The History of Pastoral Activities in S Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness


Laboratorio di Palinologia e Paleobotanica, Dip. Scienze Vita, Università degli Studi di Modena e Reggio Emilia, 41100 Modena, Italy
Sustainability 2019, 11(2), 404; https://doi.org/10.3390/su11020404
Submission received: 13 December 2018
/
Revised: 7 January 2019
/
Accepted: 11 January 2019
/
Published: 15 January 2019
(This article belongs to the Special Issue The Long-Term Perspective of Human Impact on Landscape for Environmental Change and Sustainability)
Abstract
:The present-day Mediterranean landscape is a result of the long-term human–environment–climate interactions that have driven the ecological dynamics throughout the Holocene. Pastoralism had (and still has) an important role in shaping this landscape, and contributes to maintaining the mosaic patterns of the Mediterranean habitats. Palaeoecological records provide significant multi-proxy data on environmental changes during the Holocene that are linked to human activities. In such research, the palynological approach is especially useful for detailing the complexity of anthropogenically-driven landscape transformations by discriminating past land uses and pastoral/breeding activities. This paper focuses on the palynological evidence for the impact of centuries of grazing on the vegetation of Basilicata, a region of southern Italy where animal breeding and pastoralism have a long tradition. A set of 121 pollen samples from eight archaeological sites (dated from the 6th century BC to the 15th century AD) and five modern surface soil samples were analyzed. The joint record of pollen pasture indicators and spores of coprophilous fungi suggests that continuous and intense pastoral activities have been practiced in the territory and have highly influenced its landscape. The palaeoecological results of this study provide us with better knowledge of the diachronical transformations of the habitats that were exposed to continuous grazing, with a shift toward more open vegetation and increase of sclerophyllous shrubs. The palynological approach gives insights into the vocation and environmental sustainability of this southern Italy region on a long-term basis.
1. Introduction
Since prehistoric times, humans have directly (by activities) or indirectly (by presence) influenced ecosystems and landscapes around the world [1]. Human actions have resulted in several changes in the biosphere by influencing and driving the main variations in biodiversity and ecosystem processes [2]. From a long-term perspective, human influence and then—depending on the time and space scale—impact have generated notable patterns of landscape complexity [3,4,5,6]. This is especially evident in the Mediterranean area, where, since ancient times, the continuous succession of different civilizations has had direct and indirect effects on the environment that led to changes in the vegetation cover [7,8,9]. Grazing is one of the most important factors determining patterns of vegetation in Mediterranean ecosystems [10,11,12]. In the Mediterranean basin, long-term pastoral activities have had impressive consequences for the current biodiversity [13]. It is widely recognized that grazing prevents the growth of woody vegetation and reduces the species richness, and that intensive grazing profoundly affects the succession in such systems [14,15,16,17]. Moreover, it is well-known that grazing also affects the morphological and functional traits of plants at the global level [18].
Nevertheless, grazing should not be considered a negative factor that has a detrimental impact on plant biodiversity. It actually improves the quality of vegetation by removing old plants and stimulating grass production [19]. Recent multidisciplinary studies (integrating palaeoecological, pedological, phylogenetic, and palaeontological evidence) have shown the key role that wild herbivores and domestic livestock play in grassland–forest dynamics, contributing to the maintenance of the European mosaic-pattern vegetation that was created by megaherbivore disturbances at the Pleistocene–Holocene boundary [20,21].
Over the last few decades, several ecological studies dealing with grazing’s effects on vegetation and the environment have been carried out in order to obtain information essential to improving landscape management and sustainable resources exploitation [22,23,24,25]. These studies are based on short-term ecological data that span only a few decades, and they do not provide potentially significant trends in these records, nor allow us to predict possible future scenarios of biosphere responses to global changes. Indeed, it is the study of long-term ecological records (greater than 50 years) that actually has great potential in conservation biology, as it provides us with a valuable historical perspective on the dynamics of contemporary ecological systems [26,27,28,29,30]. Palaeoecological records, which provide multi-proxy palaeoenvironmental data, are essential for understanding details of long-term human impacts on ecosystems [31,32]. Among the multi-proxy analyses, palynological investigations allow us to acquire a high-resolution data set on environmental changes during the Holocene that are linked to human activities (e.g., [4,33]). Microscopic biological remains (pollen and non-pollen palynomorphs (NPPs), such as fungal spores and algal elements) from sedimentary records are meaningful bio-indicators for palaeoecological reconstruction (e.g., [34,35,36,37]). In particular, the combined evidence of pollen and NPPs from archaeological sites is useful for tracing the impact of past human activities on the local ecosystem; these biomarkers can also be used to discriminate past land uses and pastoral/breeding practices (e.g., [38,39,40,41,42]).
A palynological approach to the study of Mediterranean landscapes is ideally suited for detecting the land-use history and ecosystem changes that gave rise to the present-day Mediterranean environment. Such challenging issues have been addressed, for example, by Jouffroy-Bapicot and colleagues in the framework of the multidisciplinary research project on the making of the Cretan Mountain Landscape, Greece [43]. Multi-proxy analyses of the Asi Gonia peat bog sediments have shown that, during the two last millennia, pastoralism had a predominant role in vegetation dynamics. Other studies dealing with palynological investigations in grazing areas have been conducted on modern analogues in the Pyrenees Mountains to characterize pollen and non-pollen palynomorphs (NPP) indicators of types of highland vegetation and grazing pressure [44,45,46,47]. In the same study area, previous palaeoenvironmental studies that were carried out in several peat basins have proved that these wetlands were grazed in the past [48,49].
Previous Studies and the Aim of This Paper
In Italy, an outstanding number of studies that include pollen or other plant remains from archaeological contexts (BRAIN database: brainplants.successoterra.net) [50] have testified to the widespread occurrence of breeding/pastoral activities on the peninsula over the last few millennia. This long-term practice can also be inferred from pollen spectra from the top cores (Holocene sediments) of biostratigraphical records [51,52,53,54,55]. However, only a few papers focused solely on past pastoral practices and their legacy on the current Italian landscape have been published thus far [56]. On this issue, palaeoenvironmental research in southern Italy—and especially research carried out in Basilicata, on which this paper is based—can serve as a most suitable example. Here, animal breeding and pastoralism have a long tradition and have certainly played an important role in shaping the landscape. The PhD project on the archaeo-environmental reconstruction of this region has provided a palynological dataset that may be of key relevance for understanding the pastoralism that has been practiced in the area over the last 2500 years [57]. Some aspects of this research have already been published, such as the environmental and economic settings of the rural sites of the Greek colonial system on the coastal plain [58,59,60,61], and the agro-pastoral characterization of the inland indigenous sites [62,63,64,65]. Besides this, some specific issues that emerged from the research have been further explored (e.g., the value of Cichorieae as a pastoral indicator [66] and the Olea pollen representativeness in the modern local olive groves [67]).
This work focuses on the palynological evidence of pastoral/breeding activities in the studied archaeological records and explores the impact of centuries of grazing on the vegetation of Basilicata. The main aim of this study is to better understand the complexity of landscape transformations that have occurred in these lands, which have continuously been exploited as pastures, thus as to improve our awareness of the biodiversity in, and the long-term human impact on, the current landscape of a typical region of southern Italy. As part of the historically underdeveloped area of Mezzogiorno, in the last few decades, Basilicata has faced several challenges due to its geological instability, peripherality, and marked depopulation. To improve its economic development, the Italian government has implemented policy interventions and development strategies that are supported by European programmes, which threaten to transform the natural and cultural assets of the region. A long-term perspective on the development of its cultural landscape is fundamental to provide strategies to avoid the loss of environmental peculiarities and biodiversity that comply with the cultural identity and vocation of this territory.
2. Materials and Methods
2.1. Study Area
This research has been carried out on samples from the main archaeological sites of the Basilicata region, which are located in a transect from the southern Apennines (1 site) to the Ionian coast along the Bradano river (7 sites). The eight sites (Figure 1, Table 1) have been studied in collaboration with the University of Basilicata (sites 1–4) [68,69] and the University of Texas at Austin (sites 5–8) [70,71,72].
Torre di Satriano (site 1) is the only high-ground site (c. 900 m asl); although it is subject to very cold winter temperatures, it is located in a territory rich in natural resources, such as a vast supply of water, extensive hilly areas for pastures and cultivations, and wood. The archaeological excavations brought to light the remains of an outstanding dwelling, which has been interpreted as the center of power within the local community between 560/550 BC and 480 BC [73]. The sites of the low-hill inland area (2–4) lie along the Bradano river, which was an important waterway for the transport of goods. These sites are fortified rural settlements that span from the Archaic/Hellenistic (4) to the Roman periods and the Middle Ages/Renaissance (2,3), and their displacement on the top of the hills is due to both defense and control reasons [68,69]. In fact, the river has always represented the way for foreign populations (such as the Greek settlers) who were attracted to the lands for farming to access the territory. The sites of the coastal plain (5–8), which all belong to the chora (rural territory) of the Greek colonial city of Metaponto, are located mainly on the first marine terraces a few meters above the plain, which is often subject to floods. The network of small farmhouses and rural villages along the Bradano river had a crucial role in setting up the local farming system and regiment the water course and its floods [71].
The climate of the study area has a typical Mediterranean seasonality. The vegetation typologies that occur in the region depend on the high environmental diversity of the territory [74]. Most of the study area is characterized by the Mesotemperate Turkey oak vegetation series of Lathyro digitati-Quercetum cerridis [75]. In the Apennine sector, anthropogenic meadows (including grassland species such as Trifolium nigrescens Viv., Medicago hispida Gaertner, Dactylis glomerata L. subsp. glomerata, Centaurea centaurium L. and Scorpiurus muricatus L.), and mixed oakwoods (mainly composed of Quercus cerris L., Q. frainetto Ten., Q. virgiliana Ten., Q. pubescens Willd., usually with Carpinus orientalis Mill., Fraxinus ornus L., Acer monspessolanum L.) are the main vegetation [75]. The low-hill inland area is covered with shrubby Mediterranean plants (Pistacia lentiscus L., Phillyrea latifolia L., Cistus creticus L., Helianthemum jonium Lacaita and other elements related to Pistacio Rhamnetalia alaterni plant communities) that are associated with grasses and annual herbs included in the Camphorosmo-monspeliacae-Lygeetum sparti and Medicago coronate-Hedysaretum glomerati vegetation series. In the Metaponto area, the relatively recent Pinus halepensis reforestations influence the dune communities, which the Salsolo kali-Cakiletum maritimae and Malcomietalia series dominate [75,76]. Vegetables and fruits, especially cherry orchards, citrus groves, and vineyards, grow in the mild climate of the coastal plain [76].
