1. Organic Potatoes Production Requires Breeding
Potato (
Solanum tuberosum) is the third most important food crop worldwide after rice and wheat, with a total production of over 300 million metric tons, as stated by the International Potato Center (CIP), Lima, Perù [
1]. Average potato yield has been erratic across the world during the past decade, ranging between 16.3 to 19.4 t/ha from 2000 to 2011, though showing an overall slight increase (FAOSTAT, 2011 [
2]). China and India are the largest potato producers: 88.35 and 42.33 MT, respectively.
Over the last decade, changing food lifestyles reflected the consumers’ growing interest in organic food, generally perceived as healthier and safer for humans and environment. Organic food is in fact produced from raw material obtained by agricultural farming systems (organic farming) relying on ecological processes, biodiversity, and cycles adapted to local conditions—rather than the use of inputs with adverse effects—sustaining the health of soils, ecosystems, and people. Its diffusion has largely increased across the entire world, assuming a relevant economic impact, with social and political implications, implemented also by the European agricultural policies (2009/128/CE Directive, [
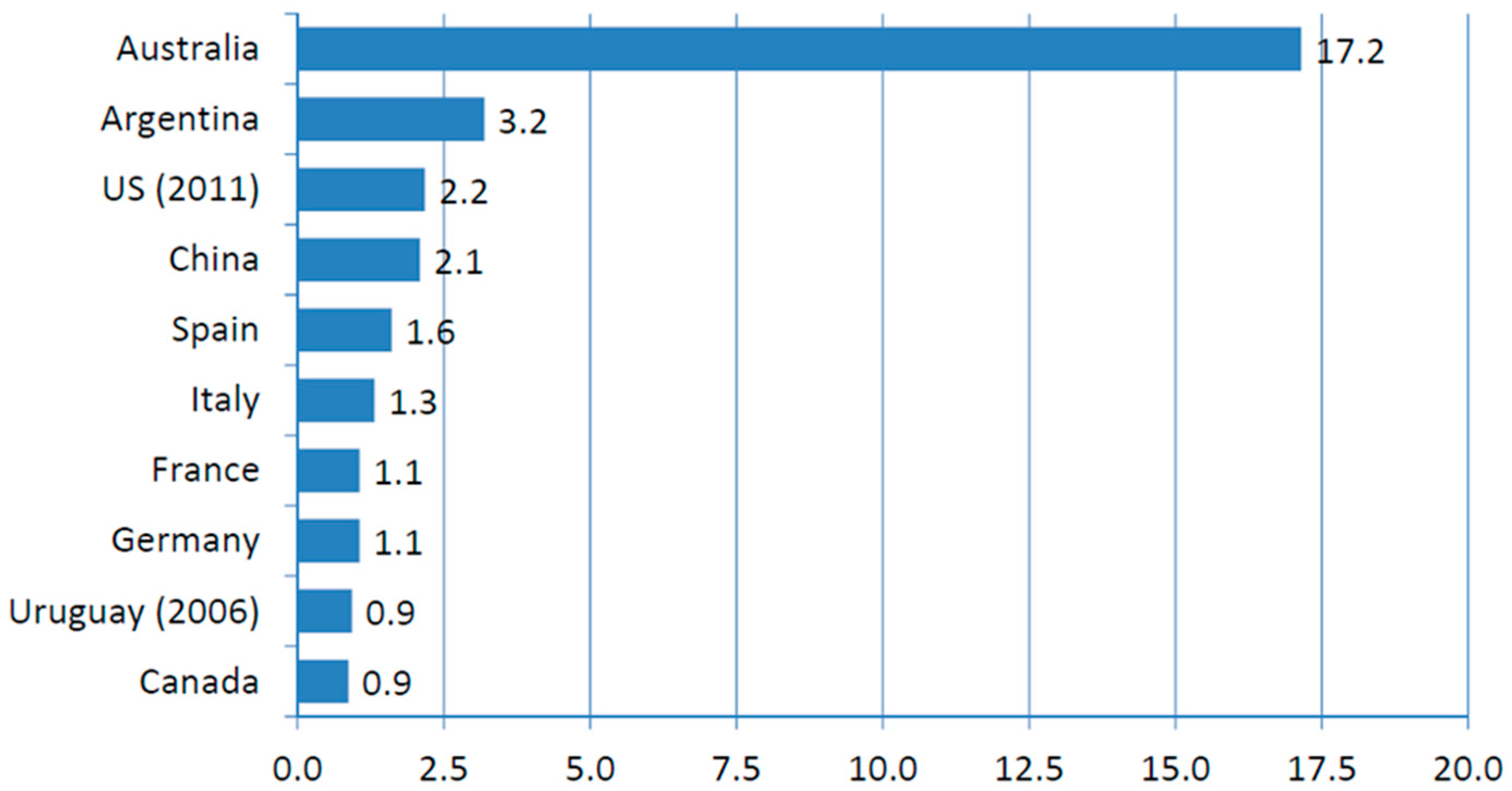
3]). In the world, Australia stands as the largest area dedicated to organic farming while Italy ranks at sixth place, with a production of 1.3 MH (data end of 2011 [
4],
Figure 1).
