
The Conservational State of Coastal Ecosystems on the Mexican Caribbean Coast: Environmental Guidelines for Their Management

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Coastal Ecosystems
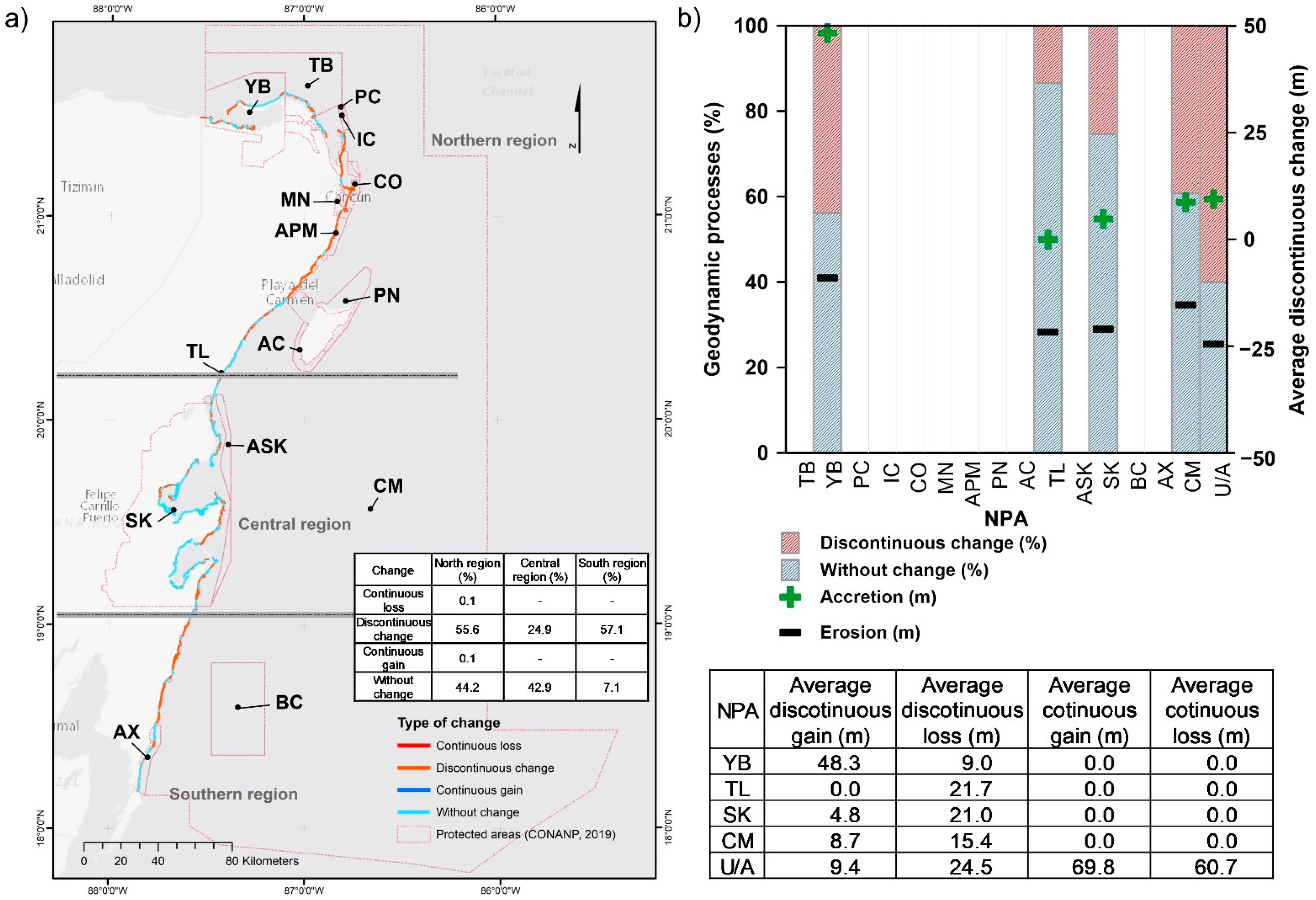
Coastline Evolution
2.2. Marine Climate
2.2.1. Waves
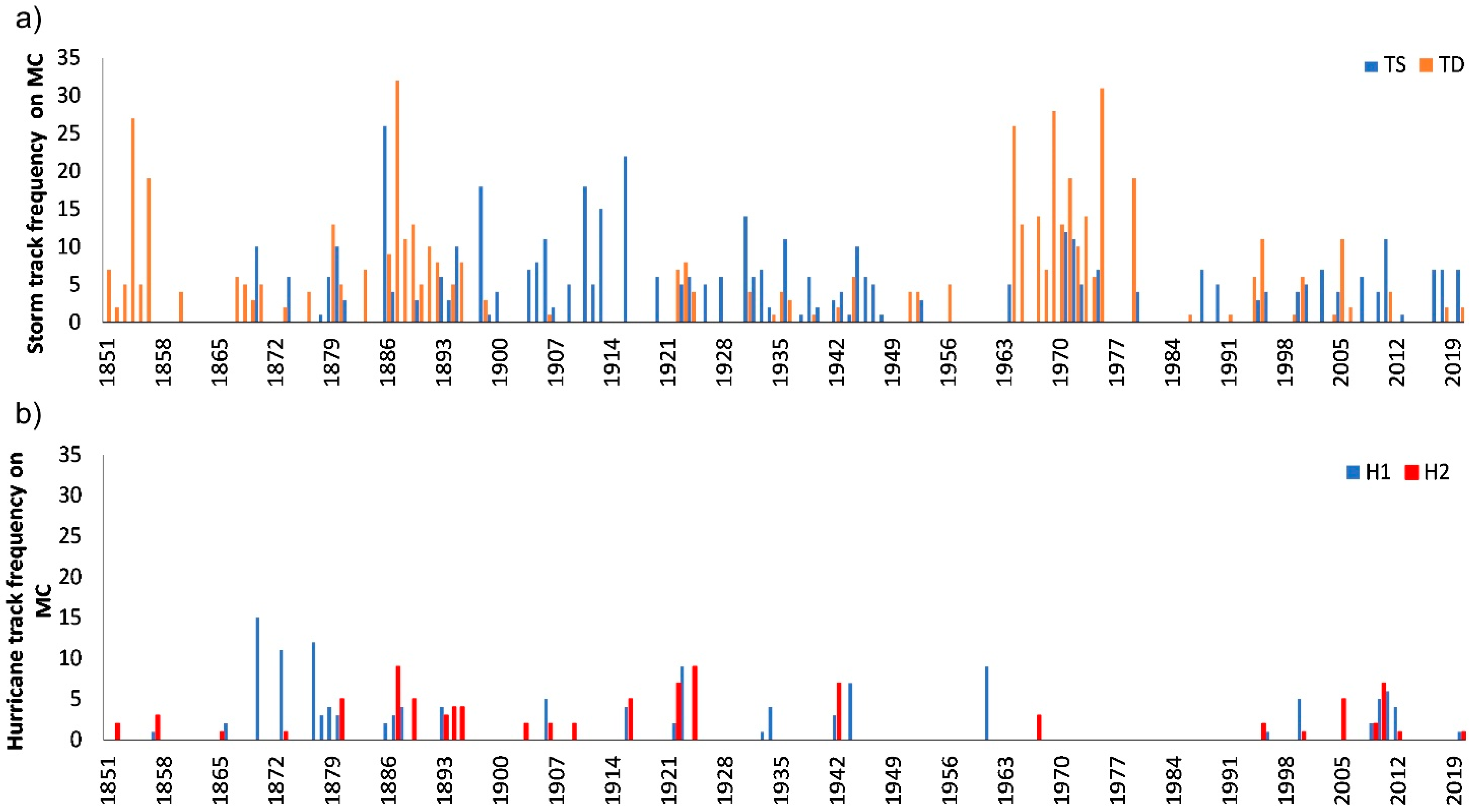
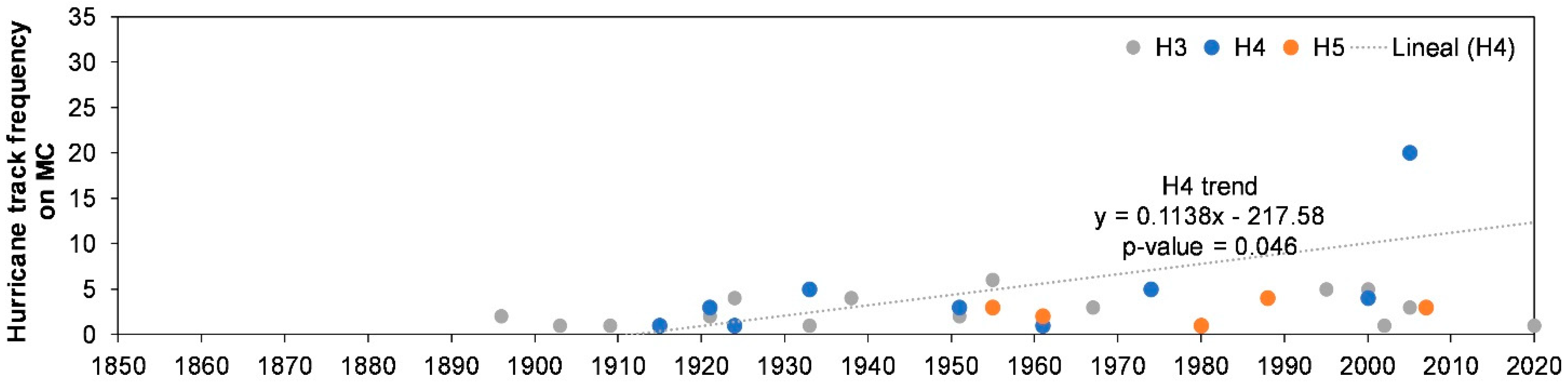
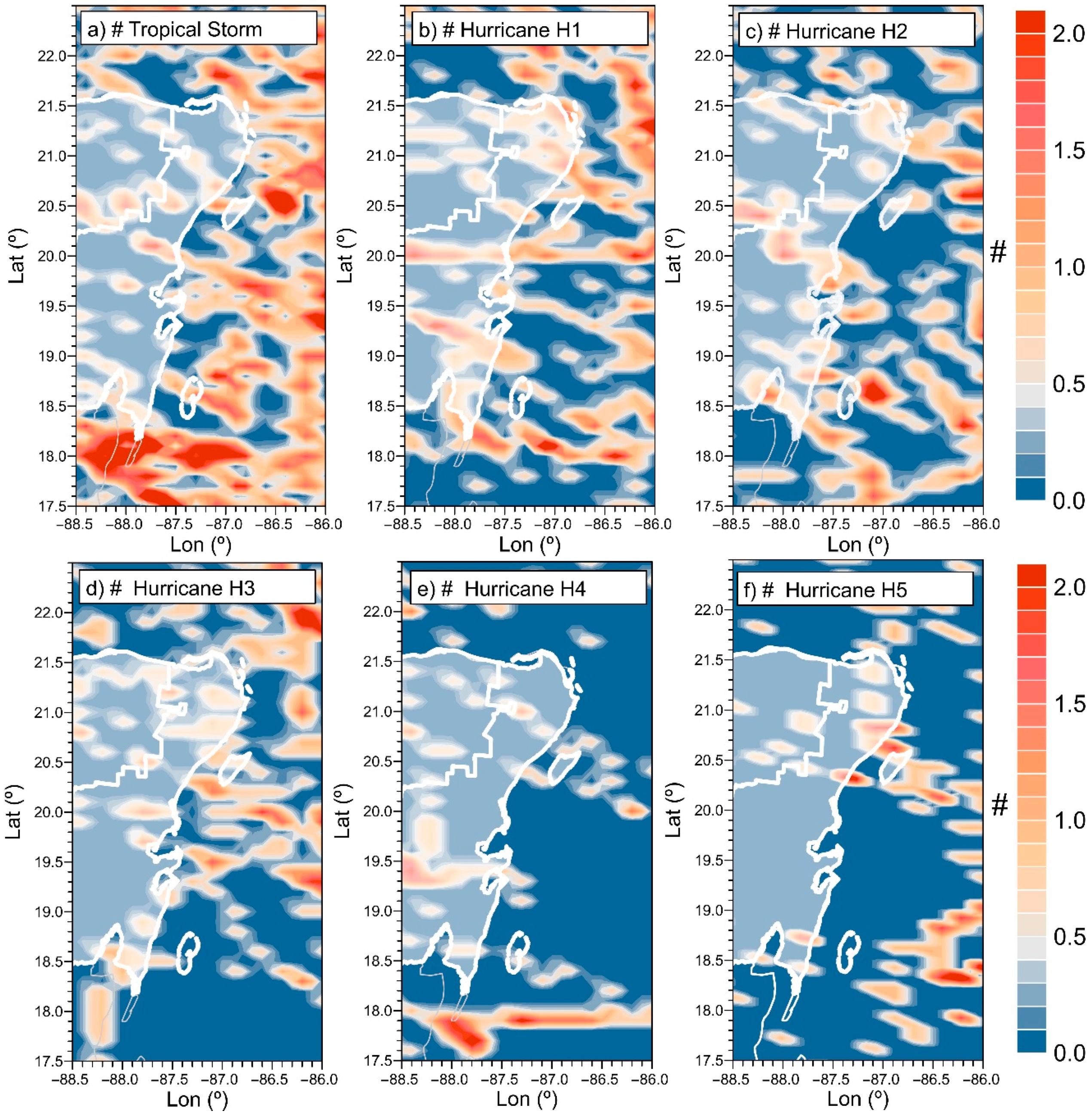
2.2.2. Hurricanes