2.2. Pollen Samples and Analysis
A set of 121 pollen samples were taken from archaeological layers—small trenches, rooms or floors of houses, and spot samples from the eight sites. Sometimes also fillings of pottery were analysed, but data were not used for palaeoenvironmental reconstructions. In addition, five pollen samples were collected from the current surface soils to obtain a reference of the modern composition of local pollen rain. According to the chronology of the archaeological sites, the temporal transect spans about 20 centuries—from the 6th century BC to the 15th century AD (Table 1).
Pollen samples, of about 8 g each, were prepared using tetra NaPyrophosphate, HCl 10%, acetolysis, separation with NaMetatungstate hydrate, HF 40%, and ethanol [77]. Lycopodium tablets were added to calculate concentrations (p/g = pollen per gram of sediment, and npp/g = npp per gram) [78]. Permanent pollen slides were mounted on glycerol jelly. Identification was made at 400× and 1000× magnification, with the help of keys, atlases [79,80,81], and the reference pollen collection of the Laboratory of Palynology and Palaeobotany of the University of Modena and Reggio Emilia. Non-pollen palynomorphs (NPPs, microfossils of a great variety of organisms—mainly fungi and algae—sensitive to various ecological parameters or to human presence) were identified according to the reference literature [82,83,84]. Among NPPs, particular attention has been paid to the coprophilous fungal spores (e.g., Sordaria, Sporormiella, Delitschia, Podospora types) as indices of the presence of dung, and therefore associated with pastoral/breeding activities [38,47].
Pollen and NPPs were counted in the same samples. On average, about 350 pollen grains and 200 NPPs per sample were counted. The percentage of pollen diagrams were basically calculated from pollen sums including all pollen counted. Five pollen groups which refer to the plant landscape and human activities indicators were calculated: (a) Mixed oakwood (including broadleaved Quercus-oaks, Carpinus betulus-hornbeam, Ostrya carpinifolia-hop hornbeam, Fraxinus-ash, Tilia-linden, Ulmus-elm, and Corylus-hazel); (b) Mediterranean plants (Quercus ilex-evergreen oak type, Daphne cf. gnidium-flax-leaved daphne, Helianthemum-rock rose, Juniperus-juniper type, Erica-heat, Olea-olive tree, Phillyrea-green olive tree, Pistacia cf. lentiscus-lentisk, Rhamnus-buckthorn type); (c) Hydro-hygrophilous plants (sum of plants from wet environments: Alnus-alder, Cyperaceae-sedges, Nymphaea cf. alba-white water lily, Phragmites cf. australis-common reed, Populus-poplar, Ranunculus cf. macrophyllus-large leaved buttercup, Sagittaria-arrowhead, Typha/Sparganium-cattail/bur-reed type); (d) cultivated/cultivable plants (the woody plants Castanea-chestnut, Corylus-hazelnut, Juglans-walnut, Olea, Prunus-plum, Malus/Pyrus-apple/pear, Vitis-grapevine, and cereals Avena/Triticum-oats/wheat group and Hordeum-barely group); (e) local pastoral pollen indicators (LPPI) [85], including plants strictly linked to local pastoral activities (Cirsium, Centaurea nigra type and other Asteroideae, Ranunculaceae, Galium type, Potentilla type, and Cichorieae—the tribe with fenestrate pollen within the Cichorioideae subfamily [66]).
The pollen diagrams were drawn with TGView [86]. On-site pollen data were chronologically ordered and grouped according to the main time phases of the sites. Following Mercuri [87,88], the chronological pollen samples groups were treated as a single ‘regional site’ to interpret the regional data set by comparing coeval samples and checking the main floristic differences among the chronological phases. Moreover, though their value of local palaeoecological indicators is well known, in the elaborations, NPPs were also treated as ‘regional proxies’ in order to compare the different sites.
3. Results and Discussion
The pollen dataset shows evidence of the diachronic evolution of the Basilicata cultural landscape over about 2500 years of (mainly pastoral) human activities. Below, after a description of the state of preservation and concentration of pollen and other palynomorphs, the main results on the vegetation cover and land-uses in the region are briefly reported; greater emphasis has been placed on the signs of pastoral/breeding practices in the landscape, which are treated in different subsections. Data are presented according to the main chronological/cultural phases at a regional scale; the principal palynological results site-by-site are summarised in Table 2.
3.1. Pollen Preservation and Concentration of Palynomorphs
Pollen was found in almost all samples in a generally good state of preservation, although many grains were crumpled, broken, or rearranged due to the physical–chemical and post-depositional disturbance processes that are fairly common in the archaeological deposits [89,90,91,92]. In addition, some pollen grains were thinned and pale, likely as a consequence of their passage through the digestive tract before their inclusion in the deposits. Several pollen clumps—small single, or mixed-type pollen clusters (Figure 2)—were observed at three sites (1, 5, and 7). These clumps could be interpreted as the remains of anthers (flowers standing in/transported to the sites) or as fecal pellets dropped by arthropods and herbivorous mammals [88,93,94,95]. Interestingly, no pollen clumps have been observed in the modern soil samples.
The pollen and the NPP concentrations were quite high in most of the archaeological samples (overall mean concentrations: about 11 × 103 p/g and 20 × 103 npp/g). The modern samples showed a notable pollen concentration (26 × 103 p/g), while NPPs were less recurring in the slides (7 × 103 npp/g).
Pollen spectra were characterized by a high number of taxa, mainly belonging to herbs (mostly Cichorieae and Poaceae wild grass group), with floristic lists that include between 31 and 152 taxa per sample. The good mean pollen concentration and the high taxa diversity suggest that any selective deterioration affected pollen grains in the deposits [62]. Herbs also prevailed in the modern samples (again, Cichorieae and Poaceae were the most recurring taxa); their floristic lists (from 52 to 71 taxa) suggest a less-diversified vegetation composition as compared to the past.
3.2. Vegetation Cover: Wood Composition, Mediterranean Habitats, and Wet Environments
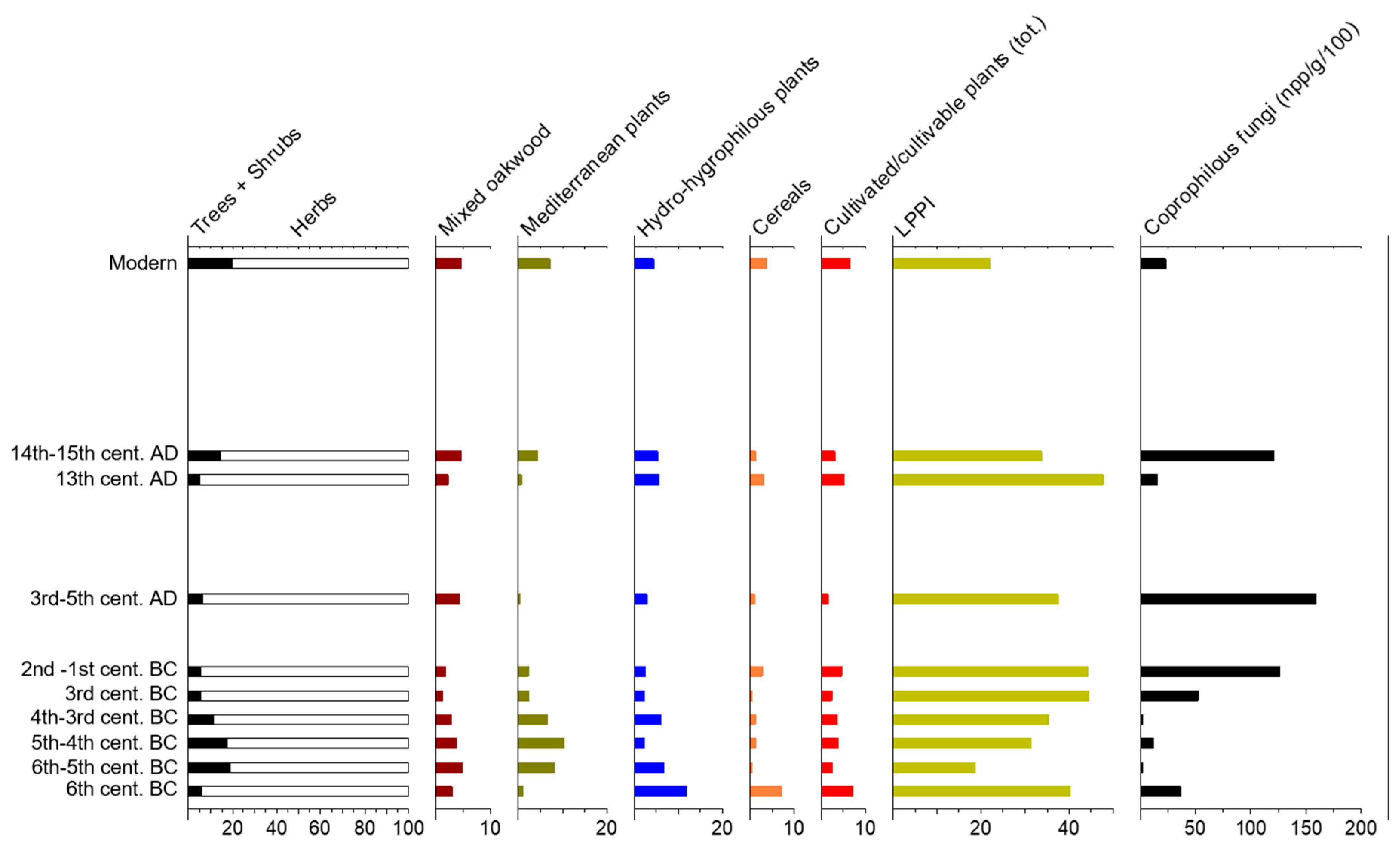
Woody plants were poorly represented in the pollen spectra (the mean percentage ratio of woodland/herbaceous plants is 11/89), and the tree cover, in particular, was very low. The forest cover was <20% at all sites and over the whole considered period (from the 6th century BC to the present; Figure 3). The woods were represented by both Mediterranean trees and shrubs (4% on average in the archaeological samples) and mixed oakwoods (3%). Fagaceae (deciduous Quercus, 1.9%) and Pinaceae (1.8%) prevailed in the forest cover, followed by Olea (1.3%), Daphne (0.9%), and Quercus ilex type (0.6%), which were the main elements of the Mediterranean vegetation.
