Impact of Nitrogen Addition on Physiological, Crop Total Nitrogen, Efficiencies and Agronomic Traits of the Wheat Crop under Rainfed Conditions



Abstract
1. Introduction
2. Materials and Methods
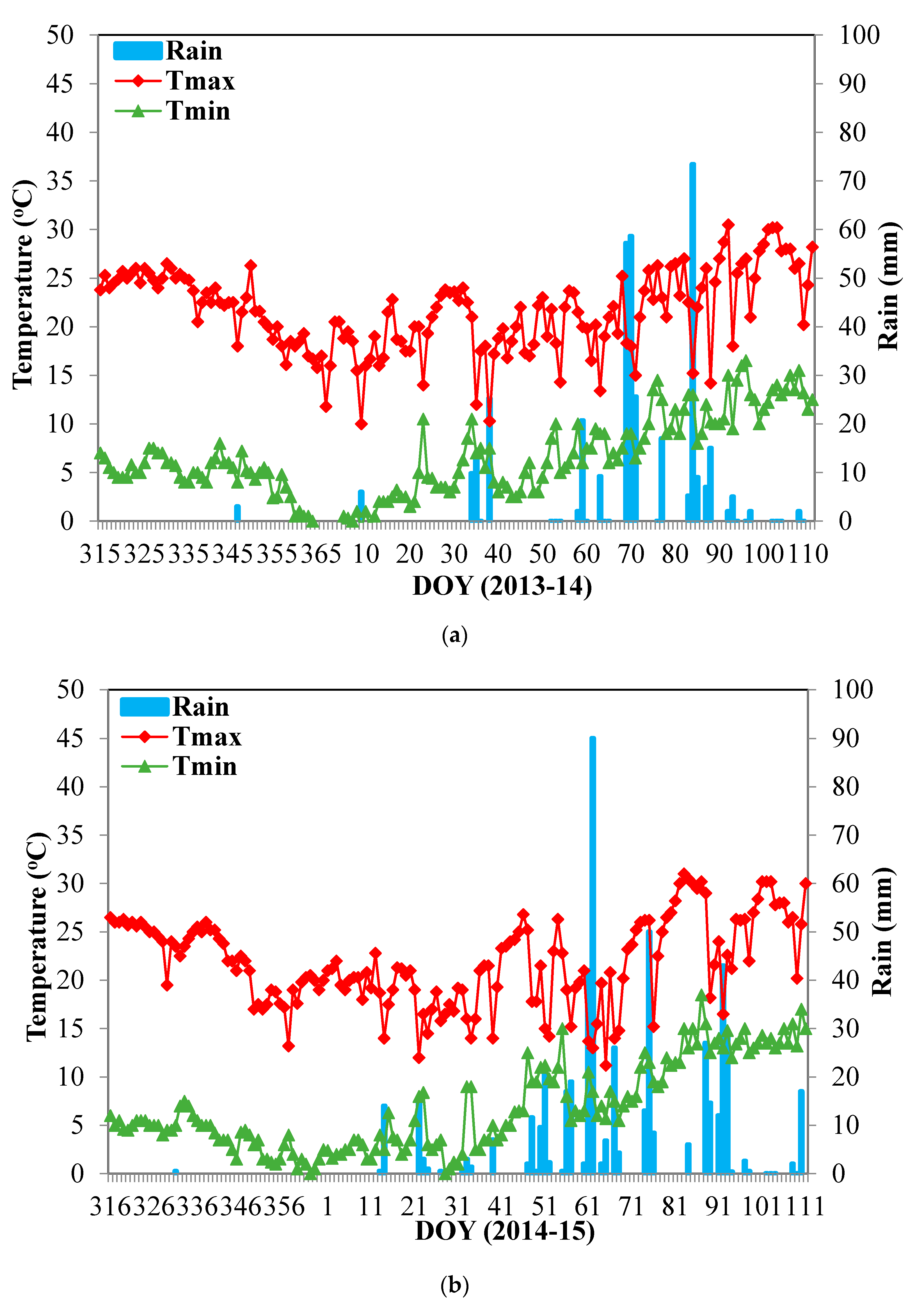
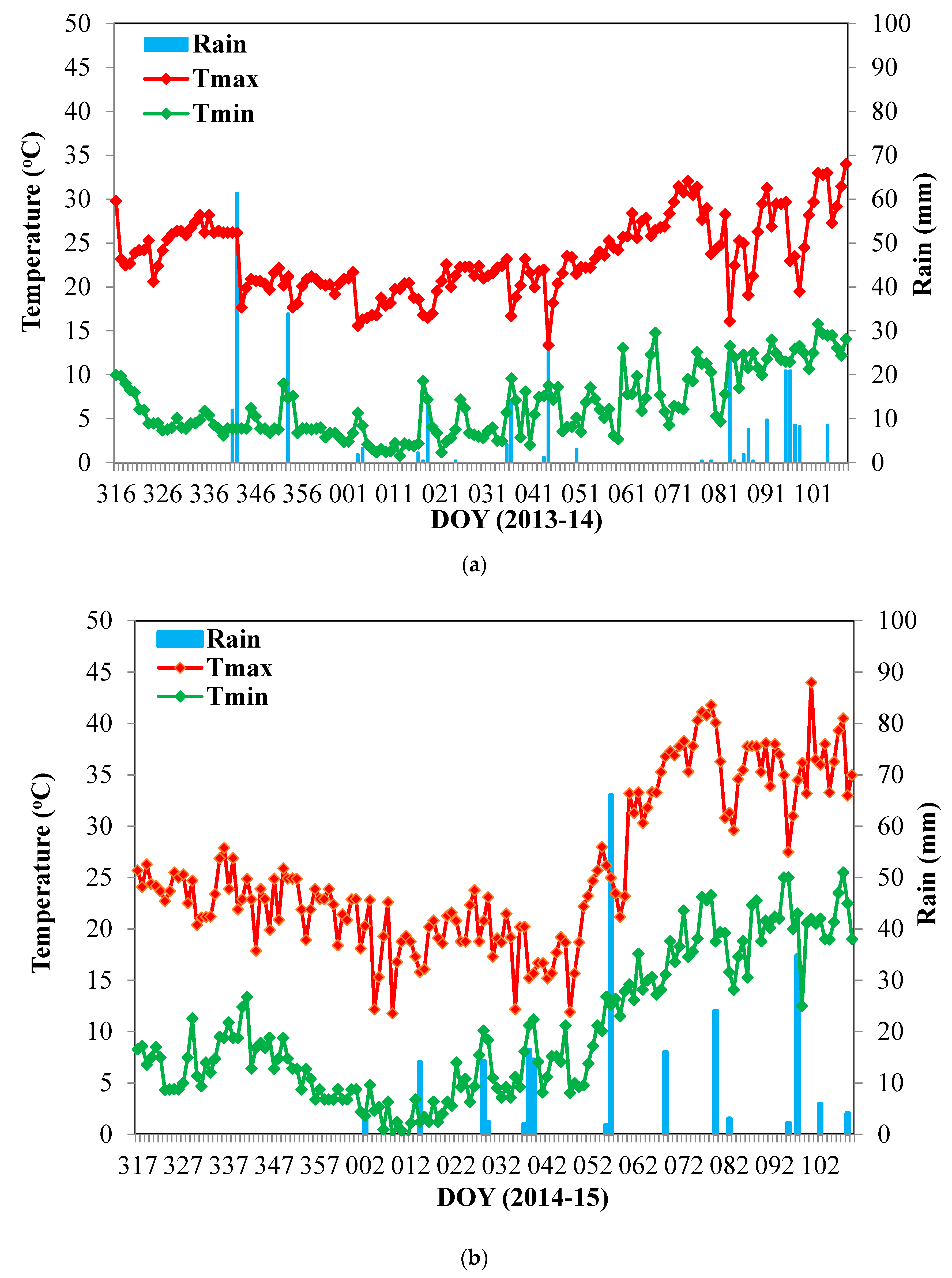
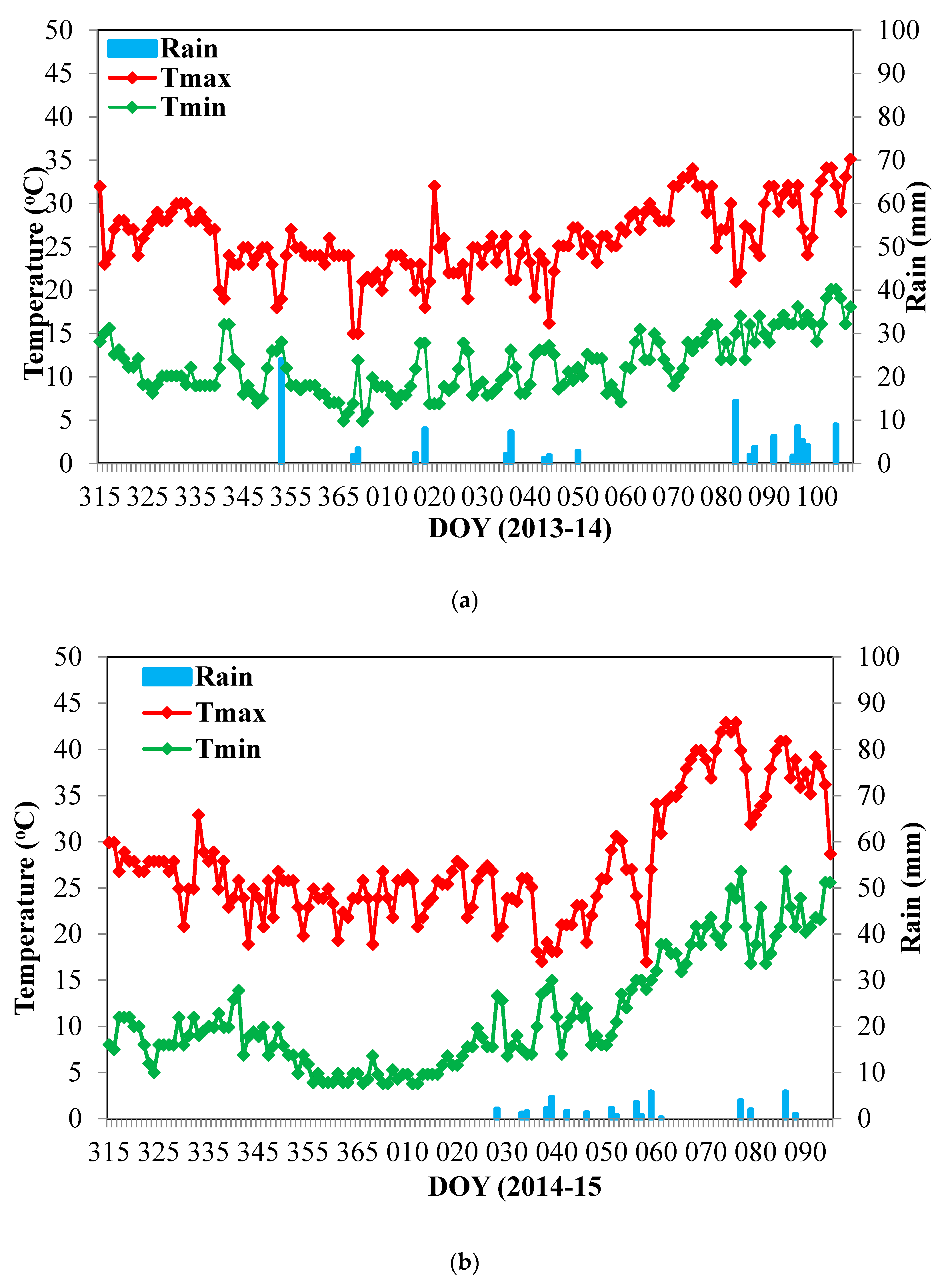
2.1. Study Sites, Treatments, Plant Material, and Experimental Design
2.2. Physiological Traits Measurements
2.3. Crop Total Nitrogen
2.4. Nitrogen Efficiencies
2.5. Agronomic Traits
2.6. Statistical Analysis
3. Results
3.1. Crop Physiological Traits
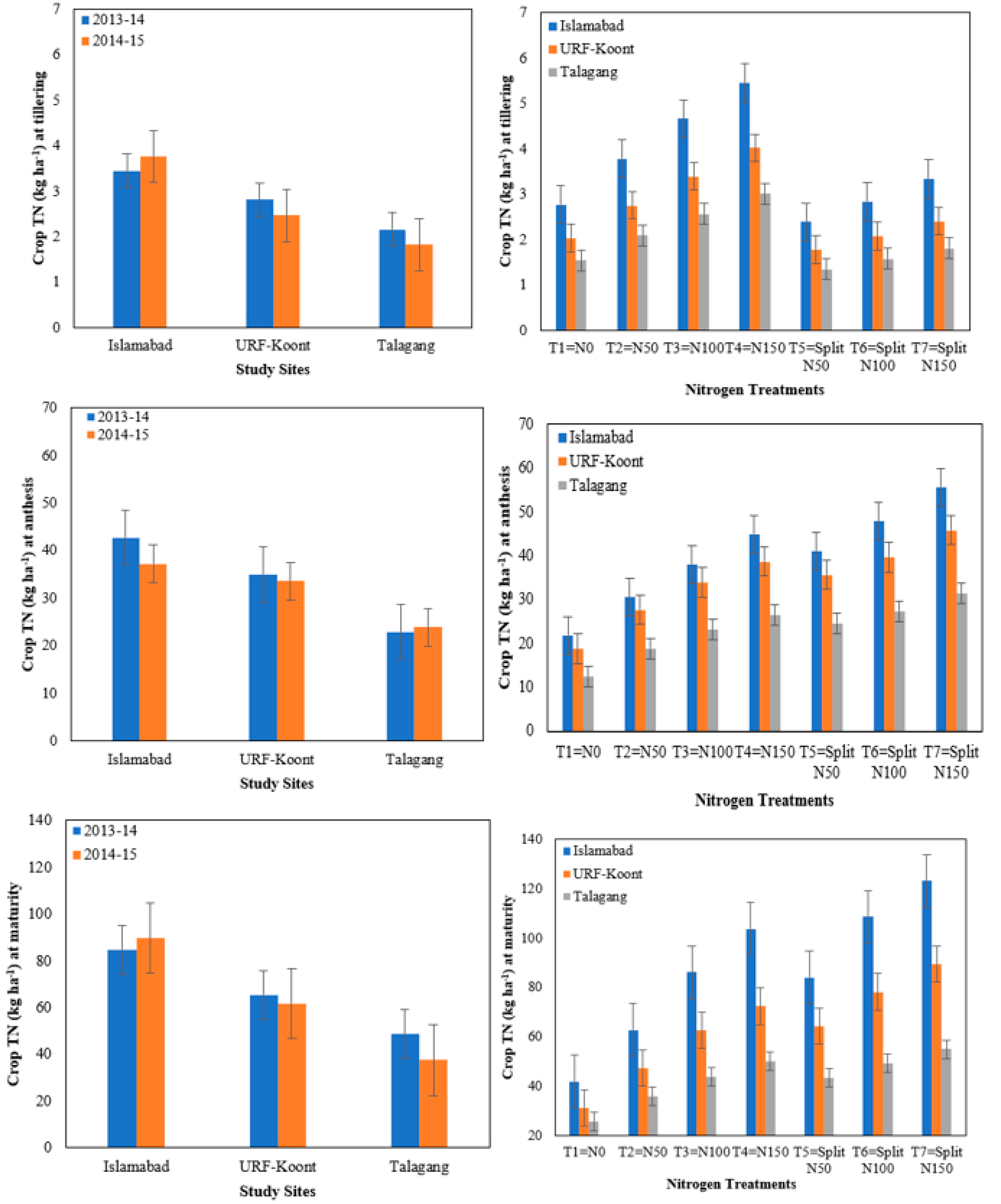
3.2. Crop Total Nitrogen
3.3. Nitrogen Efficiencies
3.3.1. Nitrogen Use Efficiency (kg kg−1)
3.3.2. Nitrogen Uptake Efficiency (NUpE)
3.3.3. Nitrogen Utilization Efficiency (NUtE) (kg kg−1)
3.4. Agronomic Traits
3.5. Relationship of Physiological Traits with Grain Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Drake, B.G.; Gonzàlez-Meler, M.A.; Long, S.P. More efficient plants: A consequence of rising atmospheric CO2? Ann. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 609–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.C.; Yu, X.F.; Ma, Y.F. Effect of nitrogen application and elevated CO2 on photosynthetic gas exchange and electron transport in wheat leaves. Photosynthetica 2013, 51, 593–602. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Isopp, H.; Frehner, M.; Long, S.P.; Nösberger, J. Sucrose-phosphate synthase responds differently to source–sink relations and to photosynthetic rates: Lolium perenne L. growing at elevated pCO2 in the field. Plant Cell Environ. 2000, 23, 597–607. [Google Scholar] [CrossRef]
- Hymus, G.J.; Baker, N.R.; Long, S.P. Growth in elevated CO2 can both decrease and increase photochemistry and photoinhibition of photosynthesis in a predictable manner. Dactylis glomerata grown in two levels of nitrogen nutrition. Plant Physiol. 2001, 127, 1204–1211. [Google Scholar]
- Warren, C.R. The photosynthetic limitation posed by internal conductance to CO2 movement is increased by nutrient supply. J. Exp. Bot. 2004, 55, 2313–2321. [Google Scholar] [CrossRef]
- Gao, J.; Luo, Q.; Sun, C.; Hu, H.; Wang, F.; Tian, Z.; Jiang, D.; Cao, W.; Dai, T. Low Nitrogen Priming Enhances Photosynthesis Adaptation to Water-Deficit Stress in Winter Wheat (Triticum aestivum L.) Seedlings. Front. Plant Sci. 2019, 10, 818. [Google Scholar] [CrossRef]
- Chaves, M.M. Effects of Water Deficits on Carbon Assimilation. J. Exp. Bot. 1991, 42, 1–16. [Google Scholar] [CrossRef]
- Yordanov, I.; Velikova, V.; Tsonev, T. Plant Responses to Drought, Acclimation, and Stress Tolerance. Photosynthetica 2000, 38, 171–186. [Google Scholar] [CrossRef]