The 8.7% of total Italian agricultural land is dedicated to organic farming (Fibl-IFOAM Survey 2012), and the sales of organic food products increased of 20% in 2015 (Coldiretti [
5], the major Italian farmers union). The main features of organic farming are claimed to be sustainability, low-impact cropping methods, use of non-chemical fertilizers, and high-quality production; nevertheless, an objective and exhaustive assessment on the supposed higher nutritional value of organic food and on the benefits on consumers’ health is still missing. Generally, organic food presents lower nitrates and higher content in vitamin C, but this is not true for all species; spinach, for example, accumulates more nitrates under organic farming than other farming systems [
6,
7,
8]. A literature-based study including potato [
7] revealed that organic vegetables are not significantly better than conventional ones and, in addition, experimental findings are often contradictory due to the influence of environmental factors and large genetic diversity among potato varieties tested. Up to date, only few studies focused on organic potatoes and most of them investigated target beneficial (vitamin C, flavonoids, vitamin B1, phenolics) and harmful compounds (heavy metals, glycoalkaloids, nitrates, residues of pesticides), organoleptic properties, and agronomic parameters (yield, tuber size, germination of tubers, starch content). Surprisingly, a high vitamin C content, typical of most organic food, is not always confirmed in organic tubers [
7,
9,
10,
11,
12] that, indeed, frequently show high content of vitamin B1 [
11]. Organic restrictions on fertilization causes a reduced nitrogen availability, resulting in a lower N content in organic tubers, that could strongly influence the vitamin C content, together with the concentration of nitrate and total proteins [
6,
7] and the content and composition in free amino acids. Lysine, the most abundant free amino acid in potato, was often found significantly higher in organic tubers [
10,
13]. Protein profiling also changes, with organic tuber proteome enriched in stress-related proteins [
14], even though other authors, such as Brazinskene et al. [
15], are not in agreement with these findings. As reported by Brandt et al. [
16], the plants managed by organic practices have necessarily to cope more frequently with pathogens, and accordingly accumulate more defense-related compounds (such as phenylpropanoids [
9,
10,
12,
17]) than conventionally produced tubers. Some phenolics are associated with health-promoting effects, mainly related to their antioxidant activity: chlorogenic acid, carotenoids, and anthocyanins have been reported to protect against degenerative diseases, cancer, heart diseases, hypertension, and viral and bacterial diseases [
18]. Instead, for other defense compounds such as glycoalkaloids (GAs) results are few and not in agreement [
7,
11]. GAs are known antinutritional compounds, unsuitable for human consumption at high concentrations (>20 mg 100 g
−1 of fresh weight), due to their toxicity. Elevated content of GAs were found mainly in the skin of immature tubers or in tubers exposed to light or stressed by different factors, like pathogens or drought [
19]. Further investigation is required to identify if there is a systematic trend for GA accumulation after organic cultivation.
Organic growers are severely limited from using synthetic chemicals (International Federation of Organic Agriculture Movements’ Norms, 2014 [
20]), so detectable residue levels of synthetic pesticides, as chloropropham, are systematically and significantly lower in organic products, with the only exception of the fungicide chlorothalonil [
7]. In order to control the late blight disease caused by oomycete
Phytophthora infestans, especially in areas where potatoes are grown on large extensions and rain occurs during the production period, copper (Cu) fungicides are exceptionally allowed since organic potatoes are very exposed to this pathogen [
21]. Cu is a persistent and toxic heavy metal, and should be minimized in the soil to reduce pollution; therefore, in 2006, after copper accumulation in the soils reached dangerous levels, EU limited its application from 8 to 6 kg/ha per year (further lowered to 3–4 kg/ha in Germany and Switzerland, under organic regulations [
22]). In countries where Cu is prohibited, 100% yield losses often occur (Finland, 2004).
