
Macroecological Patterns of Resilience Inferred from a Multinational, Synchronized Experiment



 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Lake Selection

{kind=link}
{kind=link}
{kind=link}
| Country | Lake | Coordinates | Altitude (m a.s.l.) |
|---|---|---|---|
| Sweden | Erken | 59°49′59″N | 11 |
| Estonia | Võrtsjärv | 58°12′17″N | 35 |
| Germany | Müggelsee | 52°26′0″N | 32 |
| Czech Republic | Vodňany | 49°09′14″N | 395 |
| Turkey | DSI Gölet | 39°52′38″N | 998 |
| Greece | Lysimachia | 38°33′40″N | 16 |
2.2. Experimental Design
2.3. Sampling
2.4. Statistical Analyses
2.4.1. Measuring Resilience
2.4.2. Statistical Comparisons
3. Results and Discussion
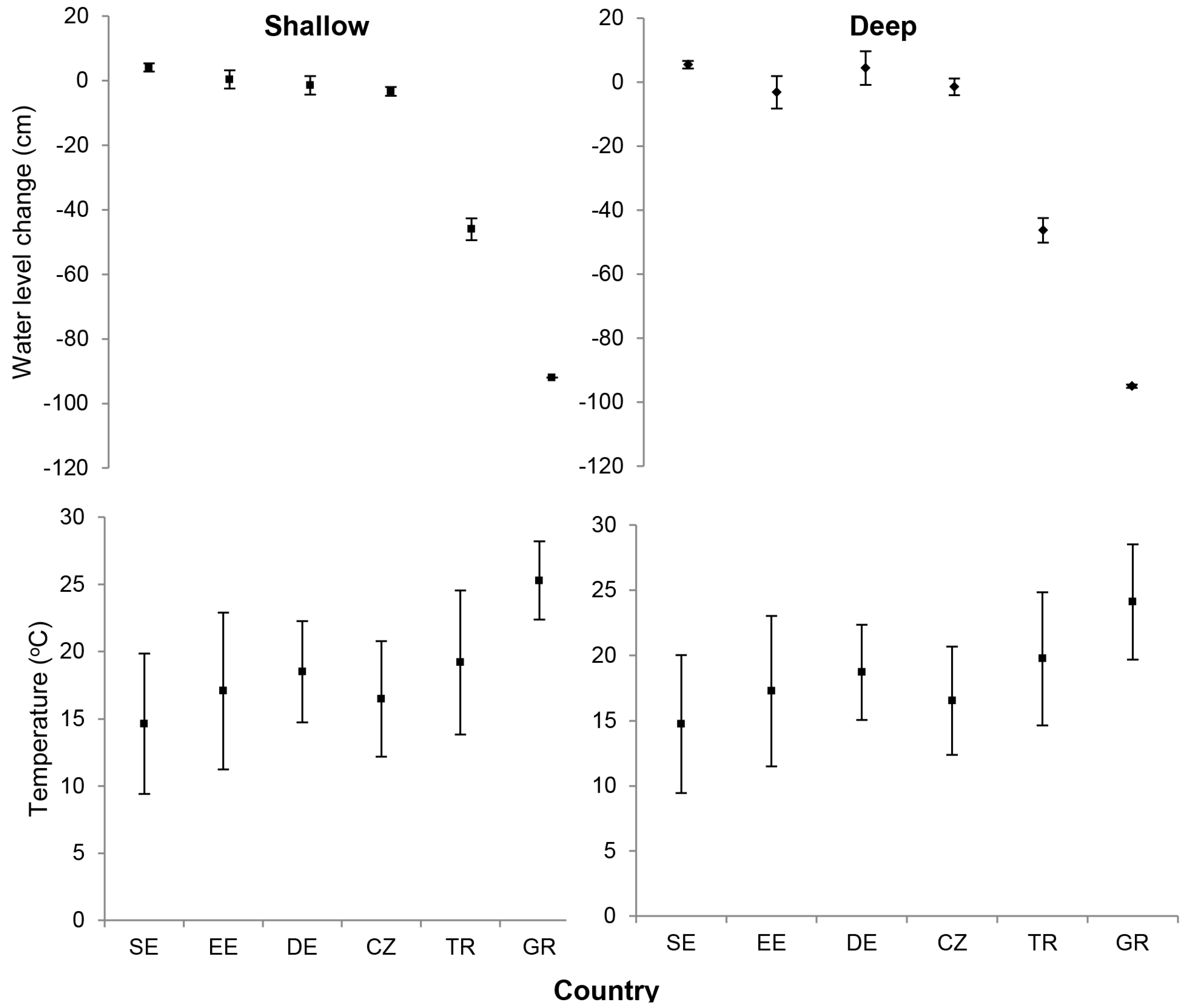
3.1. Abiotic Variables: Water Level Change and Temperature

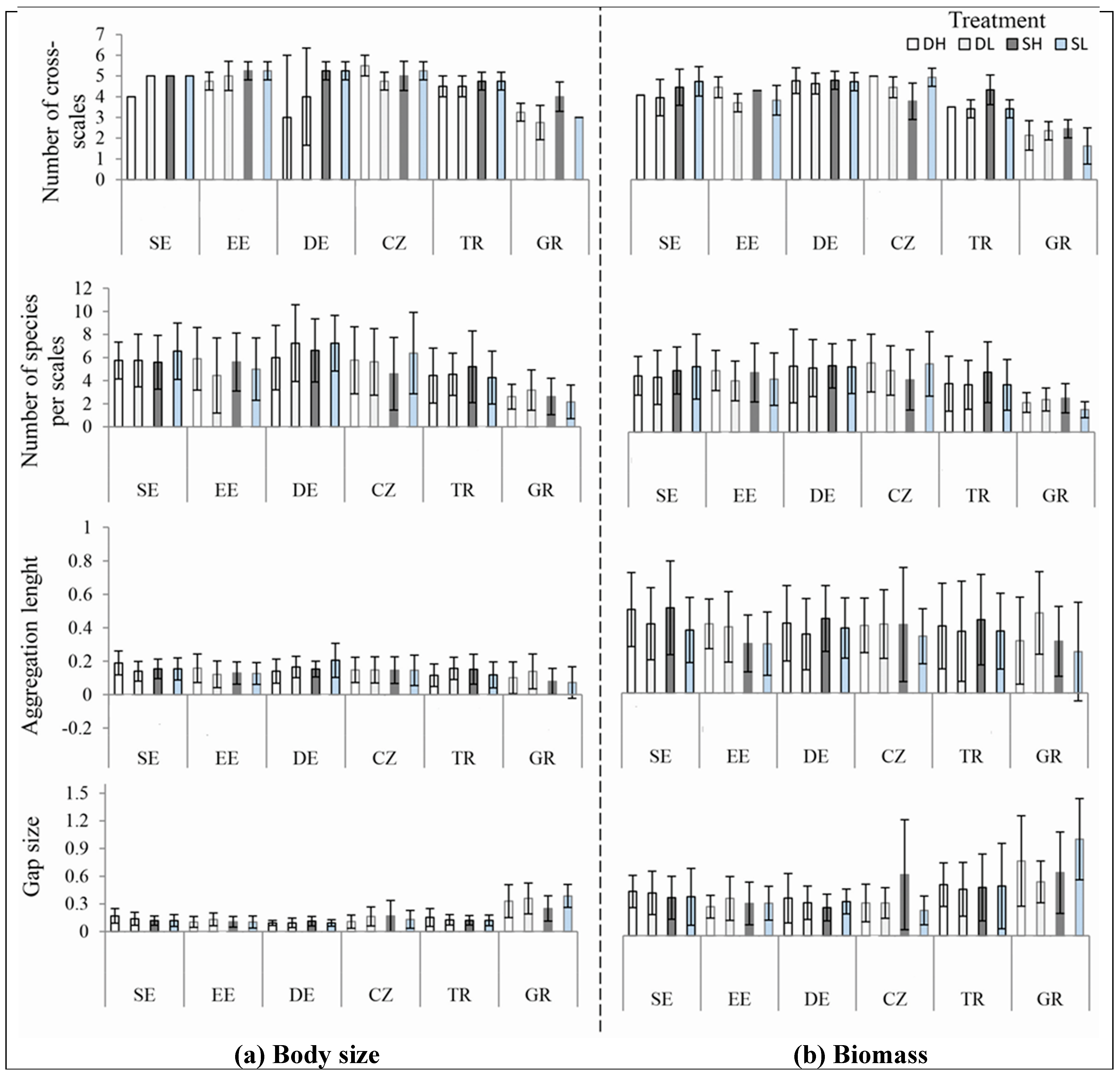
3.2. Univariate Analyses

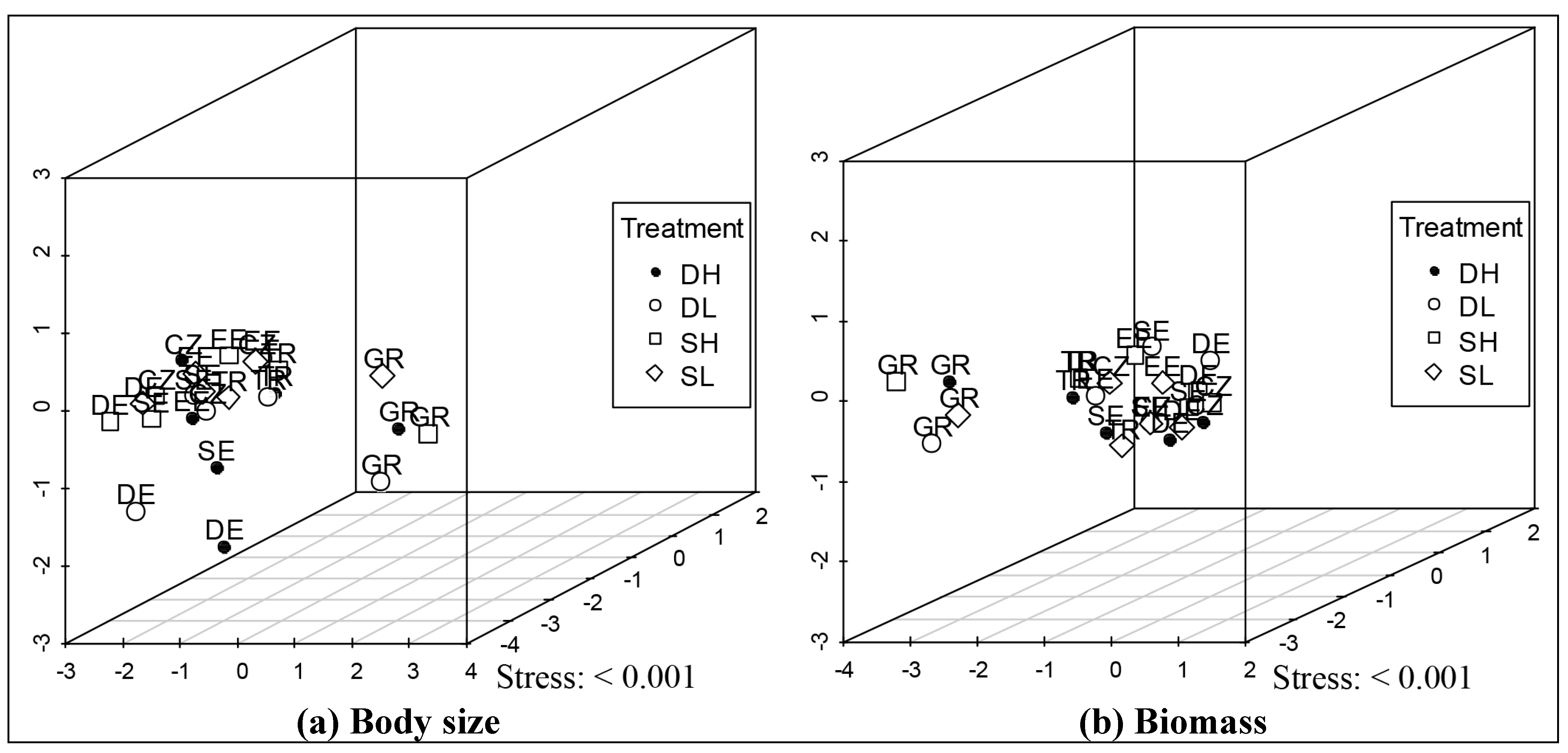
3.3. Multivariate Ordinations

3.4. Discussion
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- IPCC. Intergovernmental Panel on Climate Change, Climate Change: Contribution of Working Group iii to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007. [Google Scholar]
- Kernan, M.R.; Battarbee, R.W.; Moss, B. Climate Change Impacts on Freshwater Ecosystems; Wiley Online Library: Oxford, UK, 2010; Volume 314. [Google Scholar]
- Jeppesen, E.; Kronvang, B.; Meerhoff, M.; Sondergaard, M.; Hansen, K.M.; Andersen, H.E.; Lauridsen, T.L.; Liboriussen, L.; Beklioglu, M.; Ozen, A.; et al. Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J. Environ. Qual. 2009, 38, 1930–1941. [Google Scholar]
- Meyer, J.L.; Sale, M.J.; Mulholland, P.J.; Poff, N.L. Impacts of climate change on aquatic ecosystem functioning and health. J. Am. Water Resour. Assoc. 1999, 35, 1373–1386. [Google Scholar] [CrossRef]
- Wilhelm, S.; Adrian, R. Impact of summer warming on the thermal characteristics of a polymictic lake and consequences for oxygen, nutrients and phytoplankton. Freshw. Biol. 2008, 53, 226–237. [Google Scholar] [CrossRef]
- Arnell, N.W. The effect of climate change on hydrological regimes in europe: A continental perspective. Glob. Environ. Change-Human Policy Dimens. 1999, 9, 5–23. [Google Scholar] [CrossRef]
- Giorgi, F.; Bi, X.Q.; Pal, J. Mean, interannual variability and trends in a regional climate change experiment over europe. II: Climate change scenarios (2071–2100). Clim. Dyn. 2004, 23, 839–858. [Google Scholar]