- Xin, Y.; Tao, F. Optimizing genotype-environment-management interactions to enhance productivity and eco-efficiency for wheat-maize rotation in the North China Plain. Sci. Total Environ. 2019, 654, 480–492. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, Y.; Xu, X.; Shen, Q.; Guo, S. Light-saturated photosynthetic rate in high-nitrogen rice (Oryza sativa L.) leaves is related to chloroplastic CO2 concentration. J. Exp. Bot. 2009, 60, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Ladha, J.K.; Pathak, H.; Krupnik, T.J.; Six, J.; van Kessel, C. Efficiency of Fertilizer Nitrogen in Cereal Production: Retrospects and Prospects. In Advances in Agronomy; Academic Press: Cambridge, MA, USA, 2005; Volume 87, pp. 85–156. [Google Scholar]
- Jin, X.; Yang, G.; Tan, C.; Zhao, C. Effects of nitrogen stress on the photosynthetic CO2 assimilation, chlorophyll fluorescence, and sugar-nitrogen ratio in corn. Sci. Rep. 2015, 5, 9311. [Google Scholar] [CrossRef]
- Wong, S.-C.; Cowan, I.R.; Farquhar, G.D. Leaf Conductance in Relation to Rate of CO2 Assimilation I. Influence of Nitrogen Nutrition, Phosphorus Nutrition, Photon Flux Density, and Ambient Partial Pressure of CO2 during Ontogeny. Plant Physiol. 1985, 78, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Terashima, I.; Evans, J.R. Effects of light and nitrogen nutrition on the organization of the photosynthetic apparatus in spinach. Plant Cell Physiol. 1988, 29, 143–155. [Google Scholar]
- Rascher, U.; Liebig, M.; Lüttge, U. Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the field. Plant Cell Environ. 2000, 23, 1397–1405. [Google Scholar] [CrossRef]
- Lawlor, D.; Boyle, F.; Young, A.; Keys, A.; Kendall, A. Nitrate nutrition and temperature effects on wheat: Photosynthesis and photorespiration of leaves. J. Exp. Bot. 1987, 38, 393–408. [Google Scholar] [CrossRef]
- Sugiharto, B.; Miyata, K.; Nakamoto, H.; Sasakawa, H.; Sugiyama, T. Regulation of Expression of Carbon-Assimilating Enzymes by Nitrogen in Maize Leaf. Plant Physiol. 1990, 92, 963–969. [Google Scholar] [CrossRef]
- Sage, R.F.; Pearcy, R.W. The Nitrogen Use Efficiency of C3 and C4 Plants II. Leaf Nitrogen Effects on the Gas Exchange Characteristics of Chenopodium album (L.) and Amaranthus retroflexus (L.). Plant Physiol. 1987, 84, 959–963. [Google Scholar] [CrossRef]
- Ciompi, S.; Gentili, E.; Guidi, L.; Soldatini, G.F. The effect of nitrogen deficiency on leaf gas exchange and chlorophyll fluorescence parameters in sunflower. Plant Sci. 1996, 118, 177–184. [Google Scholar] [CrossRef]
- Khamis, S.; Lamaze, T.; Lemoine, Y.; Foyer, C. Adaptation of the Photosynthetic Apparatus in Maize Leaves as a Result of Nitrogen Limitation: Relationships between electron transport and carbon assimilation. Plant Physiol. 1990, 94, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Cao, W.; Jing, Q. Effects of nitrogen form on nitrogen absorption and photosynthesis of different wheat genotypes. Chin. J. Appl. Ecol. 2001, 12, 849–852. [Google Scholar]
- Fan, X.; Jiang, D.; Dai, T.; Jing, Q.; Cao, W. Effects of nitrogen supply on flag leaf photosynthesis and grain starch accumulation of wheat from its anthesis to maturity under drought or waterlogging. Ying Yong Sheng Tai Xue Bao/J. Appl. Ecol. 2005, 16, 1883–1888. [Google Scholar]