While targeted approaches permitted to address specific questions as those reported above, “omics” and integrated multi-omics approaches currently provide the chance to also identify unexpected nutrients or anti-nutritional compounds that differentially occur in different farming systems, highlighting their influences on the overall physiological and biochemical status of tubers. For example, a metabolome study revealed the presence of formic acid in organic tubers, probably present in bee glue usually used in organic practices [
10]. The same authors reported a great overall carbohydrates reduction in organic tubers, and suggested a putative key role of the non-protein amino acid gamma-aminobutyric acid (GABA), strongly accumulated in organic potatoes, in carbon and nitrogen portioning. In the same way, a controlled paired study on differences between organic and conventional tubers proteomes suggested that the protein pattern is much more influenced by organic fertilization regime than organic crop protection treatments, identifying the first as the major influence on the physiological status of the tuber [
14].
Even though it is still hard to draw conclusive statements about the health effects of organic food, it is becoming increasingly clearer that not only farming practices, but also the choice of genotype, strongly affects chemical composition of potato tubers. It is necessary to develop varieties suitable for a low-input agriculture, making “organic plant breeding” a challenging task to maximize the positive effects of organic farming. The biggest limit is the so-called yield gap: many studies report that the average yield in organic farming is 60% lower than yields obtained using conventional practice, with losses mainly due to biotic stresses including late blight and early blight (causal agent
Alternaria solani; [
16]). The feasibility of breeding to enable a sustainable intensification of organic farming has been recently reviewed [
23,
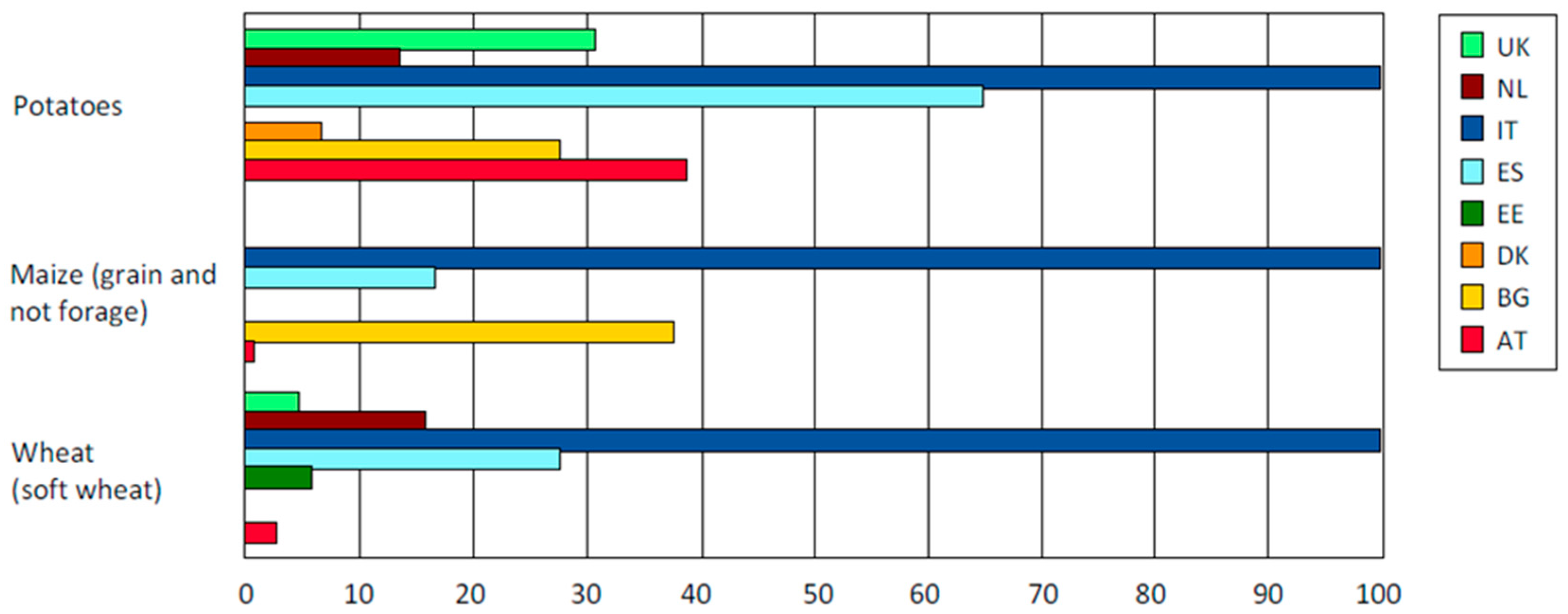
24] suggesting the necessity to introduce in crop varieties useful traits available in wild germplasm. Zero synthetic pesticides implies putting emphasis on the level of genetic resistance and the need of low level fertilizers implies to increase the nitrogen use efficiency (NUE) of varieties intended for organic farming. Weed competitiveness and the ability to establish relationships with microorganisms in the soil are also relevant because these factors can enhance the uptake of resources and its use efficiency. In addition, sensorial and nutritional characteristics of organic potato are strictly hampered from the use of varieties not selected for this purpose. Quality improvements of organic varieties regard the reduced accumulation in tubers of glycoalkaloids and allergens (such as patatin) and the reduction of the potato glycemic index. Over the time, other important quality traits are expected to be pursued, such as improved nutritional value, with direct benefits to consumers. In our opinion, an increased availability of organic varieties will also discourage farmers from using non-organic seeds in organic farming. Nowadays this practice is admitted upon adequate justification, i.e., if the variety is not available, but in Italy it is a very common practice, not only for potatoes but also for wheat and maize, making this country the worst supplier for organic seeds for these species (
Figure 2).
From a legal point of view, Regulation (EC) 889/2008 [