- Giorgi, F.; Lionello, P. Climate change projections for the mediterranean region. Glob. Planet. Change 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kraft, C.E.; Wright, R.; He, X.; Soranno, P.A.; Hodgson, J.R. Resilience and resistance of a lake phosphorus cycle before and after food web manipulation. Am. Nat. 1992, 140, 781–798. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime shifts, resilience, and biodiversity in ecosystem management. Annu. Rev. Ecol. Syst. 2004, 35, 557–581. [Google Scholar]
- Smith, V.H.; Tilman, G.D.; Nekola, J.C. Eutrophication: Impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environ. Pollut. 1999, 100, 179–196. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Walker, B.; Holling, C.S.; Carpenter, S.R.; Kinzig, A. Resilience, adaptability and transformability in social-ecological systems. Ecol. Soc. 2004, 9. Article 5. [Google Scholar]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Scheffer, M.; Nes, E. Shallow lakes theory revisited: Various alternative regimes driven by climate, nutrients, depth and lake size. Hydrobiologia 2007, 584, 455–466. [Google Scholar] [CrossRef]
- Gyllstrom, M.; Hansson, L.A.; Jeppesen, E.; Garcia-Criado, F.; Gross, E.; Irvine, K.; Kairesalo, T.; Kornijow, R.; Miracle, M.R.; Nykanen, M.; et al. The role of climate in shaping zooplankton communities of shallow lakes. Limnol. Oceanogr. 2005, 50, 2008–2021. [Google Scholar]
- Layer, K.; Riede, J.O.; Hildrew, A.G.; Woodward, G. Food web structure and stability in 20 streams across a wide pH gradient. In Advances in Ecological Research: Ecological Networks; Woodward, G., Ed.; Elsevier Academic Press Inc: San Diego, CA, USA, 2010; Volume 42, pp. 265–299. [Google Scholar]
- Rawcliffe, R.; Sayer, C.D.; Woodward, G.; Grey, J.; Davidson, T.A.; Jones, J.I. Back to the future: Using palaeolimnology to infer long-term changes in shallow lake food webs. Freshw. Biol. 2010, 55, 600–613. [Google Scholar] [CrossRef]
- Meerhoff, M.; Teixeira-de Mello, F.; Kruk, C.; Alonso, C.; Gonzalez-Bergonzoni, I.; Pacheco, J.P.; Lacerot, G.; Arim, M.; Beklioglu, M.; Brucet, S.; et al. Environmental warming in shallow lakes: A review of potential changes in community structure as evidenced from space-for-time substitution approaches. In Advances in Ecological Research; Jacob, U., Woodward, G., Eds.; Elsevier Academic Press Inc: San Diego, 2012; Volume 46, pp. 259–349. [Google Scholar]
