Assessing the Biostimulant Effects of a Novel Plant-Based Formulation on Tomato Crop

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Set Up
2.2. Preliminary Test
2.3. Main Experiment
2.4. Plant Growth and Physiological Parameters
2.5. Total Phenols Content and Antioxidant Activity
2.6. Hydrogen Peroxide, Lipid Peroxidation and Antioxidative Enzyme Activities
2.7. Plant Nutrient Content
2.8. Fresh Produce Quality
2.9. Statistical Methods
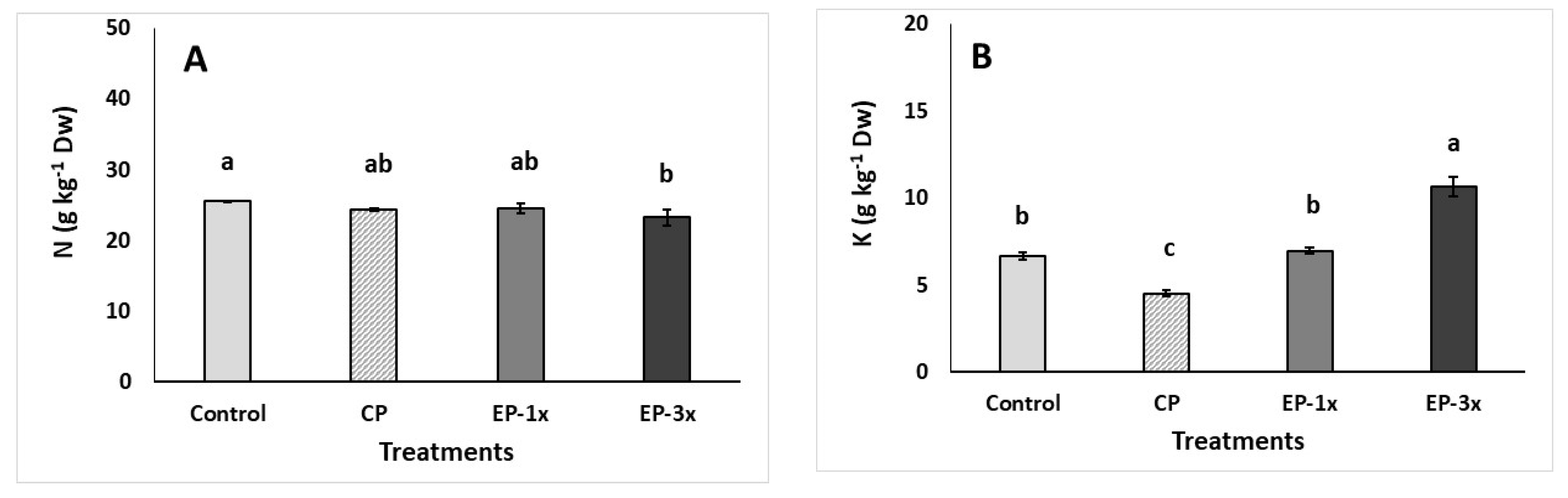
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lucini, L.; Colla, G.; Miras-Moreno, B.; Bernardo, L.; Cardarelli, M.; Terzi, V.; Bonini, P.; Rouphael, Y. Inoculation of Rhizoglomus irregulare or Trichoderma atroviride differentially modulates metabolite profiling of wheat root exudates. Phytochemistry 2019, 157, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2014, 31, 1–17. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drobek, M.; Frąc, M.; Cybulska, J. Plant Biostimulants: Importance of the Quality and Yield of Horticultural Crops and the Improvement of Plant Tolerance to Abiotic Stress—A Review. Agronmy 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.H.; Saa, S. Biostimulants in agriculture. Front. Plant Sci. 2015, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Rouphael, Y.; Cardarelli, M.; Bonini, P.; Colla, G. Synergistic Action of a Microbial-based Biostimulant and a Plant Derived-Protein Hydrolysate Enhances Lettuce Tolerance to Alkalinity and Salinity. Front. Plant Sci. 2017, 8, 131. [Google Scholar] [CrossRef] [Green Version]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Kałużewicz, A.; Gąsecka, M.; Spiżewski, T. Influence of biostimulants on phenolic content in broccoli heads directly after harvest and after storage. Folia Hortic. 2017, 29, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Mahmoud, E.A.; El-Ansary, D.O.; Mattar, M.A. Effects of Water Stress and Modern Biostimulants on Growth and Quality Characteristics of Mint. Agronmy 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Di Mattia, E.; El-Nakhel, C.; Cardarelli, M. Co-inoculation of Glomus intraradices and Trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 2015, 95, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Litskas, V.; Chrysargyris, A.; Stavrinides, M.C.; Tzortzakis, N. Water-energy-food nexus: A case study on medicinal and aromatic plants. J. Clean. Prod. 2019, 233, 1334–1343. [Google Scholar] [CrossRef]
- Hamedani, S.R.; Rouphael, Y.; Colla, G.; Colantoni, A.; Cardarelli, M. Biostimulants as a Tool for Improving Environmental Sustainability of Greenhouse Vegetable Crops. Sustainability 2020, 12, 5101. [Google Scholar] [CrossRef]
- Dudaš, S.; Šola, I.; Sladonja, B.; Erhatić, R.; Ban, D.; Poljuha, D. The Effect of Biostimulant and Fertilizer on “Low Input” Lettuce Production. Acta Bot. Croat. 2016, 75, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Anastasiou, M.; Pantelides, I.S.; Tzortzakis, N. Effects of Ascophyllum nodosum seaweed extracts on lettuce growth, physiology and fresh-cut salad storage under potassium deficiency. J. Sci. Food Agric. 2018, 98, 5861–5872. [Google Scholar] [CrossRef]