26] does not specify which breeding techniques are allowed for organic plant breeding. The only clear exclusion is gene manipulation, even though CMS-Hybrids (cytoplasmic male sterility), that use cell fusion to combine species that normally cannot crossbreed, are allowed [
25]. De facto, organic potato breeding is restricted to traditional breeding based on crossing of heterozygous tetraploid parents with different phenotypic assessments and selection of a small percentage of the offspring that contains at least some of the desirable traits of the parents. This process takes a long time to produce a new marketable potato variety from the initial cross (up to 15 years) because it requires the elimination of unwanted traits. It follows that cultivated potatoes have a narrow genetic base, resulting in high susceptibility to many pests and pathogens, with low resilience and adaptability to different environments and to different agricultural practices, and a reduced varietal choice. The feasibility of re-introducing in elite crop varieties traits available in the gene pool of Solanum could pass through the so-called new plant breeding techniques (NPTBs) [
23,
24].
2. Not All GM Potatoes Are Equal
Conventional breeding techniques and genetic engineering approaches are essential components of what we can call plant breeders’ toolbox, sharing the goal to give plants new properties (e.g., a resistance to a disease, insect or drought, tolerance to a herbicide, an improvement of food quality or nutritional value, an increased yield). All the genetic engineering processes result in genetically modified (GM) organisms or plants, but technologies have evolved to the more recent NPBTs and products obtained may have quite different characteristics. However, an increasing confusion in the public debates on GMOs is still contributing to creating misunderstanding between processes and products that we would like to clarify.
In its old sense, the term genetic engineering or modification indicates the incorporation into a plant genome of genetic elements (coding and regulatory sequences) derived from any organism (eukaryotic, prokaryotic) as well as novel sequences synthetized in vitro that are able to confer the desired phenotype. If the genetic elements incorporated derive from individuals that cannot naturally breed, the approach is called transgenesis and produces transgenic plants. If a gene (with its promoter, introns, and terminator) is taken as such from a crossable individual, without introducing any foreign material, the approach is called cisgenesis and develops cisgenic plants [
27]. Based on these definitions, even if the gene of interest derives from a crossable species, the use of selectable marker genes, such as bacterial antibiotic resistance genes, necessarily results in a transgenic plant. Only if marker genes are removed by site-specific recombinases or by a marker-free transformation, a cisgenic plant is produced. As a third case, if we use sequences (genes and regulatory sequences) taken from the same or closely related individuals but in combinations that are different from those available in nature, we develop an intragenic plant by intragenesis [
28]. Gene silencing approaches, e.g., RNA interference (RNAi), are included in intragenesis offering considerably more options than cisgenesis (even if still less compared to transgenesis). Transgenes, cisgenes, and intragenes can be delivered in the plant cells using the same method, in most cases by Agrobacterium, and therefore the process is the same, but the product can be quite different. Only a cisgenic plant can, in principle, be produced also through traditional breeding.