- Pickett, S.T. Space-for-time substitution as an alternative to long-term studies. In Long-Term Studies in Ecology; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Moss, B.; Stephen, D.; Balayla, D.M.; Bécares, E.; Collings, S.E.; Fernández-Aláez, C.; Fernández-Aláez, M.; Ferriol, C.; García, P.; Gomá, J.; et al. Continental-scale patterns of nutrient and fish effects on shallow lakes: Synthesis of a pan-european mesocosm experiment. Freshw. Biol. 2004, 49, 1633–1649. [Google Scholar]
- Stephen, D.; Balayla, D.M.; Becares, E.; Collings, S.E.; Fernandez-Alaez, C.; Fernandez-Alaez, M.; Ferriol, C.; Garcia, P.; Goma, J.; Gyllstrom, M.; et al. Continental-scale patterns of nutrient and fish effects on shallow lakes: Introduction to a pan-european mesocosm experiment. Freshw. Biol. 2004, 49, 1517–1524. [Google Scholar]
- Fraser, L.H.; Henry, H.A.L.; Carlyle, C.N.; White, S.R.; Beierkuhnlein, C.; Cahill, J.F.; Casper, B.B.; Cleland, E.; Collins, S.L.; Dukes, J.S.; et al. Coordinated distributed experiments: An emerging tool for testing global hypotheses in ecology and environmental science. Front. Ecol. Environ. 2012, 11, 147–155. [Google Scholar]
- Gannon, J.E.; Stemberger, R.S. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. Am. Fish. Soc. 1978, 97, 16–35. [Google Scholar]
- Attayde, J.L.; Bozelli, R.L. Assessing the indicator properties of zooplankton assemblages to disturbance gradients by canonical correspondence analysis. Can. J. Fish. Aquat. Sci. 1998, 55, 1789–1797. [Google Scholar] [CrossRef]
- Burns, C.W.; Galbraith, L.M. Relating planktonic microbial food web structure in lentic freshwater ecosystems to water quality and land use. J. Plankton Res. 2007, 29, 127–139. [Google Scholar] [CrossRef]
- Pinto-Coelho, R.; Bezerra-Neto, J. Effects of eutrophication on size and biomass of crustacean zooplankton in a tropical reservoir. Braz. J. Biol. 2005, 65, 325–338. [Google Scholar] [PubMed]
- Pinel-Alloul, B.; Méthot, G.; Verreault, G.; Vigneault, Y. Zooplankton species associations in Quebec lakes: Variation with abiotic factors, including natural and anthropogenic acidification. Can. J. Fish. Aquat. Sci. 1990, 47, 110–121. [Google Scholar] [CrossRef]
- Dodson, S.I.; Lillie, R.A.; Will-Wolf, S. Land use, water chemistry, aquatic vegetation, and zooplankton community structure of shallow lakes. Ecol. Appl. 2005, 15, 1191–1198. [Google Scholar] [CrossRef]
