
Sustainability in Natural Grassland in the Brazilian Pampa Biome: Livestock Production with CO2 Absorption

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Pasture Management
2.3. Instrumentation and Data Processing
2.4. Estimates of Uncertainty in NEE
2.5. Energy Balance Closure
3. Results
3.1. Pasture Management
3.2. Climate and Soil Conditions
3.3. CO2 Flux Dynamics
3.4. Cumulative NEE
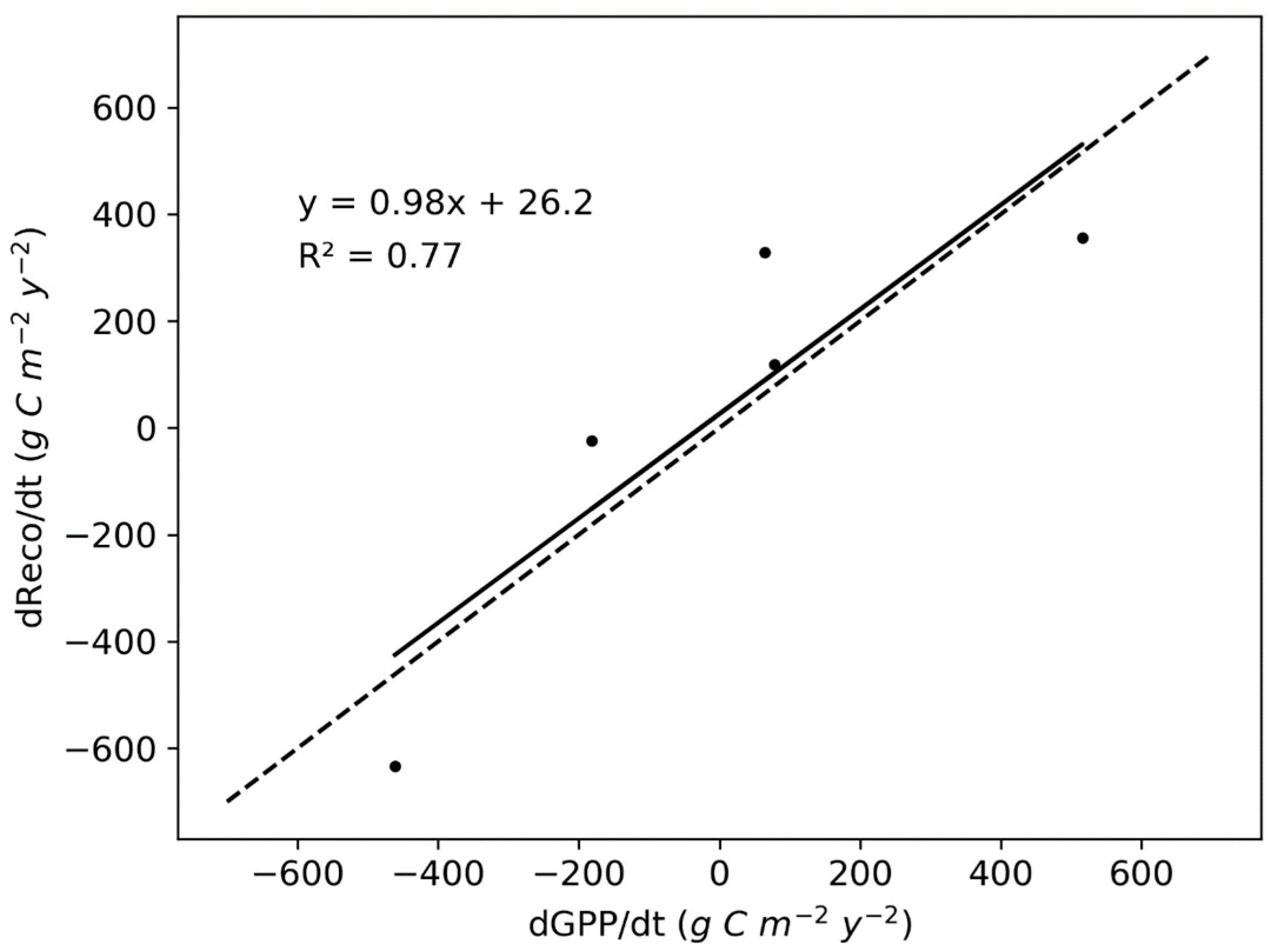
3.5. Environmental Controls of NEP, GPP, and Reco
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boldrini, I.L.O.B.B. Bioma Pampa: Diversidade Florística e Fisionômica; Pallotti: Porto Alegre, Brazil, 2010; ISBN 9788562689345. [Google Scholar]
- Jaurena, M.; Durante, M.; Devincenzi, T.; Savian, J.V.; Bendersky, D.; Moojen, F.G.; Pereira, M.; Soca, P.; Quadros, F.L.F.; Pizzio, R.; et al. Native Grasslands at the Core: A New Paradigm of Intensification for the Campos of Southern South America to Increase Economic and Environmental Sustainability. Front. Sustain. Food Syst. 2021, 5, 547834. [Google Scholar] [CrossRef]
- Baggio, R.; Overbeck, G.E.; Durigan, G.; Pillar, V.D. To Graze or Not to Graze: A Core Question for Conservation and Sustainable Use of Grassy Ecosystems in Brazil. Perspect. Ecol. Conserv. 2021, 19, 256–266. [Google Scholar] [CrossRef]
- Bencke, G. Diversidade e Conservação Da Fauna Dos Campos Do Sul Do Brasil. In Campos Sulinos: Conservação e Uso Sustentável da Biodiversidade; Pillar, V.D., Ed.; Ministério do Meio Ambiente: Brasília, Brazil, 2009; pp. 101–121. [Google Scholar]
- Baeza, S.; Vélez-Martin, E.; De Abelleyra, D.; Banchero, S.; Gallego, F.; Schirmbeck, J.; Veron, S.; Vallejos, M.; Weber, E.; Oyarzabal, M.; et al. Two Decades of Land Cover Mapping in the Río de La Plata Grassland Region: The MapBiomas Pampa Initiative. Remote Sens. Appl. 2022, 28, 100834. [Google Scholar] [CrossRef]
- De Oliveira, T.E.; de Freitas, D.S.; Gianezini, M.; Ruviaro, C.F.; Zago, D.; Mércio, T.Z.; Dias, E.A.; Lampert, V.D.N.; Barcellos, J.O.J. Agricultural Land Use Change in the Brazilian Pampa Biome: The Reduction of Natural Grasslands. Land Use Policy 2017, 63, 394–400. [Google Scholar] [CrossRef]
- MapBiomas Mapeamento Anual de Cobertura e Uso Da Terra No Pampa—Coleção 7. Available online: http://alerta.mapbiomas.org/ (accessed on 19 March 2024).
- Tiscornia, G.; Jaurena, M.; Baethgen, W. Drivers, Process, and Consequences of Native Grassland Degradation: Insights from a Literature Review and a Survey in Río de La Plata Grasslands. Agronomy 2019, 9, 239. [Google Scholar] [CrossRef]
