
Alterations of Amino Acid Concentrations and Photosynthetic Indices in Light Irradiated Arabidopsis thaliana during Phytoextraction

Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Plant
2.2. Light Irradiation Treatment
2.3. Cd Analysis and Quality Control
2.4. Measurement of Chlorophyll Fluorescence
2.5. Measurement of MDA, H2O2
2.6. Measurement of Amino Acids
2.7. Statistical Analysis
3. Results and Discussion
3.1. Plant Growth
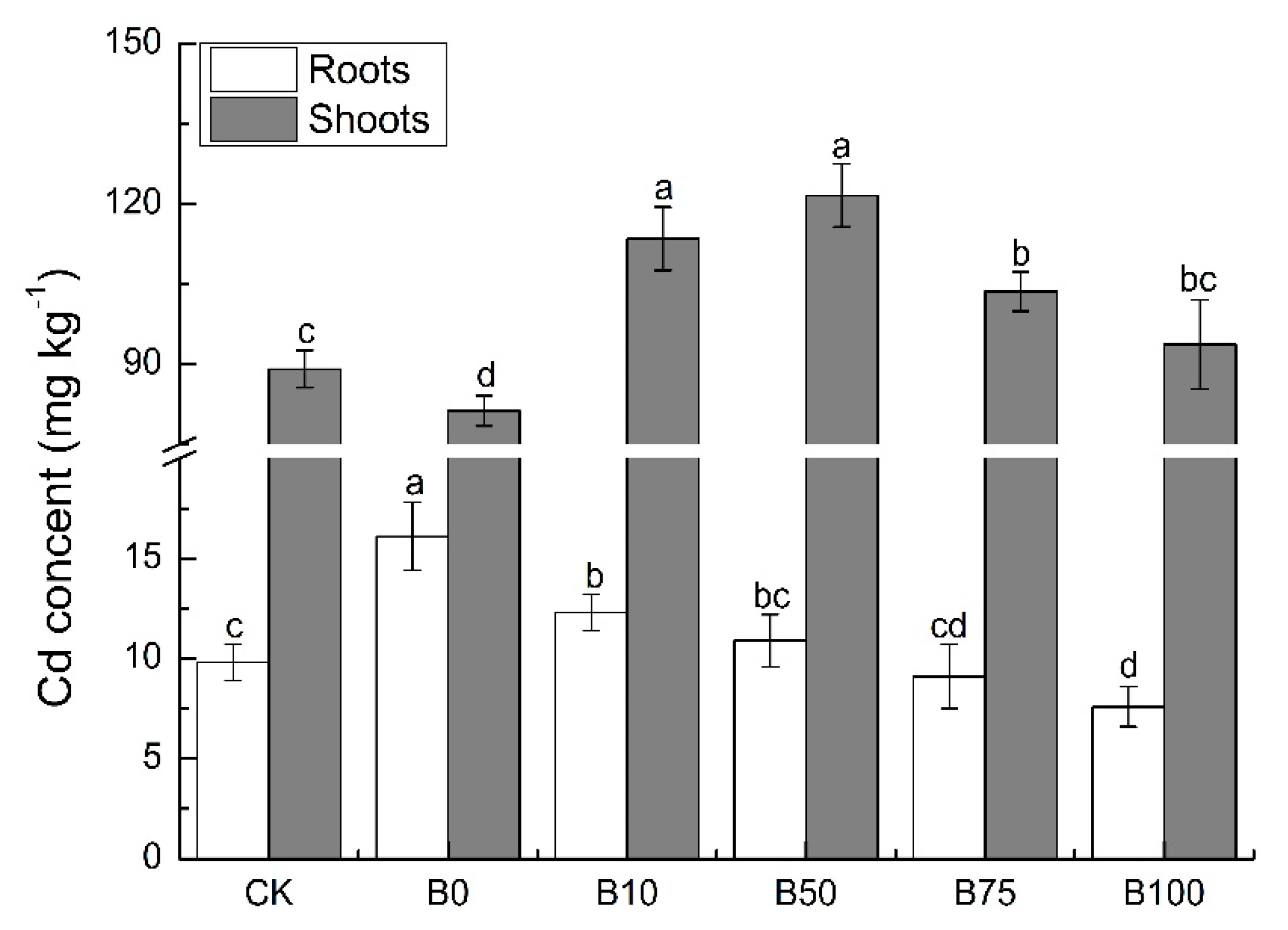
3.2. Cd Content
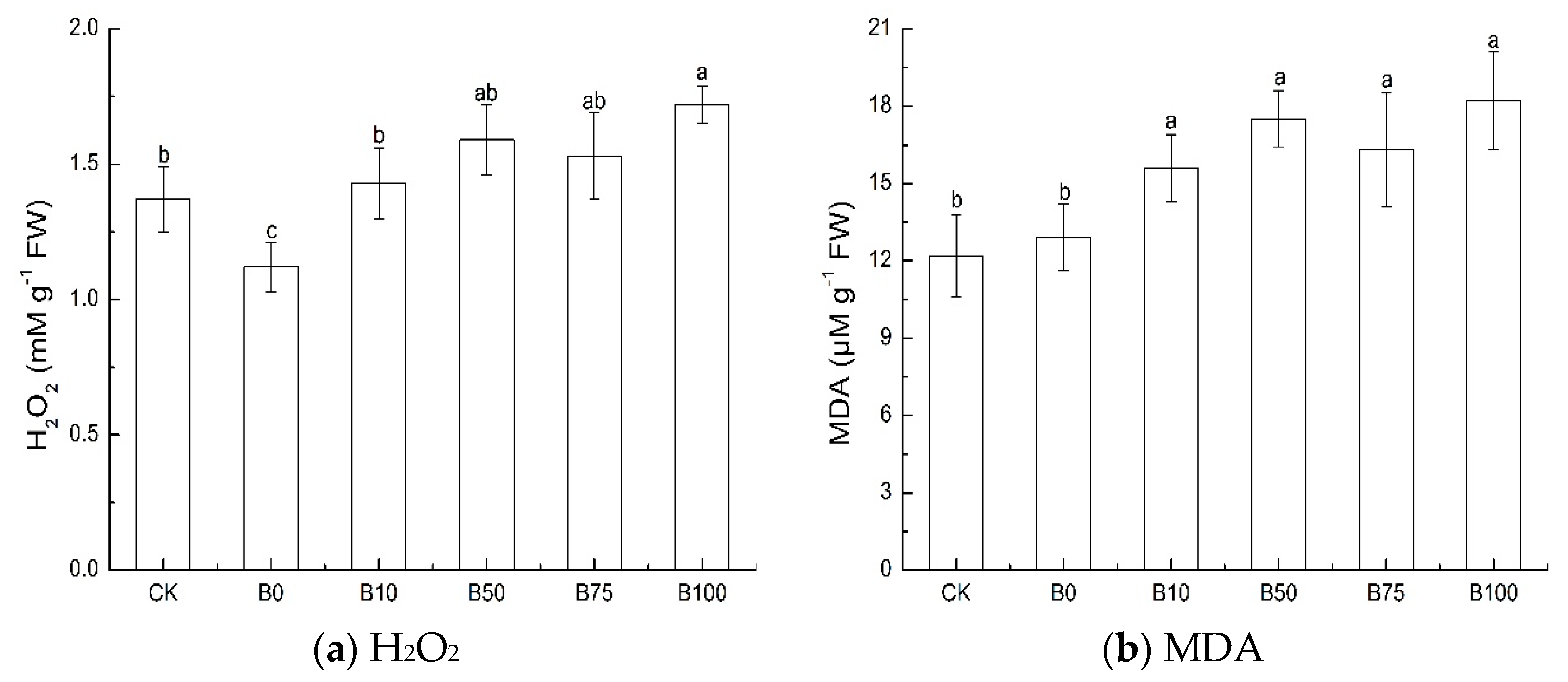
3.3. MDA and H2O2 Content
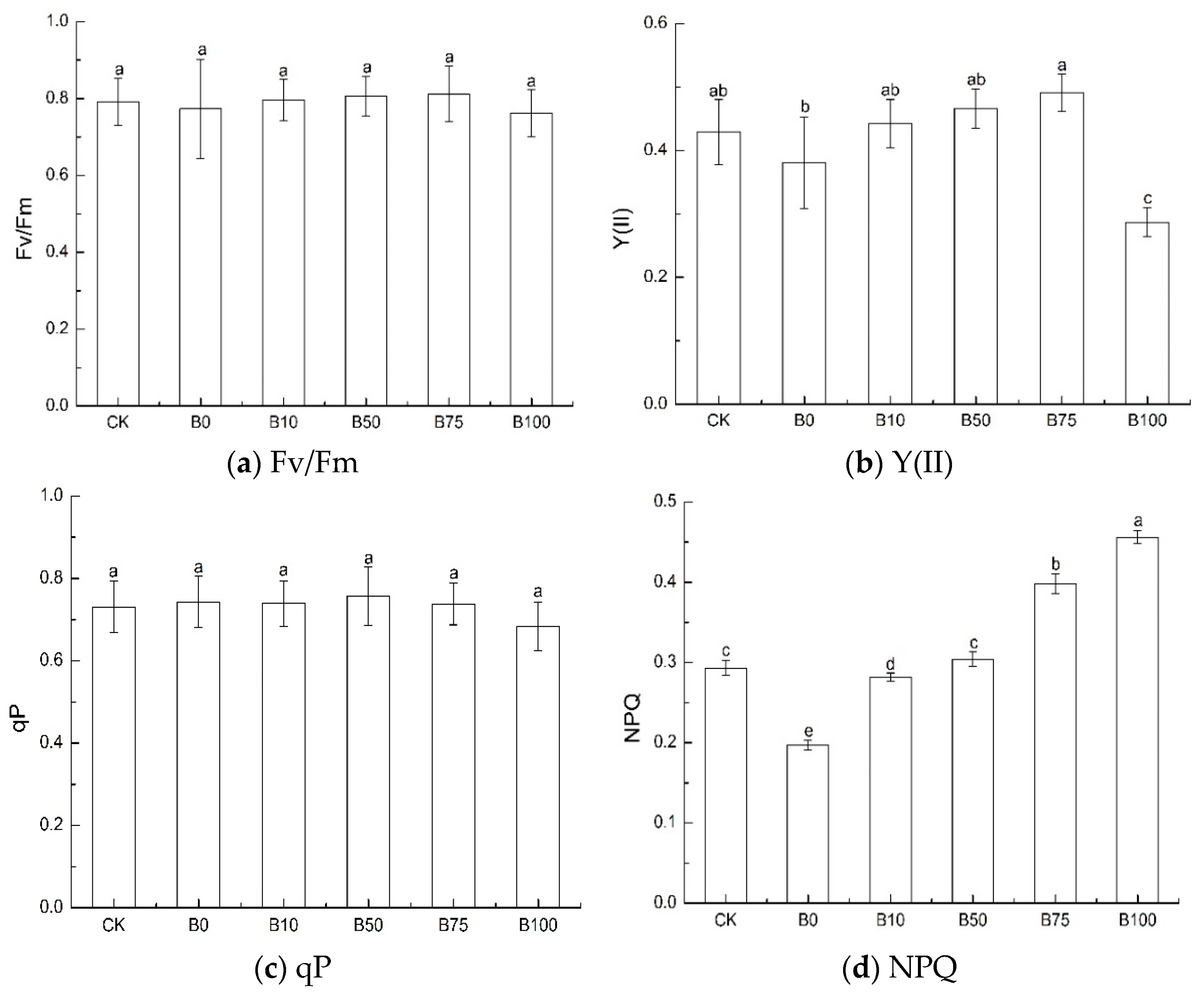
3.4. Chlorophyll Fluorescence Characteristic
3.5. Amino Acid
3.6. Phytoremediation Efficiency
4. Discussion
4.1. Responses of Biomass Yield and Cd Accumulation to Light Irradiation
4.2. Influences of Light Irradiation on Cd Accumulation
4.3. Relationship among Oxidative Damage, Photosynthetic Indices, and Amino Acid Content
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric acid assisted phytoremediation of cadmium by Brassica napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef]
- Gallego, S.M.; Pena, L.B.; Barcia, R.A.; Azpilicueta, C.E.; Iannone, M.F.; Rosales, E.P.; Zawoznik, M.S.; Groppa, M.D.; Benavides, M.P. Unravelling cadmium toxicity and tolerance in plants: Insight into regulatory mechanisms. Environ. Exp. Bot. 2012, 83, 33–46. [Google Scholar] [CrossRef]
- Wu, Q.; Du, Y.; Huang, Z.; Gu, J.; Leung, J.Y.S.; Mai, B.; Xiao, T.; Liu, W.; Fu, J. Vertical profile of soil/sediment pollution and microbial community change by e-waste recycling operation. Sci. Total Environ. 2019, 669, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Xu, X.; Zhang, Y.; Cong, X.; Lu, X.; Huo, X. Elevated lead levels from e-waste exposure are linked to sensory integration difficulties in preschool children. Neuro Toxicol. 2019, 71, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Mushtaq, H.; Ali, H.; Munis, M.F.H.; Javed, M.T.; Chaudhary, H.J. Diazotrophs-assisted phytoremediation of heavy metals: A novel approach. Environ. Sci. Pollut. Res. 2015, 22, 2505–2514. [Google Scholar] [CrossRef] [PubMed]
- Niñerola, V.B.; Navarro-Pedreño, J.; Lucas, I.G.; Pastor, I.M.; Vidal, M.M.J. Geostatistical assessment of soil salinity and cropping systems used as soil phytoremediation strategy. J. Geochem. Explor. 2017, 174, 53–58. [Google Scholar] [CrossRef]