- Sousa, W.; Attayde, J.L.; Rocha, E.D.S.; Eskinazi-Sant’Anna, E.M. The response of zooplankton assemblages to variations in the water quality of four man-made lakes in semi-arid northeastern Brazil. J. Plankton Res. 2008, 30, 699–708. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Between Hazards of Starvation and Risk of Predation: The Ecology of Offshore Animals; International Ecology Institute: Hamburg, Germany, 2003. [Google Scholar]
- Angeler, D.G.; Allen, C.R.; Rojo, C.; Alvarez-Cobelas, M.; Rodrigo, M.A.; Sanchez-Carrillo, S. Inferring the relative resilience of alternative states. PloS One 2013, 8, e77338. [Google Scholar] [CrossRef] [PubMed]
- Angeler, D.G.; Göthe, E.; Johnson, R.K. Hierarchical dynamics of ecological communities: Do scales of space and time match? PloS One 2013, 8, e69174. [Google Scholar] [CrossRef] [PubMed]
- Holling, C.S. Cross-scale morphology, geometry, and dynamics of ecosystems. Ecol. Monogr. 1992, 62, 447–502. [Google Scholar] [CrossRef]
- Stow, C.; Allen, C.R.; Garmestani, A.S. Evaluating discontinuities in complex systems: Toward quantitative measures of resilience. Ecol. Soc. 2007, 12. Article 26. [Google Scholar]
- Allen, C.R.; Holling, C.S. Cross-scale structure and the generation of innovation and novelty in discontinuous complex systems. In Discontinuities in Ecosystems and Other Complex Systems; Allen, C.R., Holling, C.S., Eds.; Columbia University Press: New York, NY, USA, 2008; pp. 219–233. [Google Scholar]
- Nash, K.L.; Allen, C.R.; Angeler, D.G.; Barichievy, C.; Eason, T.; Garmestani, A.S.; Graham, N.A.J.; Granholm, D.; Knutson, M.; Nelson, R.J.; et al. Discontinuities, cross-scale patterns, and the organization of ecosystems. Ecology 2014, 95, 654–667. [Google Scholar]
- Nash, K.L.; Graham, N.A.J.; Wilson, S.K.; Bellwood, D.R. Cross-scale habitat structure drives fish body size distributions on coral reefs. Ecosystems 2013, 16, 478–490. [Google Scholar] [CrossRef]
- Allen, C.R.; Garmestani, A.S.; Havlicek, T.D.; Marquet, P.A.; Peterson, G.D.; Restrepo, C.; Stow, C.A.; Weeks, B.E. Patterns in body mass distributions: Sifting among alternative hypotheses. Ecol. Lett. 2006, 9, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.R.; Gunderson, L.; Johnson, A.R. The use of discontinuities and functional groups to assess relative resilience in complex systems. Ecosystems 2005, 8, 958–966. [Google Scholar] [CrossRef]
- Peterson, G.; Allen, C.R.; Holling, C.S. Ecological resilience, biodiversity, and scale. Ecosystems 1998, 1, 6–18. [Google Scholar] [CrossRef]
- Allen, C.R.; Forys, E.A.; Holling, C.S. Body mass patterns predict invasions and extinctions in transforming landscapes. Ecosystems 1999, 2, 114–121. [Google Scholar] [CrossRef]