The striking trait of the vegetation cover corresponded to a Mediterranean shrubland that was widespread during the Hellenistic period (>5% on average; up to 10.2% in the 5th–4th century BC), while its minimum values (0.2%) were observed during the 3rd–5th century AD, when the lowest values of the mixed oakwoods (1.2%) were recorded (Figure 3). In particular, Mediterranean plants recurred in the pollen spectra from the Greek rural sites 5 (9%) and 7 (5.7%) (Table 2) and were also significant in the Medieval samples from site 2 (7.1%) (Figure 3). Although the spread of sclerophyllous shrubs is linked to dryer environmental conditions, there was no local evidence of a concurrent significant reduction in wet environments (see below). This suggests that this was mostly a degraded environment rather than natural vegetation. In fact, the Mediterranean plants (Olea may have been cultivated, and the shrubs developed under continued grazing pressure) may have been the result of several human activities that were practised in the area for a long time.
In the surface sediment samples, the oakwoods and Mediterranean vegetation were well-represented. The modern pollen spectra indicate a prevalence of pines and Mediterranean shrubs in the current wood composition. The increase of sclerophyllous shrubs in the modern samples compared to the post-Hellenistic ones may be reasonably explained by a less-diversified land-use rather than by a true environmental change. Likewise, the extension of pinewoods is due to the modern afforestation of the abandoned farmlands in the region.
In past pollen spectra, wet environments were represented by Alnus, Populus, Salix (0.2%) and to a lesser extent Populus (0.1%), among trees, and by Cyperaceae (2.7%), Ranunculus cf. macrophyllus (0.8%), Typha types (0.3%) and Nymphaea alba type (0.2%), accompanied by other plants of wetlands in traces (e.g., Lemna, Potamogeton, Hydrocharis, <0.1%). These pollen grains are ubiquitous, and their highest mean value dated to the first Hellenistic phases (11.9%). All Roman samples have pollen from wet environments that never exceeds 3%, and then rises up to 5.6% in the Medieval Ages (Figure 3). Aquatic and hygrophilous plants matched the presence of the HdV-181, Zygnemataceae and Pseudoschizaea algae, suggesting that there were some small springs or stagnant shallow water pools not far from the sites [83,96,97,98].
In modern samples, pollen from plants living near or in the water showed fairly similar percentages (4.5% on average), while there are few algae. This is evidence of the occurrence of a low number of wet environments in the area.
Overall, the past pollen spectra describe open areas with scanty woodlands, shrubby grasslands, and a very local presence of wet environments. These data agree with the assumption that the area was largely treeless, with few variations in the wood composition. The Mediterranean elements that recur in the spectra could reflect the impact of pastoralism on the landscape.
Today, as in the past, the thin forest cover has a strong Mediterranean imprint, and the wet environments are similar to those that were present in the area after the Greek period. The ensemble of this pollen evidence, together with that from human environments (the following paragraphs), indicates that the natural environment of the region has always had a strong ‘contamination imprint’ by a human presence.
3.3. Crops and Fields
Cultivated plants were mainly represented by Olea (1.1%; up to 2.8% in the 5th–4th century BC samples from the sites 5, 6, and 7) and Juglans (0.2%) and traces of Castanea and Vitis (<0.1%) among woody plants. Unlike the other mentioned taxa, Olea was well-represented in all samples, and could reasonably have resulted from a process of cultivation instead of from the wild subspecies living in the area. In fact, Olea pollen is known to have been spreading since about 3600 cal. years BP in southern Italy, when a strong human impact was recorded (e.g., [99,100]). Moreover, olive tree cultivation has been well-attested to at the sites of Basilicata, where evidence of plant processing has also been found (e.g., olive presses at site 4) [62]. Other trees and shrubs that produce edible fruits that may be evidence of possibly cultivated plants were Corylus (0.3%), Ficus (0.1%), and Prunus (0.03%). Their low total amount (4% on average) suggests that they were grown/cultivated fairly far away from the sites.
Cereals were represented by the oat/wheat type (1.8%; up to 6.4% in the 6th century BC samples from site 1), barley type (0.3%), and traces of rye (0.04%). Therefore, wheat, rather than other cereals, were mainly cultivated in the fields in all phases. Weeds, such as Papaveraceae (0.1%), Anagallis, Aphanes type, Cerastium, Convolvulus, Mercurialis, Polygonum, and Solanum (total: <0.2%), are other indicators of crop practices [101]. Overall, the low amount of pollen from fields suggests that these cultivations were carried out quite far away from the sites (with the exception of site 1, where the cereal farming should have been very well-developed; Table 2). Other cultivated herbs may have been Fabaceae (3.4%, including Astragalus, Lotus, Medicago, and Trifolium type, legumes possibly cultivated for fodder) and some Apiaceae (0.6%, including aromatic species). Interestingly, Cannabis was attested to at site 7 (only in 2 samples dated 3rd century BC, 0.2% each), suggesting that some hemp was grown on the Metaponto plain during the Hellenistic period.
Agricultural activities were also well-attested to in the modern spectra (6.6%) by orchard crops, cereal fields, and olive groves. The cultivated woody plants (mainly olives) had values comparable to those obtained from the archaeological samples, while slightly higher percentages of cereals were observed in the current surface soils (3.9%). This suggests a continuity of the agricultural practices in the area, probably linked to and in line with the traditional land-uses of the territory, and despite the introduction of mechanized farming in recent years.
3.4. Grazing and Pasturelands
3.4.1. Pollen Pasture Indicators
All of the archaeological samples were characterized by plants and other evidence from grazing environments (Figure 4). Pollen spectra were dominated by Cichorieae (26.5%) and Poaceae wild grass group (18.7%), together with other Asteraceae (8.4%), Chenopodiaceae (5.5%), and Brassicaceae (4.5%). All of these taxa are indicative of dry pastures and are commonly included in the pollen assemblage that is employed for detecting the impact of pastoralism on the landscape (Figure 5) [44,45]. In particular, Cichorieae was recognized as one of the main pasture pollen indicators reflecting animal breeding and pasturelands [66]. The controversial significance of the overrepresentation of Cichorieae in pollen records from the Mediterranean context (as an effect of selective pollen deterioration in the deposits or due to the selection by animal browsing?) had been resolved in the studied sites through an integrated approach comparing the main results from palaeoecological and ecological studies that were carried out in the region. The study showed that Cichorieae prevail in secondary pastures and some types of primary open habitats; the recovery of high percentages of this pollen is, therefore, a good indicator of these habitats even in past environments [66] (p. 163).
Besides Cichorieae, the recovery of high percentages of Poaceae wild grass group—strongly correlated with open and grazed areas [44]—is further evidence of open habitats characterized by dry pastures. These xeric environments are, in fact, favored by the overexploitation of thinned plant resources, including overgrazing [62,102].
Cichorieae had prevalent values in the Roman (32%) and Medieval phases (31%; up to 41% in the 13th century samples). In the Hellenistic samples (23%), there was a steady increase in Cichorieae percentages from the 5th (9%) to the 2nd–1st century BC (34%) after a first peak (24%) in the 6th century BC (Figure 4). Similarly, the most significant values of Poaceae were found in the Roman (31%) and Medieval samples (18%), with high percentages also in the Greek phases (17%). These pollen data, along with the constant presence of Asteraceae (e.g., Carduus, Centaurea nigra type, Cirsium), Chenopodiaceae and Brassicaceae, indicate that dry grasslands/pastures might have been well-extended, probably with seasonal oscillations, in the past landscape.
Constantly recorded in the spectra were other herbs linked to grazing/breeding activities, such as Plantago (2.5%, recurring in trampled areas) [103,104] and Fabaceae (3.4%, palatable herbs present in meadow-pastures or used for fodder). Plantain had its maximum values in the Roman samples (5.6%) when legumes were less represented (0.8%). This probably means that animals grazed in open pastures without any additional feed, maybe due to the mild climate of this period. Fabaceae and Plantago instead had quite comparable values in almost all of the other studied phases (Figure 4).
According to Mazier [45,85], the LPPI—Local Pastoral Pollen Indicators is a sum that is helpful when investigating the response of plants to browsing and the local presence of animal breeding or pastoral activities in a territory. In the archaeological samples, the sum of LPPI had mostly high percentages (37%, up to 48% in the samples dated 13th century AD), suggesting that there were widespread breeding activities in the sites’ surroundings. In particular, this sum reached its highest values in the samples from sites 8 and 6 (c. 48%; Table 2); due to their peculiar archaeological contexts (a dump and a drainage channel, respectively), these samples record and provide detailed information on the human activities in the area.
Pollen pasture indicators were similarly represented in the current pollen spectra. Poaceae was the most-recurring pollen taxa (18%), followed by Cichorieae (11%) and Brassicaceae (5.5%). This seems to be in line with the reduction in the grazed areas in the region at present. Conversely, the good values of Fabaceae (3.8%) suggest the use of such plants as fodder (cultivated and then grazed or stored).
The amount of LPPI plants was decidedly lower in the modern surface soils than in the archaeological samples (22% vs. 37%; Figure 4), indicating that there were more breeding activities in the past than today.
3.4.2. Non-Pollen Palynomorphs as Grazing Markers
The dung-loving fungi are powerful environmental proxies used to assess the presence of fauna in the past, in particular, herbivores [38,105,106,107,108]. In the life-cycle of saprobic fungi growing on animal dung, the spores of coprophilous ascomycetes are accidentally eaten by herbivores, survive passage through the intestinal tract, germinate, and then grow in the deposited dung. The fungi then release their spores, which are deposited on vegetation, that is then consumed by grazing animals to complete the cycle. Therefore, the coprophilous fungi are indices of the presence of dung and are associated with pastoral/breeding activities.
The main dung-loving fungi are Sordaria (HdV-55a), Apiosordaria verruculosa (169), Sporormiella (HdV-113), Cercophora (HdV-112), Delitschia, and Podospora (HdV-368) types (Figure 5). As shown by recent studies on modern analogues in grazed areas [44,47,105,106], Sporormiella (including obligate coprophilous species) and Sordaria (a non-strictly coprophilous genus) can be considered the most reliable indicators of the local presence of herbivores.