- López-González, F.A.; Allende, R.; de Lima, J.M.S.; Canozzi, M.E.A.; Sessim, A.G.; Barcellos, J.O.J. Intensification of Cow-Calf Production: How Does the System Respond Biologically to Energy Inputs in a Long-Term Horizon? Livest. Sci. 2020, 237, 104058. [Google Scholar] [CrossRef]
- Modernel, P.; Dogliotti, S.; Alvarez, S.; Corbeels, M.; Picasso, V.; Tittonell, P.; Rossing, W.A.H. Identification of Beef Production Farms in the Pampas and Campos Area That Stand out in Economic and Environmental Performance. Ecol. Indic. 2018, 89, 755–770. [Google Scholar] [CrossRef]
- Ruviaro, C.F.; da Costa, J.S.; Florindo, T.J.; Rodrigues, W.; de Medeiros, G.I.B.; Vasconcelos, P.S. Economic and Environmental Feasibility of Beef Production in Different Feed Management Systems in the Pampa Biome, Southern Brazil. Ecol. Indic. 2016, 60, 930–939. [Google Scholar] [CrossRef]
- Azadi, H.; Taheri, F.; Burkart, S.; Mahmoudi, H.; De Maeyer, P.; Witlox, F. Impact of Agricultural Land Conversion on Climate Change. Environ. Dev. Sustain. 2021, 23, 3187–3198. [Google Scholar] [CrossRef]
- Kuinchtner, B.C.; Ferreira De Quadros, F.L.; Casanova, P.T.; Marin, L.; Seeger, B.B.; de Moura Steinhorst, D.; Ongaratto, F. Thermal Sum (Degrees-Days) Rest Period as a Grazing Management Tool in Natural Grasslands: Effects on Animal Performance and Forage Production. Environ. Sustain. Indic. 2021, 10, 100125. [Google Scholar] [CrossRef]
- Gomes, S.; Lopes, J.M.; Nogueira, S. Willingness to Pay More for Green Products: A Critical Challenge for Gen Z. J. Clean. Prod. 2023, 390, 136092. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Bork, E.W.; Richter, G.M.; Chen, C.; Hussain Shah, S.H.; Mezbahuddin, S. Effects of Grazing Management on Spatio-Temporal Heterogeneity of Soil Carbon and Greenhouse Gas Emissions of Grasslands and Rangelands: Monitoring, Assessment and Scaling-Up. J. Clean. Prod. 2021, 288, 125737. [Google Scholar] [CrossRef]
- Smith, K.A.; Conen, F. Impacts of Land Management on Fluxes of Trace Greenhouse Gases. Soil. Use Manag. 2004, 20, 255–263. [Google Scholar] [CrossRef]
- Guan, K.; Jin, Z.; Peng, B.; Tang, J.; DeLucia, E.H.; West, P.C.; Jiang, C.; Wang, S.; Kim, T.; Zhou, W.; et al. A Scalable Framework for Quantifying Field-Level Agricultural Carbon Outcomes. Earth Sci. Rev. 2023, 243, 104462. [Google Scholar] [CrossRef]
- Baldocchi, D.D. Assessing the Eddy Covariance Technique for Evaluating Carbon Dioxide Exchange Rates of Ecosystems: Past, Present and Future. Glob. Chang. Biol. 2003, 9, 479–492. [Google Scholar] [CrossRef]
- Gomez-Casanovas, N.; Delucia, N.J.; Bernacchi, C.J.; Boughton, E.H.; Sparks, J.P.; Chamberlain, S.D.; Delucia, E.H. Grazing Alters Net Ecosystem C Fluxes and the Global Warming Potential of a Subtropical Pasture. Ecol. Appl. 2018, 28, 557–572. [Google Scholar] [CrossRef] [PubMed]