- Guo, Q.; Meng, L.; Zhang, Y.N.; Mao, P.C.; Tian, X.X.; Li, S.S.; Zhang, L. Antioxidative systems, metal ion homeostasis and cadmium distribution in Iris lactea exposed to cadmium stress. Ecotoxicol. Environ. Saf. 2017, 139, 50–55. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.; Goltsev, V. Effects of different metals on photosynthesis: Cadmium and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [Green Version]
- Arena, C.; Figlioli, F.; Sorrentino, M.C.; Izzo, L.G.; Capozzi, F.; Giordano, S.; Spagnuolo, V. Ultrastructural, protein and photosynthetic alterations induced by Pb and Cd in Cynara cardunculus L., and its potential for phytoremediation. Ecotoxicol. Environ. Saf. 2017, 145, 83–89. [Google Scholar] [CrossRef]
- Gupta, D.K.; Tripathi, R.D.; Rai, U.N.; Dwivedi, S.; Mishra, S.; Srivastava, S.; Inouhe, M. Changes in amino acid profile and metal content in seeds of Cicer arietinum L. (chickpea) grown under various fly-ash amendments. Chemosphere 2006, 65, 939–945. [Google Scholar] [CrossRef]
- Lanoue, J.; Leonardos, E.D.; Grodzinski, B. Effects of Light Quality and Intensity on Diurnal Patterns and Rates of Photo-Assimilate Translocation and Transpiration in Tomato Leaves. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Xiao, H.; Guo, Q.; Zhang, Z.; Zhao, J.; Yang, D. Effects of cadmium stress on growth and amino acid metabolism in two Compositae plants. Ecotoxicol. Environ. Saf. 2018, 158, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, A.; Peng, S.; Wei, Z.; Liu, G. Identification and application of amino acids as chelators in phytoremediation of rare earth elements lanthanum and yttrium. Plant Soil 2013, 373, 329–338. [Google Scholar] [CrossRef]
- Bayçu, G.; Moustaka, J.; Gevrek, N.; Moustakas, M. Chlorophyll Fluorescence Imaging Analysis for Elucidating the Mechanism of Photosystem II Acclimation to Cadmium Exposure in the Hyperaccumulating Plant Noccaea caerulescens. Materials 2018, 11, 2580. [Google Scholar] [CrossRef] [Green Version]
- Jan, L.; Fefer, D.; Košmelj, K.; Gaberščik, A.; Jerman, I. Geomagnetic and strong static magnetic field effects on growth and chlorophyll a fluorescence in Lemna minor. Bioelectromagnetics 2015, 36, 190–203. [Google Scholar] [CrossRef]