Altogether, these proxies of pastoral activities were found in the archaeological samples in variable concentrations (Figure 4). Coprophilous spores (mostly Sordaria and Sporormiella types) recurred in the Late Hellenistic, Roman, and Medieval samples (from c. 12,000 up to c. 16,000 npp/g) when the highest LPPI values were observed. The significant values of LPPI, together with the coprophilous fungal spores, strongly suggest that pastoral/breeding activities were widely practiced in the study area.
The largest amount of coprophilous fungi was observed at sites 2 and 4 (c. 17,600 npp/g and 9000 npp/g; Table 2) in samples that were mainly from layers within natural or artificial enclosures where animals were housed or moved to graze. Interestingly, in those samples, several spores of Diporotheca rhizophila have also been found (c. 2000 npp/g as the average of the two sites; up to c. 9000 npp/g in the 3rd century BC samples from the site 4). The genus D. rhizophila is a root parasite that grows in meso- to eutrophic conditions and wet habitats with extensive soil erosion due to the impact of agricultural activities and livestock trampling [109]. The spores of this fungus may generally be a valuable soil disturbance indicator for pastoral activities. In particular, in highly trampled or overgrazed areas, the soil is left barren and exposed to erosion with the dispersion of Diporotheca spores. The combined evidence of D. rhizophila and coprophilous fungal spores suggests that these enclosed areas (probably with the water sources needed to supply livestock) had been intensively grazed.
Sporormiella and Cercophora types reached their highest values at site 2 (c. 13,000 npp/g and 8300 npp/g), where Sordaria was also quite well-attested to (c. 3500 npp/g). Delitschia and Podospora were the lowest represented morphotypes in all the archaeological samples (always <200 npp/g).
Other microfossils that are related to the presence of animals at the sites are eggs of intestinal parasites, such as Dicrocoelium, which is a parasite of ruminants [77,110]. Parasitological remains were quite rare in the samples (few eggs of Dicrocoelium were found in late Hellenistic and Roman phases at the sites 2 and 7) but are further evidence that some animal excrements became incorporated into these deposits.
Overall, the NPP record evinces the continual presence of animals in the past landscapes, supporting the evidence of pastures and pastoral activities in the region.
In the modern samples, the NPP indicators of grazing pressure were less-recorded (Figure 4). The coprophilous fungal spore concentration was decidedly lower (c. 2500 npp/g) than that found in the archaeological layers. Nevertheless, the morphotypes observed were the same and with a proportionately similar presence: mostly Sordaria, Sordariaceae, and Sporormiella (c. 1600 npp/g in total), and few Cercophora (c. 150 npp/g), Delitschia and Podospora types (<100 npp/g each). No intestinal parasite eggs were found. Altogether, these microfossils data indicate that animals frequent the area less as compared to the past, probably as an effect of the reduction in pastoral practices in the region at present.
3.5. The Relevance of Past Pastoral Activities and Their Legacy on Today’s Landscape
The integrated analyses of microscopic records from the studied archaeological sites indicate wide and continuous pastoral activities practiced in the region. In particular, the combined evidence from pollen pasture indicators and NPP markers of grazing (mainly coprophilous fungal spores) point out that pastures were the main type of land-use in the territory surrounding each of the eight study sites. As evidenced by the high values of the LPPI sums in the past pollen records, this region has long been a grazed area, with more intense pastoral activities documented from the end of the Hellenistic age to the Medieval and Renaissance periods; besides this, land used for animal feed was also attested to by fodder plants pollen.
The interpretation of the archaeological pollen data was actually based on a comparison of several contexts, chronologies, geomorphologies, and vegetation belts, offering different environmental contexts from the sites located on mountains and hills to those placed on the coastal plain. Interestingly, the pollen spectra suggest that similar agrarian systems exist along this transect, mainly with pastoral activities and cereal and olive cultivations. According to the palynological dataset, during ancient Greek times, crop and tree cultivation was the prevalent activity in the chora of Metaponto, and agriculture was performed close to the settlements along with pastoral/herding practices. The same land-use had continued throughout the late Hellenistic/early-Roman phases up to the Medieval period, when pastoral farming was the predominant form of land-use, especially in the inland areas. Here, the determining factor of such a type of farming was probably the conformation of the land, since ploughing is not practicable on hilly slopes. The geomorphology may have also influenced the types of livestock that were used in the territory, where—in accordance with the zooarchaeological record [64]—sheep and goats rather than cattle were reared and grazed. Moreover, the widespread shrubby vegetation that was documented in the pollen record is further evidence that the prevalent domestic animals may have been ovicaprines, as most of the Mediterranean shrub species contain secondary metabolites such as tannins, terpenes, and volatile oils that are non-edible for many other herbivores [111].
Conversely, in the modern spectra, pollen and NPP indicators of grazing are not as common as in previous times. The current landscape is characterized by intense agricultural activities, including orchard crops, cereal fields, and olive groves, and by shrubby grasslands and a well-developed Mediterranean macchia. The modern development of Mediterranean vegetation may also reasonably be a response to overgrazing in the past [12,13]. Grazing is regarded as one of the main actions responsible for deforestation as well as the major factor preventing the evolution of macchia to forests [14,16]. Overgrazing or continual grazing pressure by sheep and goats has changed most of the once-forested areas of the Mediterranean into the maquis shrublands that typically occur in the current landscapes. This environmental change has been documented in most of the reference biostratigraphical records from the central Mediterranean. For example, the Holocene pollen record from Lago Grande di Monticchio in Basilicata revealed a forest cover decrease from c. 3000 cal. years BP to the present, and a concurrent increase in the relative abundance of Pistacia and other Mediterranean plants, Olea, Juglans, Castanea (OJC group, evidence of human-induced environments) [6], Poaceae wild grass group, Artemisia, and Plantago (PAZ 1b-1a) [112]. Similarly, a sharp reduction in forested areas, and an increase in synanthropic plants and Mediterranean vegetation was systematically documented in the mid—late Holocene sections of off-site cores from southern Italy [52,53,54,55,99,100,113,114,115,116]. Overall, these biostratigraphical records suggest that a human impact on vegetation has occurred since about 3500 cal. years BP. Therefore, it is not surprising that the natural environment had been shaped by the human presence in the study area since ancient times, as shown by the very low tree cover that was expressed in the 6th century BC samples. In addition, the pollen sums indicate that intense pastoral activities and broad agrarian practices have highly influenced the landscape surrounding the sites. In the modern pollen spectra, the low forest cover is thereby the result of millennia of human presence and activities in the region. The prevailing Mediterranean vegetation can be considered to be a degraded environment related to the long-term impact of livestock grazing.
The current plant biodiversity of the region mainly depends on past human activities, closely intertwined with environmental and climatic factors. The microfossil records suggest that continuous livestock activity had greatly contributed to a shift in the floristic composition towards the current plant biodiversity and a patchwork of habitats, including open areas. Therefore, the conservation and sustainable use of this biodiversity can be achieved mainly by recognizing the important role of pastoral practices in regional cultures, traditions, and livelihoods.
This study can support biodiversity awareness by providing useful information on the vocation and environmental sustainability of the Basilicata region. For example, the current environmental resource management strategies intend to replace the abandoned grazing areas with woodlands, as well as introduce new cost-effective crops in a major renovation and development of the territory. These actions may carry significant risks of biodiversity alteration in the context of unsustainable development. Palynological data from the regional archaeological sites provide insights into anthropogenically-driven landscape dynamics and inform us about a landscape adaptation and biodiversity changes over time. In addition, the long-term perspective of this research provides a knowledge base on the effects of many different forms of farming (cultivations or grazing) on local biodiversity and also testifies to the impact of agricultural intensification or abandonment on the ecosystem, which the region has already experienced in the past. Understanding the responses of ecosystems to anthropogenic disturbance and the long-term scale of these processes are essential points to the development of sustainable management strategies that can help to reduce biodiversity loss.
4. Conclusions
The palynological dataset from eight archaeological sites in Basilicata (southern Italy) is relevant to understanding the farming activities that have been practised in the area over the last 2500 years. The data point to an open plant landscape that has been continuously dominated by pastures, cereal fields, and olive groves. Important evidence of pastoral farming has arisen from the joint record of pollen pasture indicators and NPP markers of grazing (mainly spores of coprophilous fungi). The combined evidence from these microfossils points out that the region’s environmental setting has been greatly influenced by intense pastoral/breeding activities since ancient times.
Pastoral farming was the predominant activity from the late Hellenistic period to the Middle Ages/Renaissance, especially in the inland areas, where the geomorphological features may have favoured the breeding of ovicaprines. The high values of both LPPI sums and coprophilous fungal spores suggest that pastures were widespread in the sites’ surroundings. In addition, the high percentages of the Cichorieae and Poaceae wild grass group indicate open habitats characterized by dry pastures. Besides these grazing lands, the good percentages of Fabaceae suggest a combination of pastures and land used for animal feed production. In addition, pollen from sclerophyllous plants (especially shrubs) recurs in all of the spectra. Mediterranean vegetation is commonly viewed as a degraded landscape and could be considered to be a long-term pasture marker. While pollen and NPP indicators of grazing are not common in modern spectra, Mediterranean plants are quite well-attested to; these data suggest a reduction in pastures in current land-uses and support the idea of the importance of past pastoral practices as a major agent of the regional landscape’s transformation. To improve environmental sustainability, we cannot ignore both the landscape evolution and bio-cultural value of a territory. The palaeoecological results from the study sites are useful information, especially for the understanding of the various environmental conservation measures. By knowing the past biodiversity and ecosystem dynamics, future scenarios can be taken into account by stakeholders and decision makers, helping them figure out the consequences of different conservation strategies. In order to better understand the dynamics and processes that govern modern ecosystems and habitats, an integrated discussion of the main results obtained from palaeoecological and ecological studies carried out in a certain region should be pursued. In fact, the insights into long-term events greatly contribute to improving modern concepts of community organization, evaluating climate dynamics, and understanding disturbance processes that cannot be understood by analyzing modern systems alone [117]. In the same way, the study of modern analogues could provide further information on pollen and NPP vegetation and land-use relationships and contribute to a more comprehensive landscape reconstruction (e.g., [118]). Future developments of this research will move in this direction and try to support both modern biodiversity awareness and cultural landscape conservation for sustainable regional resources management and exploitation. Future landscape management, resources exploitation, and restoration planning could benefit from the palaeoecological insights that this research provides, which take into account new factors, including both past and future environmental (climate) and social (history) changes.