- de la Motte, L.G.; Mamadou, O.; Beckers, Y.; Bodson, B.; Heinesch, B.; Aubinet, M. Rotational and Continuous Grazing Does Not Affect the Total Net Ecosystem Exchange of a Pasture Grazed by Cattle but Modifies CO2 Exchange Dynamics. Agric. Ecosyst. Environ. 2018, 253, 157–165. [Google Scholar] [CrossRef]
- Wall, A.M.; Campbell, D.I.; Mudge, P.L.; Schipper, L.A. Temperate Grazed Grassland Carbon Balances for Two Adjacent Paddocks Determined Separately from One Eddy Covariance System. Agric. For. Meteorol. 2020, 287, 107942. [Google Scholar] [CrossRef]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World Map of the Köppen-Geiger Climate Classification Updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. World Reference Base for Soil Resources 2014: International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; FAO: Rome, Italy, 2014; ISBN 9789251083697. [Google Scholar]
- Rubert, G.; Roberti, D.; Pereira, L.S.; Quadros, F.; Velho, H.C.; de Moraes, O. Evapotranspiration of the Brazilian Pampa Biome: Seasonality and Influential Factors. Water 2018, 10, 1864. [Google Scholar] [CrossRef]
- Andrade, B.O.; Bonilha, C.L.; Overbeck, G.E.; Vélez-Martin, E.; Rolim, R.G.; Bordignon, S.A.L.; Schneider, A.A.; Vogel Ely, C.; Lucas, D.B.; Garcia, É.N.; et al. Classification of South Brazilian Grasslands: Implications for Conservation. Appl. Veg. Sci. 2019, 22, 168–184. [Google Scholar] [CrossRef]
- Confortin, A.C.C.; Quadros, F.L.F.; Santos, A.B.; Seibert, L.; Severo, P.O.; Ribeiro, B.S.R. Leaf Tissue Fluxes of Pampa Biome Native Grasses Submitted to Two Grazing Intervals. Grass Forage Sci. 2017, 72, 654–662. [Google Scholar] [CrossRef]
- Barbieri, C.W.; de Quadros, F.L.F.; Jochims, F.; Kuinchtner, B.C.; de Carvalho, T.H.N.; Casanova, P.T.; Fernandes, A.M.; Pereira, J.B. Beef Heifers Grazing Behavior and Herbage Intake in Natural Grassland under Rotational Grazing. Ciência Rural. 2015, 45, 2056–2062. [Google Scholar] [CrossRef]
- Haydock, K.P.; Shaw, N. The Comparative Yield Method for Estimating Dry Matter Yield of Pasture. Aust. J. Exp. Agric. 1975, 15, 663–670. [Google Scholar]
- Hersbach, H.; Bell, B.; Berrisford, P.; Biavati, G.; Horányi, A.; Muñoz Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Rozum, I.; et al. ERA5 Hourly Data on Pressure Levels from 1940 to Present. Copernicus Climate Change Service (C3S) Climate Data Store (CDS). Available online: https://cds.climate.copernicus.eu/cdsapp#!/dataset/reanalysis-era5-pressure-levels?tab=overview (accessed on 5 January 2023).