- Tripathi, R.D.; Singh, R.; Tripathi, P.; Dwivedi, S.; Chauhan, R.; Adhikari, B.; Trivedi, P.K. Arsenic accumulation and tolerance in rootless macrophyte Najas indica are mediated through antioxidants, amino acids and phytochelatins. Aquat. Toxicol. 2014, 157, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zou, D.; Zhu, M.; Yang, Y. Effects of CO2 levels and light intensities on growth and amino acid contents in red seaweed Gracilaria lemaneiformis. Aquac. Res. 2017, 48, 2683–2690. [Google Scholar] [CrossRef]
- Jeong, B.R.; Sivanesan, I. Impact of light quality and sucrose on adventitious shoot regeneration and bioactive compound accumulation in Ajuga multiflora Bunge. Sci. Hortic. 2018, 236, 222–228. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Zou, T.; Wu, B.; Wu, W.; Ge, L.; Xu, Y. Effects of different spectra from LED on the growth, development and reproduction of Arabidopsis Thaliana. Phyton 2020, 89, 275–289. [Google Scholar] [CrossRef]
- Luo, J.; Cao, M.; Zhang, C.; Wu, J.; Gu, X.W.S. The in fluence of light combination on the physicochemical characteristics and enzymatic activity of soil with multi-metal pollution in phytoremediation. J. Hazard. Mater. 2020, 393, 1–10. [Google Scholar] [CrossRef]
- Vandenbrink, J.P.; Herranz, R.; Medina, F.J.; Edelmann, R.E.; Kiss, J.Z. A novel blue-light phototropic response is revealed in roots of Arabidopsis thaliana in microgravity. Planta 2016, 244, 1201–1215. [Google Scholar] [CrossRef]
- Lin, Y.F.; Hassan, Z.; Talukdar, S.; Schat, H.; Aarts, M.G.M. Expression of the Znt1 zinc transporter from the metal hyperaccumulator noccaea caerulescens confers enhanced zinc and cadmium tolerance and accumulation to Arabidopsis thaliana. PLoS ONE 2016, 11, e0149750. [Google Scholar] [CrossRef] [PubMed]
- Szopiński, M.; Sitko, K.; Gieroń, Ż.; Rusinowski, S.; Corso, M.; Hermans, C.; Verbruggen, N.; Małkowski, E. Toxic effects of cd and zn on the photosynthetic apparatus of the Arabidopsis halleri and Arabidopsis arenosa pseudo-metallophytes. Front. Plant Sci. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ok, Y.S.; Kim, S.C.; Kim, D.K.; Skousen, J.G.; Lee, J.S.; Cheong, Y.W.; Kim, S.J.; Yang, J.E. Ameliorants to immobilize Cd in rice paddy soils contaminated by abandoned metal mines in Korea. Environ. Geochem. Health 2011, 33, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, W.; Yang, T.; Chen, Y.; Ni, W. Effects of Cadmium Stress on Leaf Chlorophyll Fluorescence and Photosynthesis of Elsholtzia argyi—A Cadmium Accumulating Plant. Int. J. Phytoremed. 2015, 17, 85–92. [Google Scholar] [CrossRef]