- Carpenter, S.; Walker, B.; Anderies, J.M.; Abel, N. From metaphor to measurement: Resilience of what to what? Ecosystems 2001, 4, 765–781. [Google Scholar] [CrossRef]
- Havlicek, T.D.; Carpenter, S.R. Pelagic species size distributions in lakes: Are they discontinuous? Limnol. Oceanogr. 2001, 46, 1021–1033. [Google Scholar] [CrossRef]
- Raffaelli, D.; Hall, S.; Emes, C.; Manly, B. Constraints on body size distributions: An experimental approach using a small-scale system. Oecologia 2000, 122, 389–398. [Google Scholar] [CrossRef]
- Angeler, D.G.; Allen, C.R.; Johnson, R.K. Insight on invasions and resilience derived from spatiotemporal discontinuities of biomass at local and regional scales. Ecol. Soc. 2012, 17. Article 32. [Google Scholar]
- Carpenter, S.R.; Brock, W.A.; Hanson, P.C. Ecological and social dynamics in simple models of ecosystem management. Conserv. Ecol. 1999, 3. Article 4. [Google Scholar]
- Carpenter, S.R.; Ludwig, D.; Brock, W.A. Management of eutrophication for lakes subject to potentially irreversible change. Ecol. Appl. 1999, 9, 751–771. [Google Scholar] [CrossRef]
- Landkildehus, F.; Søndergaard, M.; Beklioğlu, M.; Adrian, R.; Angeler, D.G.; Hejzlar, J.; Papastergiadou, E.; Zingel, P.; Çakiroğlu, A.İ.; Scharfenberger, U.; et al. Climate change effects on shallow lakes: Design and preliminary results of a cross-european climate gradient mesocosm experiment. Est. J. Ecol. 2014, 63, 71–89. [Google Scholar]
- González Sagrario, M.A.; Jeppesen, E.; Gomà, J.; Søndergaard, M.; Jensen, J.P.; Lauridsen, T.; Landkildehus, F. Does high nitrogen loading prevent clear-water conditions in shallow lakes at moderately high phosphorus concentrations? Freshw. Biol. 2005, 50, 27–41. [Google Scholar] [CrossRef]
- Jeppesen, E.; Sondergaard, M.; Meerhoff, M.; Lauridsen, T.L.; Jensen, J.P. Shallow lake restoration by nutrient loading reduction—Some recent findings and challenges ahead. Hydrobiologia 2007, 584, 239–252. [Google Scholar] [CrossRef]
- Bottrell, H.H.; Duncan, A.; Gliwicz, Z.M.; Grygierek, E.; Herzig, A.; Hillbricht-Ilkowska, A.; Kurasawa, H.; Larsson, P.; Weglenska, T. A review of some problems in zooplankton production studies. Nor. J. Zool. 1976, 24, 419–456. [Google Scholar]
- Culver, D.A.; Boucherle, M.M.; Bean, D.J.; Fletcher, J.W. Biomass of freshwater crustacean zooplankton from length–weight regressions. Can. J. Fish. Aquat. Sci. 1985, 42, 1380–1390. [Google Scholar] [CrossRef]
