2.1. Hybrid Evaluation
Results demonstrated that there were differences between hybrids for many phenomorphological traits and competitive factors, that some phenomorphological traits accounted for differences in competitive ability in the hybrids, and that, for some of these traits, hybrid performance can be predicted based on parentage.
Consistent differences between hybrids existed for most phenomorphological traits and competitive factors. Hybrid effects existed for all traits except ear length, ear length stability, and ear width stability (
Table 1,
Table 2). Hybrid-by-environment (H × E) effects existed for 100 kernel mass, ears per plant, early leaf area, late leaf area, average leaf height, early height, mid-season height, and yield stability. However, Spearman rank-change H × E effects existed only for ears per plant and average leaf height. In all traits that had hybrid effects, GCA effects also existed, except for 100 kernel mass stability. In all traits that had hybrid effects, SCA effects also existed, except for 100 kernel mass stability, average leaf height, average number of tillers, mid-season height, ear number stability, and early sorghum biomass.
Table 1.
Significance of mean squares from analysis of variance for phenological and morphological traits of 19 hybrids from a seven line diallel and Jubilee, a check, measured in Arlington, WI 2007, and West Madison, WI 2008.
Table 1.
Significance of mean squares from analysis of variance for phenological and morphological traits of 19 hybrids from a seven line diallel and Jubilee, a check, measured in Arlington, WI 2007, and West Madison, WI 2008.
| Source | Hybrid | Environment | Hybrid × Environment | Environment × Treatment | GCA | SCA | Treatment | Hybrid × Treatment |
|---|
| Yield | ** | ** | | | ** | ** | ** | |
| 100 kernel mass | ** | | ** | * | ** | | | * |
| Ear length | | * | | | ** | | * | |
| Ear Width | ** | | | | ** | * | ** | |
| Ears per plant | ** | | ** | * | ** | * | ** | * |
| GDD to anthesis | ** | ** | | | ** | ** | | |
| Early-season leaf area | ** | ** | ** | | ** | * | | |
| Late-season leaf area | ** | ** | ** | ** | ** | ** | ** | |
| Average leaf height | ** | ** | ** | | ** | | | |
| Early-season plant height | ** | ** | ** | | ** | * | | |
| Mid-season plant height | ** | ** | ** | | ** | ** | | |
| Late-season plant height | ** | | | | ** | ** | | |
| Tiller number | ** | | | | ** | | ** | * |
| Above-ear leaf angle | ** | * | | | ** | * | ** | |
| Yield stability | ** | * | * | ** | ** | ** | ** | ** |
| Ear length stability | | * | | ** | * | | ** | ** |
| Ear width stability | | | | ** | * | | ** | ** |
| 100 kernel mass stability | * | * | | ** | | | ** | ** |
| Ear number stability | * | * | | ** | ** | | ** | ** |
| Early sorghum biomass | * | ** | | ** | ** | | ** | ** |
| Late sorghum biomass | ** | | | ** | ** | | ** | ** |
Table 2.
Means for phenological and morphological traits for 19 hybrids from a seven line diallel and Jubilee, a check, averaged across one or more of the following locations: Arlington WI, 2007, and West Madison, WI, 2008.
Table 2.
Means for phenological and morphological traits for 19 hybrids from a seven line diallel and Jubilee, a check, averaged across one or more of the following locations: Arlington WI, 2007, and West Madison, WI, 2008.