2.3. Water Quality
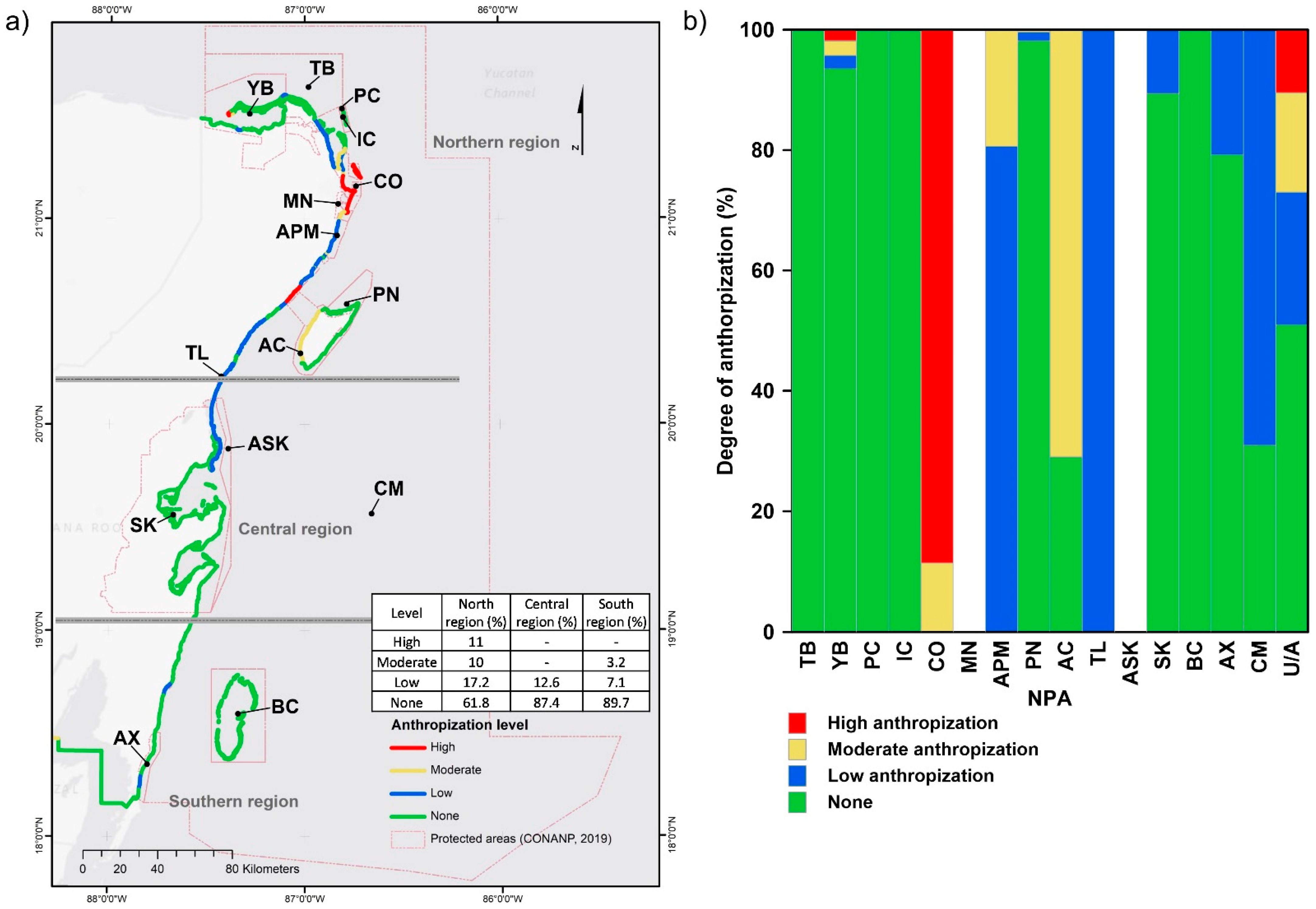
2.4. Anthropization
3. Results
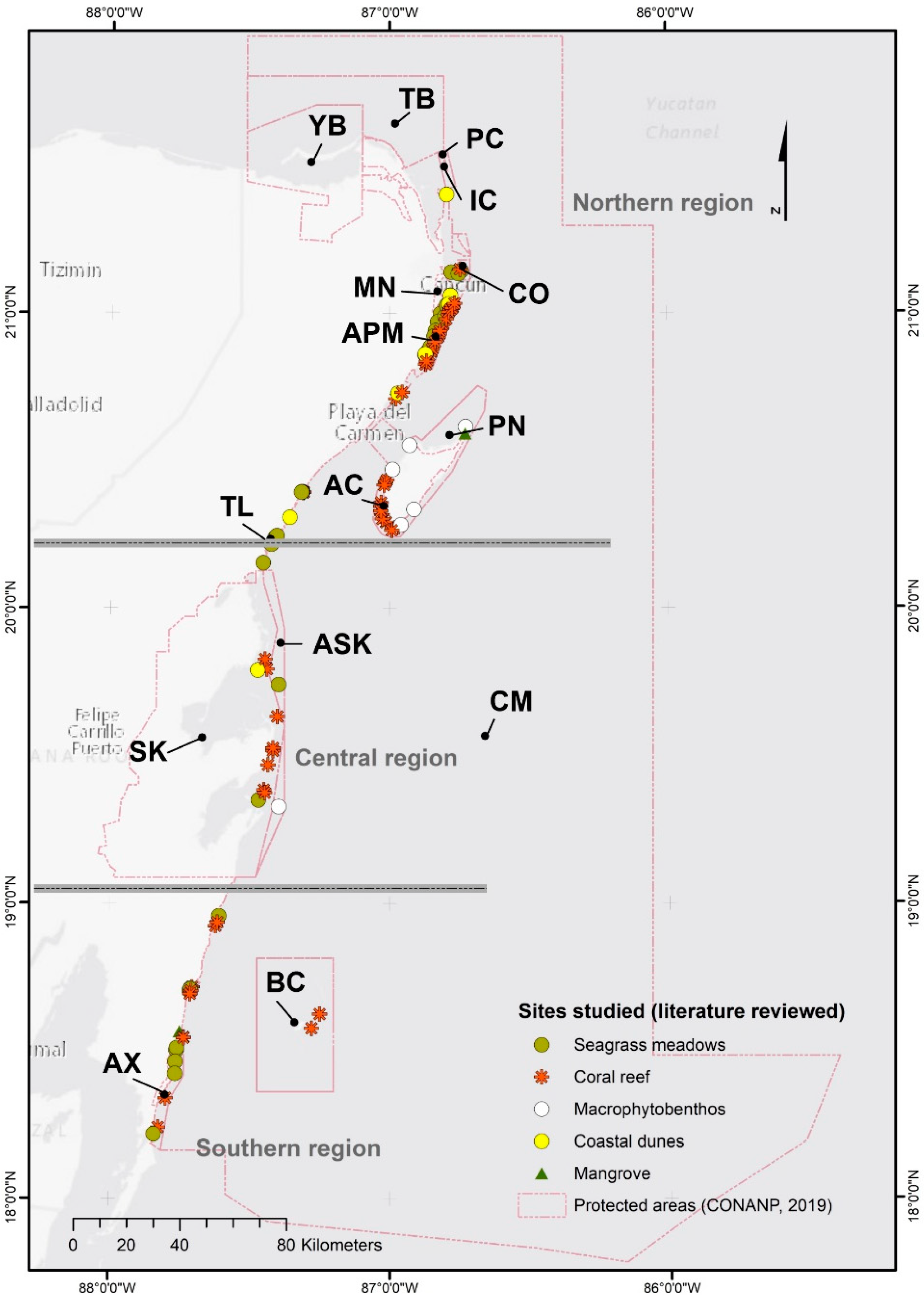
3.1. Coastal Ecosystems
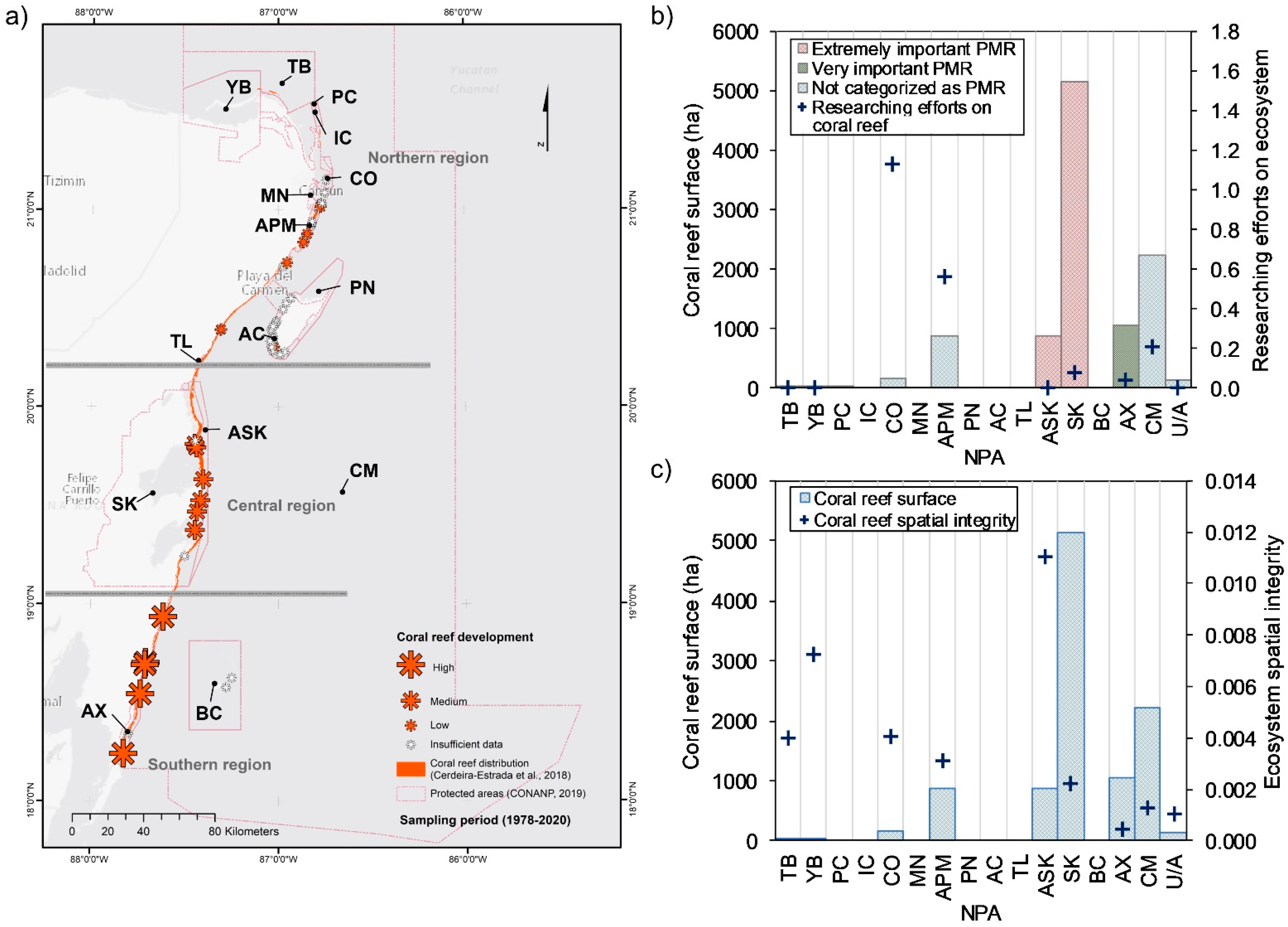
3.1.1. Coral Reefs
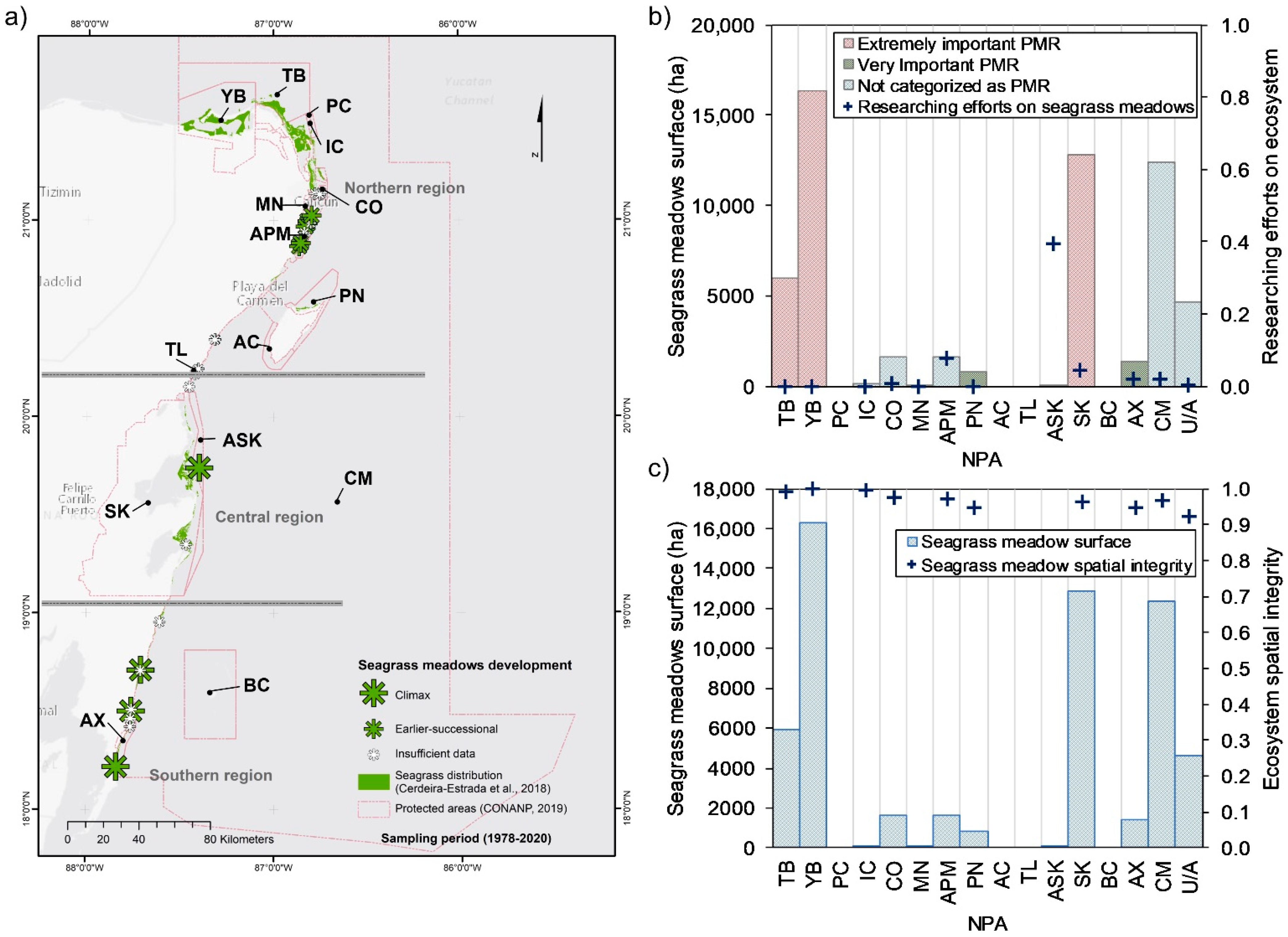
3.1.2. Seagrass Meadows
3.1.3. Mangrove
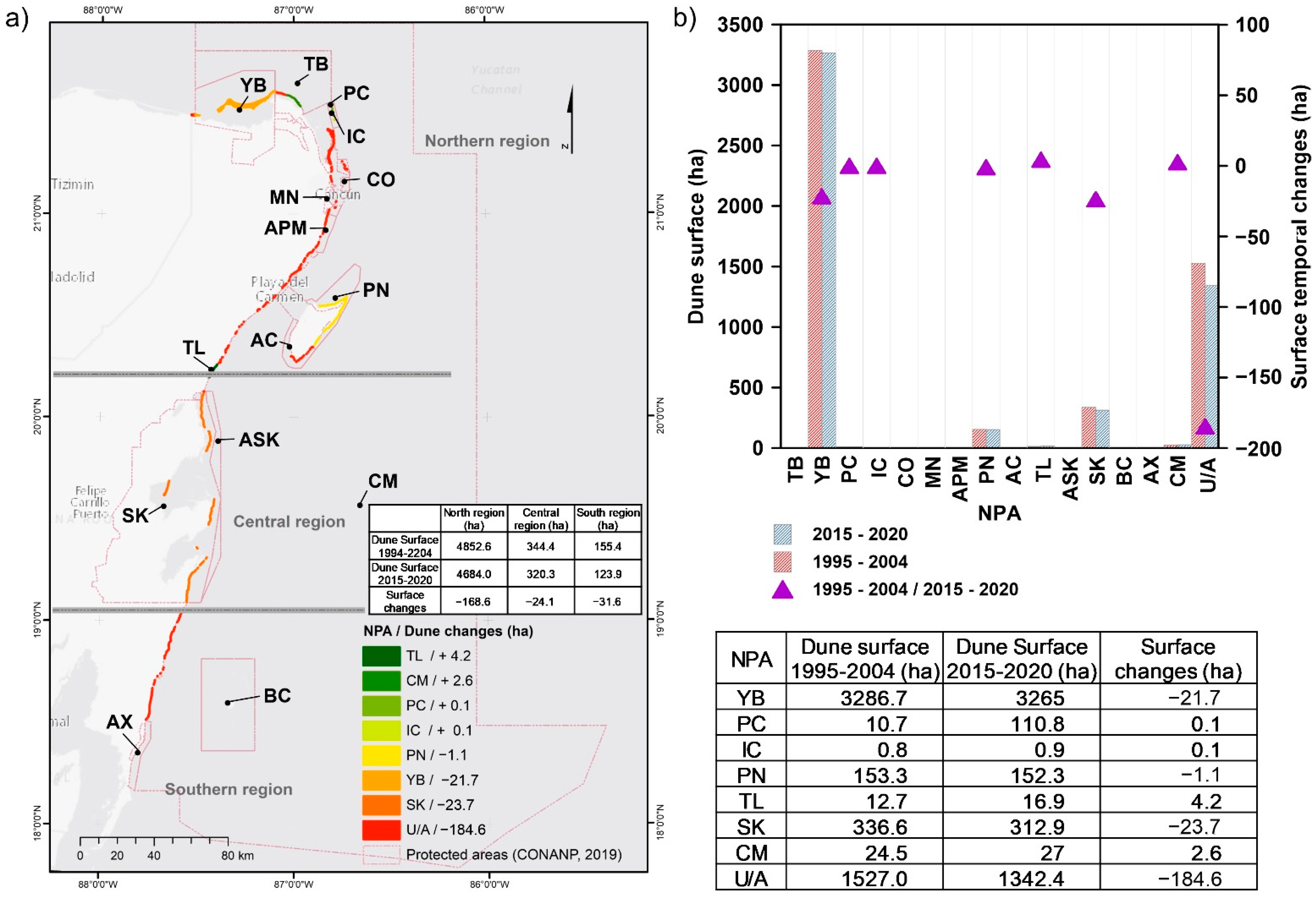
3.1.4. Coastal Dunes
3.1.5. Coastline
3.2. Stressors
3.2.1. Wave and Wind Climate
3.2.2. Water Quality
