.75 | 5.00 | 2.33 | 3.29 | 2.38 |
| 15–30 cm Soil Depth | |||||
| Agroforestry | 5.00 | 5.60 | 31.5 | 10.2 | 3.10 |
| Grassland | 0.18 | 0.23 | 4.23 | 5.25 | 30.3 |
| Agriculture | 0.88 | 3.75 | 36.5 | 0.48 | 1.75 |
| Eroded | 9.50 | 4.25 | 3.50 | 0.80 | 5.20 |
| LSD (0.05) | 5.25 | 3.00 | 6.71 | 4.34 | 3.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhowmik, A.; Kukal, S.S.; Saha, D.; Sharma, H.; Kalia, A.; Sharma, S. Potential Indicators of Soil Health Degradation in Different Land Use-Based Ecosystems in the Shiwaliks of Northwestern India. Sustainability 2019, 11, 3908. https://doi.org/10.3390/su11143908
Bhowmik A, Kukal SS, Saha D, Sharma H, Kalia A, Sharma S. Potential Indicators of Soil Health Degradation in Different Land Use-Based Ecosystems in the Shiwaliks of Northwestern India. Sustainability. 2019; 11(14):3908. https://doi.org/10.3390/su11143908
Chicago/Turabian StyleBhowmik, Arnab, Surinder Singh Kukal, Debasish Saha, Harmandeep Sharma, Anu Kalia, and Sandeep Sharma. 2019. "Potential Indicators of Soil Health Degradation in Different Land Use-Based Ecosystems in the Shiwaliks of Northwestern India" Sustainability 11, no. 14: 3908. https://doi.org/10.3390/su11143908