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic Anemometer Tilt Correction Algorithms. Bound. Layer. Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of Flux Measurements for Density Effects Due to Heat and Water Vapour Transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Gash, J.H.C.; Culf, A.D. Applying a Linear Detrend to Eddy Correlation Data in Realtime. Bound. Layer. Meteorol. 1996, 79, 301–306. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; De Bruin, H.; Elbers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A System to Measure Surface Fluxes of Momentum, Sensible Heat, Water Vapour and Carbon Dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Moncrieff, J.; Clement, R.; Finnigan, J.; Meyers, T. Averaging, Detrending, and Filtering of Eddy Covariance Time Series. In Handbook of Micrometeorology: A Guide for Surface Flux Measurement and Analysis; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Vickers, D.; Mahrt, L. Quality Control and Flux Sampling Problems for Tower and Aircraft Data. J. Atmos. Ocean. Technol. 1997, 14, 512–526. [Google Scholar] [CrossRef]
- Foken, T.; Gockede, M.; Mauder, M.; Mahrt, L.; Amiro, B.; Munger, W. Chapter 9: Post-Field Data Quality Control. In Handbook of Micrometeorology: A Guide for Surface Flux Measurement and Analysis; Springer: Dordrecht, The Netherlands, 2004; ISBN 978-1-4020-2264-7. [Google Scholar]
- Béziat, P.; Ceschia, E.; Dedieu, G. Carbon Balance of a Three Crop Succession over Two Cropland Sites in South West France. Agric. For. Meteorol. 2009, 149, 1628–1645. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a Standardized Processing of Net Ecosystem Exchange Measured with Eddy Covariance Technique: Algorithms and Uncertainty Estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef]
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A Simple Two-Dimensional Parameterisation for Flux Footprint Prediction (FFP). Geosci. Model. Dev. 2015, 8, 3695–3713. [Google Scholar] [CrossRef]
- Wutzler, T.; Lucas-Moffat, A.; Migliavacca, M.; Knauer, J.; Sickel, K.; Šigut, L.; Menzer, O.; Reichstein, M. Basic and Extensible Post-Processing of Eddy Covariance Flux Data with REddyProc. Biogeosciences 2018, 15, 5015–5030. [Google Scholar] [CrossRef]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the Separation of Net Ecosystem Exchange into Assimilation and Ecosystem Respiration: Review and Improved Algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hollinger, D.Y.; Burba, G.G.; Davis, K.J.; Flanagan, L.B.; Katul, G.G.; Munger, J.W.; Ricciuto, D.M.; Stoy, P.C.; Suyker, A.E.; et al. A Multi-Site Analysis of Random Error in Tower-Based Measurements of Carbon and Energy Fluxes. Agric. For. Meteorol. 2006, 136, 1–18. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hollinger, D.Y. Statistical Modeling of Ecosystem Respiration Using Eddy Covariance Data: Maximum Likelihood Parameter Estimation, and Monte Carlo Simulation of Model and Parameter Uncertainty, Applied to Three Simple Models. Agric. For. Meteorol. 2005, 131, 191–208. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hollinger, D.Y. A Method to Estimate the Additional Uncertainty in Gap-Filled NEE Resulting from Long Gaps in the CO2 Flux Record. Agric. For. Meteorol. 2007, 147, 199–208. [Google Scholar] [CrossRef]