| Hybrid | Yield | 100 kernelmass | Earlength | Earwidth | Ear number | GDD toanthesis | Early-seasonleaf area | Late-season leaf area |
|---|
| All (kg ha−1) | All (g) | All (cm) | All (cm) | A07 | WM08 | All | All (cm) | All (cm) |
|---|
| Ia5125 × C68 | 4842 | 18.4 | 14.7 | 4.4 | 0.96 | 0.93 | 975 | 4842 | 18.4 |
| Ia5125 × P39 | 3605 | 14.6 | 14.0 | 4.1 | 0.92 | 0.98 | 972 | 3605 | 14.6 |
| Ia5125 × P51 | 4179 | 16.6 | 13.4 | 3.9 | 1.03 | 1.03 | 936 | 4179 | 16.6 |
| Ia5125 × We10 | 2833 | 18.3 | 12.8 | 4.0 | 0.78 | 0.65 | 759 | 2833 | 18.3 |
| Ia5125 × IL101t | 4162 | 19.0 | 14.6 | 3.9 | 0.97 | 0.98 | 947 | 4162 | 19.0 |
| Ia5125 × C40 | 4685 | 23.4 | 15.0 | 4.2 | 0.91 | 1.02 | 907 | 4685 | 23.4 |
| C68 × P39 | 4059 | 17.3 | 14.3 | 3.8 | 0.95 | 0.95 | 1045 | 4059 | 17.3 |
| C68 × P51 | 4394 | 19.3 | 13.6 | 3.8 | 1.10 | 0.96 | 1150 | 4394 | 19.3 |
| C68 × We10 | 4089 | 21.4 | 13.9 | 4.0 | 0.92 | 0.93 | 1022 | 4089 | 21.4 |
| C68 × IL101t | 4681 | 21.1 | 14.9 | 3.9 | 0.98 | 0.98 | 1138 | 4681 | 21.1 |
| C68 × C40 | 4742 | 24.4 | 14.6 | 4.1 | 0.88 | 0.95 | 1162 | 4742 | 24.4 |
| P39 × P51 | 2553 | 14.2 | 12.6 | 3.3 | 0.92 | 1.05 | 967 | 2553 | 14.2 |
| P39 × We10 | 2663 | 15.8 | 12.9 | 3.6 | 0.78 | 0.88 | 1033 | 2663 | 15.8 |
| P39 × IL101t | 3389 | 16.5 | 13.8 | 3.4 | 1.04 | 1.03 | 970 | 3389 | 16.5 |
| P39 × C40 | 3834 | 18.7 | 14.3 | 3.6 | 0.95 | 1.03 | 1264 | 3834 | 18.7 |
| P51 × We10 | 3549 | 20.0 | 13.8 | 3.6 | 0.98 | 0.97 | 1055 | 3549 | 20.0 |
| P51 × IL101t | 3770 | 21.0 | 13.2 | 3.4 | 1.17 | 1.07 | 1108 | 3770 | 21.0 |
| C40 × P51 | 3448 | 21.8 | 14.4 | 3.5 | 0.92 | 0.95 | 1122 | 3448 | 21.8 |
| C40 × We10 | 4134 | 24.6 | 15.3 | 4.0 | 0.93 | 0.92 | 1165 | 4134 | 24.6 |
| Jubilee | 3659 | 14.1 | 14.5 | 3.9 | 0.97 | 1.10 | 731 | 3659 | 14.1 |
| CV | 0.15 | 0.10 | 0.16 | 0.06 | 0.19 | 0.13 | 0.18 | 0.15 | 0.10 |
| LSD (0.05) | 482 | 1.7 | 1.9 | 0.2 | 0.15 | 0.17 | 152 | 482 | 1.7 |