- Nunes, M.A.; Ramalho, J.C.; Dias, M.A. Effect of nitrogen supply on the photosynthetic performance of leaves from coffee plants exposed to bright light. J. Exp. Bot. 1993, 44, 893–899. [Google Scholar] [CrossRef]
- Verhoeven, A.S.; Demmig-Adams, B.; Adams, W.W., III. Enhanced Employment of the Xanthophyll Cycle and Thermal Energy Dissipation in Spinach Exposed to High Light and N Stress. Plant Physiol. 1997, 113, 817–824. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Ecophysiology of photosynthesis. Ecol. Stud. 1994, 100, 49–70. [Google Scholar]
- Bungard, R.A.; McNeil, D.; Morton, J.D. Effects of nitrogen on the photosynthetic apparatus of Clematis vitalba grown at several irradiances. Funct. Plant Biol. 1997, 24, 205–214. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Photosynthetic CO2 assimilation, chlorophyll fluorescence and photoinhibition as affected by nitrogen deficiency in maize plants. Plant Sci. 2000, 151, 135–143. [Google Scholar] [CrossRef]
- Zhang, L.; Shangguan, Z.; Mao, M.; Yu, G. Effects of long-term application of nitrogen fertilizer on leaf chlorophyll fluorescence of upland winter wheat. Ying Yong Sheng Tai Xue Bao/J. Appl. Ecol. 2003, 14, 695–698. [Google Scholar]
- Cai, R.-G.; Zhang, M.; Yin, Y.-P.; Ping, W.; Zhang, T.B.; Feng, G.; Dai, Z.-M.; Liang, T.-B.; Wu, Y.-H.; Wang, Z.-L. Photosynthetic characteristics and antioxidative metabolism of flag leaves in responses to nitrogen application during grain filling of field-grown wheat. Agric. Sci. China 2008, 7, 157–167. [Google Scholar] [CrossRef]
- Filella, I.; Serrano, L.; Serra, J.; Penuelas, J. Evaluating wheat nitrogen status with canopy reflectance indices and discriminant analysis. Crop Sci. 1995, 35, 1400–1405. [Google Scholar] [CrossRef]
- Moran, J.A.; Mitchell, A.K.; Goodmanson, G.; Stockburger, K.A. Differentiation among effects of nitrogen fertilization treatments on conifer seedlings by foliar reflectance: A comparison of methods. Tree Physiol. 2000, 20, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, S.; Brueck, H.; Asch, F. Chlorophyll index, photochemical reflectance index and chlorophyll fluorescence measurements of rice leaves supplied with different N levels. J. Photochem. Photobiol. B Biol. 2012, 113, 7–13. [Google Scholar] [CrossRef]
- Zhong, S.; Xu, Y.; Meng, B.; Loik, M.E.; Ma, J.-Y.; Sun, W. Nitrogen Addition Increases the Sensitivity of Photosynthesis to Drought and Re-watering Differentially in C3 Versus C4 Grass Species. Front. Plant Sci. 2019, 10, 815. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Garcia, P.C.; López-Lefebre, L.R.; Rivero, R.M.; Ruiz, J.M.; Romero, L. Proline metabolism in response to nitrogen deficiency in French Bean plants (Phaseolus vulgaris L. cv Strike). Plant Growth Regul. 2002, 36, 261–265. [Google Scholar] [CrossRef]
- Rare, E. Stress physiology: The functional significance of the accumulation of nitrogen-containing compounds. J. Hortic. Sci 1990, 65, 231–243. [Google Scholar] [CrossRef]