- Zeri, M.; Sá, L.D.A. The Impact of Data Gaps and Quality Control Filtering on the Balances of Energy and Carbon for a Southwest Amazon Forest. Agric. For. Meteorol. 2010, 150, 1543–1552. [Google Scholar] [CrossRef]
- Foken, T. The Energy Balance Closure Problem: An Overview. Ecol. Appl. 2008, 18, 1351–1367. [Google Scholar] [CrossRef]
- Barr, A.G.; van der Kamp, G.; Black, T.A.; McCaughey, J.H.; Nesic, Z. Energy Balance Closure at the BERMS Flux Towers in Relation to the Water Balance of the White Gull Creek Watershed 1999–2009. Agric. For. Meteorol 2012, 153, 3–13. [Google Scholar] [CrossRef]
- Grimm, A.M.; Barros, V.R.; Doyle, M.E. Climate Variability in Southern South America Associated with El Niño and La Niña Events. J. Clim. 2000, 13, 35–58. [Google Scholar] [CrossRef]
- Zimmer, T.; Buligon, L.; de Arruda Souza, V.; Romio, L.C.; Roberti, D.R. Influence of Clearness Index and Soil Moisture in the Soil Thermal Dynamic in Natural Pasture in the Brazilian Pampa Biome. Geoderma 2020, 378, 114582. [Google Scholar] [CrossRef]
- Hoeppner, S.S.; Dukes, J.S. Interactive Responses of Old-Field Plant Growth and Composition to Warming and Precipitation. Glob. Chang. Biol. 2012, 18, 1754–1768. [Google Scholar] [CrossRef]
- Gourlez de la Motte, L.; Jérôme, E.; Mamadou, O.; Beckers, Y.; Bodson, B.; Heinesch, B.; Aubinet, M. Carbon Balance of an Intensively Grazed Permanent Grassland in Southern Belgium. Agric. For. Meteorol. 2016, 228–229, 370–383. [Google Scholar] [CrossRef]
- Rutledge, S.; Mudge, P.L.; Campbell, D.I.; Woodward, S.L.; Goodrich, J.P.; Wall, A.M.; Kirschbaum, M.U.F.; Schipper, L.A. Carbon Balance of an Intensively Grazed Temperate Dairy Pasture over Four Years. Agric. Ecosyst. Environ. 2015, 206, 10–20. [Google Scholar] [CrossRef]
- Feigenwinter, I.; Hörtnagl, L.; Zeeman, M.J.; Eugster, W.; Fuchs, K.; Merbold, L.; Buchmann, N. Large Inter-Annual Variation in Carbon Sink Strength of a Permanent Grassland over 16 Years: Impacts of Management Practices and Climate. Agric. For. Meteorol. 2023, 340, 109613. [Google Scholar] [CrossRef]
- Baldocchi, D.; Chu, H.; Reichstein, M. Inter-Annual Variability of Net and Gross Ecosystem Carbon Fluxes: A Review. Agric. For. Meteorol. 2018, 249, 520–533. [Google Scholar] [CrossRef]
- Ammann, C.; Neftel, A.; Jocher, M.; Fuhrer, J.; Leifeld, J. Effect of Management and Weather Variations on the Greenhouse Gas Budget of Two Grasslands during a 10-Year Experiment. Agric. Ecosyst. Environ. 2020, 292, 106814. [Google Scholar] [CrossRef]
- Rogger, J.; Hörtnagl, L.; Buchmann, N.; Eugster, W. Carbon Dioxide Fluxes of a Mountain Grassland: Drivers, Anomalies and Annual Budgets. Agric. For. Meteorol. 2022, 314, 108801. [Google Scholar] [CrossRef]