| Hybrid | Early-season plant height | Mid-season plant height | Late-season plant height | Upper leaf angle | 100 kernel mass stability | Ears per plant stability | Early-season sorg. mass | Late-seasonsorg. mass |
| All (cm) | All (cm) | All (cm) | All (°) | All (%) | All (%) | All (g) | All (g) |
| Ia5125 × C68 | 14.7 | 4.4 | 4.4 | 58 | 125 | 112 | 13 | 53 |
| Ia5125 × P39 | 14.0 | 4.1 | 4.1 | 76 | 91 | 86 | 16 | 72 |
| Ia5125 × P51 | 13.4 | 3.9 | 3.9 | 64 | 95 | 90 | 14 | 71 |
| Ia5125 × We10 | 12.8 | 4.0 | 4.0 | 58 | 102 | 63 | 18 | 68 |
| Ia5125 × IL101t | 14.6 | 3.9 | 3.9 | 56 | 96 | 99 | 14 | 71 |
| Ia5125 × C40 | 15.0 | 4.2 | 4.2 | 56 | 97 | 97 | 15 | 66 |
| C68 × P39 | 14.3 | 3.8 | 3.8 | 62 | 92 | 79 | 15 | 74 |
| C68 × P51 | 13.6 | 3.8 | 3.8 | 53 | 94 | 83 | 13 | 67 |
| C68 × We10 | 13.9 | 4.0 | 4.0 | 46 | 98 | 87 | 14 | 68 |
| C68 × IL101t | 14.9 | 3.9 | 3.9 | 46 | 106 | 94 | 12 | 65 |
| C68 × C40 | 14.6 | 4.1 | 4.1 | 44 | 99 | 69 | 14 | 50 |
| P39 × P51 | 12.6 | 3.3 | 3.3 | 69 | 73 | 64 | 17 | 74 |
| P39 × We10 | 12.9 | 3.6 | 3.6 | 60 | 97 | 67 | 16 | 77 |
| P39 × IL101t | 13.8 | 3.4 | 3.4 | 61 | 98 | 78 | 17 | 78 |
| P39 × C40 | 14.3 | 3.6 | 3.6 | 65 | 108 | 83 | 15 | 65 |
| P51 × We10 | 13.8 | 3.6 | 3.6 | 57 | 122 | 77 | 16 | 75 |
| P51 × IL101t | 13.2 | 3.4 | 3.4 | 52 | 96 | 83 | 15 | 70 |
| C40 × P51 | 14.4 | 3.5 | 3.5 | 54 | 100 | 84 | 15 | 74 |
| C40 × We10 | 15.3 | 4.0 | 4.0 | 54 | 102 | 93 | 14 | 58 |
| Jubilee | 14.5 | 3.9 | 3.9 | 57 | 84 | 74 | 18 | 82 |
| CV | 0.16 | 0.06 | 0.06 | 0.10 | 0.21 | 0.23 | 0.20 | 0.17 |
| LSD (0.05) | 1.9 | 0.2 | 0.2 | 4.6 | 27 | 26 | 3.4 | 13 |
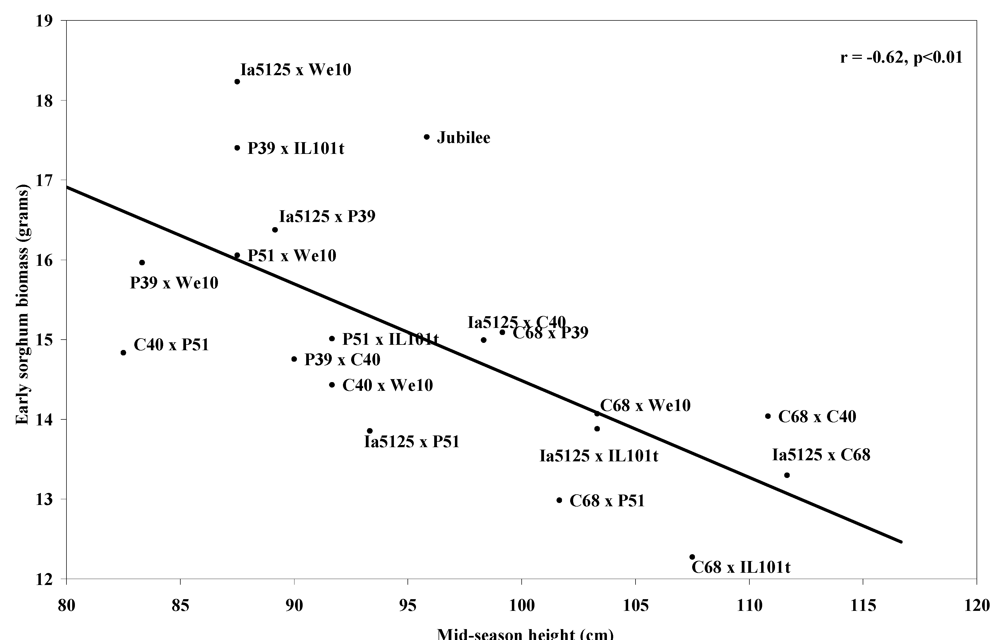
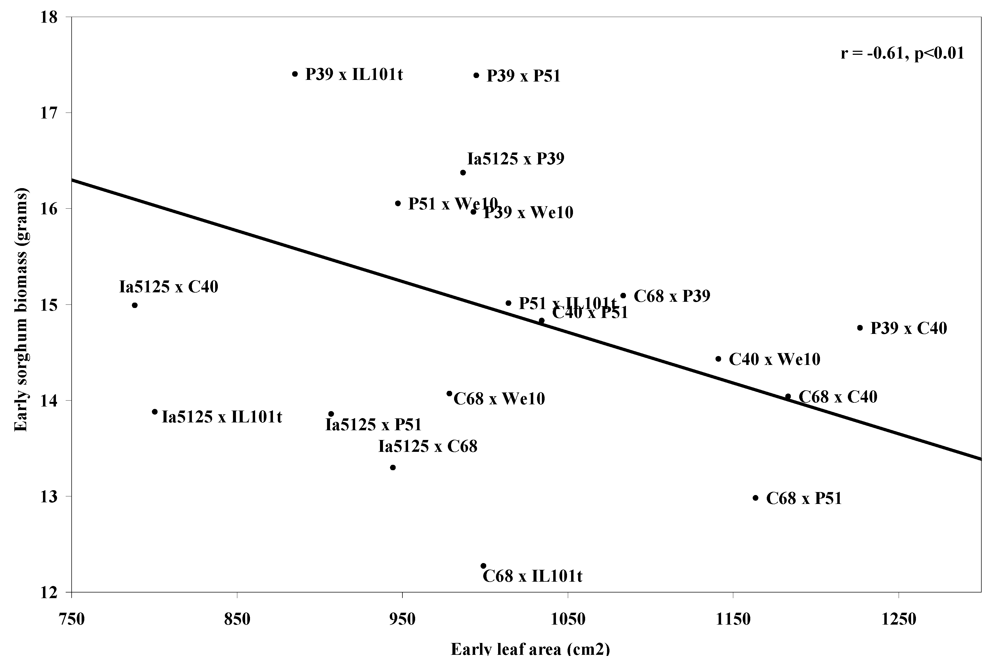
Correlations were seen between the sorghum-free plot means of many of the phenomorphological traits and the three factors used to assess competitive ability: early sorghum biomass, late sorghum biomass, and yield stability (