Among NPBTs, genome editing by site-directed nucleases (SDN) has great potential in plant breeding. In general, SDNs introduce a double-strand break in a target DNA sequence in cells, that is repaired by the cell’s repair system itself and can result in different outcomes: gene knockout of site-specific random mutations are produced by non-homologous end joining (SDN-1), gene correction/replacement if a donor DNA is used for the repair by homologous recombination (SDN-2); and, finally, gene addition (SDN-3) to introduce long fragments (e.g., transgenes, cisgenes) [
29]). Delivery of the component necessary to break the DNA can be accomplished by transient or stable transformation methods; the first are preferred to avoid the presence of exogenous DNA sequences. If sequences encoding nucleases and other components are not present in the final edited products, plants obtained by targeted mutagenesis are indistinguishable from plants obtained by conventional mutagenesis or natural variations.
NPBTs are all promising tools for the further improvement of elite crop varieties, especially in crops that are vegetatively propagated to maintain their genetic composition—such as apple, grape, potato, cassava, and banana—for which the long time and high cost required for traditional breeding would be drastically reduced. Potato is an intensively managed crop, which requires irrigation, fertilization, and frequent pesticide applications in order to assure the highest possible yield and requires a great genetic improvement effort. Despite this and its worldwide importance, the predominant variety used in the United States (Russet Burbank) was developed more than 130 years ago and suffers from multiple biotic and abiotic diseases. Most commercial potato varieties are autotetraploids, highly heterozygous, and suffer acute inbreeding depression; these attributes seriously hampered potato improvement by traditional breeding approaches. Fortunately, potato is amenable to in vitro culture and it is relatively easy to introduce new traits to through genetic transformation of diverse genotypes. This makes potato one of the first successful (scientifically speaking) GM crops, and it still remains as the forefront of plant biotechnology (
Table 1). Transgenic potatoes were among the first GM plants produced. The story began at the end of 1980s in Monsanto laboratories, St. Louis, MO, USA. At that time, Russet Burbank was the most economically important cultivated potato variety, but it was highly vulnerable to production losses due to viruses (especially potato leafroll virus, PLRV) and insects (mainly the Colorado potato beetle or
Leptinotarsa decemlineata). To face these problems, two teams of Monsanto scientists, the Virus Team and the Insect control Group, began working extensively on resistance to viruses and insects respectively, by using transgenic expression of viral regions (taken from coat protein or replicase genes) and/or of a synthetic Bacillus thuringensis Bt gene [
30]. Between 1995 and 2000, the first transgenic potato lines carrying single or combined resistance traits in the same genotype were developed, registered under the brand name NewLeaf™ and marketed by NatureMark, a wholly-owned subsidiary of Monsanto. GM potatoes received great initial acceptance in the United States because they were assessed as safe by USDA and produced healthy potatoes, free of necrosis and with a reduced or zero requirement for application of insecticides, with great benefits for farmers, processors, consumers, and the environment. Farmers saved money especially on insecticides, no more needing to control both CPB and insect vectors of PLRV. Processors benefitted of high-quality raw material, free of internal discoloring caused by PLRV, and consumers received superior quality potatoes at no additional cost. The environment also benefitted for the significant reduction in pesticides applications [
31]. These GM potatoes were also approved for export to Japan, Mexico, Australia, Bulgaria, Romania, and Russia, at least until a successful anti-GM campaign against transgenic potatoes in 1999 began. At that time, quick service restaurants decided to ban GM foods from their food chain for image reasons, and Europe did not approve import of transgenic potatoes. Even if fresh market was favorable to biotech potatoes, the closure of processed potatoes caused GM potato production to come to a halt, and public and private research on potato improvement by advanced technologies virtually ceased. Acceptance of transgenic potatoes and development of new lines to address local pests (like potato tuber moth) were however encouraged in many developing countries, where costs precluded large-scale applications of pesticides, and that therefore greatly benefited from the GM resistance crops.