- Pereira, C.; Dias, M.I.; Petropoulos, S.A.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Calhelha, R.C.; Soković, M.; Stojković, D.; Barros, L.; et al. The Effects of Biostimulants, Biofertilizers and Water-Stress on Nutritional Value and Chemical Composition of Two Spinach Genotypes (Spinacia oleracea L.). Molecules 2019, 24, 4494. [Google Scholar] [CrossRef] [Green Version]
- Fan, D.; Hodges, D.M.; Zhang, J.; Kirby, C.W.; Ji, X.; Locke, S.J.; Critchley, A.T.; Prithiviraj, B. Commercial extract of the brown seaweed Ascophyllum nodosum enhances phenolic antioxidant content of spinach (Spinacia oleracea L.) which protects Caenorhabditis elegans against oxidative and thermal stress. Food Chem. 2011, 124, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Rady, M.M.; Mohamed, G.F. Modulation of salt stress effects on the growth, physio-chemical attributes and yields of Phaseolus vulgaris L. plants by the combined application of salicylic acid and Moringa oleifera leaf extract. Sci. Hortic. 2015, 193, 105–113. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O’Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Cuciniello, A.; Cenvinzo, V.; Bonini, P.; Colla, G.; Rouphael, Y. Yield and Nutritional Quality of Vesuvian Piennolo Tomato PDO as Affected by Farming System and Biostimulant Application. Agronmy 2019, 9, 505. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Koleška, I.; Hasanagić, D.; Todorović, V.; Murtić, S.; Klokić, I.; Parađiković, N.; Kukavica, B. Biostimulant prevents yield loss and reduces oxidative damage in tomato plants grown on reduced NPK nutrition. J. Plant Interact. 2017, 12, 209–218. [Google Scholar] [CrossRef] [Green Version]
- González-González, M.F.; Ocampo-Alvarez, H.; Santacruz-Ruvalcaba, F.; Sánchez-Hernández, C.V.; Casarrubias-Castillo, K.; Becerril-Espinosa, A.; Castañeda-Nava, J.J.; Hernández-Herrera, R.M. Physiological, Ecological, and Biochemical Implications in Tomato Plants of Two Plant Biostimulants: Arbuscular Mycorrhizal Fungi and Seaweed Extract. Front. Plant Sci. 2020, 11, 999. [Google Scholar] [CrossRef] [PubMed]
- Souri, M.K.; Bakhtiarizade, M. Biostimulation effects of rosemary essential oil on growth and nutrient uptake of tomato seedlings. Sci. Hortic. 2019, 243, 472–476. [Google Scholar] [CrossRef]
- Zanin, L.; Tomasi, N.; Cesco, S.; Varanini, Z.; Pinton, R. Humic Substances Contribute to Plant Iron Nutrition Acting as Chelators and Biostimulants. Front. Plant Sci. 2019, 10, 675. [Google Scholar] [CrossRef] [Green Version]
- Sfara, V.; Zerba, E.N.; Alzogaray, R.A. Fumigant insecticidal activity and repellent effect of five essential oils and seven monoterpenes on first-instar nymphs of Rhodnius prolixus. J. Med Entomol. 2009, 46, 511–515. [Google Scholar] [CrossRef]
- Sun, Y.; Cai, X.; Cao, J.; Wu, Z.; Pan, D. Effects of 1,8-cineole on Carbohydrate Metabolism Related Cell Structure Changes of Salmonella. Front. Microbiol. 2018, 9, 1078. [Google Scholar] [CrossRef]