Table 3). The three traits most correlated with reduced early-season sorghum mass were mid-season plant height (r = −0.62) (
Figure 1), early-season leaf area (r = −0.61) (
Figure 2), and yield (r = −0.60) (
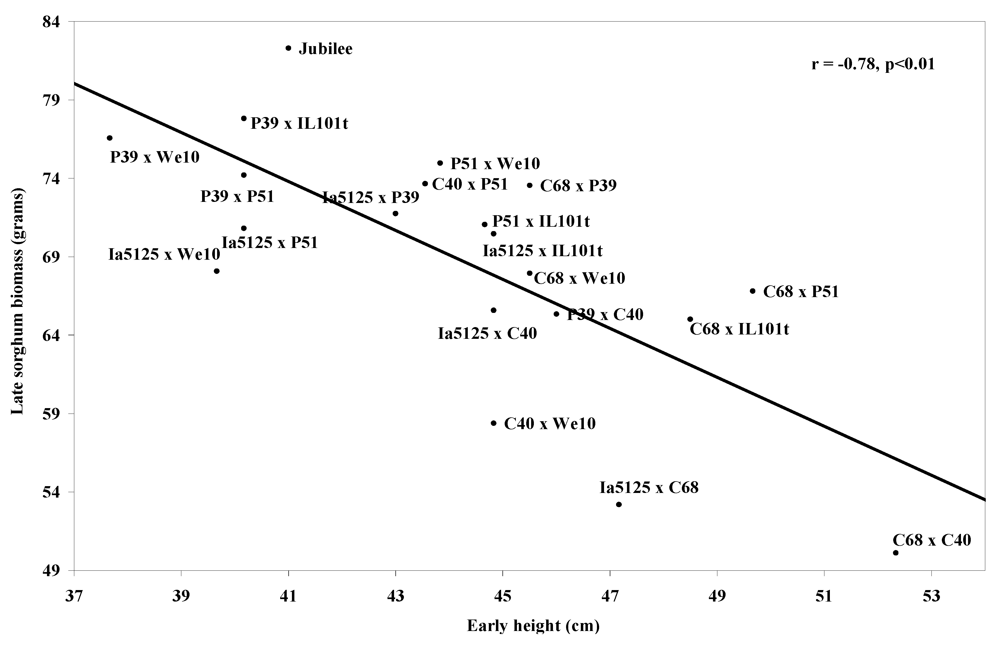
Table 3). Stepwise regression found that early-season plant height and early-season leaf area combined accounted for 67% of the variation present in early-season sorghum mass (p < 0.01), and additional traits did not significantly improve the model (data not shown). Early-season plant height (r = −0.78) (
Figure 3), 100 kernel mass (r = −0.62), and yield (r = −0.61) were most correlated with reduced late-season sorghum biomass. Early-season plant height accounted for 61% of variation in late-season sorghum biomass, with additional traits not significantly improving the regression model. Mid-season plant height (r = 0.58) (
Figure 4), late-season plant height (r = 0.56), and upper leaf angle (r = −0.53) were the three factors most correlated with yield stability. Mid-season plant height accounted for 33% of variation in yield stability, with additional traits not significantly improving the regression model.
Table 3.
Phenotypic correlation coefficients between the means of 19 hybrids from a seven line diallel and Jubilee, a check, for all morphological and phenological traits and the means of traits related to weed competitiveness, measured in Arlington WI, 2007 and West Madison, WI, 2008. Means for all traits except early-season sorghum mass and late-season sorghum mass taken only from non-sorghum control blocks.
Table 3.
Phenotypic correlation coefficients between the means of 19 hybrids from a seven line diallel and Jubilee, a check, for all morphological and phenological traits and the means of traits related to weed competitiveness, measured in Arlington WI, 2007 and West Madison, WI, 2008. Means for all traits except early-season sorghum mass and late-season sorghum mass taken only from non-sorghum control blocks.