3.2.3. Anthropization
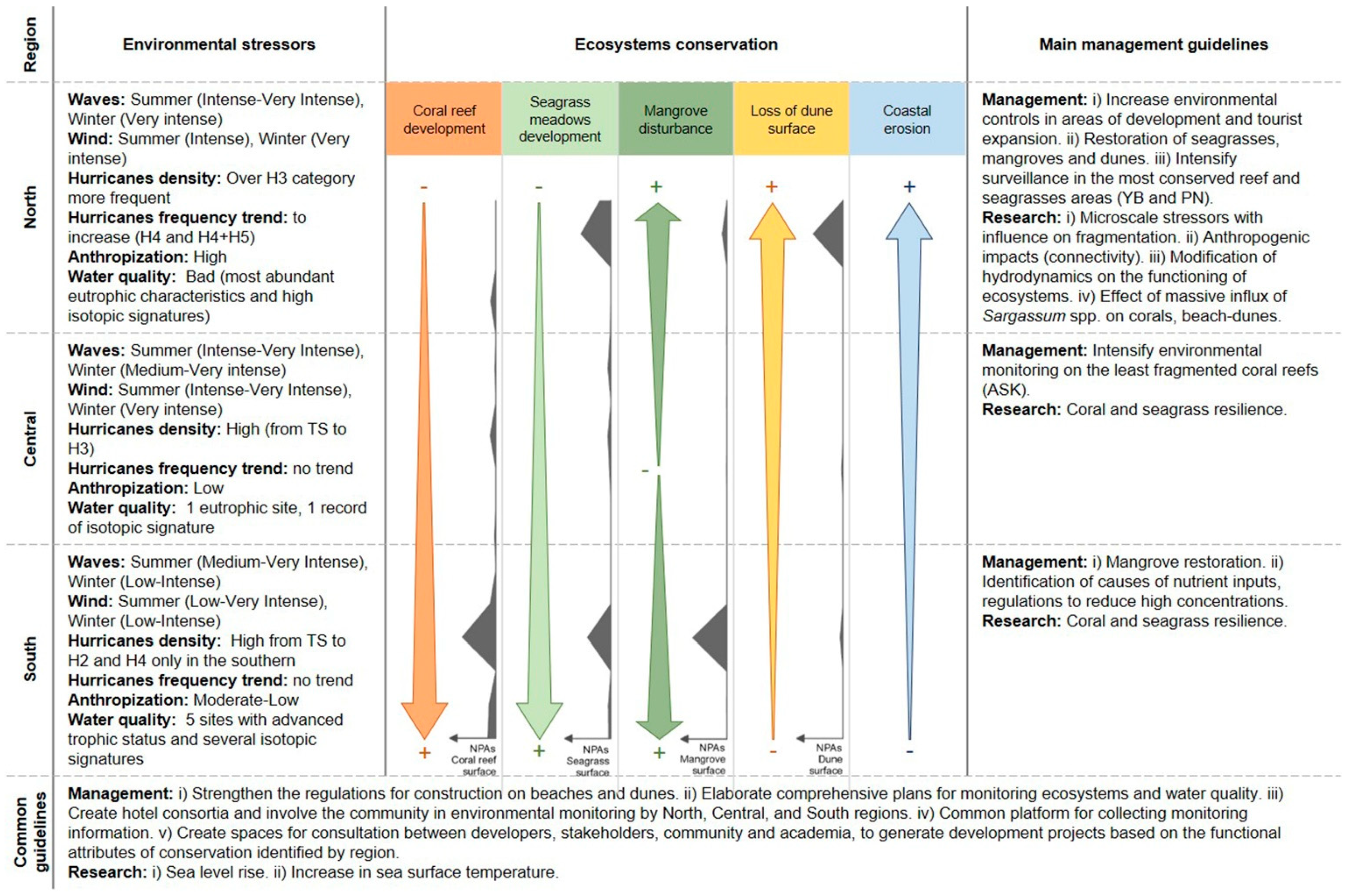
4. Discussion
4.1. Coastal Ecosystems and Wave Climate
4.2. Coastal Ecosystems and Water Quality
4.3. Coastal Ecosystems and Anthropization
4.4. Coastal Ecosystems and NPAs
4.5. Environmental Guidelines
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cortés, J. Coral reefs of the Americas: An introduction to Latin American Coral Reefs. In Latin American Coral Reefs; Cortés, J., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2003; pp. 1–7. [Google Scholar]
- Agard, J.B.R.; Cropper, A. Caribbean Sea Ecosystem Assessment (CARSEA). Caribb. Mar. Stud. 2007, 8, 1–85. [Google Scholar]
- Comisión Nacional de Áreas Naturales Protegidas (CONANP). Áreas Naturales Protegidas Federales de la República ArcGIS Metadata. Available online: http://sig.conanp.gob.mx/website/pagsig/info_shape.htm (accessed on 8 April 2020).