Acknowledgments
This research was the PhD project of AF at the Earth System Science Doctorate School of the University of Modena and Reggio Emilia. The main investigation was carried out in collaboration with the Institute of Classical Archaeology of the University of Texas at Austin within the ‘Metaponto project’ (director: Prof. Joseph Coleman Carter), a vast integrated study of Greek colonies and rural populations of Basilicata. The indigenous sites were investigated in the framework of the multidisciplinary research lead by the Postgraduate School in Archaeological Heritage at Matera of the University of Basilicata (scientific officers: Profs. Massimo Osanna and Dimitris Roubis). The author is grateful to the anonymous reviewers for their constructive comments.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kareiva, P.; Watts, S.; McDonald, R.; Boucher, T. Domesticated Nature: Shaping Landscapes and Ecosystems for Human Welfare. Science 2007, 316, 1866–1869. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, P.M.; Mooney, H.A.; Lubchenco, J.; Melillo, J.M. Human domination of earth’s ecosystems. Science 1997, 277, 494–499. [Google Scholar] [CrossRef]
- Butzer, K.W. Environmental history in the Mediterranean world: Cross-disciplinary investigation of cause-and-effect for degradation and soil erosion. J. Archaeol. Sci. 2005, 32, 1773–1800. [Google Scholar] [CrossRef]
- Mercuri, A.M. Genesis and evolution of the cultural landscape in central Mediterranean: The ‘where, when and how’ through the palynological approach. Landsc. Ecol. 2014, 29, 1799–1810. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Sadori, L.; Blasi, C. Editorial: Archaeobotany for cultural landscape and human impact reconstructions. Plant Biosyst. 2010, 144, 860–864. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E. Olea, Juglans and Castanea: The OJC group as pollen evidence of the development of human-induced environments in the Italian peninsula. Quat. Int. 2013, 303, 24–42. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Sadori, L.; Uzquiano, P. Mediterranean and North-African cultural adaptations to mid-Holocene environmental and climatic changes. Holocene 2011, 21, 189–206. [Google Scholar] [CrossRef]
- Kouli, K.; Masi, A.; Mercuri, A.M.; Florenzano, A.; Sadori, L. Regional vegetation histories: An overview of the pollen evidence from the Central Mediterranean. Late Antiq. Archaeol. 2015, 11, 69–82. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Florenzano, A.; Burjachs, F.; Giardini, M.; Kouli, K.; Masi, A.; Picornell-Gelabert, L.; Revelles, J.; Sadori, L.; Servera-Vives, G.; et al. From influence to impact: The multifunctional land-use in Mediterranean prehistory emerging from palynology of archaeological sites (8.0-2.8 ka BP). Holocene. in press.
- Naveh, Z.; Whittaker, R.H. Structural and floristic diversity of shrublands and woodlands in northern Israel and other Mediterranean areas. Vegetatio 1979, 41, 171–190. [Google Scholar] [CrossRef]
- Davis, F.W.; Goetz, S. Modelling vegetation pattern using digital terrain data. Landsc. Ecol. 1990, 4, 69–80. [Google Scholar] [CrossRef]
- Carmel, Y.; Kadmon, R. Effects of grazing and topography on long-term vegetation changes in a Mediterranean ecosystem in Israel. Plant Ecol. 1999, 145, 243–254. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J. Biology and Wildlife of the Mediterranean Region; Oxford University Press: Oxford, UK, 1999; ISBN 9780198500360. [Google Scholar]
- Joffre, R.J.; Vacher, C.; Long, G. The dehesa: An agrosilvopastoral system on the Mediterranean region with special reference to the Sirera Morena area of Spain. Agrofor. Syst. 1988, 6, 71–96. [Google Scholar] [CrossRef]
- Callaway, R.M.; Davis, F.W. Vegetation dynamics, fire, and the physical environment in coastal central California. Ecology 1993, 74, 1567–1578. [Google Scholar] [CrossRef]
- Seligman, N.G.; Perevolotsky, A. Has intensive grazing by domestic livestock degraded Mediterranean Basin rangelands? In Plant-Animal interactions in Mediterranean-Type Ecosystems; Arianoutsou-Faraggitaki, M., Groves, R.H., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 93–103. ISBN 9789401109086. [Google Scholar]
- Amiri, F.; Ariapour, A.; Fadai, S. Effects of livestock grazing on vegetation composition and soil moisture properties in grazed and non-grazed range site. J. Biol. Sci. 2008, 8, 1289–1297. [Google Scholar] [CrossRef]
- Díaz, S.; Lavorel, S.; McIntyre, S.; Falczuk, V.; Casanoves, F.; Milchunas, D.G.; Skarpe, C.; Rusch, G.; Sternberg, M.; Noy-Meyr, I.; et al. Plant trait responses to grazing—A global synthesis. Glob. Chang. Biol. 2007, 13, 313–341. [Google Scholar] [CrossRef]
- Georgiadis, N.J.; Ruess, R.W.; McNaughton, S.J.; Western, D. Ecological conditions that determine when grazing stimulates grass production. Oecologia 1989, 81, 316–322. [Google Scholar] [CrossRef]
- Feurdean, A.; Ruprecht, E.; Molnár, Z.; Hutchinson, S.M.; Hickler, T. Biodiversity-rich European grasslands: Ancient, forgotten ecosystems. Biol. Conserv. 2018, 228, 224–232. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bond, W.J. Humboldt and the reinvention of nature. J. Ecol. 2018. [Google Scholar] [CrossRef]
- Montalvo, J.; Casado, M.A.; Levassor, C.; Pineda, F.D. Species diversity patterns in Mediterranean grasslands. J. Veg. Sci. 1993, 4, 213–222. [Google Scholar] [CrossRef]
- Biondini, M.E.; Manske, L. Grazing frequency and ecosystem processes in a northern mixed prairie, USA. Ecol. Appl. 1996, 6, 239–256. [Google Scholar] [CrossRef]
- Pickup, G. Estimating the effects of land degradation and rainfall variation on productivity in rangelands: An approach using remote sensing and models of grazing and herbage dynamics. J. Appl. Ecol. 1996, 33, 819–832. [Google Scholar] [CrossRef]
- Han, G.; Hao, X.; Zhao, M.; Wang, M.; Ellert, B.H.; Willms, W.; Wang, M. Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 125, 21–32. [Google Scholar] [CrossRef]
- Birks, H.J.B. Contributions of Quaternary palaeoecology to nature conservation. J. Veg. Sci. 1996, 7, 89–98. [Google Scholar] [CrossRef]
- Willis, K.J.; Birks, H.J.B. What Is Natural? The Need for a Long-Term Perspective in Biodiversity Conservation. Science 2006, 314, 1261–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, K.J.; Araújo, M.B.; Bennett, K.D.; Figueroa-Rangel, B.; Froyd, C.A.; Myers, N. How can a knowledge of the past help to conserve the future? Biodiversity conservation and the relevance of long-term ecological studies. Philos. Trans. R. Soc. B 2007, 362, 175–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacPherson, J. Applying Palaeoecology to Conservation: A long-term perspective for informed management of a fynbos nature reserve. Plymouth Stud. Sci. 2009, 2, 218–269. [Google Scholar]
- Barnosky, A.D.; Hadly, E.A.; Gonzalez, P.; Head, J.; Polly, P.D.; Lawing, A.M.; Eronen, J.T.; Ackerly, D.D.; Alex, K.; Biber, E.; et al. Merging paleobiology with conservation biology to guide the future of terrestrial ecosystems. Science 2017, 355, eaah4787. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Marignani, M.; Sadori, L. Palaeoecology and long-term human impact in plant biology. Plant Biosyst. 2015, 149, 136–143. [Google Scholar] [CrossRef]
- Beneš, J.; Mercuri, A.M. CEA 2018: The 14th Conference of Environmental Archaeology in Modena and the special issue of IANSA. IANSA. in press.