- Petrović, J.; Glamočlija, J.; Marin, P.D.; Brkić, D.; Van Griensven, L.J. Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an In Vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef] [Green Version]
- Bedoya-Pérez, M.A.; Isler, I.; Banks, P.B.; McArthur, C. Roles of the volatile terpene, 1,8-cineole, in plant–herbivore interactions: A foraging odor cue as well as a toxin? Oecologia 2013, 174, 827–837. [Google Scholar] [CrossRef]
- Tzortzakis, N. Maintaining postharvest quality of fresh produce with volatile compounds. Innov. Food Sci. Emerg. Technol. 2007, 8, 111–116. [Google Scholar] [CrossRef]
- Caldas, G.F.R.; Oliveira, A.R.D.S.; Araújo, A.V.; Lafayette, S.S.L.; Albuquerque, G.; Silva-Neto, J.D.C.; Costa-Silva, J.H.; Ferreira, F.; Da Costa, J.G.M.; Wanderley, A.G. Gastroprotective Mechanisms of the Monoterpene 1,8-Cineole (Eucalyptol). PLOS ONE 2015, 10, e0134558. [Google Scholar] [CrossRef] [Green Version]
- Chrysargyris, A.; Xylia, P.; Akinci, G.; Moustakas, K.; Tzortzakis, N. Printed Paper Waste as an Alternative Growing Medium Component to Produce Brassica Seedlings under Nursery Conditions. Sustainability 2020, 12, 5992. [Google Scholar] [CrossRef]
- Inouye, S.; Takizawa, T.; Yamaguchi, H. Antibacterial activity of essential oils and their major constituents against respiratory tract pathogens by gaseous contact. J. Antimicrob. Chemother. 2001, 47, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrysargyris, A.; Nikolaidou, E.; Stamatakis, A.; Tzortzakis, N. Vegetative, physiological, nutritional and antioxidant behavior of spearmint (Mentha spicata L.) in response to different nitrogen supply in hydroponics. J. Appl. Res. Med. Aromat. Plants 2017, 6, 52–61. [Google Scholar] [CrossRef]
- Chondraki, S.; Tzerakis, C.; Tzortzakis, N. Influence of sodium chloride and calcium foliar spray on hydroponically grown parsley in nutrient film technique system. J. Plant Nutr. 2012, 35, 1457–1467. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Papakyriakou, E.; Petropoulos, S.A.; Tzortzakis, N. The combined and single effect of salinity and copper stress on growth and quality of Mentha spicata plants. J. Hazard. Mater. 2019, 368, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Antoniou, O.; Tzionis, A.; Prasad, M.; Tzortzakis, N. Alternative soilless media using olive-mill and paper waste for growing ornamental plants. Environ. Sci. Pollut. Res. 2018, 25, 35915–35927. [Google Scholar] [CrossRef]
- Xylia, P.; Clark, A.; Chrysargyris, A.; Romanazzi, G.; Tzortzakis, N. Quality and safety attributes on shredded carrots by using Origanum majorana and ascorbic acid. Postharvest Biol. Technol. 2019, 155, 120–129. [Google Scholar] [CrossRef]
- Tzortzakis, N.; Saridakis, C.; Chrysargyris, A. Treated Wastewater and Fertigation Applied for Greenhouse Tomato Cultivation Grown in Municipal Solid Waste Compost and Soil Mixtures. Sustainability 2020, 12, 4287. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Przygocka-Cyna, K.; Grzebisz, W. Effect of biofertilizers on nutrient uptake by vegetables grown in a short cropping sequence. J. Elem. 2018, 23, 807–823. [Google Scholar] [CrossRef]