Meanwhile, in Europe the BASF company developed the transgenic variety Amflora (also known as EH92-527-1), a high amylopectin variety suitable for industrial applications, obtained by silencing the granule bound starch synthase (GBSS) gene controlling the amylose synthesis [
40]. Amflora contained the antibiotic resistance gene
nptII as well, of bacterial origin. Nevertheless, it was evaluated as safe for cultivation because the presence of
nptII gene in GM plants did not lead to higher chance of (medically relevant) bacteria developing resistance against antibiotics [
41]. After 13 years, Amflora received European approval for commercial cultivation in EU in 2010. The following year, BASF proposed the late blight resistant variety Fortuna for EU authorization for commercialization [
42]. Even if Fortuna carried cisgenes (two resistance genes from a wild Mexican potato relative), it could not be defined as cisgenic because it contained the herbicide resistance gene
ahas, from a non-crossable donor plant. The reaction of GM opponents was so strong to demand the EU to authorize each European country to ban GM crops for cultivation. In January 2012, Amflora and Fortuna were withdrawn by BASF from the EU market due to the uncertainty in the regulations and threats of field destructions.
Learning from market difficulties encountered by Monsanto and BASF, J.R. Simplot company focused on ‘all native’ DNA transformation or intragenesis, using only potato genes for trait introduction in order to address public concerns. The first goal was the elimination of undesired native traits from French fries and potato chips, such as the production of acrylamide and black spot bruising through gene silencing. The substrates for the production of acrylamide are reducing sugars (glucose and fructose), and the amino acid asparagine. Therefore, strategies to reduce acrylamide forming potential were the silencing of the acid invertase gene [
43] and asparagine synthase-1 (StAst1) gene [
35], through intragenic modifications [
36,
37,
44]. Black spot bruise, successfully prevented by silencing polyphenol oxidase (PPO) genes [
45], was also obtained by intragenesis, without inserting any foreign DNA in the plant genome. The first generation of Innate™ Simplot potato combined low acrylamide potential (low asparagine) and black spot bruise resistance. In 2014, the US Department of Agriculture (USDA) and the US Food and Drug Administration (FDA) approved its cultivation in the USA, and in 2015 it also received clearance from Canadian government; therefore, these potatoes could be sold on US and Canadian market since the beginning of 2016. The Canadian Food Inspection Agency determined that these potatoes are “safe and nutritious as traditional potato varieties”, and that they do not present an increased risk to the environment when compared to currently cultivated potato varieties in Canada. A second generation of Innate™ potatoes was further developed by Simplot, and has been recently deregulated by the USDA [
46]. This clone contains the same traits as the first generation (low black spot bruise and low asparagine), plus two additional traits: resistance to late blight and low reducing sugars derived from wild or cultivated potatoes. These improved clones provide significant sustainability benefits for potato growers, processors, and consumers. For example, if all fresh Russet potatoes were converted, the U.S. would have the potential to save 400 million lbs. of potato waste each year. Levels of acrylamide could be reduced up to 90% percent when these potatoes are cooked at high temperatures, which cannot be achieved until today. Besides, low reducing sugars enable cold storage at 4 °C for more than six months without the build-up of sugars, reducing sprout suppressants application and improving potato quality.
Big efforts to obtain durable resistance in potato against late blight disease were done through cisgenesis (DuRPh project, [
47]), by combining multiple resistance genes. The sustainable control of this pathogen can be achieved only through genetic modification. Different late blight resistance genes were cloned from wild relatives (
Solanum stoloniferum,
S. bulbocastanum,
S. demissum, and
S. venturii) and introduced in several potato cisgenic lines [
32,
33,
47,
48] using a marker-free transformation approach [
32,
34]. Stacking of multiple R genes maximized durability of resistance and a correct spatial and temporal management where different improved lines may subsequently be introduced in a cultivation rotation scheme, supporting disease management and reducing fungicide use by over 80%. The cost to get durable late blight resistance in potato through dynamic varieties was estimated as being lower than the cost of traditional late blight management and of traditional breeding [
47].