- Carvalho, P.C.D.F.; Batello, C. Access to Land, Livestock Production and Ecosystem Conservation in the Brazilian Campos Biome: The Natural Grasslands Dilemma. Livest. Sci. 2009, 120, 158–162. [Google Scholar] [CrossRef]
- Carnevalli, R.; Silva, S.C.; Bueno, A.A.O.; Uebele, M.C.; Bueno, F.O.; Hodgson, J.; Silva, G.N.; Morais, J. Herbage Production and Grazing Losses in Panicum Maximum Cv. Mombaça under Four Grazing Managements. Trop. Grassl. 2006, 40, 165–176. [Google Scholar]
- Cezimbra, I.M.; de Albuquerque Nunes, P.A.; de Souza Filho, W.; Tischler, M.R.; Genro, T.C.M.; Bayer, C.; Savian, J.V.; Bonnet, O.J.F.; Soussana, J.-F.; de Faccio Carvalho, P.C. Potential of Grazing Management to Improve Beef Cattle Production and Mitigate Methane Emissions in Native Grasslands of the Pampa Biome. Sci. Total Environ. 2021, 780, 146582. [Google Scholar] [CrossRef] [PubMed]
- de Souza Filho, W.; de Albuquerque Nunes, P.A.; Barro, R.S.; Kunrath, T.R.; de Almeida, G.M.; Genro, T.C.M.; Bayer, C.; de Faccio Carvalho, P.C. Mitigation of Enteric Methane Emissions through Pasture Management in Integrated Crop-Livestock Systems: Trade-Offs between Animal Performance and Environmental Impacts. J. Clean. Prod. 2019, 213, 968–975. [Google Scholar] [CrossRef]
- Vasconcelos, K.; Farinha, M.; Bernardo, L.; Lampert, V.D.N.; Gianezini, M.; da Costa, J.S.; Filho, A.S.; Genro, T.C.M.; Ruviaro, C.F. Livestock-Derived Greenhouse Gas Emissions in a Diversified Grazing System in the Endangered Pampa Biome, Southern Brazil. Land Use Policy 2018, 75, 442–448. [Google Scholar] [CrossRef]
- Gatti, L.V.; Basso, L.S.; Miller, J.B.; Gloor, M.; Gatti Domingues, L.; Cassol, H.L.G.; Tejada, G.; Aragão, L.E.O.C.; Nobre, C.; Peters, W.; et al. Amazonia as a Carbon Source Linked to Deforestation and Climate Change. Nature 2021, 595, 388–393. [Google Scholar] [CrossRef]
- Skidmore, M.E.; Moffette, F.; Rausch, L.; Christie, M.; Munger, J.; Gibbs, H.K. Cattle Ranchers and Deforestation in the Brazilian Amazon: Production, Location, and Policies. Glob. Environ. Chang. 2021, 68, 102280. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Warm Season | No. of Days | Pasture | Cattle | |||
|---|---|---|---|---|---|---|
| Average Height (cm) | Total Dry Mass (kg ha−1) | Leaves (%) | Average Occupancy (kg LW ha−1) | Weight Gain (kg LW ha−1) | ||
| 23/10/2015–22/03/2016 | 152 | 22.7 | 4197.2 | 40 | 885.9 | 136.8 |
| 29/09/2016–30/03/2017 | 183 | 33.2 | 4951.1 | 40 | 923.4 | 146.4 |
| 25/10/2017–22/03/2018 | 149 | 30.4 | 4777.0 | 35 | 1262.4 | 119.2 |
| 29/09/2018–19/03/2019 | 172 | 29.2 | 3428.8 | 55 | 826.3 | 154.8 |
| 06/11/2019–21/03/2020 | 137 | 29.2 | 2858.6 | 45 | 822.7 | 95.9 |
| Periods | No. of Days | Rg (W m−2) | Ta (°C) [No. of Frost] | Prec (mm) | NEE (g C m−2) | GPP (g C m−2) | Reco (g C m−2) |
|---|---|---|---|---|---|---|---|
| Warm season | |||||||
| 23/10/2015–22/03/2016 | 152 | 224.5 | 23.1 | 933.8 | −164.5 | 1664.9 | 1500.4 |
| 29/09/2016–30/03/2017 | 183 | 238.6 | 22.7 | 1420.8 | −209.9 | 2245.3 | 2035.4 |
| 25/10/2017–22/03/2018 | 149 | 270.0 | 23.3 | 560.0 | −220.5 | 1922.2 | 1701.7 |