| Trait | Yield Stability | Early-season sorghum mass | Late-season sorghum mass |
|---|
| Yield | 0.51* | −0.60** | −0.61** |
| 100 kernel mass | -- | -- | −0.62** |
| Ear length | -- | -- | -- |
| Ear width | 0.49* | -- | −0.49* |
| GDD to anthesis | -- | -- | -- |
| Early-season leaf area | -- | −0.61** | −0.50* |
| Late-season leaf area | -- | -- | -- |
| Early-season corn plant height | 0.48* | −0.59** | −0.78** |
| Mid-season corn plant height | 0.57** | −0.62** | −0.57** |
| Late-season corn plant height | 0.56* | -- | −0.45* |
| Corn tiller number | -- | -- | -- |
| Upper leaf angle | −0.53* | -- | -- |
Figure 1.
Means of sorghum-free control plot means of mid-season corn plant height vs. means of early sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 1.
Means of sorghum-free control plot means of mid-season corn plant height vs. means of early sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 2.
Means of sorghum-free control plot means of corn plant early leaf area vs.means of early sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 2.
Means of sorghum-free control plot means of corn plant early leaf area vs.means of early sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 3.
Means of sorghum-free control plot means of corn plant early height vs.means of late sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 3.
Means of sorghum-free control plot means of corn plant early height vs.means of late sorghum biomass, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 4.
Means of sorghum-free control plot means of corn plant mid-season height vs.means of yield stability, at Arlington, WI 2007 and West Madison, WI 2008.
Figure 4.
Means of sorghum-free control plot means of corn plant mid-season height vs.means of yield stability, at Arlington, WI 2007 and West Madison, WI 2008.