- Burns, I.G. Nitrogen supply, growth and development. Acta Hortic. 1996, 428, 21–30. [Google Scholar] [CrossRef]
- Makino, A. Photosynthesis, Grain Yield, and Nitrogen Utilization in Rice and Wheat. Plant Physiol. 2011, 155, 125–129. [Google Scholar] [CrossRef]
- Van Ogtrop, F.; Ahmad, M.; Moeller, C. Principal components of sea surface temperatures as predictors of seasonal rainfall in rainfed wheat growing areas of Pakistan. Meteorol. Appl. 2014, 21, 431–443. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Bremner, J.M.; Breitenbeck, G.A. A simple method for determination of ammonium in semimicro-Kjeldahl analysis of soils and plant materials using a block digester. Commun. Soil Sci. Plant Anal. 1983, 14, 905–913. [Google Scholar] [CrossRef]
- Rahimizadeh, M.; Kashani, A.; Zare-Feizabadi, A.; Koocheki, A.R.; Nassiri-Mahallati, M. Nitrogen use efficiency of wheat as affected by preceding crop, application rate of nitrogen and crop residues. Aust. J. Crop Sci. 2010, 4, 363–368. [Google Scholar]
- Ahmed, M.; Hassan, F.-U.; Aslam, M.; Akram, M.N.; Aslam, M.A. Photosynthesis of spring wheat (Triticum aestivum) in rainfed ecology of Pakistan. Afr. J. Biotechnol. 2010, 9, 7495–7503. [Google Scholar]
- Yu, O.; Goudriaan, J.; Wang, T.-D. Modelling Diurnal Courses of Photosynthesis and Transpiration of Leaves on the Basis of Stomatal and Non-Stomatal Responses, Including Photoinhibition. Photosynthetica 2001, 39, 43–51. [Google Scholar] [CrossRef]
- Kimball, B.A.; Kobayashi, K.; Bindi, M. Responses of Agricultural Crops to Free-Air CO2 Enrichment. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2002; Volume 77, pp. 293–368. [Google Scholar]
- Gyuga, P.; Demagante, A.L.; Paulsen, G.M. Photosynthesis and grain growth of wheat under extreme nitrogen nutrition regimes during maturation. J. Plant Nutr. 2002, 25, 1281–1290. [Google Scholar] [CrossRef]
- Abid, M.; Tian, Z.; Ata-Ul-Karim, S.T.; Cui, Y.; Liu, Y.; Zahoor, R.; Jiang, D.; Dai, T. Nitrogen Nutrition Improves the Potential of Wheat (Triticum aestivum L.) to Alleviate the Effects of Drought Stress during Vegetative Growth Periods. Front. Plant Sci. 2016, 7, 981. [Google Scholar] [CrossRef]
- Zlatev, Z.; Lidon, F.C. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emir. J. Food Agric. 2012, 24, 57–72. [Google Scholar]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Grassi, G.; Magnani, F. Stomatal, mesophyll conductance and biochemical limitations to photosynthesis as affected by drought and leaf ontogeny in ash and oak trees. Plant Cell Environ. 2005, 28, 834–849. [Google Scholar] [CrossRef]
- Ahmed, M.; ul Hassan, F. Response of Spring Wheat (Triticum aestivum L.) Quality Traits and Yield to Sowing Date. PLoS ONE 2015, 10, e0126097. [Google Scholar] [CrossRef] [PubMed]