- Comisión Nacional de Áreas Naturales Protegidas (CONANP). Actualización de la Cobertura de Vacíos y Omisiones de Conservación en México por las Áreas Protegidas Federales, Estatales y Municipales Hasta 2016. Available online: http://sig.conanp.gob.mx/website/pagsig/mapas_serie.htm (accessed on 8 April 2020).
- Rioja-Nieto, R.; Álvarez-Filip, L. Coral reef systems of the Mexican Caribbean: Status, recent trends and conservation. Mar. Pollut. Bull. 2019, 140, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Long-term region-wide declines in Caribbean corals. Science 2003, 301, 958–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, C. Status of Coral Reefs of the World 2004; Australian Institute Marine Science: Townsville, QLD, Australia, 2004; Volume 1, pp. 1–547.
- Casarin, R.S.; Martinez, G.R.; Mariño-Tapia, I.; Vanegas, G.P.; Baldwin, E.M.; Mancera, E.E. Manmade Vulnerability of the Cancun Beach System: The Case of Hurricane Wilma. Clean Soil Air Water 2012, 40, 911–919. [Google Scholar] [CrossRef]
- Rioja-Nieto, R.; Garza-Pérez, R.; Álvarez-Filip, L.; Mariño-Tapia, I.; Enríquez, C. The Mexican Caribbean: From Xcalak to Holbox, 2nd ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Knutson, T.; Camargo, S.J.; Chan, J.C.L.; Emanuel, K.; Ho, C.H.; Kossin, J.; Mohapatra, M.; Satoh, M.; Sugi, M.; Walsh, K.; et al. Tropical cyclones and climate change assessment. Bull. Am. Meteorol. Soc. 2019, 100, 1987–2007. [Google Scholar] [CrossRef] [Green Version]
- Franks, J.S.; Johnson, D.R.; Ko, D.S.; Rubio, G.S.; Hendon, J.R.; Lay, M. Unprecedented Influx of Pelagic Sargassum along Caribbean Island Coastlines during Summer 2011. Gulf Caribb. Fish. Inst. 2011, 64, 6–8. [Google Scholar]
- Smetacek, V.; Zingone, A. Green and golden seaweed tides on the rise. Nature 2013, 504, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.; Martínez, M.L.; Odériz, I.; Mendoza, E.; Feagin, R.A. Response of vegetated dune-beach systems to storm conditions. Coast. Eng. 2016, 109, 53–62. [Google Scholar] [CrossRef]
- Uribe-Martínez, A.; Guzmán-Ramírez, A.; Arreguín-Sánchez, F.; Cuevas, E. El sargazo en el Caribe mexicano, revisión de una historia impensable. In Gobernanza y Manejo de las Costas y Mares Ante la Incertidumbre: Una Guía para Tomadores de Decisiones; Rivera-Arriaga, E., Azuz-Adeath, I., Rosas, O.D.C., Espinoza-Tenorio, A., Casarín, R.S., Ortega-Rubio, A., Botello, A.V., Vega-Serratos, B.E., Eds.; Universidad Autónoma de Campeche, RICOMAR: Campeche, Mexico, 2020; pp. 743–768. [Google Scholar]
- Chávez, V.; Uribe-Martínez, A.; Cuevas, E.; Rodríguez-Martínez, R.E.; van Tussenbroek, B.I.; Francisco, V.; Estévez, M.; Celis, L.B.; Monroy-Velázquez, L.V.; Leal-Bautista, R.; et al. Massive influx of pelagic sargassum spp. on the coasts of the mexican caribbean 2014–2020: Challenges and opportunities. Water 2020, 12, 2908. [Google Scholar] [CrossRef]
- van Tussenbroek, B.I.; Hernández Arana, H.A.; Rodríguez-Martínez, R.E.; Espinoza-Avalos, J.; Canizales-Flores, H.M.; González-Godoy, C.E.; Barba-Santos, M.G.; Vega-Zepeda, A.; Collado-Vides, L. Severe impacts of brown tides caused by Sargassum spp. on near-shore Caribbean seagrass communities. Mar. Pollut. Bull. 2017, 122, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, R.E.; van Tussenbroek, B.; Jordán-Dahlgren, E. Afluencia masiva de sargazo pelágico a la costa del Caribe mexicano (2014-2015). In Florecimientos Algales Nocivos en México, 1st ed.; CICESE: Ensenada, BC, Mexico, 2016; pp. 352–365. [Google Scholar]
- Hewitt, J.E.; Ellis, J.I.; Thrush, S.F. Multiple stressors, nonlinear effects and the implications of climate change impacts on marine coastal ecosystems. Glob. Chang. Biol. 2016, 22, 2665–2675. [Google Scholar] [CrossRef]
- Reaka-Kudla, M.L. Biodiversity of Caribbean coral reefs. In Caribbean Marine Biodiversity; Miloslavich, P., Klein, E., Eds.; Des Tech Publishers: Lancaster, CA, USA, 2005; pp. 259–276. [Google Scholar]
- van Tussenbroek, B.I. Dynamics of seagrasses and associated algae in coral reef lagoons. Hidrobiologica 2011, 21, 293–310. [Google Scholar]
- van Tussenbroek, B.I.; Cortés, J.; Collin, R.; Fonseca, A.C.; Gayle, P.M.H.; Guzmán, H.M.; Jácome, G.E.; Juman, R.; Koltes, K.H.; Oxenford, H.A.; et al. Caribbean-wide, long-term study of seagrass beds reveals local variations, shifts in community structure and occasional collapse. PLoS ONE 2014, 9, e90600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirales-Cota, M.; Espinoza-Avalos, J.; Schmook, B.; Ruiz-Luna, A.; Ramos-Reyes, R. Drivers of mangrove deforestation in Mahahual-Xcalak, Quintana Roo, southeast Mexico. Ciencias Mar. 2010, 36, 147–159. [Google Scholar] [CrossRef]
- Rodríguez-Zúñiga, M.T.; Troche-Souza, C.; Vázquez-Lule, A.D.; Márquez-Mendoza, J.D.; Balderas, B.V.; Valderrama-Landeros, L.; Velázquez-Salazar, S.; Cruz-López, M.I.; Ressl, R.; Uribe-Martínez, A.; et al. Manglares de México/Extensión, Distribución y Monitoreo; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Distrito Federal, Mexico, 2013; p. 128.
- Seingier, G.; Espejel, I.; Fermán-Almada, J.L. Cobertura vegetal y marginación en la costa mexicana. Investig. Ambient. 2009, 1, 54–69. [Google Scholar]
- Martínez, M.L.; Moreno-Casasola, P.; Espejel, I.; Jiménez-Orocio, O.; Infante Mata, D.; Rodríguez-Revelo, N. Diagnóstico de las Dunas Costeras de México; CONAFOR: Ciudad de Mexico, Mexico, 2014; p. 350. [Google Scholar]
- Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). Distribución de los Manglares en México en 2015, Escala: 1:50,000. Edición: 1. Available online: http://www.conabio.gob.mx/informacion/metadata/gis/mx_man15gw.xml?_httpcache=yes&_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no (accessed on 8 April 2020).