Large differences were seen in the potential combining abilities for the inbred parents both for the factors measuring competitive ability, and for the phenomorphological traits that correlate with competitive ability. For example, averaged across all environments, the range of GCA for yield,
i.e., the difference between the inbred with the highest GCA for yield and the inbred with the most negative GCA for yield, was 1341 kg ha
−1 (
Table 4), the range for early leaf area GCA was 256 cm
2 (
Table 4), the range for late plant height GCA was 26 cm (
Table 5), the range for early height was 8.4 cm (
Table 5), the range for early sorghum biomass GCA was 3.0 grams (
Table 6), the range for late sorghum biomass was 26 grams (
Table 6), and the range for yield stability GCA was 21% (
Table 7). By comparing the mean squares of GCA to total hybrid mean squares (2MSGCA: 2MSGCA + MSSCA), it can be shown that most of the variation in competitive ability and related traits is predictable based on inbred GCA alone (
Table 8).
Table 4.
General and specific combining abilities for corn grain yield† in kg ha-1 (above diagonal) and corn early leaf area‡ in cm2 (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, average was calculated from Arlington, WI 2007 and West Madison, WI 2008.
Table 4.
General and specific combining abilities for corn grain yield† in kg ha-1 (above diagonal) and corn early leaf area‡ in cm2 (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, average was calculated from Arlington, WI 2007 and West Madison, WI 2008.
| | SCA | GCA |
|---|
| Ia5125 | C68 | P39 | P51 | We10 | IL101t | C40 |
|---|
| SCA | Ia5125 | | 52.61 | 156.82 | 372.51 | −747.65 | −44.28 | 209.99 | 204.18 |
| C68 | 37.10 | | 110.12 | 87.39 | 7.96 | −25.49 | −232.60 | 704.20 |
| P39 | 81.57 | −43.91 | | −412.51 | −77.61 | 23.51 | 199.66 | −636.58 |
| P51 | 28.69 | 43.20 | −91.41 | | 450.36 | 46.25 | −544.00 | −278.54 |
| We10 | −92.64 | −28.71 | 30.84 | 35.29 | | . | 366.95 | −504.13 |
| IL101t | 36.25 | 27.51 | −92.19 | 28.43 | . | | . | 121.10 |
| C40 | −90.96 | −35.19 | 115.11 | −44.19 | 55.23 | . | | 389.76 |
| GCA | −151.53 | 47.54 | −0.76 | 16.54 | −39.05 | 20.40 | 106.85 | |
Table 5.
General and specific combining abilities for corn plant late height† in cm (above diagonal) and corn plant early height‡ in cm (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007, West Madison, WI 2007, and West Madison, WI 2008.
Table 5.
General and specific combining abilities for corn plant late height† in cm (above diagonal) and corn plant early height‡ in cm (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007, West Madison, WI 2007, and West Madison, WI 2008.
| | SCA | GCA |
|---|
| Ia5125 | C68 | P39 | P51 | We10 | IL101t | C40 |
|---|
| SCA | Ia5125 | | 3.42 | 0.33 | 6.67 | −14.35 | −0.27 | 4.20 | 12.05 |
| C68 | 0.49 | | −1.70 | 1.38 | 2.95 | −1.80 | −4.25 | 16.25 |
| P39 | 1.16 | −0.31 | | −4.20 | 0.20 | 3.87 | 1.50 | −7.33 |
| P51 | −0.41 | 1.13 | −0.87 | | 5.29 | −1.80 | −7.34 | −8.41 |
| We10 | −0.97 | −0.19 | −0.61 | 0.58 | | . | 5.90 | −0.98 |
| IL101t | 1.18 | −0.87 | −1.37 | 1.06 | . | | . | −10.06 |
| C40 | −1.45 | −0.25 | 2.00 | −1.49 | 1.20 | . | | 12.05 |
| GCA | −0.73 | 4.82 | −3.59 | −0.94 | −2.54 | 0.39 | 2.65 | |
Table 6.
General and specific combining abilities for early sorghum dry weight† in g (above diagonal) and late sorghum dry weight‡ in g (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007 and West Madison, WI 2008.
Table 6.