- Boretti, A.; Florentine, S. Atmospheric CO2 Concentration and Other Limiting Factors in the Growth of C3 and C4 Plants. Plants 2019, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Hassan, F.U.; Qadir, G.; Shaheen, F.A.; Aslam, M.A. Response of proline accumulation in bread wheat (Triticum aestivum L.) under rainfed conditions. J. Agric. Meteorol. 2017. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Ben Rejeb, K.; Abdelly, C.; Savouré, A. How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Filippou, P.; Bouchagier, P.; Skotti, E.; Fotopoulos, V. Proline and reactive oxygen/nitrogen species metabolism is involved in the tolerant response of the invasive plant species Ailanthus altissima to drought and salinity. Environ. Exp. Bot. 2014, 97, 1–10. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Panda, R.K.; Chakraborty, A.; Halder, D. Enhancing grain yield, biomass and nitrogen use efficiency of maize by varying sowing dates and nitrogen rate under rainfed and irrigated conditions. Field Crops Res. 2018, 221, 339–349. [Google Scholar] [CrossRef]
- Guarda, G.; Padovan, S.; Delogu, G. Grain yield, nitrogen-use efficiency and baking quality of old and modern Italian bread-wheat cultivars grown at different nitrogen levels. Eur. J. Agron. 2004, 21, 181–192. [Google Scholar] [CrossRef]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef]
- Raun, W.R.; Solie, J.B.; Johnson, G.V.; Stone, M.L.; Mullen, R.W.; Freeman, K.W.; Thomason, W.E.; Lukina, E.V. Improving nitrogen use efficiency in cereal grain production with optical sensing and variable rate application. Agron. J. 2002, 94, 815–820. [Google Scholar] [CrossRef]
- Meng, Q.; Yue, S.; Hou, P.; Cui, Z.; Chen, X. Improving Yield and Nitrogen Use Efficiency Simultaneously for Maize and Wheat in China: A Review. Pedosphere 2016, 26, 137–147. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Griffiths, S. Exploiting genetic variation in nitrogen use efficiency for cereal crop improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, H.; Lei, Q.; Luo, J.; Lindsey, S.; Zhang, J.; Zhai, L.; Wu, S.; Zhang, J.; Liu, X.; et al. Optimizing the nitrogen application rate for maize and wheat based on yield and environment on the Northern China Plain. Sci. Total Environ. 2018, 618, 1173–1183. [Google Scholar] [CrossRef]
- Faralli, M.; Matthews, J.; Lawson, T. Exploiting natural variation and genetic manipulation of stomatal conductance for crop improvement. Curr. Opin. Plant Biol. 2019, 49, 1–7. [Google Scholar] [CrossRef]
- Ahmed, M.; Aslam, M.A.; Hassan, F.; Hayat, R.; Ahmad, S. Biochemical, physiological and agronomic response of wheat to changing climate of reinfed areas of pakistan. Pak. J. Bot. 2018, 51, 535–551. [Google Scholar]
- Saud, S.; Fahad, S.; Yajun, C.; Ihsan, M.Z.; Hammad, H.M.; Nasim, W.; Amanullah, Jr.; Arif, M.; Alharby, H. Effects of Nitrogen Supply on Water Stress and Recovery Mechanisms in Kentucky Bluegrass Plants. Front. Plant Sci. 2017, 8, 983. [Google Scholar] [CrossRef]