- Cerdeira-Estrada, S.; Martínez-Clorio, M.I.; Rosique-De-La-Cruz, L.O.; Kolb, M.; Gonzales-Posada, A.M.; Uribe-Martínez, A.; Martell-Dubois, R.; Garza-Pérez, J.R.; Alvarez-Filip, L.; Cruz-López, M.I.; et al. Cobertura Bentónica de los Ecosistemas Marinos del Caribe Mexicano: Cabo Catoche—Xcalak. 2018’, escala: 1:8000. edición: 2. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad; Universidad Nacional Autónoma de México. México. Available online: http://www.conabio.gob.mx/informacion/metadata/gis/covertv2gw.xml?_httpcache=yes&_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no (accessed on 8 April 2020).
- Argüelles-Jiménez, J.; Alva-Basurto, J.C.; Pérez-España, H.; Zetina-Rejón, M.J.; Arias-González, J.E. The measurement of ecosystem development in Caribbean coral reefs through topological indices. Ecol. Indic. 2020, 110, 105866. [Google Scholar] [CrossRef]
- Ferdie, M.; Fourqurean, J.W. Responses of seagrass communities to fertilization along a gradient of relative availability of nitrogen and phosphorus in a carbonate environment. Limnol. Oceanogr. 2004, 49, 2082–2094. [Google Scholar] [CrossRef] [Green Version]
- Valderrama-Landeros, L.H.; Martell-Dubois, R.; Ressl, R.; Silva-Casarín, R.; Cruz-Ramírez, C.J.; Muñoz-Pérez, J.J. Dynamics of coastline changes in Mexico. J. Geogr. Sci. 2019, 29, 1637–1654. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, P.S.; Eklöf, J.S.; van Katwijk, M.M.; O’Brien, K.R.; de la Torre-Castro, M.; Boström, C.; Bouma, T.J.; Krause-Jensen, D.; Unsworth, R.K.F.; van Tussenbroek, B.I.; et al. The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems—A review. Biol. Rev. 2017, 92, 1521–1538. [Google Scholar] [CrossRef] [Green Version]
- Hernández, A.L.M.; van Tussenbroek, B.I. Patch dynamics and species shifts in seagrass communities under moderate and high grazing pressure by green sea turtles. Mar. Ecol. Prog. Ser. 2014, 517, 143–157. [Google Scholar] [CrossRef]
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Schepers, D.; et al. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- European Centre for Medium-Range Weather Forecasts (ECMWF). ERA5: Data Documentation. Available online: https://confluence.ecmwf.int/display/CKB/ERA5%3A+data+documentation (accessed on 8 September 2020).
- National Oceanic and Atmospheric Administration (NOAA). International Best Track Archive for Climate Stewardship (IBTrACS). Available online: https://www.ncei.noaa.gov/data/international-best-track-archive-for-climate-stewardship-ibtracs/v04r00/access/shapefile/ (accessed on 3 December 2020).
- Hirsch, R.M.; Slack, J.R.; Smith, R.A. Techniques of trend analysis for monthly water quality data. Water Resour. Res. 1982, 18, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Westmacott, J.R.; Burn, D.H. Climate change effects on the hydrologic regime within the Churchill-Nelson River Basin. J. Hydrol. 1997, 202, 263–279. [Google Scholar] [CrossRef]
- Sen, P.K. Estimates of the Regression Coefficient Based on Kendall’s Tau. J. Am. Stat. Assoc. 1968, 63, 1379–1389. [Google Scholar] [CrossRef]
- Yue, S.; Wang, C. The Mann-Kendall Test Modified by Effective Sample Size to Detect Trend in Serially Correlated Hydrological Series. Water Resour. Manag. 2004, 18, 201–218. [Google Scholar] [CrossRef]
- Comisión Nacional del Agua (CONAGUA). Calidad del agua en México. Available online: https://www.gob.mx/conagua/articulos/calidad-del-agua (accessed on 2 December 2020).
- EEA (European Environmental Agency). Nutrients in European Ecosystems. Environmental Assessment Report No. 4; Office for official publications of the European Communities: Copenhagen, Denmark, 1999; p. 155.
- Cruz, C.J.; Mendoza, E.; Silva, R.; Chávez, V. Assessing Degrees of Anthropization on the Coast of Mexico from Ecosystem Conservation and Population Growth Data. J. Coast. Res. 2019, 92, 136–144. [Google Scholar] [CrossRef]
- Barranco, L.M.; Carriquiry, J.D.; Rodríguez-Zaragoza, F.A.; Cupul-Magaña, A.L.; Villaescusa, J.A.; Calderón-Aguilera, L.E. Spatiotemporal variations of live coral cover in the northern Mesoamerican Reef System, Yucatan Peninsula, Mexico. Sci. Mar. 2016, 80, 143–150. [Google Scholar]
- Medina-Gómez, I.; Madden, C.J.; Herrera-Silveira, J.; Kjerfve, B. Response of Thalassia Testudinum morphometry and distribution to environmental drivers in a pristine tropical lagoon. PLoS ONE 2016, 11, 1–24. [Google Scholar]
- Gutiérrez-Aguirre, M.A.; De La Fuente-Betancourt, M.G.; Cervantes-Martínez, A. Biomasa y densidad de dos especies de pastos marinos en el sur de Quintana Roo, México. Rev. Biol. Trop. 2000, 48, 313–316. [Google Scholar] [CrossRef]
- Sánchez, A.; Ortiz-Hernández, M.C.; Talavera-Sáenz, A.; Aguíñiga-García, S. Stable nitrogen isotopes in the turtle grass Thalassia testudinum from the Mexican Caribbean: Implications of anthropogenic development. Estuar. Coast. Shelf Sci. 2013, 135, 86–93. [Google Scholar] [CrossRef]
- Sánchez, A.; Anguas-Cabrera, D.; Camacho-Cruz, K.; Ortiz-Hernández, M.C.; Aguíñiga-García, S. Spatial and temporal variation of the δ15N in Thalassia testudinum in the Mexican Caribbean (2009–2017). Mar. Freshw. Res. 2020, 71, 905–912. [Google Scholar] [CrossRef]
- Mutchler, T.; Dunton, K.H.; Townsend-Small, A.; Fredriksen, S.; Rasser, M.K. Isotopic and elemental indicators of nutrient sources and status of coastal habitats in the Caribbean Sea, Yucatan Peninsula, Mexico. Estuar. Coast. Shelf Sci. 2007, 74, 449–457. [Google Scholar] [CrossRef]
- Mutchler, T.; Mooney, R.; Wallace, S.; Podsim, L.; Fredriksen, S.; Dunton, K. Origins and Fate of Inorganic Nitrogen from Land to Coastal Ocean on the Yucatan Peninsula, Mexico. Coast. Lagoons 2010, 12, 283–305. [Google Scholar]
- Odériz, I.; Gómez, I.; Ventura, Y.; Díaz, V.; Escalante, A.; Gómez, D.T.; Bouma, T.J.; Silva, R. Understanding Drivers of Connectivity and Resilience under Tropical Cyclones in Coastal Ecosystems at Puerto Morelos, Mexico. J. Coast. Res. 2020, 95, 128–132. [Google Scholar] [CrossRef]
- Mendoza, E.; Odériz, I.; Martínez, M.L.; Silva, R. Measurements and Modelling of Small Scale Processes of Vegetation Preventing Dune Erosion. J. Coast. Res. 2017, 77, 19–27. [Google Scholar] [CrossRef]
- Guimarais, M.; Zúñiga, A.; Pina, F.; Matos, F. Hurricane Paloma’s effects on seagrasses along Jardines de la Reina archipelago, Cuba. Rev. Biol. Trop. 2013, 61, 1425–1432. [Google Scholar]
- Montefalcone, M.; Parravicini, V.; Vacchi, M.; Albertelli, G.; Ferrari, M.; Morri, C.; Bianchi, C.N. Human influence on seagrass habitat fragmentation in NW Mediterranean Sea. Estuar. Coast. Shelf Sci. 2010, 86, 292–298. [Google Scholar] [CrossRef]
- El Allaoui, N.; Serra, T.; Colomer, J.; Soler, M.; Casamitjana, X.; Oldham, C. Interactions between fragmented seagrass canopies and the local hydrodynamics. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Cruz-Palacios, V.; van Tussenbroek, B.I. Simulation of hurricane-like disturbances on a Caribbean seagrass bed. J. Exp. Mar. Biol. Ecol. 2005, 324, 44–60. [Google Scholar] [CrossRef]
- James, R.K.; Silva, R.; van Tussenbroek, B.I.; Escudero-Castillo, M.; Mariño-Tapia, I.; Dijkstra, H.A.; Van Westen, R.M.; Pietrzak, J.D.; Candy, A.S.; Katsman, C.A.; et al. Maintaining Tropical Beaches with Seagrass and Algae: A Promising Alternative to Engineering Solutions. Bioscience 2019, 69, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Boehlert, G.W. Larval dispersal and survival in tropical reef fishes. In Reef Fisheries; Polunin, N.V.C., Roberts, C.M., Eds.; Fish and Fisheries Series; Chapman & Hall: London, UK, 1996; Volume 20, pp. 61–84. [Google Scholar]