- Agrawal, D.; Allakhverdiev, S.I.; Jajoo, A. Cyclic electron flow plays an important role in protection of spinach leaves under high temperature stress. Физиoлoгия Растений 2016, 63, 225–230. [Google Scholar] [CrossRef]
- Zhang, X.H.; Liu, Y.H.; Liu, B.W.; Liu, Q.; Wen, S.Y.; Ao, B.; Lin, Z.Q.; Zheng, Y.L.; Yang, W.Z.; Chu, X.T.; et al. Arbuscular mycorrhiza fungus improved growth, antioxidant defense, and endogenous hormones in tall fescue under low-light stress. South Afr. J. Bot. 2019, 127, 43–50. [Google Scholar] [CrossRef]
- Xu, Y.; Xiao, H. Free amino acid concentrations and nitrogen isotope signatures in Pinus massoniana (Lamb.) needles of different ages for indicating atmospheric nitrogen deposition. Environ. Pollution. 2017, 221, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Tlustoš, P.; Břendová, K.; Száková, J.; Najmanová, J.; Koubová, K. The long-term variation of Cd and Zn hyperaccumulation by Noccaea spp and Arabidopsis halleri plants in both pot and field conditions. Int. J. Phytoremed. 2016, 18, 110–115. [Google Scholar] [CrossRef]
- McGrath, S.P.; Lombi, E.; Gray, C.W.; Caille, N.; Dunham, S.J.; Zhao, F.J. Field evaluation of Cd and Zn phytoextraction potential by the hyperaccumulators Thlaspi caerulescens and Arabidopsis halleri. Environ. Pollut. 2006, 141, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Gurajala, H.K.; Wu, L.H.; Van Der Ent, A.; Qiu, R.L.; Baker, A.J.M.; Tang, Y.T.; Yang, X.E.; Shu, W.S. Hyperaccumulator Plants from China: A Synthesis of the Current State of Knowledge. Environ. Sci. Technol. 2018, 52, 11980–11994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewes, L.J.; Stolpe, C.; Kerim, A.; Krämer, U.; Müller, C. Metal hyperaccumulation in the Brassicaceae species Arabidopsis halleri reduces camalexin induction after fungal pathogen attack. Environ. Exp. Bot. 2018, 153, 120–126. [Google Scholar] [CrossRef]
- Aghababaei, F.; Raiesi, F.; Hosseinpur, A. The significant contribution of mycorrhizal fungi and earthworms to maize protection and phytoremediation in Cd-polluted soils. Pedobiologia 2014, 57, 223–233. [Google Scholar] [CrossRef]