- Dumont, H.; Velde, I.; Dumont, S. The dry weight estimate of biomass in a selection of cladocera, copepoda and rotifera from the plankton, periphyton and benthos of continental waters. Oecologia 1975, 19, 75–97. [Google Scholar] [CrossRef]
- McCauley, E. The estimation of the abundance and biomass of zooplankton in samples. In A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters, 2nd ed.; Blackwell Scientific Publications: Oxford, UK; London, UK; Edinburgh, UK; Boston, MA, USA; Melbourne, Australia, 1984; pp. 228–265. [Google Scholar]
- Michaloudi, E. Dry weights of the zooplankton of Lake Mikri Prespa (Macedonia, Greece). Belg. J. Zool. 2005, 135, 223–227. [Google Scholar]
- Rosen, R.A. Length-dry weight relationships of some freshwater zooplanktona. J. Freshw. Ecol. 1981, 1, 225–229. [Google Scholar] [CrossRef]
- Ruttner-Kolisko, A. Suggestions for biomass calculations of plankton rotifers. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1977, 8, 71–76. [Google Scholar]
- Vuille, T. Abundance, standing crop and production of microcrustacean population (Cladocera, Copepoda) in the littoral-zone of Lake Biel, Switzerland. Archiv Fur Hydrobiologie 1991, 123, 165–185. [Google Scholar]
- Bremner, A.P.; Taplin, R.H. Performance of localized regression tree splitting criteria on data with discontinuities. Aust. N. Z. J. Stat. 2004, 46, 367–381. [Google Scholar] [CrossRef]
- Chipman, H.A.; George, E.I.; McCulloch, R.E. Bayesian cart model search. J. Am. Stat. Assoc. 1998, 93, 935–948. [Google Scholar] [CrossRef]
- Chipman, H.A.; George, E.I.; McCulloch, R.E. Bayesian Cart. Available at: http://www.rob-mcculloch.org/code/CART/index.html/ (accessed on 15 January 2015).
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Legendre, P.; Legendre, L. Ordination in reduced space. In Developments in Environmental Modelling; Pierre, L., Louis, L., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 20, pp. 387–480. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Allen, C.R.; Saunders, D.A. Variability between scales: Predictors of nomadism in birds of an australian mediterranean-climate ecosystem. Ecosystems 2002, 5, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Stewart, R.I.; Dossena, M.; Bohan, D.A.; Jeppesen, E.; Kordas, R.L.; Ledger, M.E.; Meerhoff, M.; Moss, B.; Mulder, C.; Shurin, J.B. Mesocosm experiments as a tool for ecological climate-change research. Adv. Ecol. Res. 2013, 48, 71–181. [Google Scholar]