- Alcérreca-Huerta, J.C.; Encarnacion, J.I.; Ordoñez-Sánchez, S.; Callejas-Jiménez, M.; Barroso, G.G.D.; Allmark, M.; Mariño-Tapia, I.; Casarín, R.S.; O’Doherty, T.; Johnstone, C.; et al. Energy yield assessment from ocean currents in the insular shelf of Cozumel Island. J. Mar. Sci. Eng. 2019, 7, 147. [Google Scholar] [CrossRef] [Green Version]
- Van Dijk, J.K.; van Tussenbroek, B.I.; Jiménez-Durán, K.; Márquez-Guzman, G.J.; Ouborg, J. High levels of gene flow and low population genetic structure related to high dispersal potential of a tropical marine angiosperm. Mar. Ecol. Prog. Ser. 2009, 390, 67–77. [Google Scholar] [CrossRef]
- Porto-Hannes, I.; Zubillaga, A.L.; Shearer, T.L.; Bastidas, C.; Salazar, C.; Coffroth, M.A.; Szmant, A.M. Population structure of the corals Orbicella faveolata and Acropora palmata in the Mesoamerican Barrier Reef System with comparisons over Caribbean basin-wide spatial scale. Mar. Biol. 2015, 162, 81–98. [Google Scholar] [CrossRef]
- Carriquiry, D.J.; Barranco-Servin, M.L.; Villaescusa, A.J.; Camacho-Ibar, F.V.; Reyes-Bonilla, H.; Cupul-Magaña, L.A. Conservation and Sustainability of Mexican Caribbean Coral Reefs and the Threats of a Human-Induced Phase-Shift. In Environmental Change Sustainability; Silvern, S., Young, S., Eds.; InTech: London, UK, 2013; pp. 29–51. [Google Scholar]
- Camacho-Cruz, K.A.; Ortiz-Hernández, M.C.; Sánchez, A.; Carrillo, L.; De Jesús Navarrete, A. Water quality in the eastern karst region of the Yucatan Peninsula: Nutrients and stable nitrogen isotopes in turtle grass, Thalassia testudinum. Environ. Sci. Pollut. Res. 2020, 27, 15967–15983. [Google Scholar] [CrossRef]
- Ladd, M.C.; Collado-Vides, L. Practical applications of monitoring results to improve managing for coral reef resilience: A case study in the Mexican Caribbean. Biodivers. Conserv. 2013, 22, 1591–1608. [Google Scholar] [CrossRef]
- van Tussenbroek, B.; Hermus, K.; Tahey, T. Biomass and Growth of the Turtle Grass Thalassia testudinum (Banks ex Koníg) in a Shallow Tropical Lagoon System, in Relation to Tourist Development. Caribb. J. Sci. 1996, 32, 357–364. [Google Scholar]
- Hernández-Terrones, L.; Rebolledo-Vieyra, M.; Merino-Ibarra, M.; Soto, M.; Le-Cossec, A.; Monroy-Ríos, E. Groundwater pollution in a karstic region (NE Yucatan): Baseline nutrient content and flux to coastal ecosystems. Water Air Soil Pollut. 2011, 218, 517–528. [Google Scholar] [CrossRef]
- Carruthers, T.J.B.; van Tussenbroek, B.I.; Dennison, W.C. Influence of submarine springs and wastewater on nutrient dynamics of Caribbean seagrass meadows. Estuar. Coast. Shelf Sci. 2005, 64, 191–199. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Langton, R.; Bedford, B.J.; Potts, A.C.; Day, O.; Hu, C. Land-based nutrient enrichment of the Buccoo Reef Complex and fringing coral reefs of Tobago, West Indies. Mar. Pollut. Bull. 2010, 60, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, B.E.; Littler, M.M.; Littler, D.S. A comparison of nutrient-limited productivity in macroalgae from a Caribbean barrier reef and from a mangrove ecosystem. Aquat. Bot. 1987, 28, 243–255. [Google Scholar] [CrossRef]
- Lapointe, B.E. Nutrient thresholds for bottom-up control of macroalgal blooms on coral reefs in Jamaica and southeast Florida. Limnol. Oceanogr. 1997, 42, 1119–1131. [Google Scholar] [CrossRef] [Green Version]
- Mumby, P.J. The impact of exploiting grazers (Scaridae) on the dynamics of Caribbean coral reefs. Ecol. Appl. 2006, 16, 747–769. [Google Scholar] [CrossRef] [Green Version]
- Burkepile, D.E.; Hay, M.E. Herbivore species richness and feeding complementarity affect community structure and function on a coral reef. Proc. Natl. Acad. Sci. USA 2008, 105, 16201–16206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchley, A.; McField, M.D.; Alvarez-Filip, L. Rapidly increasing macroalgal cover not related to herbivorous fishes on Mesoamerican reefs. PeerJ 2016, 4, e2084. [Google Scholar] [CrossRef] [Green Version]
- Duarte, C.M.; Martín, M.; Margarita, G. Evidence of iron deficiency in seagrasses growing above carbonate sediments. Limnol. Oceanogr. 1995, 40, 1153–1158. [Google Scholar] [CrossRef]
- Moore, K.A.; Wetzel, R.L. Seasonal variations in eelgrass (Zostera marina L.) responses to nutrient enrichment and reduced light availability in experimental ecosystems. J. Exp. Mar. Biol. Ecol. 2000, 244, 1–28. [Google Scholar] [CrossRef]
- Silberstein, K.; Chiffings, A.W.; McComb, A.J. The loss of seagrass in cockburn sound, Western Australia. III. The effect of epiphytes on productivity of Posidonia australis Hook. F. Aquat. Bot. 1986, 24, 355–371. [Google Scholar] [CrossRef]
- Tomasko, D.A.; Lapointe, B.E. Productivity and biomass of Thalassia testudinum as related to water column nutrient availability and epiphyte levels: Field observations and experimental studies. Mar. Ecol. Prog. Ser. 1991, 75, 9–17. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Tomasko, D.A.; Matzie, W.R. Eutrophication and trophic state classification of seagrass communities in the Florida Keys. Bull. Mar. Sci. 1994, 54, 696–717. [Google Scholar]
- Suchley, A.; Alvarez-Filip, L. Local human activities limit marine protection efficacy on Caribbean coral reefs. Conserv. Lett. 2018, 11, e12571. [Google Scholar] [CrossRef]
- Denny, M.W. Biology and the Mechanics of the Wave-Swept Environment; Princeton University Press: Princeton, NJ, USA, 1988; p. 329. [Google Scholar]
- Koch, E.W.; Gust, G. Water flow in tide- and wave-dominated beds of the seagrass Thalassia testudinum. Mar. Ecol. Prog. Ser. 1999, 184, 63–72. [Google Scholar] [CrossRef]
- Wenger, A.S.; Williamson, D.H.; da Silva, E.T.; Ceccarelli, D.M.; Browne, N.K.; Petus, C.; Devlin, M.J. Effects of reduced water quality on coral reefs in and out of no-take marine reserves. Conserv. Biol. 2016, 30, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Pollock, F.J.; Lamb, J.B.; Field, S.N.; Heron, S.F.; Schaffelke, B.; Shedrawi, G.; Bourne, D.G.; Willis, B.L. Sediment and turbidity associated with offshore dredging increase coral disease prevalence on nearby reefs. PLoS ONE 2014, 9, e102498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, D.I.; Carruthers, T.J.B.; Morrison, P.F.; McComb, A.J. Experimental manipulation of canopy density in a temperate seagrass Amphibolis griffithii (Black) den Hartog] meadow: Effects on sediments. In Seagrass Biology; Kuo, J., Phillips, R.C., Walker, D.I., Kirkman, H., Eds.; Proceedings of an International Workshop; University of Western Australia: Nedlans, WA, Australia, 1996; pp. 117–122. [Google Scholar]
- Creed, J.C.; Amado Filho, G.M. Disturbance and recovery of the macroflora of a seagrass (Halodule wrightii Ascherson) meadow in the Abrolhos Marine National Park, Brazil: An experimental evaluation of anchor damage. J. Exp. Mar. Biol. Ecol. 1999, 235, 285–306. [Google Scholar] [CrossRef]
- Orth, R.J.; Fishman, J.R.; Wilcox, D.J.; Moore, K.A. Identification and Management of Fishing Gear Impacts in a Recovering Seagrass System in the Coastal Bays of the Delmarva Peninsula, USA. J. Coast. Res. 2002, 111–129. [Google Scholar]