- Heber, U.; Bukhov, N.G.; Shuvalov, V.A.; Kobayashi, Y.; Lange, O.L. Protection of the photosynthetic apparatus against damage by excessive illumination in homoiohydric leaves and poikilohydric mosses and lichens. J. Exp. Bot. 2001, 52, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; Ai, H.; Xu, X.; Chen, K.; Niu, H.; Zhu, H.; Sun, J.; Du, D.; Chen, L. Nitric oxide alleviates toxicity of hexavalent chromium on tall fescue and improves performance of photosystem II. Ecotoxicol. Environ. Saf. 2018, 164, 32–40. [Google Scholar] [CrossRef]
- Fatima, A.; Singh, A.A.; Mukherjee, A.; Agrawal, M.; Agrawal, S.B. Variability in defence mechanism operating in three wheat cultivars having different levels of sensitivity against elevated ozone. Environ. Exp. Bot. 2018, 155, 66–78. [Google Scholar] [CrossRef]
- Semer, J.; Navrátil, M.; Špunda, V.; Štroch, M. Chlorophyll fluorescence parameters to assess utilization of excitation energy in photosystem II independently of changes in leaf absorption. J. Photochem. Photobiol. B Biol. 2019, 197, 1–11. [Google Scholar] [CrossRef]
- Pietrini, F.; Iori, V.; Beone, T.; Mirabile, D.; Zacchini, M. Effects of a ladle furnace slag added to soil on morpho-physiological and biochemical parameters of Amaranthus paniculatus L. plants. J. Hazard. Mater. 2017, 329, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Li, P.; Shang, B.; Liu, S.; Yang, A.; Wang, Y.; Feng, Z. Differential responses of peach (Prunus persica) seedlings to elevated ozone are related with leaf mass per area, antioxidant enzymes activity rather than stomatal conductance. Environ. Pollut. 2017, 227, 380–388. [Google Scholar] [CrossRef]
- Fu, W.; Liu, D.; Yin, Q.; Wu, Y.; Li, P. Suspended sediment in tidal currents: An often-neglected pollutant that aggravates mangrove degradation. Mar. Pollut. Bull. 2014, 84, 164–171. [Google Scholar] [CrossRef]
- Kaur, P.; Bali, S.; Sharma, A.; Vig, A.P.; Bhardwaj, R. Effect of earthworms on growth, photosynthetic efficiency and metal uptake in Brassica juncea L. plants grown in cadmium-polluted soils. Environ. Sci. Pollut. Res. 2017, 24, 13452–13465. [Google Scholar] [CrossRef] [PubMed]