- Lake, P.S. Disturbance, patchiness, and diversity in streams. J. N. Am. Benthol. Soc. 2000, 19, 573–592. [Google Scholar] [CrossRef]
- Bond, N.R.; Lake, P.; Arthington, A.H. The impacts of drought on freshwater ecosystems: An australian perspective. Hydrobiologia 2008, 600, 3–16. [Google Scholar] [CrossRef]
- Legler, N.D.; Johnson, T.B.; Heath, D.D.; Ludsin, S.A. Water temperature and prey size effects on the rate of digestion of larval and early juvenile fish. Trans. Am. Fish. Soc. 2010, 139, 868–875. [Google Scholar] [CrossRef]
- Wang, N.; Xu, X.; Kestemont, P. Effect of temperature and feeding frequency on growth performances, feed efficiency and body composition of pikeperch juveniles (Sander lucioperca). Aquaculture 2009, 289, 70–73. [Google Scholar] [CrossRef]
- Petchey, O.L.; McPhearson, P.T.; Casey, T.M.; Morin, P.J. Environmental warming alters food-web structure and ecosystem function. Nature 1999, 402, 69–72. [Google Scholar] [CrossRef]
- Gasith, A.; Resh, V.H. Streams in mediterranean climate regions: Abiotic influences and biotic responses to predictable seasonal events. Annu. Rev. Ecol. Syst. 1999, 30, 51–81. [Google Scholar] [CrossRef]
- Hershkovitz, Y.; Gasith, A. Resistance, resilience, and community dynamics in mediterranean-climate streams. Hydrobiologia 2013, 719, 59–75. [Google Scholar] [CrossRef]
- Washington-Allen, R.A.; Ramsey, R.D.; West, N.E.; Norton, B.E. Quantification of the ecological resilience of drylands using digital remote sensing. Ecol. Soc. 2008, 13. Article 33. [Google Scholar]
- Heinrich, G.; Gobiet, A. The future of dry and wet spells in europe: A comprehensive study based on the ensembles regional climate models. Int. J. Climatol. 2012, 32, 1951–1970. [Google Scholar] [CrossRef]
- Trenberth, K.E. Changes in precipitation with climate change. Clim. Res. 2011, 47, 123–138. [Google Scholar] [CrossRef]
- Yang, T.; Wang, X.Y.; Zhao, C.Y.; Chen, X.; Yu, Z.B.; Shao, Q.X.; Xu, C.Y.; Xia, J.; Wang, W.G. Changes of climate extremes in a typical arid zone: Observations and multimodel ensemble projections. J. Geophys. Res.-Atmos. 2011, 116, D19106. [Google Scholar] [CrossRef]
- Bogan, M.T.; Lytle, D.A. Severe drought drives novel community trajectories in desert stream pools. Freshw. Biol. 2011, 56, 2070–2081. [Google Scholar] [CrossRef]
- Foti, R.; del Jesus, M.; Rinaldo, A.; Rodriguez-Iturbe, I. Hydroperiod regime controls the organization of plant species in wetlands. Proc. Natl. Acad. Sci. USA 2012, 109, 19596–19600. [Google Scholar] [CrossRef] [PubMed]