- Souri, M.K.; Roemheld, V. Split daily application of ammonium cannot ameliorate ammonium toxicity in tomato plants. Hortic. Environ. Biotechnol. 2009, 50, 384–391. [Google Scholar]
- Rose, M.T.; Patti, A.F.; Little, K.R.; Brown, A.L.; Jackson, W.R.; Cavagnaro, T.R. A meta-analysis and review of plant-growth response to humic substances: Practical implications for agriculture. Adv. Agron. 2014, 124, 37–89. [Google Scholar]
- Denre, M.; Ghanti, S.; Sarkar, K. Effect of humic acid application on accumulation of mineral nutrition and pungency in garlic (Allium sativum L.). Int. J. Biotechnol. Mol. Biol. Res. 2014, 5, 7–12. [Google Scholar]
- Petropoulos, S.A.; Fernandes, Â.; Plexida, S.; Chrysargyris, A.; Tzortzakis, N.; Barreira, J.C.M.; Barros, L.; Ferreira, I.C. Biostimulants Application Alleviates Water Stress Effects on Yield and Chemical Composition of Greenhouse Green Bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Urban, J.; Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal conductance increases with rising temperature. Plant Signal. Behav. 2017, 12, e1356534. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Mou, B. Drench Application of Fish-derived Protein Hydrolysates Affects Lettuce Growth, Chlorophyll Content, and Gas Exchange. HortTechnology 2017, 27, 539–543. [Google Scholar] [CrossRef]
- Xu, C.; Mou, B. Chitosan as Soil Amendment Affects Lettuce Growth, Photochemical Efficiency, and Gas Exchange. HortTechnology 2018, 28, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Wozniak, E.; Blaszczak, A.; Wiatrak, P.; Canady, M. Biostimulant mode of action: Impact of biostimulant on cellular level. In the Chemical Biology of Plant Biostimulants; Geelen, D., Xu, L., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2020; pp. 229–243. [Google Scholar]
- Fanasca, S.; Colla, G.; Maiani, G.; Venneria, E.; Rouphael, Y.; Azzini, E.; Saccardo, F. Changes in Antioxidant Content of Tomato Fruits in Response to Cultivar and Nutrient Solution Composition. J. Agric. Food Chem. 2006, 54, 4319–4325. [Google Scholar] [CrossRef]
- Bramley, P.M. Regulation of carotenoid formation during tomato fruit ripening and development. J. Exp. Bot. 2002, 53, 2107–2113. [Google Scholar] [CrossRef] [Green Version]
- Tzortzakis, N.; Borland, A.M.; Singleton, I.; Barnes, J. Impact of atmospheric ozone-enrichment on quality-related attributes of tomato fruit. Postharvest Biol. Technol. 2007, 45, 317–325. [Google Scholar] [CrossRef]
- Guillén, F.; Castillo, S.; Zapata, P.J.; Martínez-Romero, D.; Serrano, M.; Valero, D. Efficacy of 1-MCP treatment in tomato fruit. 1. Duration and concentration of 1-MCP treatment to gain an effective delay of postharvest ripening. Postharvest Biol. Technol. 2007, 43, 23–27. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Growth | Control | CP | EP-1x | EP-3x |
|---|---|---|---|---|