- Ahmed, M.; Hassan, F.-U. Cumulative effect of temperature and solar radiation on wheat yield. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2011, 39, 146–152. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Islamabad | URF-Koont | Talagang | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Determinations (2013–14) | Units | 0–15 | 15–30 | 30–45 | 0–15 | 15–30 | 30–45 | 0–15 | 15–30 | 30–45 |
| pH | 1:01 | 7.5 | 7.6 | 8.3 | 8.1 | 8.6 | 8.8 | 8.3 | 8.4 | 8.5 |
| EC | dSm−1 | 0.24 | 0.2 | 0.21 | 0.32 | 0.34 | 0.26 | 0.28 | 0.27 | 0.29 |
| Nitrogen | % | 0.042 | 0.041 | 0.039 | 0.04 | 0.36 | 0.33 | 0.032 | 0.031 | 0.029 |
| Nitrate-N | mg Kg−1 | 7.86 | 7.28 | 6.5 | 4.52 | 4.21 | 4.04 | 3.16 | 3.34 | 3.12 |
| AV.P | mg kg−1 | 3.68 | 3.46 | 3.41 | 2.8 | 3 | 2.86 | 2 | 2.16 | 2.32 |
| K | mg kg−1 | 156 | 175 | 177 | 114 | 150 | 157 | 118 | 108 | 109 |
| Organic Carbon | % | 0.89 | 0.76 | 0.68 | 0.72 | 0.47 | 0.45 | 0.64 | 0.75 | 0.72 |
| Silt | % | 0.35 | 0.35 | 0.35 | 23 | 21 | 20 | 27 | 28 | 28 |
| Sand | % | 0.31 | 0.31 | 0.31 | 56 | 56 | 56 | 58 | 56 | 55 |
| Clay | % | 0.34 | 0.34 | 0.34 | 21 | 23 | 24 | 15 | 16 | 17 |
| Texture | Loam | Loam | Loam | Sandy clay loam | Sandy clay loam | Sandy clay loam | sandy loam | sandy loam | sandy loam | |
| B. Density | gcm−3 | 1.24 | 1.42 | 1.46 | 1.31 | 1.58 | 1.66 | 1.51 | 1.59 | 1.68 |
| SLL | mmmm−1 | 0.07 | 0.09 | 0.198 | 0.061 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 |
| SDUL | mmmm−1 | 0.38 | 0.30 | 0.30 | 0.28 | 0.23 | 0.22 | 0.18 | 0.18 | 0.15 |
| Saturated SW | mmmm−1 | 0.417 | 0.424 | 0.387 | 0.38 | 0.35 | 0.31 | 0.36 | 0.33 | 0.3 |
| SW at tillering (BBCH20) | mmmm−1 | 0.30 | 0.24 | 0.23 | 0.24 | 0.18 | 0.18 | 0.15 | 0.15 | 0.14 |
| SW at anthesis (BBCH60) | mmmm−1 | 0.33 | 0.25 | 0.23 | 0.24 | 0.19 | 0.19 | 0.16 | 0.16 | 0.15 |
| Islamabad | URF-Koont | Talagang | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Determinations (2014–15) | Units | 0–15 | 15–30 | 30–45 | 0–15 | 30–45 | 45–60 | 15–30 | 30–45 | 45–60 |
| pH | 1:01 | 7.4 | 7.5 | 7.9 | 8.2 | 8.1 | 8 | 7.9 | 8 | 7.7 |
| EC | dSm−1 | 0.23 | 0.2 | 0.2 | 0.28 | 0.31 | 0.31 | 0.21 | 0.23 | 0.23 |
| Nitrogen | % | 0.041 | 0.042 | 0.039 | 0.03 | 0.02 | 0.02 | 0.031 | 0.036 | 0.035 |
| Nitrate-N | mg Kg−1 | 6.4 | 5.9 | 5.3 | 2.58 | 2.77 | 2.64 | 2.61 | 2.51 | 2.4 |
| AV.P | mg kg−1 | 3.1 | 2.9 | 3.3 | 2.38 | 2.55 | 2.43 | 1.92 | 2.07 | 2.23 |
| K | mg kg−1 | 120 | 135 | 159 | 109 | 143 | 149 | 115 | 101 | 106 |
| Organic Carbon | % | 0.72 | 0.69 | 0.5 | 0.68 | 0.45 | 0.43 | 0.64 | 0.75 | 0.72 |
| Silt | % | 0.35 | 0.35 | 0.35 | 23 | 21 | 20 | 27 | 28 | 28 |
| Sand | % | 0.31 | 0.3 | 0.31 | 56 | 56 | 56 | 58 | 56 | 55 |
| Clay | % | 0.34 | 0.35 | 0.34 | 21 | 23 | 24 | 15 | 16 | 17 |
| Texture | Loam | Loam | Loam | Sandy clay loam | Sandy clay loam | Sandy clay loam | sandy loam | sandy loam | sandy loam | |
| BD | gcm−3 | 1.22 | 1.4 | 1.44 | 1.29 | 1.55 | 1.65 | 1.53 | 1.61 | 1.7 |
| SLL | mmmm−1 | 0.09 | 0.198 | 0.186 | 0.057 | 0.076 | 0.076 | 0.07 | 0.08 | 0.08 |