General and specific combining abilities for early sorghum dry weight† in g (above diagonal) and late sorghum dry weight‡ in g (below diagonal), from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007 and West Madison, WI 2008.
| | SCA | GCA |
|---|
| Ia5125 | C68 | P39 | P51 | We10 | IL101t | C40 |
|---|
| SCA | Ia5125 | | −0.21 | −0.18 | −1.33 | 2.22 | −0.91 | 0.41 | 0.15 |
| C68 | −6.76 | | 0.31 | −0.42 | −0.17 | −0.75 | 1.23 | −1.62 |
| P39 | −0.74 | 5.83 | | 0.94 | −1.32 | 1.34 | −1.10 | 1.42 |
| P51 | 0.00 | 0.76 | −4.37 | | 0.14 | 0.32 | 0.35 | 0.05 |
| We10 | −0.21 | 4.41 | 0.51 | 0.57 | | . | −0.88 | 0.88 |
| IL101t | 2.23 | 0.97 | 1.25 | −4.45 | . | | . | −0.34 |
| C40 | 5.49 | −5.21 | −2.49 | 7.48 | −5.28 | . | | −0.54 |
| GCA | −1.72 | −6.50 | 6.02 | 4.37 | 1.84 | 2.36 | −6.37 | |
Table 7.
General and specific combining abilities for yield stability in percent†, from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007 and West Madison, WI 2008.
Table 7.
General and specific combining abilities for yield stability in percent†, from an incomplete half diallel among seven sweet corn inbreds, measured in Arlington, WI 2007 and West Madison, WI 2008.
| | SCA | GCA |
|---|
| C68 | P39 | P51 | We10 | IL101t | C40 |
|---|
| SCA | Ia5125 | 5.39 | 7.86 | 5.49 | −18.09 | 3.43 | −4.07 | 2.85 |
| C68 | | −3.64 | −3.08 | 3.30 | 2.72 | −4.69 | 11.56 |
| P39 | | | 0.74 | −3.26 | −2.64 | 0.95 | −9.64 |
| P51 | | | | 5.30 | −3.50 | −4.94 | 0.57 |
| We10 | | | | | . | 12.75 | −3.69 |
| IL101t | | | | | | . | −2.33 |
| C40 | | | | | | | 0.67 |
Table 8.
Predictability, calculated as the ratio of (2 MSGCA):(2 MSGCA + MSSCA), of GCA for traits measured on 19 hybrids from an incomplete half-diallel, measured in Arlington, WI 2007, West Madison, WI 2007, and West Madison, WI 2008.
Table 8.
Predictability, calculated as the ratio of (2 MSGCA):(2 MSGCA + MSSCA), of GCA for traits measured on 19 hybrids from an incomplete half-diallel, measured in Arlington, WI 2007, West Madison, WI 2007, and West Madison, WI 2008.
| Trait | Ratio of (2 MSGCA):(2 MSGCA + MSSCA) |
|---|
| Yield | 0.94 |
| Ear width | 0.98 |
| Number of ears | 0.89 |
| GDD to anthesis | 0.96 |
| Early leaf area | 0.91 |
| Late leaf area | 0.85 |
| Early-season height | 0.98 |
| Mid-season height | 0.98 |
| Late-season height | 0.96 |
| Upper leaf angle | 0.98 |
| Yield stability | 0.86 |
2.2. General Discussion
Some of the genotype by environment interaction effects seen in many of the traits may be due to the variable growing conditions experienced between environments. Rainfall and temperature may have contributed to creating these different growing environments.
Some previous conclusions about traits relating to competitive ability were supported by the results of this study. Although, unlike Roggenkamp
et al. [
20], upright leaves were not found to be correlated with WSA, upright leaves were found to be correlated with increased yield stability. While Makus [
21] and Williams
et al. [
12,
13] found greater WSA in late hybrids and Begna [
22] found greater WSA in early hybrids, no differences in WSA based on maturity were observed in this study. Although late-season leaf area was not found to be correlated to WSA, early-season leaf area was, which supports Woolley and Smith’s [
23] finding that leafy corn had greater WSA. In contrast to Makus’s [
21] finding that yield was reduced more in a taller cultivar than a shorter one, this study found that, in general, plant height correlated with both increased CT and increased WSA. This study had more entries present than some previous work [
8,
12,
13,
19,
20,
21,
22,
24,
25], allowing greater legitimacy to inferences made relating traits such as plant height to competitive ability.