- Pérez-Villegas, G.; Carrascal, E. Tourism development in Cancun, Quintana Roo and its consequences on vegetation. Investig. Geogr. 2000, 43, 145–166. [Google Scholar]
- Pethick, J.; Ruitenbeek, J. Coastal Geomorphology and Climate Change Adaptation. In Opportunities for Environmentally Healthy, Inclusive, and Resilient Growth in Mexico’s Yucatán Peninsula; Sánchez-Triana, E., Ruitenbeek, J., Enriquez, S., Siegmann, K., Eds.; International Development in Focus/World Bank Group: Washington, DC, USA, 2020; pp. 23–40. [Google Scholar]
- Bozec, Y.M.; Alvarez-Filip, L.; Mumby, P.J. The dynamics of architectural complexity on coral reefs under climate change. Glob. Chang. Biol. 2015, 21, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Zaragoza, F.A.; Ortiz, M.; Berrios, F.; Campos, L.; De Jesús-Navarrete, A.; Castro-Pérez, J.; Hernández-Flores, A.; García-Rivas, M.; Fonseca-Peralta, F.; Gallegos-Aguilar, E. Trophic models and short-term dynamic simulations for benthic-pelagic communities at banco chinchorro biosphere reserve (Mexican Caribbean): A conservation case. Community Ecol. 2016, 17, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Sleeman, J.C.; Kendrick, G.A.; Boggs, G.S.; Hegge, B.J. Measuring fragmentation of seagrass landscapes: Which indices are most appropriate for detecting change? Mar. Freshw. Res. 2005, 56, 851–864. [Google Scholar] [CrossRef]
- Alvarez-Filip, L.; Estrada-Saldívar, N.; Pérez-Cervantes, E.; Molina-Hernández, A.; González-Barrios, F.J. A rapid spread of the stony coral tissue loss disease outbreak in the Mexican Caribbean. PeerJ 2019, 7, e8069. [Google Scholar] [CrossRef] [Green Version]
- Unsworth, R.K.F.; Collier, C.J.; Waycott, M.; Mckenzie, L.J.; Cullen-Unsworth, L.C. A framework for the resilience of seagrass ecosystems. Mar. Pollut. Bull. 2015, 100, 34–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.S.; Fonseca, M.S.; Stafford, N.B. Seagrass ecology: New contributions from a landscape perspective. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 625–645. [Google Scholar]
- Duarte, C.M.; Fourqurean, J.W.; Krause-Jensen, D.; Olesen, B. Dynamics of Seagrass Stability and Change. In Seagrasses: Biology, Ecology and Conservation; Larkum, A.W.D., Orth, R., Duarte, C.M., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 271–294. [Google Scholar]
- Brenner, L.; Engelbauer, M.; Job, H. Mitigating tourism-driven impacts on mangroves in Cancún and the Riviera Maya, Mexico: An evaluation of conservation policy strategies and environmental planning instruments. J. Coast. Conserv. 2018, 22, 755–767. [Google Scholar] [CrossRef]
- Vázquez-Lule, A.D.; Díaz-Gallegos, J.; Adame, M.F. Caracterización del sitio de manglar Cozumel. In Sitios de Manglar con Relevancia Biológica y con Necesidades de Rehabilitación Ecológica; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO): Distrito Federal, Mexico, 2009; pp. 1–17. [Google Scholar]
- Cinco-Castro, S.; Herrera-Silveira, J. Vulnerability of mangrove ecosystems to climate change effects: The case of the Yucatan Peninsula. Ocean Coast. Manag. 2020, 192, 105196. [Google Scholar] [CrossRef]
- Liu, Y.; Kerkering, H.; Weisberg, R.H. Coastal Ocean Observing Systems; Elsevier: London, UK, 2015; pp. 1–461. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Official Names of the NPAs [3] | Management Category [3] | Abbreviations | PMR [4] |
|---|---|---|---|
| Tiburón Ballena | Biosphere Reserve | TB | Extreme importance |
| Yum Balam | Flora and Fauna Protection Area | YB | Extreme importance |
| Playa de la Isla Contoy | Sanctuary | PC | |
| Isla Contoy | National Park | IC | |
| Costa Occidental de Isla Mujeres, Punta Cancún y Punta Nizuc | National Park | CO | |
| Manglares de Nichupté | Flora and Fauna Protection Area | MN | |
| Arrecife de Puerto Morelos | National Park | APM | |
| Porción norte y la franja costera oriental, terrestres y marinas de la Isla de Cozumel | Flora and Fauna Protection Area | PN | Very important |
| Arrecifes de Cozumel | National Park | AC | Very important |
| Tulum | National Park | TL | Very important |
| Arrecifes de Sian Ka’an | Biosphere Reserve | ASK | Extreme importance |
| Sian Ka’an | Biosphere Reserve | SK | Extreme importance |
| Banco Chinchorro | Biosphere Reserve | BC | Extreme importance |
| Arrecifes de Xcalak | National Park | AX | Very important |
| Caribe Mexicano | Biosphere Reserve | CM | |
| Unprotected areas | U/A |
| Energy Level | Wave Power (kW/m) | Wind Power (kW/m2) |
|---|---|---|
| Low | <5.3 | <0.21 |
| Medium | 5.3 < P < 5.9 | 0.21 < P < 0.24 |
| Intense | 5.3 < P < 6.5 | 0.24 < P < 0.26 |
| Very intense | >6.5 | >0.26 |
| Region | Minimum Latitude | Maximum Latitude | Wave Power (kW/m) | Mean Wave Direction (°) | Wind Power (kW/m2) | Mean Wind Direction (°) |
|---|---|---|---|---|---|---|
| North | 21.75 | 22.25 | Intense | 42 | Medium | 77 |
| 21.25 | 21.75 | Very intense | 62 | Very intense | 80 | |
| 20.75 | 21.25 | Very intense | 74 | Very intense | 78 | |
| Central | 20.25 | 20.75 | Low | 99 | Low | 80 |
| 19.75 | 20.25 | Medium | 91 | Intense | 74 | |
| 19.25 | 19.75 | Very intense | 83 | Very intense | 75 | |
| South | 18.75 | 19.25 | Low | 86 | Low | 76 |
| 18.25 | 18.75 | Low | 84 | Low | 74 | |
| 17.75 | 18.25 | Intense | 80 | Intense | 73 |
| Region | Minimum Latitude | Maximum Latitude | Wave Power (kW/m) | Mean Wave Direction (°) | Wind Power (kW/m2) | Mean Wind Direction (°) |
|---|---|---|---|---|---|---|
| North | 21.75 | 22.25 | Low | 85 | Low | 95 |
| 21.25 | 21.75 | Intense | 100 | Intense | 98 | |
| 20.75 | 21.25 | Very intense | 103 | Intense | 99 | |
| Central | 20.25 | 20.75 | Low | 116 | Low | 104 |
| 19.75 | 20.25 | Intense | 107 | Very intense | 96 | |
| 19.25 | 19.75 | Very intense | 102 | Very intense | 97 | |
| South | 18.75 | 19.25 | Medium | 100 | Low | 96 |
| 18.25 | 18.75 | Medium | 99 | Intense | 94 | |
| 17.75 | 18.25 | Very intense | 95 | Very intense | 93 |
| Saffir-Simpson Scale | Mann-K tau (τ) | p-Value * | Sen’s Slope | Trend Estimation |
|---|---|---|---|---|
| TD | 0.033 | 0.762 | 0.000 | No trend |
| TS | −0.090 | 0.323 | 0.000 | No trend |
| H1 | −0.079 | 0.285 | 0.000 | No trend |
| H2 | −0.187 | 0.150 | −0.009 | No trend |
| H3 | 0.179 | 0.216 | 0.012 | No trend |
| H4 | 0.509 | 0.046 | 0.062 | Trend |
| H5 | 0.105 | 1.000 | 0.011 | No trend |
| H4 + H5 | 0.357 | 0.044 | 0.023 | Trend |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarais, M.; Zúñiga-Ríos, A.; Cruz-Ramírez, C.J.; Chávez, V.; Odériz, I.; van Tussenbroek, B.I.; Silva, R. The Conservational State of Coastal Ecosystems on the Mexican Caribbean Coast: Environmental Guidelines for Their Management. Sustainability 2021, 13, 2738. https://doi.org/10.3390/su13052738
Guimarais M, Zúñiga-Ríos A, Cruz-Ramírez CJ, Chávez V, Odériz I, van Tussenbroek BI, Silva R. The Conservational State of Coastal Ecosystems on the Mexican Caribbean Coast: Environmental Guidelines for Their Management. Sustainability. 2021; 13(5):2738. https://doi.org/10.3390/su13052738
Chicago/Turabian StyleGuimarais, Mayrene, Adán Zúñiga-Ríos, Cesia J. Cruz-Ramírez, Valeria Chávez, Itxaso Odériz, Brigitta I. van Tussenbroek, and Rodolfo Silva. 2021. "The Conservational State of Coastal Ecosystems on the Mexican Caribbean Coast: Environmental Guidelines for Their Management" Sustainability 13, no. 5: 2738. https://doi.org/10.3390/su13052738