| 29/09/2018–19/03/2019 | 172 | 245.1 | 23.5 | 941.4 | −290.8 | 2267.0 | 1976.2 |
| 06/11/2019–21/03/2020 | 137 | 280.0 | 24.0 | 495.2 | −220.3 | 1867.4 | 1647.1 |
| 29/09/2020–21/03/2021 | 174 | 259.7 | 22.5 | 409.6 | −348.8 | 2049.9 | 1701.1 |
| Cold season | |||||||
| 12/03/2015–22/10/2015 | 225 | 142.6 | 17.5 [12] | 1434.0 | −58.3 | 1354.0 | 1295.7 |
| 23/03/2016–28/09/2016 | 190 | 132.6 | 15.3 [11] | 663.4 | 13.3 | 834.4 | 847.7 |
| 31/03/2017–24/10/2017 | 208 | 135.9 | 17.9 [4] | 1210.4 | −49.5 | 1333.1 | 1283.5 |
| 23/03/2018–30/09/2018 | 192 | 126.4 | 16.5 [16] | 904.6 | 189.5 | 989.4 | 1178.9 |
| 20/03/2019–05/11/2019 | 231 | 148.8 | 17.9 [18] | 1259.2 | 9.3 | 1425.5 | 1434.9 |
| 22/03/2020–28/09/2020 | 191 | 147.8 | 16.3 [15] | 654.8 | 128.5 | 828.3 | 956.8 |
| 22/03/2021–22/09/2021 | 185 | 139.2 | 15.6 [7] | 722.6 | NA | NA | NA |
| Annual | |||||||
| 2015 | 365 | 174.6 | 19.6 | 2347.8 | −385.3 ± 19 | 2838.5 | 2453.2 |
| 2016 | 366 | 186.0 | 18.9 | 1722.8 | −120.7 ± 7 | 2902.7 | 2781.9 |
| 2017 | 365 | 186.3 | 20.3 | 2114.6 | −280.2 ± 31 | 3418.4 | 3138.3 |
| 2018 | 365 | 188.7 | 19.6 | 1721.6 | −122.5 ± 8 | 3236.9 | 3114.4 |
| 2019 | 365 | 188.8 | 20.2 | 1853.6 | −82.0 ± 5 | 3314.7 | 3232.7 |
| 2020 | 366 | 205.2 | 19.4 | 1130.4 | −254.7 ± 14 | 2853.5 | 2598.7 |
| Variables | Monthly Scale | Annual Scale | Meteorological Seasons | Management Seasons | ||||
|---|---|---|---|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | Warm | Cold | |||
| Rg | −0.8 | 0.3 | 0.2 | −0.1 | 0.2 | −0.2 | −0.3 | −0.3 (ns) |
| Ta | −0.6 | −0.1 | −0.1 | −0.4 | 0.6 (ns) | −0.7 (ns) | −0.4 | −0.5 (ns) |
| VPD | −0.6 | −0.2 | 0.5 | −0.3 | −0.2 (ns) | −0.6 (ns) | 0.0 | −0.5 (ns) |
| Ts | −0.5 | −0.6 | −0.8 | 0.4 | 0.8 (ns) | −0.1 (ns) | 0.0 | −0.1 (ns) |
| SWC | 0.2 | 0.2 | 0.4 | −0.5 | 0.1 (ns) | −0.4 (ns) | −0.3 | −0.3 (ns) |
| Prec | −0.1 | −0.4 | 0.1 | 0.2 | −0.5 | −0.2 | −0.1 | −0.6 |
| Pasture height | - | - | - | - | - | −0.6 | - | |
| Total dry mass | - | - | - | - | - | 0.4 | - | |
| Weight gain | - | - | - | - | - | −0.3 | - | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberti, D.R.; Mergen, A.; Gotuzzo, R.A.; Veeck, G.P.; Bremm, T.; Marin, L.; de Quadros, F.L.F.; Jacques, R.J.S. Sustainability in Natural Grassland in the Brazilian Pampa Biome: Livestock Production with CO2 Absorption. Sustainability 2024, 16, 3672. https://doi.org/10.3390/su16093672
Roberti DR, Mergen A, Gotuzzo RA, Veeck GP, Bremm T, Marin L, de Quadros FLF, Jacques RJS. Sustainability in Natural Grassland in the Brazilian Pampa Biome: Livestock Production with CO2 Absorption. Sustainability. 2024; 16(9):3672. https://doi.org/10.3390/su16093672
Chicago/Turabian StyleRoberti, Débora Regina, Alecsander Mergen, Ricardo Acosta Gotuzzo, Gustavo Pujol Veeck, Tiago Bremm, Luciana Marin, Fernando Luiz Ferreira de Quadros, and Rodrigo Josemar Seminoti Jacques. 2024. "Sustainability in Natural Grassland in the Brazilian Pampa Biome: Livestock Production with CO2 Absorption" Sustainability 16, no. 9: 3672. https://doi.org/10.3390/su16093672