- Galop, D.; Rius, D.; Cugny, C.; Mazier, F. A history of long-term human–environment interactions in the French Pyrenees inferred from the pollen data. In Continuity and Change Cultural Adaptation to Mountain Environments Studies in Human Ecology and Adaptation; Lozny, L.R., Ed.; Springer Science + Business Media: New York, NY, USA, 2013; Volume 7, pp. 19–30. ISBN 9781461457022. [Google Scholar]
- Ejarque, A.; Julià, R.; Riera, S.; Palet, J.M.; Orengo, H.A.; Miras, Y.; Gascón, C. Tracing the history of highland human management in the Eastern Pre-Pyrenees: An interdisciplinary palaeoenvironmental study at the Pradell fen, Spain. Holocene 2009, 19, 1241–1255. [Google Scholar] [CrossRef]
- Gauthier, E.; Bichet, V.; Massa, C.; Petit, C.; Vannière, B.; Richard, H. Pollen and non-pollen palynomorph evidence of medieval farming activities in southwestern Greenland. Veg. Hist. Archaeobot. 2010, 19, 427–438. [Google Scholar] [CrossRef]
- Kouli, K. Plant landscape and land use at the Neolithic lake settlement of Dispilió (Macedonia, Northern Greece). Plant Biosyst. 2015, 149, 145–204. [Google Scholar] [CrossRef]
- Bellotti, P.; Calderoni, G.; Dall’Aglio, P.L.; D’Amico, C.; Davoli, L.; Di Bella, L.; D’Orefice, M.; Esu, D.; Ferrari, K.; Bandini Mazzanti, M.; et al. Middle-to late-Holocene environmental changes in the Garigliano delta plain (Central Italy): Which landscape witnessed the development of the Minturnae Roman colony? Holocene 2016, 26, 1457–1471. [Google Scholar] [CrossRef]
- Van Geel, B.; Buurman, J.; Brinkkemper, O.; Schelvis, J.; Aptroot, A.; van Reenen, G.; Hakbijl, T. Environmental reconstruction of a Roman Period settlement site in Uitgeest (The Netherlands), with special reference to coprophilous fungi. J. Archaeol. Sci. 2003, 30, 873–883. [Google Scholar] [CrossRef]
- Florenzano, A.; Mercuri, A.M.; Carter, J.C. Economy and environment of the Greek colonial system in southern Italy: Pollen and NPPs evidence of grazing from the rural site of Fattoria Fabrizio (6th-4th cent. BC; Metaponto, Basilicata). Ann. Bot. 2013, 3, 173–181. [Google Scholar] [CrossRef]
- Bowes, K.; Mercuri, A.M.; Rattigheri, E.; Rinaldi, R.; Arnoldus-Huyzendveld, A.; Ghisleni, M.; Grey, C.; MacKinnon, M.; Vaccaro, E. Peasant Agricultural Strategies in Southern Tuscany: Convertible Agriculture and the Importance of Pasture. In The Economic Integration of Roman Italy: Rural Communities in a Globalising World; de Haas, T.C.A., Tol, G., Eds.; Brill: Leiden, The Netherlands, 2017; Volume 404, pp. 170–199. ISBN 9789004345027. [Google Scholar]
- Cremaschi, M.; Mercuri, A.M.; Torri, P.; Florenzano, A.; Pizzi, C.; Marchesini, M.; Zerboni, A. Climate change versus land management in the Po Plain (Northern Italy) during the Bronze Age: New insights from the VP/VG sequence of the Terramara Santa Rosa di Poviglio. Quat. Sci. Rev. 2016, 136, 153–172. [Google Scholar] [CrossRef]
- Bosi, G.; Labate, D.; Rinaldi, R.; Montecchi, M.C.; Mazzanti, M.; Torri, P.; Riso, F.M.; Mercuri, A.M. A survey of the Late Roman period (3rd-6th century AD): Pollen, NPPs and seeds/fruits for reconstructing environmental and cultural changes after the floods in Northern Italy. Quat. Int. 2018. [Google Scholar] [CrossRef]
- Jouffroy-Bapicot, I.; Vannière, B.; Iglesias, V.; Debret, M.; Delarras, J.-F. 2000 Years of Grazing History and the Making of the Cretan Mountain Landscape, Greece. PLoS ONE 2016, 11, e0156875. [Google Scholar] [CrossRef]
- Ejarque, A.; Miras, Y.; Riera, S. Pollen and non-pollen palynomorph indicators of vegetation and highland grazing activities obtained from modern surface and dung datasets in the eastern Pyrenees. Rev. Palaeobot. Palynol. 2011, 167, 123–139. [Google Scholar] [CrossRef]
- Mazier, F.; Galop, D.; Brun, C.; Buttler, A. Modern pollen assemblages from grazed vegetation in the western Pyrenees, France: A numerical tool for more precise reconstruction of past cultural landscapes. Holocene 2006, 16, 91–103. [Google Scholar] [CrossRef]
- Mazier, F.; Galop, D.; Gaillard, M.J.; Rendu, C.; Cugny, C.; Legaz, A.; Peyron, O.; Buttler, A. Multidisciplinary approach to reconstructing local pastoral activities—An example from the Pyrenean Mountains (Pays Basque). Holocene 2009, 19, 171–188. [Google Scholar] [CrossRef]
- Cugny, C.; Mazier, F.; Galop, D. Modern and fossil non-pollen palynomorphs from the Basque mountains (western Pyrenees, France): The use of coprophilous fungi to reconstruct pastoral activity. Veg. Hist. Archaeobot. 2010, 19, 391–408. [Google Scholar] [CrossRef]
- Ejarque, A.; Miras, Y.; Riera, S.; Palet, J.M.; Orengo, H.A. Testing microregional variability in the Holocene shaping of high mountain cultural landscapes: A palaeoenvironmental case-study in the eastern Pyrenees. J. Archaeol. Sci. 2010, 37, 1468–1479. [Google Scholar] [CrossRef]
- Miras, Y.; Ejarque, A.; Orengo, H.A.; Riera, S.; Palet, J.M.; Poiraud, A. Prehistoric impact on landscape and vegetation at high altitudes: An integrated palaeoecological and archaeological approach in the eastern Pyrenees (Perafita valley, Andorra). Plant Biosyst. 2010, 144, 946–961. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Allevato, E.; Arobba, D.; Bandini Mazzanti, M.; Bosi, G.; Caramiello, R.; Castiglioni, E.; Carra, M.L.; Celant, A.; Costantini, L.; et al. Pollen and macroremains from Holocene archaeological sites: A dataset for the understanding of the bio-cultural diversity of the Italian landscape. Rev. Palaeobot. Palynol. 2015, 218, 250–266. [Google Scholar] [CrossRef] [Green Version]
- Kleine, E.; Woldring, H.; Cappers, R.; Attema, P.; Delvigne, J. Il carotaggio del Lago Forano presso Alessandria del Carretto (Calabria, Italia). Nuovi dati sulla vegetazione olocenica e sulla storia dell’uso del suolo nella Sibaritide interna. In Proceedings of the Preistoria e Protostoria della Calabria: Scavi e Ricerche 2003, Atti delle giornate di studio, Pellaro, Reggio Calabria, Italy, 25–26 October 2003. [Google Scholar]
- Di Rita, F.; Simone, O.; Caldara, M.; Gehrels, W.R.; Magri, D. Holocene environmental changes in coastal Tavoliere Plain (Apulia, southern Italy): A multiproxy approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 139–151. [Google Scholar] [CrossRef]
- Di Rita, F.; Lirer, F.; Bonomo, S.; Cascella, A.; Ferraro, L.; Florindo, F.; Insinga, D.D.; Lurcock, P.C.; Margaritelli, G.; Petrosino, P.; et al. Late Holocene forest dynamics in the Gulf of Gaeta (central Mediterranean) in relation to NAO variability and human impact. Quat. Sci. Rev. 2018, 179, 137–152. [Google Scholar] [CrossRef]
- Joannin, S.; Brugiapaglia, E.; de Beaulieu, J.-L.; Bernardo, L.; Magny, M.; Peyron, O.; Goring, S.; Vannière, B. Pollen-based reconstruction of Holocene vegetation and climate in southern Italy: The case of Lago Trifoglietti. Clim. Past 2012, 8, 1973–1996. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; Massamba N’siala, I. A marine/terrestrial integration for mid-late Holocene vegetation history and the development of the cultural landscape in the Po Valley as a result of human impact and climate change. Veg. Hist. Archaeobot. 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Maggi, R.; Nisbet, R.; Barker, G. The Archeology of Pastoralism in Southern Europe; Rivista di Studi Liguri; Istituto Internazionale di Studi Liguri-Museo Bicknell: Bordighera, Italy, 1990; Volume LVI. [Google Scholar]
- Florenzano, A. Evolution of a Mediterranean Landscape as Shown by the Archaeo-Environmental Reconstruction of Lucanian sites. Ph.D. Thesis, Università di Modena e Reggio Emilia, Modena, Italy, 2013. [Google Scholar]
- Florenzano, A.; Mercuri, A.M. Palynology of archaeological sites: The example of economy and human impact of the Metaponto area (6th-1st century BC). Rendiconti Online 2012, 21, 750–752. [Google Scholar]
- Florenzano, A. Archaeobotany at Fattoria Fabrizio. In The Chora of Metaponto 5: A Greek Farmhouse at Ponte Fabrizio; Lanza Catti, E., Swift, K., Carter, J.C., Eds.; University of Texas Press: Austin, TX, USA, 2014; Volume 5, pp. 113–138. ISBN 9780292758643. [Google Scholar]
- Florenzano, A. Archaeobotanical Analysis. In The Chora of Metaponto 6: A Greek Settlement at Sant’Angelo Vecchio; Silvestrelli, F., Edlund-Berry, I.E.M., Eds.; University of Texas Press: Austin, TX, USA, 2016; Volume 6, pp. 159–171. ISBN 9781477309476. [Google Scholar]
- Florenzano, A. Palynological approach to reconstruct cultural landscape evolution: Case studies from South Italy. In Proceedings of the 2018 IEEE International Workshop on Metrology for Archaeology and Cultural Heritage, Cassino, Italy, 22–24 October 2018; ISBN 9781538652756. [Google Scholar]
- Mercuri, A.M.; Florenzano, A.; Massamba N’siala, I.; Olmi, L.; Roubis, D.; Sogliani, F. Pollen from archaeological layers and cultural landscape reconstruction: Case studies from the Bradano Valley (Basilicata, southern Italy). Plant Biosyst. 2010, 144, 888–901. [Google Scholar] [CrossRef]
- Florenzano, A.; Mercuri, A.M. Dal polline nei sedimenti alla ricostruzione del paesaggio e dell’economia di Torre di Satriano. In Segni del Potere: Oggetti di lusso dal Mediterraneo nell’Appennino Lucano di età Arcaica; Osanna, M., Vullo, M., Eds.; Osanna Edizioni: Venosa, Italy, 2013; pp. 163–168. ISBN 9788881674015. [Google Scholar]
- Roubis, D.; Colacino, C.; Fascetti, S.; Pascale, S.; Pastore, V.; Sdao, F.; De Venuto, G.; Florenzano, A.; Mercuri, A.M.; Miola, A.; et al. The archaeology of ancient pastoral sites in the territory of Montescaglioso (4th–1st century BC). An interdisciplinary approach from the Bradano valley (Basilicata- southern Italy). SIRIS 2013, 13, 117–136. [Google Scholar]
- Florenzano, A. La pastorizia nell’economia e nel modellamento del paesaggio mediterraneo. Esempi da siti archeologici del sud Italia. In Storia e Archeologia Globale 2. I Pascoli, i Campi, il Mare. Paesaggi di altura e di Pianura in Italia dall’Età del Bronzo al Medioevo; Cambi, F., De Venuto, G., Goffredo, R., Eds.; Edipuglia: Bari, Italy, 2015; pp. 245–252. ISBN 9788872287750. [Google Scholar]
- Florenzano, A.; Marignani, M.; Rosati, L.; Fascetti, S.; Mercuri, A.M. Are Cichorieae an indicator of open habitats and pastoralism in current and past vegetation studies? Plant Biosyst. 2015, 149, 154–165. [Google Scholar] [CrossRef]
- Florenzano, A.; Mercuri, A.M.; Rinaldi, R.; Rattighieri, E.; Fornaciari, R.; Messora, R.; Arru, L. The representativeness of Olea pollen from olive groves and the Late Holocene landscape reconstruction in central Mediterranean. Front. Earth Sci. 2017, 5, 85. [Google Scholar] [CrossRef]