- Kovács, H.; Aleksza, D.; Baba, A.I.; Hajdu, A.; Király, A.M.; Zsigmond, L.; Tóth, S.Z.; Kozma-Bognár, L.; Szabados, L. Light Control of Salt-Induced Proline Accumulation Is Mediated by ELONGATED HYPOCOTYL 5 in Arabidopsis. Front. Plant Sci. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Growth, Photosynthetic Characteristics, Antioxidant Capacity and Biomass Yield and Quality of Wheat (Triticum aestivum L.) Exposed to LED Light Sources with Different Spectra Combinations. J. Agron. Crop Sci. 2014, 200, 219–230. [Google Scholar] [CrossRef]
- Luo, J.; He, W. Sensitivity of Eucalyptus globulus to red and blue light with different combinations and their influence on its efficacy for contaminated soil phytoremediation. J. Environ. Manag. 2019, 241, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Rainbow, P.S. Subcellular partitioning and the prediction of cadmium toxicity to aquatic organisms. Environ. Chem. 2006, 3, 395–399. [Google Scholar] [CrossRef]
- Kwon, H.K.; Jeon, J.Y.; Oh, S.J. Potential for heavy metal (copper and zinc) removal from contaminated marine sediments using microalgae and light emitting diodes. Ocean Sci. J. 2017, 52, 57–66. [Google Scholar] [CrossRef]
- Wan, X.M.; Lei, M.; Chen, T.B.; Yang, J.X.; Liu, H.T.; Chen, Y. Role of transpiration in arsenic accumulation of hyperaccumulator Pteris vittata L. Environ. Sci. Pollut. Res. 2015, 22, 16631–16639. [Google Scholar] [CrossRef]
- Žaltauskaite, J.; Sodiene, I. Effects of cadmium and lead on the life-cycle parameters of juvenile earthworm Eisenia fetida. Ecotoxicol. Environ. Saf. 2014, 103, 9–16. [Google Scholar] [CrossRef]
- Lin, R.; Wang, X.; Luo, Y.; Du, W.; Guo, H.; Yin, D. Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings (Triticum aestivum L.). Chemosphere 2007, 69, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Islam, E.; Irem, S.; Akhtar, K.; Ashraf, M.Y.; Iqbal, J.; Liu, D. Pb-induced phytotoxicity in para grass (Brachiaria mutica) and Castorbean (Ricinus communis L.): Antioxidant and ultrastructural studies. Chemosphere 2018, 200, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Luo, Q.; Liu, S.; Zhao, Y.; Long, Y.; Pan, Y. Screening ornamental plants to identify potential Cd hyperaccumulators for bioremediation. Ecotoxicol. Environ. Saf. 2018, 162, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Blomster, T.; Salojärvi, J.; Sipari, N.; Brosché, M.; Ahlfors, R.; Keinänen, M.; Overmyer, K.; Kangasjärvi, J. Apoplastic reactive oxygen species transiently decrease auxin signaling and cause stress-induced morphogenic response in Arabidopsis. Plant Physiol. 