| SDUL | mmmm−1 | 0.29 | 0.28 | 0.26 | 0.26 | 0.21 | 0.21 | 0.23 | 0.23 | 0.25 |
| Saturated SW | mmmm−1 | 0.424 | 0.419 | 0.359 | 0.38 | 0.33 | 0.29 | 0.36 | 0.33 | 0.3 |
| SW at tillering (BBCH20) | mmmm−1 | 0.23 | 0.25 | 0.21 | 0.23 | 0.17 | 0.17 | 0.20 | 0.20 | 0.22 |
| SW at anthesis (BBCH60) | mmmm−1 | 0.26 | 0.26 | 0.22 | 0.24 | 0.18 | 0.18 | 0.21 | 0.21 | 0.23 |
| gs (mole H2O m−2 s−1) | Rs (m2 s mole−1) | An μmole CO2 m−2 s−1 | E (mmole H2O m−2 s−1) | Ci (μmole CO2 mol−1 air) | Chlorophyll Contents (SPAD) | Proline Content (μg g−1) | |
|---|---|---|---|---|---|---|---|
| Years (Y) | |||||||
| 2013–14 | 0.65 a | 0.73 b | 18.70 a | 7.52 a | 298.41 a | 54.672 a | 30.77 b |
| 2014–15 | 0.50 b | 1.05 a | 16.24 b | 6.32 b | 272.60 b | 45.56 b | 38.25 a |
| Study Sites/Locations (L) | |||||||
| Islamabad | 0.67 a | 0.66 c | 18.88 a | 7.95 a | 317.79 a | 51.80 a | 27.85 c |
| URF-Koont | 0.56 b | 0.87 b | 16.77 b | 6.89 b | 290.12 b | 46.79 b | 33.86 b |
| Talagang | 0.46 c | 1.07 a | 15.55 c | 6.13 c | 252.60 c | 40.29 c | 40.13 a |
| Nitrogen Treatments (T) | |||||||
| T1 = N0 | 0.60 c | 0.71 a | 15.25 c | 6.82 c | 290.79 d | 44.51 d | 27.97 d |
| T2 = N50 | 0.73 b | 0.56 b | 17.50 b | 8.15 b | 310.18 c | 48.39 c | 29.52 c |
| T3 = N100 | 0.78 a | 0.49 c | 18.10 b | 8.75 b | 318.33 b | 54.55 b | 32.15 b |
| T4 = N150 | 0.79 a | 0.48 c | 19.49 a | 9.22 a | 320.23 b | 57.28 a | 33.82 b |
| T5 = Split N50 | 0.73 b | 0.56 b | 17.52 b | 8.15 b | 310.19 c | 54.39 b | 30.74 c |
| T6 = Split N100 | 0.80 a | 0.48 c | 20.07 a | 9.58 a | 329.25 a | 58.86 a | 35.42 a |
| T7 = Split N150 | 0.80 a | 0.48 c | 19.79 a | 9.31 a | 325.27 a | 58.88 a | 35.42 a |
| Interactions | |||||||
| Y × L | *** | *** | *** | *** | NS | NS | *** |
| Y × T | NS | NS | NS | NS | NS | NS | NS |
| L × T | NS | NS | NS | NS | NS | *** | NS |
| Y × L × T | NS | NS | NS | NS | NS | NS | NS |
| Crop N at Tillering (BBCH20) (kg N ha−1) | Crop N at Anthesis (BBCH60) (kg N ha−1) | Crop N at Maturity (BBCH90) (kg N ha−1) | |
|---|---|---|---|
| Years (Y) | |||
| 2013–14 | 2.81 a | 33.53 a | 66.417 a |
| 2014–15 | 2.68 b | 31.51 b | 62.95 b |
| Study Sites/Locations (L) | |||
| Islamabad | 3.61 a | 39.93 a | 87.22 a |
| URF-Koont | 2.64 b | 34.25 b | 63.60 b |
| Talagang | 1.99 c | 23.38 c | 43.22 c |
| Nitrogen Treatments (T) | |||
| T1 = N0 | 2.11 e | 17.64 f | 32.91 e |
| T2 = N50 | 2.87 c | 25.64 e | 48.60 d |
| T3 = N100 | 3.54 b | 31.61 d | 64.17 c |
| T4 = N150 | 4.16 a | 36.62 b | 75.32 b |
| T5 = Split N50 | 1.84 f | 33.70 c | 63.81 c |
| T6 = Split N100 | 2.17 e | 38.18 b | 78.74 b |
| T7 = Split N150 | 2.52 d | 44.23 a | 89.20 a |
| Interactions | |||
| Y × L | *** | *** | *** |
| Y × T | NS | NS | *** |
| L × T | *** | *** | *** |
| Y × L × T | NS | NS | *** |
| Years (Y) | NUE | NUtE | NUpE |
|---|---|---|---|
| 2013–14 | 18.35 a | 44.18 b | 0.41 a |
| 2014–15 | 16.63 b | 44.35 a | 0.39 b |
| Study Sites/Locations (L) | |||