If breeders are choose a few traits to use as a basis of selection for improved competitive ability, which should they be? Plant height, especially early- and mid-season plant height, showed the greatest correlation with both reduced sorghum biomass and yield stability (
Table 3,
Figure 1,
Figure 3 and
Figure 4), and that hybrid plant height was highly predictable based on the heights of the inbred parents (
Table 8). Correlation is not causation and plant height may simply be the measured trait that is linked to other unmeasured factors that cause increases in competitive ability. For example, a recent modeling study [
26] suggested that root characteristics may have been more important than canopy characteristics in the improvement of stress tolerance in commercial field corn hybrids. However, the results of this study suggest that regardless of the underlying factors responsible for competitive ability, selection for early- and mid-season plant height may lead to more competitive sweet corn.
Can a diallel analysis help breeders identify superior inbreds for competitive ability? Taking this study as an example, it can be shown that C68 stands out as a competitive inbred. It produced hybrids which on average had the highest yield, the highest yield stability, and the lowest early- and late-season sorghum biomass (
Table 4,
Table 6 and
Table 7). C68 also had the largest GCA in the positive direction for early- and late-season plant height and for vertical leaves (
Table 5, data not shown). C68 produced both the tallest and the most tolerant hybrids, which is consistent with So
et al.’s [
27] finding that plant height, along with other ‘late canopy and maturity’ factors, was most effective in explaining the differences in CT in sweet corn.
While C68 stands out as most competitive, P39 stands out as least competitive. P39 produced hybrids which had the lowest yield, yield stability and the most early- and late-season sorghum biomass (
Table 4,
Table 6 and
Table 7). P39 also contrasts with C68 in that P39 produced hybrids which were, on average, shortest in the early season and had the most horizontal leaves (
Table 5, data not shown).
Two key parameters influence the context of this study and should be considered when making inferences from this work about competitive ability in other systems. First, the interaction of two organisms depends partially on the relative fitness of each organism in a particular environment. The combination of the interactions that the two organisms have with each other, and the interactions that each have with the environment can make results obtained from evaluating the relationship between two organisms more sensitive to environmental variability than research focused on a single organism. In this study, competitive ability was most influenced by plant height. However, care must be taken when making any generalizations to other environments, especially those environments in which other resources, such as water or nutrients, may be more limited. In order to broaden the scope of the conclusions, similar experiments could be conducted in varied environments.
The second parameter that influences the context of this study is the choice of model weed. Forage sorghum is a good model due to its high competitiveness, accelerated growth and substantial biomass production and shading ability. It also offers the advantage of being able to be managed similarly to corn. However, can information gained from research with sorghum be applied to other weeds? Sorghum is a C4 grass similarly to corn. It has narrow, long leaves and has upright growth ability. But sorghum has a determinate flowering pattern, which limits plant height. Many broadleaf weeds have an indeterminate growth habit resulting in greater plasticity in plant height. Despite the differences between sorghum and broadleaf weeds, some inferences may be justifiable. Previous studies have found that, despite large differences in morphology, weed species can present similar competitive pressures. Moechnig
et al. [
28,
29] found that common lambsquarters (
Chenopodium album) and giant foxtail (
Setaria faberi) had similar competitive abilities in corn, depending on the environment.
An additional difference between sorghum and wild weeds is that sorghum will germinate consistently. It is useful for a model weed, as it allows uniform pressure to be created. However, weeds and other wild plants generally have extended germination periods, emerging throughout the season. Corn competitive traits that act on young weed seedlings would only be important against sorghum at the beginning of the season, but would continue to be important through-out the season when in competition with wild weeds. Conducting a similar experiment with weeds which had different emergence patterns would allow broader conclusions to be made.


{kind=link}
{kind=link}
{kind=link}
{kind=link}