2011, 157, 1866–1883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.F.; Gao, L.M.; Han, R. He–Ne laser illumination ameliorates photochemical impairment in ultraviolet-B stressed-wheat seedlings via detoxifying ROS cytotoxicity. Russ. J. Plant Physiol. 2017, 64, 766–775. [Google Scholar] [CrossRef]
- Konishi, N.; Ishiyama, K.; Matsuoka, K.; Maru, I.; Hayakawa, T.; Yamaya, T.; Kojima, S. NADH-dependent glutamate synthase plays a crucial role in assimilating ammonium in the Arabidopsis root. Physiol. Plant. 2014, 152, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Krishnani, K.K.; Singh, N.P. Oxidative and cellular stress as bioindicators for metal contamination in freshwater mollusk Lamellidens marginalis. Environ. Sci. Pollut. Res. 2017, 24, 16137–16147. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glu | Gln | Asp | Asn | Pro | |
|---|---|---|---|---|---|
| CK | 973 ± 11 a | 2331 ± 161 b | 1012 ± 89 cd | 3129 ± 73 c | 363 ± 12 d |
| B0 | 981 ± 31 a | 2591 ± 87 a | 1087 ± 76 c | 3256 ± 161 bc | 337 ± 13 e |
| B10 | 902 ± 20 b | 2234 ± 107 b | 1352 ± 119 ab | 3579 ± 113 a | 387 ± 11 c |
| B50 | 863 ± 19 c | 1981 ± 126 c | 1434 ± 56 a | 3432 ± 95 ab | 412 ± 9 b |
| B75 | 817 ± 17 d | 1862 ± 89 c | 1223 ± 89 bc | 3316 ± 63 b | 391 ± 7 c |
| B100 | 726 ± 29 e | 1573 ± 102 d | 932 ± 51 d | 2671 ± 89 d | 439 ± 13 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Liang, L.; Chen, D.; Gan, T.; Cao, M.; Luo, J. Alterations of Amino Acid Concentrations and Photosynthetic Indices in Light Irradiated Arabidopsis thaliana during Phytoextraction. Sustainability 2021, 13, 7720. https://doi.org/10.3390/su13147720
Chen Y, Liang L, Chen D, Gan T, Cao M, Luo J. Alterations of Amino Acid Concentrations and Photosynthetic Indices in Light Irradiated Arabidopsis thaliana during Phytoextraction. Sustainability. 2021; 13(14):7720. https://doi.org/10.3390/su13147720
Chicago/Turabian StyleChen, Yanmei, Longfei Liang, Dan Chen, Tian Gan, Min Cao, and Jie Luo. 2021. "Alterations of Amino Acid Concentrations and Photosynthetic Indices in Light Irradiated Arabidopsis thaliana during Phytoextraction" Sustainability 13, no. 14: 7720. https://doi.org/10.3390/su13147720