| ISLAMABAD | 22.96 a | 43.53 b | 0.53 a |
| URF-Koont | 18.22 b | 47.08 a | 0.39 b |
| Talagang | 11.31 c | 42.20 c | 0.27 c |
| Nitrogen Treatments (T) | |||
| T1 = N0 | 21.72 a | 52.78 b | 0.41 bc |
| T2 = N50 | 20.03 b | 53.98 a | 0.37 d |
| T3 = N100 | 16.49 c | 46.33 c | 0.35 de |
| T4 = N150 | 13.48 d | 41.38 d | 0.32 e |
| T5 = Split N50 | 20.03 b | 40.84 e | 0.49 a |
| T6 = Split N100 | 17.31 c | 39.42 f | 0.43 b |
| T7 = Split N150 | 13.39 d | 35.15 g | 0.38 cd |
| Interactions | |||
| Y × L | NS | *** | *** |
| Y × T | NS | NS | *** |
| L × T | NS | *** | *** |
| Y × L × T | NS | NS | *** |
| No of Tillers m−2 | 1000 Grain Weight (gm) | Biological Yield (kg ha−1) | Grain Yield (kg ha−1) | Harvest Index | |
|---|---|---|---|---|---|
| Years (Y) | |||||
| 2013–14 | 222.4 NS | 31.3 NS | 9380.9 a | 3001.9 a | 0.32 a |
| 2014–15 | 212.7 | 30.7 | 8704.7 b | 2611.4 b | 0.3 b |
| Study Sites/Locations (L) | |||||
| Islamabad | 236.5 a | 34.3 a | 10,450.1 a | 3957.5 a | 0.35 a |
| URF-Koont | 226.9 a | 33.9 a | 8578.2 b | 2830.8 b | 0.33 b |
| Talagang | 189.2 b | 24.7 b | 7042.4 c | 1760.6 c | 0.25 c |
| Nitrogen Treatments (T) | |||||
| T1 = N0 | 184.5 c | 23.5 c | 5992.4 c | 1737.8 c | 0.29 c |
| T2 = N50 | 205.8 b | 28.1 b | 8140.6 b | 2605 b | 0.32 a |
| T3 = N100 | 218.8 ab | 31.7 a | 9275.9 a | 2968.3 a | 0.32 a |
| T4 = N150 | 229.3 a | 33.1 a | 10,942.8 a | 3501.7 a | 0.32 a |
| T5 = Split N50 | 221.1 ab | 31.6 a | 8982.7 b | 2605 b | 0.29 c |
| T6 = Split N100 | 234.4 a | 34.5 a | 11,345.5 a | 3517.2 a | 0.31 b |
| T7 = Split N150 | 229.1 a | 34.3 a | 10,976.3 a | 3512.4 a | 0.32 a |
| Interactions | |||||
| Y × L | *** | NS | *** | NS | *** |
| Y × T | NS | NS | NS | NS | *** |
| L × T | NS | *** | NS | *** | *** |
| Y × L × T | NS | NS | NS | NS | *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qadeer, U.; Ahmed, M.; -Hassan, F.-u.; Akmal, M. Impact of Nitrogen Addition on Physiological, Crop Total Nitrogen, Efficiencies and Agronomic Traits of the Wheat Crop under Rainfed Conditions. Sustainability 2019, 11, 6486. https://doi.org/10.3390/su11226486
Qadeer U, Ahmed M, -Hassan F-u, Akmal M. Impact of Nitrogen Addition on Physiological, Crop Total Nitrogen, Efficiencies and Agronomic Traits of the Wheat Crop under Rainfed Conditions. Sustainability. 2019; 11(22):6486. https://doi.org/10.3390/su11226486
Chicago/Turabian StyleQadeer, Umara, Mukhtar Ahmed, Fayyaz-ul -Hassan, and Muhammad Akmal. 2019. "Impact of Nitrogen Addition on Physiological, Crop Total Nitrogen, Efficiencies and Agronomic Traits of the Wheat Crop under Rainfed Conditions" Sustainability 11, no. 22: 6486. https://doi.org/10.3390/su11226486
APA StyleQadeer, U., Ahmed, M., -Hassan, F.-u., & Akmal, M. (2019). Impact of Nitrogen Addition on Physiological, Crop Total Nitrogen, Efficiencies and Agronomic Traits of the Wheat Crop under Rainfed Conditions. Sustainability, 11(22), 6486. https://doi.org/10.3390/su11226486