| Height (m) | 1.47 ± 0.03b | 1.59 ± 0.05ab | 1.75 ± 0.05a | 1.63 ± 0.09ab |
| Leaf number | 28.33 ± 0.42a | 30.00 ± 1.29a | 32.17 ± 1.25a | 29.00 ± 1.65a |
| Plant biomass (g) | 433.90 ± 30.39a | 510.75 ± 41.27a | 524.48 ± 70.38a | 421.33 ± 42.99a |
| Plant dry weight (g) | 83.88 ± 3.98b | 107.38 ± 6.41a | 93.43 ± 8.15ab | 82.93 ± 5.86b |
| Yield (kg plant−1) | 0.58 ± 0.06b | 0.74 ± 0.05ab | 0.71 ± 0.06ab | 0.79 ± 0.06a |
| Fruit number | 9.58 ± 0.89a | 9.46 ± 0.63a | 9.10 ± 0.79a | 10.00 ± 0.41a |
| Blossom end rot (BER) (%) | 4.13 ± 2.80a | 6.59 ± 2.16a | 0.00 ± 0.00b | 4.20 ± 2.03a |
| Fruit cracking (%) | 8.23 ± 2.06b | 7.97 ± 2.68b | 12.77 ± 3.05ab | 18.43 ± 2.688a |
| Plant Physiology | Control | CP | EP-1x | EP-3x |
|---|---|---|---|---|
| Stomatal conductance (s cm−1) | 1.18 ± 0.16b | 1.56 ± 0.13ab | 1.81 ± 0.21a | 1.59 ± 0.21ab |
| Chlorophyll fluorescence (Fv/Fm) | 0.82 ± 0.01a | 0.83 ± 0.00a | 0.82 ± 0.01a | 0.83 ± 0.00a |
| Chlorophyll a (mg g−1 Fw) | 0.31 ± 0.07b | 0.29 ± 0.03b | 0.65 ± 0.06a | 0.25 ± 0.03b |
| Chlorophyll b (mg g−1 Fw) | 0.08 ± 0.02b | 0.07 ± 0.01b | 0.17 ± 0.02a | 0.06 ± 0.01b |
| Total Chlorophylls (mg g−1 Fw) | 0.39 ± 0.09b | 0.37 ± 0.03b | 0.82 ± 0.08a | 0.31 ± 0.04b |
| Quality Attributes | Control | CP | EP-1x | EP-3x |
|---|---|---|---|---|
| Fruit fresh weight-Fw (g) | 80.93 ± 3.4a | 94.4 ± 5.36a | 95.22 ± 6.65a | 81.49 ± 4.69a |
| Firmness (Newton) | 12.75 ± 0.97a | 10.27 ± 0.32b | 10.21 ± 0.54b | 10.74 ± 0.75ab |
| Total soluble solids-TSS (°Brix) | 5.30 ± 0.26a | 5.10 ± 0.18a | 5.08 ± 0.14a | 5.08 ± 0.22a |
| Titratable acidity-TA (citric acid g L−1) | 5.26 ± 0.24a | 4.14 ± 0.11b | 3.58 ± 0.12c | 3.95 ± 0.16bc |
| TSS/TA | 1.01 ± 0.04c | 1.23 ± 0.02b | 1.42 ± 0.03a | 1.28 ± 0.03b |
| Ascorbic acid-AA (mg 100 g−1 Fw) | 73.80 ± 3.71b | 79.87 ± 8.11b | 147.02 ± 4.66a | 85.21 ± 3.15b |
| Phenols (μmol GAE g−1 Fw) | 3.10 ± 0.20a | 2.59 ± 0.20b | 2.51 ± 0.04b | 2.51 ± 0.11b |
| Marketability (1–10) | 7.83 ± 0.49a | 8.58 ± 0.40a | 8.67 ± 0.17a | 7.75 ± 0.31a |
| Aroma (1–10) | 5.50 ± 0.53a | 7.00 ± 0.41a | 6.75 ± 0.28a | 6.33 ± 0.69a |
| Appearance (1–10) | 7.25 ± 0.54bc | 8.25 ± 0.40ab | 8.50 ± 0.26a | 7.00 ± 0.26c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrysargyris, A.; Charalambous, S.; Xylia, P.; Litskas, V.; Stavrinides, M.; Tzortzakis, N. Assessing the Biostimulant Effects of a Novel Plant-Based Formulation on Tomato Crop. Sustainability 2020, 12, 8432. https://doi.org/10.3390/su12208432
Chrysargyris A, Charalambous S, Xylia P, Litskas V, Stavrinides M, Tzortzakis N. Assessing the Biostimulant Effects of a Novel Plant-Based Formulation on Tomato Crop. Sustainability. 2020; 12(20):8432. https://doi.org/10.3390/su12208432
Chicago/Turabian StyleChrysargyris, Antonios, Savvas Charalambous, Panayiota Xylia, Vassilis Litskas, Menelaos Stavrinides, and Nikos Tzortzakis. 2020. "Assessing the Biostimulant Effects of a Novel Plant-Based Formulation on Tomato Crop" Sustainability 12, no. 20: 8432. https://doi.org/10.3390/su12208432