- Roubis, D. Ricerche archeologiche nell’abitato indigeno di Difesa San Biagio (Montescaglioso). In Proceedings of the Ricerche Sulla Casa in Magna Grecia e in Sicilia, Atti del Colloquio, Lecce, Italy, 22–24 June 1992; D’Andria, F., Mannino, K., Eds.; Congedo Editore: Galatina, Italy, 1996; pp. 235–253, ISBN 8880861506. [Google Scholar]
- Osanna, M.; Roubis, D.; Sogliani, F. Ricerche archeologiche ad Altojanni (Grottole –MT) e nel suo territorio. Rapporto preliminare (2005–2007). SIRIS 2007, 8, 137–156. [Google Scholar]
- Carter, J.C. Living off the Chora: Diet and Nutrition at Metaponto; Institute of Classical Archaeology, The University of Texas at Austin: Austin, TX, USA, 2003; ISBN 0970887957. [Google Scholar]
- Carter, J.C. Discovering the Greek Countryside at Metaponto; The University of Michigan Press: Ann Arbor, MI, USA, 2006; ISBN 9780472114771. [Google Scholar]
- Carter, J.V.; Swift, K. The Chora of Metaponto 7: A Greek Sanctuary at Pantanello; University of Texas Press: Austin, TX, USA, 2018; ISBN 9781477314234. [Google Scholar]
- Osanna, M.; Colangelo, L.; Carollo, G. Lo Spazio del Potere. La Residenza ad Abside, L’anaktoron, L’episcopio a Torre di Satriano; Osanna Edizioni: Venosa, Italy, 2009; ISBN 9788881672790. [Google Scholar]
- Blasi, C. La Vegetazione d’Italia, Carta Delle Serie di Vegetazione, Scala 1:500.000; Palombi & Partner S.r.l.: Roma, Italy, 2010; ISBN 98788606062909. [Google Scholar]
- Di Pietro, R.; Fascetti, S.; Filibeck, G.; Blasi, C. Le serie di vegetazione della Regione Basilicata. In La Vegetazione d’Italia, Carta Delle Serie di Vegetazione, Scala 1:500.000; Blasi, C., Ed.; Palombi & Partner S.r.l.: Roma, Italy, 2010; pp. 375–389. ISBN 98788606062909. [Google Scholar]
- De Capua, E.L.; Nigro, C.; Labriola, F. Boschi, biodiversità, territorio e variazioni ambientali. Interventi e attività della provincia di Matera. Forest 2005, 2, 110–129. [Google Scholar]
- Florenzano, A.; Mercuri, A.M.; Pederzoli, A.; Torri, P.; Bosi, G.; Olmi, L.; Rinaldi, R.; Bandini Mazzanti, M. The significance of intestinal parasite remains in pollen samples from Medieval pits in the Piazza Garibaldi of Parma, Emilia Romagna, Northern Italy. Geoarchaeology 2012, 27, 34–47. [Google Scholar] [CrossRef]
- Stockmarr, J. Tablets with spores used in absolute pollen analysis. Pollen Spores 1971, 13, 614–621. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis; Blackwell: London, UK, 1991; ISBN 9780632021765. [Google Scholar]
- Beug, H.J. Leitfaden der Pollen Bestimmung für Mittleleuropa und Angrenzende Gebeite; Gustav Fisher Verlag: Stuttgart, Germany, 2004; ISBN 9783899370430. [Google Scholar]
- Reille, M. Pollen et Spores d’Europe et d’Afrique du Nord, 2nd ed.; Laboratoire de Botanique Historique et Palynologie: Marseille, France, 1999; ISBN 2950717535. [Google Scholar]
- Van Geel, B. Non-pollen palynomorphs. In Tracking Environmental Change Using Lake Sediments, Vol. 3, Terrestrial, Algal, and Siliceous Indicators; Smol, J.P., Birks, H.J.B., Last, W.M., Eds.; Kluwer: Dordrecht, The Netherlands, 2001; pp. 99–119. ISBN 9781402006814. [Google Scholar]
- Mudie, P.J.; Marret, F.; Rochon, A.; Aksu, A.E. Non-pollen palynomorphs in the Black Sea corridor. Veg. Hist. Archaeobot. 2010, 19, 531–544. [Google Scholar] [CrossRef]
- Miola, A. Tools for Non-Pollen Palynomorphs (NPPs) analysis: A list of Quaternary NPP types and reference literature in English language (1972–2011). Rev. Palaeobot. Palynol. 2012, 186, 142–161. [Google Scholar] [CrossRef]
- Mazier, F. Modélisation de la Relation Entre Pluie Pollinique Actuelle, Végétation et Pratiques Pastorales en Moyenne Montagne (Pyrenées et Jura). Application Pour L’interprétation des Données Polliniques Fossiles. Ph.D. Thesis, U.F.R. des Sciences et Techniques, Université de Franche Comté, Besançon, France, 2006. [Google Scholar]
- Grimm, E.D. TGView; Illinois State Museum, Research and Collection Center: Springfield, MA, USA, 2004. [Google Scholar]
- Mercuri, A.M. Human influence, plant landscape evolution and climate inferences from the archaeobotanical records of the Wadi Teshuinat area (Libyan Sahara). J. Arid Environ. 2008, 72, 1950–1967. [Google Scholar] [CrossRef]
- Mercuri, A.M. Plant exploitation and ethnopalynological evidence from the Wadi Teshuinat (Tadrart Acacus, Libyan Sahara). J. Archaeol. Sci. 2008, 35, 1619–1642. [Google Scholar] [CrossRef]
- Dimbleby, G.W. The Palynology of Archaeological Sites; Academic Press: London, UK, 1985; ISBN 9780122164804. [Google Scholar]
- Hall, S.A. Deteriorated pollen grains and the interpretation of quaternary pollen diagrams. Rev. Palaeobot. Palynol. 1981, 32, 193–206. [Google Scholar] [CrossRef]
- Horowitz, A. Palynology of Arid Lands; Elsevier: Amsterdam, The Netherlands, 1992; ISBN 9780444882776. [Google Scholar]
- Traverse, A. Sedimentation of Organic Particles; Cambridge University Press: Cambridge, UK, 2005; ISBN 9780521675505. [Google Scholar]
- Davis, O.K.; Anderson, R.S. Pollen in packrat (Neotoma) middens: Pollen transport and the relationship of pollen to vegetation. Palynology 1987, 11, 185–198. [Google Scholar] [CrossRef]
- Davis, O.K.; Buchmann, S.L. Insect sources of pollen clumps in archeological sites in Southwestern U.S.A.: Ground-nesting bees and mites. AASP Contrib. Ser. 1994, 29, 63–74. [Google Scholar]
- Robbins, E.I.; Cuomo, M.C.; Haberyan, K.A.; Mudie, P.J.; Chen, Y.Y.; Head, E. Chapter 27. Fecal pellets. In Palynology: Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Salt Lake City, UT, USA, 1996; Volume 3, pp. 1085–1097. ISBN 9780931871030. [Google Scholar]
- Grenfell, H.R. Probable fossil zygnematacean algal spore genera. Rev. Palaeobot. Palynol. 1995, 84, 201–220. [Google Scholar] [CrossRef]
- Medeanic, S. Freshwater algal palynomorph records from Holocene deposits in the coastal plain of Rio Grande do Sul, Brazil. Rev. Palaeobot. Palynol. 2006, 141, 83–101. [Google Scholar] [CrossRef]
- Miola, A.; Bondesan, A.; Corain, L.; Favaretto, S.; Mozzi, P.; Piovan, S.; Sostizzo, I. Wetlands in the Venetian Po Plain (northeastern Italy) during the Last Glacial Maximum: Interplay between vegetation, hydrology and sedimentary environment. Rev. Palaeobot. Palynol. 2006, 141, 53–81. [Google Scholar] [CrossRef]
- Di Rita, F.; Magri, D. Holocene drought, deforestation, and evergreen vegetation development in the central Mediterranean: A 5,500 year record from Lago Alimini Piccolo, Apulia, southeast Italy. Holocene 2009, 19, 295–306. [Google Scholar] [CrossRef]
- Sadori, L.; Narcisi, B. The postglacial record of environmental history from Lago di Pergusa, Sicily. Holocene 2001, 11, 655–670. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Bandini Mazzanti, M.; Florenzano, A.; Montecchi, M.C.; Rattighieri, E.; Torri, P. Anthropogenic Pollen Indicators (API) from archaeological sites as local evidence of human-induced environments in the Italian peninsula. Ann. Bot. 2013, 3, 143–153. [Google Scholar] [CrossRef]
- Jalut, G.; Dedoubat, J.J.; Fontugne, M.; Otto, T. Holocene circum-Mediterranean vegetation changes: Climate forcing and human impact. Quat. Int. 2009, 200, 4–18. [Google Scholar] [CrossRef]
- Noë, R.; Blom, C.W.P.M. Occurrence of three Plantago species in Coastal dune grasslands in relation to pore-volume and organic matter content of the soil. J. Appl. Ecol. 1981, 19, 177–182. [Google Scholar] [CrossRef]
- Brun, C. Anthropogenic indicators in pollen diagrams in eastern France: A critical review. Veg. Hist. Archaeobot. 2011, 20, 135–142. [Google Scholar] [CrossRef]
- Blackford, J.J.; Innes, J.B. Linking current environments and processes to fungal spore assemblages: Surface NPM data from woodland environments. Rev. Palaeobot. Palynol. 2006, 141, 179–187. [Google Scholar] [CrossRef]
- Graf, M.T.; Chmura, G.L. Development of modern analogues for natural, mowed and grazed grasslands using pollen assemblages and coprophilous fungi. Rev. Palaeobot. Palynol. 2006, 141, 139–149. [Google Scholar] [CrossRef]
- Baker, A.G.; Bhagwat, S.A.; Willis, K.J. Do dung fungal spores make a good proxy for past distribution of large herbivores? Quat. Sci. Rev. 2013, 62, 21–31. [Google Scholar] [CrossRef]
- Davis, O.K.; Shafer, D. Sporormiella fungal spores, a palynological means of detecting herbivore density. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 237, 40–50. [Google Scholar] [CrossRef]
- Hillbrand, M.; Hadorn, P.; Cugny, C.; Hasenfratz, A.; Galop, D.; Haas, J.N. The palaeoecological value of Diporotheca rhizophila ascospores (Diporothecaceae, Ascomycota) found in Holocene sediments from Lake Nussbaumersee, Switzerland. Rev. Palaeobot. Palynol. 2012, 186, 62–68. [Google Scholar] [CrossRef]
- Le Bailly, M.; Bouchet, F. Ancient dicrocoeliosis: Occurrence, distribution and migration. Acta Trop. 2010, 115, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Bartolome, J.; Franch, J.; Plaixats, J.; Seligman, N.G. Diet selection by sheep and goats on Mediterranean heath-woodland range. J. Range Manag. 1998, 51, 383–391. [Google Scholar] [CrossRef]
- Allen, J.R.M.; Watts, W.A.; McGee, E.; Huntley, B. Holocene environmental variability–the record from Lago Grande di Monticchio, Italy. Quat. Int. 2002, 88, 69–80. [Google Scholar] [CrossRef]
- Russo Ermolli, E.; Di Pasquale, G. Vegetation dynamics of south-western Italy in the last 28 kyr inferred from pollen analysis of a Tyrrhenian Sea core. Veg. Hist. Archaeobot. 2002, 11, 211–219. [Google Scholar] [CrossRef]
- Di Rita, F.; Fletcher, W.J.; Aranbarri, J.; Margaritelli, G.; Lirer, F.; Magri, D. Holocene forest dynamics in central and western Mediterranean: Periodicity, spatio-temporal patterns and climate influence. Sci. Rep. 2018, 8, 8929. [Google Scholar] [CrossRef] [PubMed]
- Caroli, I.; Caldara, M. Vegetation history of Lago Battaglia (Eastern Gargano coast, Apulia Italy) during the Middle-Late Holocene. Veg. Hist. Archaeobot. 2007, 16, 317–327. [Google Scholar] [CrossRef]
- Tinner, W.; van Leeuven, J.F.N.; Colombaroli, D.; Vescovi, E.; van der Knaap, W.O.; Henne, P.D.; Pasta, S.; D’Angelo, S.; La Mantia, T. Holocene environmental and climatic changes at Gorgo Basso, a coastal lake in southern Sicily, Italy. Quat. Sci. Rev. 2009, 28, 1498–1510. [Google Scholar] [CrossRef]
- Foster, D.R.; Schoonmaker, P.K.; Pickett, S.T.A. Insights from palaeoecology to community ecology. Trends Ecol. Evol. 1990, 5, 119–122. [Google Scholar] [CrossRef]
- Davis, B.A.S.; Zanon, M.; Collins, P.; Mauri, A.; Bakker, J.; Barboni, D.; Barthelmes, A.; Beaudouin, C.; Bjune, A.E.; Bozilova, E.; et al. The European Modern Pollen Database (EMPD) project. Veg. Hist. Archaeobot. 2013, 22, 521–530. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Study area: (a) Location map of the eight archaeological sites studied for pollen and non-pollen palynomorphs (NPPs) in Basilicata, southern Italy; (b) the current landscapes in which the sites are located.