- Ireland, A.W.; Booth, R.K. Hydroclimatic variability drives episodic expansion of a floating peat mat in a North American kettlehole basin. Ecology 2011, 92, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Loverde-Oliveira, S.M.; Huszar, V.L.M.; Mazzeo, N.; Scheffer, M. Hydrology-driven regime shifts in a shallow tropical lake. Ecosystems 2009, 12, 807–819. [Google Scholar] [CrossRef]
- Biggs, R.; Carpenter, S.R.; Brock, W.A. Turning back from the brink: Detecting an impending regime shift in time to avert it. Proc. Natl. Acad. Sci. USA 2009, 106, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Palmer, M.A.; Reidy Liermann, C.A.; Nilsson, C.; Flörke, M.; Alcamo, J.; Lake, P.S.; Bond, N. Climate change and the world’s river basins: Anticipating management options. Front. Ecol. Environ. 2008, 6, 81–89. [Google Scholar] [CrossRef]
- Allen, C.R.; Angeler, D.G.; Garmestani, A.S.; Gunderson, L.H.; Holling, C.S. Panarchy: Theory and application. Ecosystems 2014, 17, 578–589. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baho, D.L.; Tavşanoğlu, Ü.N.; Šorf, M.; Stefanidis, K.; Drakare, S.; Scharfenberger, U.; Agasild, H.; Beklioğlu, M.; Hejzlar, J.; Adrian, R.; et al. Macroecological Patterns of Resilience Inferred from a Multinational, Synchronized Experiment. Sustainability 2015, 7, 1142-1160. https://doi.org/10.3390/su7021142
Baho DL, Tavşanoğlu ÜN, Šorf M, Stefanidis K, Drakare S, Scharfenberger U, Agasild H, Beklioğlu M, Hejzlar J, Adrian R, et al. Macroecological Patterns of Resilience Inferred from a Multinational, Synchronized Experiment. Sustainability. 2015; 7(2):1142-1160. https://doi.org/10.3390/su7021142
Chicago/Turabian StyleBaho, Didier L., Ülkü Nihan Tavşanoğlu, Michal Šorf, Kostantinos Stefanidis, Stina Drakare, Ulrike Scharfenberger, Helen Agasild, Meryem Beklioğlu, Josef Hejzlar, Rita Adrian, and et al. 2015. "Macroecological Patterns of Resilience Inferred from a Multinational, Synchronized Experiment" Sustainability 7, no. 2: 1142-1160. https://doi.org/10.3390/su7021142
APA StyleBaho, D. L., Tavşanoğlu, Ü. N., Šorf, M., Stefanidis, K., Drakare, S., Scharfenberger, U., Agasild, H., Beklioğlu, M., Hejzlar, J., Adrian, R., Papastergiadou, E., Zingel, P., Søndergaard, M., Jeppesen, E., & Angeler, D. G. (2015). Macroecological Patterns of Resilience Inferred from a Multinational, Synchronized Experiment. Sustainability, 7(2), 1142-1160. https://doi.org/10.3390/su7021142