Figure 1.
Study area: (a) Location map of the eight archaeological sites studied for pollen and non-pollen palynomorphs (NPPs) in Basilicata, southern Italy; (b) the current landscapes in which the sites are located.

Figure 2.
Pollen clumps from the archaeological layers of site 7: (a,b) Single-type pollen clumps; (c) mixed-type cluster, including more than one pollen type. The scale bar is 10 μm.
Figure 2.
Pollen clumps from the archaeological layers of site 7: (a,b) Single-type pollen clumps; (c) mixed-type cluster, including more than one pollen type. The scale bar is 10 μm.

Figure 3.
Percentage pollen diagram (average data from the eight sites, and modern soil samples): Main sums useful for palaeoenvironmental reconstruction, and concentration of coprophilous fungal spores (npp/g/100). The samples are grouped according to their chronology.
Figure 3.
Percentage pollen diagram (average data from the eight sites, and modern soil samples): Main sums useful for palaeoenvironmental reconstruction, and concentration of coprophilous fungal spores (npp/g/100). The samples are grouped according to their chronology.

Figure 4.
Percentage pollen diagram (average data from the eight sites, and modern soil samples): Mediterranean plants, pollen taxa indicators of (or linked to) pastures, and concentration of coprophilous fungal spores (npp/g/100). The samples are grouped according to their chronology.
Figure 4.
Percentage pollen diagram (average data from the eight sites, and modern soil samples): Mediterranean plants, pollen taxa indicators of (or linked to) pastures, and concentration of coprophilous fungal spores (npp/g/100). The samples are grouped according to their chronology.

Figure 5.
Main pollen and non-pollen palynomorphs indicators of grazing environments from the studied archaeological sites: (a) Cichorieae; (b) Aster type; (c) Poaceae wild grass group; (d) Plantago; (e) Centaurea nigra type; (f) Trifolium type; (g) Chenopodiaceae; (h) Brassica type; (i) Sordaria type; (j) Sporormiella type; (k) Delitschia type; (l) Podospora type. The scale bar is 10 μm.
Figure 5.
Main pollen and non-pollen palynomorphs indicators of grazing environments from the studied archaeological sites: (a) Cichorieae; (b) Aster type; (c) Poaceae wild grass group; (d) Plantago; (e) Centaurea nigra type; (f) Trifolium type; (g) Chenopodiaceae; (h) Brassica type; (i) Sordaria type; (j) Sporormiella type; (k) Delitschia type; (l) Podospora type. The scale bar is 10 μm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The studied sites in the Basilicata region, S Italy: Geographical coordinates, chronology, cultural phase, archaeological context, and the number of pollen samples.
Table 1.
The studied sites in the Basilicata region, S Italy: Geographical coordinates, chronology, cultural phase, archaeological context, and the number of pollen samples.
| Site | Geographical Coordinates | Chronology | Cultural Phase | Archaeological Context | Samples |
|---|---|---|---|---|---|
| 1 - Torre di Satriano–TS | 40°34′12.28″N; 15°38′15.26″E 930 m asl | 6th cent. BC | Archaic | indigenous rural settlement | 5 |
| 2 - Altojanni–ALJ | 40°39′73″N; 16°20′57″E 375 m asl | 3rd-5th and 12th-15th cent. AD | Roman; Medieval | fortified rural village; aggers | 26 |
| 3 - Miglionico–MGL | 40°34′03.42″N; 16°29′58.97″E 445 m asl | 14th/15th cent. AD | Medieval | castle | 10 |
| 4 - Difesa San Biagio–DSB | 40°30′21.82″N; 16°40′51.03″E 138 m asl | 5th-1st cent. BC | Hellenistic | indigenous rural settlement | 24 |
| 5 - Fattoria Fabrizio–FF | 40°24′46.58″N; 16°44′28.33″E 57 m asl | 6th-4th cent. BC | Archaic-Hellenistic | Greek farmhouse | 14 |
| 6 - Pizzica–PZZ | 40°24′45.09″N; 16°47′28.14″E 36 m asl | 5th/4th cent. BC | Archaic-Hellenistic | drainage channel in Greek necropolis | 5 |
| 7 - Sant’Angelo Vecchio–SAV | 40°23′39.96″N; 16°43′10.71″E 46 m asl | 6th/5th-1st cent. BC | Archaic-Hellenistic | Greek rural settlement | 29 |
| 8 - Pantanello–PNT | 40°23′21.53″N; 16°47′11.89″E 8 m asl | 2nd-1st cent. BC | Hellenistic | dump of a productive area | 13 |
Table 2.
Summary of pollen and NPP data from each of the eight studied sites.
| Site | Concentr. | AP | Mixed Oakwood | Mediterr. Plants | Hydro-hygroph. Plants | Cereals | Cultiv. Plants (tot.) | LPPI | Coproph. Fungi | Main Features |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 - TS | c. 2100 p/g; c. 7000 npp/g | 6.6% | 2.5% | 1.5% | 11.6% | 6.4% | 6.7% | 38.1% | c. 3500 npp/g | Open environment, with spread wet environments; cereal fields and pastures widespread |
| 2 - ALJ | c. 16,200 p/g; c. 62,200 npp/g | 13.7% | 6.4% | 4.9% | 4.1% | 0.7% | 2.8% | 34.4% | c. 17,600 npp/g | Thin oakwood combined with Mediterranean vegetation; dry grasslands/pastures widespread |
| 3 - MGL | c. 1640 p/g; <100 npp/g | 11% | 1.9% | 1.6% | 5.5% | 1.6% | 2.6% | 35.2% | <50 npp/g | Reduced oakwood and Mediterranean habitats; land-use with cereal fields and pastures |
| 4 - DSB | c. 29.200 p/g; c. 87,000 npp/g | 7.5% | 0.3% | 4.5% | 0.1% | 0.2% | 2.7% | 39% | c. 9000 npp/g | Open vegetation, in which Mediterranean shrubs and dry grasslands/pastures predominate |
| 5 - FF | c. 16.700 p/g; c. 5400 npp/g | 19.4% | 4.8% | 9% | 2.3% | 1.7% | 4.6% | 23.9% | c. 900 npp/g | Thin oakwood and local growing of Mediterranean shrubs; economy based on cereals and pastures |
| 6 - PZZ | c. 1900 p/g; c. 300 npp/g | 11.6% | 0.7% | 10.1% | 2.5% | - | 0.1% | 47.8% | <100 npp/g | Mediterranean habitats widespread; no evidence of local cultivations; dry grasslands/pastures widespread |
| 7 - SAV | c. 5500 p/g; c. 2600 npp/g | 12% | 3.4% | 5.7% | 6.9% | 0.7% | 3.2% | 34.6% | c. 550 npp/g | Mediterranean habitats combined with wet environments; land-use with pastures and reduced cereal fields |
| 8 - PNT | c. 22,000 p/g; <100 npp/g | 3% | 1.5% | 0.8% | 1.3% | 4.5% | 4.9% | 48.2% | <50 npp/g | Open vegetation predominates; dry-grasslands/pastures spread and land-use with cereal fields |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Florenzano, A. The History of Pastoral Activities in S Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness. Sustainability 2019, 11, 404. https://doi.org/10.3390/su11020404
AMA Style
Florenzano A. The History of Pastoral Activities in S Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness. Sustainability. 2019; 11(2):404. https://doi.org/10.3390/su11020404
Chicago/Turabian StyleFlorenzano, Assunta. 2019. "The History of Pastoral Activities in S Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness" Sustainability 11, no. 2: 404. https://doi.org/10.3390/su11020404
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.