New technologies for mutagenesis by SDNs were also successfully applied in potato, such as Transcription Activator-Like Effector Nuclease TALEN [
49] and the Clustered Regular Interspaced Short Palindromic Repeats (CRISPR)-associated (Cas) system [
50] that are quite simple and efficient. Sawai applied TALEN through a transgenic approach with stable transformation to disrupt the sterol side chain reductase 2 (
St SSR2) gene, involved in glycoalkaloid synthesis, obtaining potatoes with a significant reduction in these toxic compounds [
39]. Nicolia proposed a pipeline for tetraploid potato genome editing based on transient expression of TALEN in protoplasts, potentially usable also for other SDNs (e.g.,
Cas), assuring no T-DNA insertion in the plant genome [
51]. Meanwhile, a similar approach was applied at the biotech Company Calyxt, to improve cold storage and processing traits of the variety Ranger Russet, through the knockout of potato vacuolar invertase gene (
St Vlnv), involved in the production of reducing sugars in cold-stored tubers, resulting in a significant reduction of acrylamide levels in potato chips [
38]. The complete knockout lines were free of foreign DNA and therefore indistinguishable from the variety except for small deletions in the targeted gene. In other cases, targeted mutations by CRISPR/Cas9 were only induced in reporter genes and/or studied in vitro at molecular level [
52,
53].
3. Cisgenic Potatoes in Organic Farming: Scientific, Legislative, Ethical, and Social Aspects
Organic farming with GM plants seems to be a pact with the devil [
54]. According to Lammerts Van Bueren et al. [
55], gene manipulation is legally and morally not acceptable in organic agriculture. Since 1993, the Basic Standards for Organic Production and Processing of IFOAM includes a strict ban on GMOs (Position Paper on genetic engineering in 2002 [
56], revised in a draft of 2016 [
57]). Also the EU Regulation for organic agriculture 2092/91/EC [
58] prohibits the use of GMOs and the products obtained through their use. This position is mainly based on three statements. First, the possibility of unintended effects caused, for example, by transgene integration in random sites (e.g., interruption of important open reading frames or regulatory sequences), which could result in modified metabolism, novel fusion proteins, or other pleiotropic effects that could compromise safety, including the production of new allergens or toxins [
59,
60]; second point, GMOs are not compatible with the ecological principle of sustainability and, third point, the intellectual property rights and economic independence of farmers are threatened by large-scale use of GMOs. The first two statements are mainly based on the assumption that GM plants and foods are different from non-modified ones, and therefore potentially risky. But is it true?
“Substantial equivalence”, a concept introduced in 1993 by the Organization for Economic Development (OECD, [
61]), is used as starting point to structure current food safety assessment and suggests comparison of intended differences between the GM plant and progenitor cultivar. If a GM food is found to be substantially equivalent to an already existing food product, with a history of safe use, it can be treated in the same way [
62]. According to the World Food Safety Organization, a case-by-case consideration should be also adopted on the nature of the genetic modification, the plant modified, and the environment in which it will be introduced. Based on scientific evidence, the first generation of transgenic potato tubers did not differ significantly from conventional non-modified crop varieties, except for the inserted trait. There was much less variation in the proteome of GM tubers and their non-GM control, compared with that found between different varieties and landraces. Therefore, in respect to the natural variation observed in the non-GM samples, the unintended effects of transformation were considerably less pronounced [
63,
64,
65]. In addition, no new proteins unique to individual GM lines were observed. Despite this, organic supporters do not care if a GM plant or product is substantially equivalent or not to originals, focusing instead on the processes used for plant breeding. This implies an ethical choice: the intrinsic argument of “unnaturalness” of genetic engineering applications automatically does not support a scientific risk-benefit analysis and a final product-based assessment. In this view, it is hard to think that cisgenic potatoes will defuse GM opponents. However, technologies for genetic engineering are so diverse and products obtained from them can have so many useful traits that generalization is no longer acceptable.
The transgenic variety Fortuna was considered safe for cultivation after years of evaluation. However, the presence of a selectable marker gene of bacterial origin makes it not substantially equivalent to the original one. To avoid this issue, bacterial marker genes and marker genes from non-crossable plant species should be eliminated by recombination or avoided in favor of selectable cisgenes, as already reported for the apple Myb10 gene [
66,
67]. Potato is advantaged with respect to other species because cisgenesis can be obtained without the use of any selectable marker gene, due the high transformation and regeneration rates that are necessary for this strategy. This is the case of the more recent potato genotypes carrying late blight resistance genes [
32]. Also, the intragenic potatoes produced by Simplot are not indistinguishable from native ones, as they carry a combination of sequences not available in nature, but they are safe and not risky both for humans and for the environment. Again, consumers do not seem really aware of that, generally perceiving any type of organic food as healthier and safer than any type of GM food. Surprisingly, organic protocol accepts varieties obtained by mutagenesis derived from totally random modification events in the starting genome, even when the precise number and type of mutations are not known. Induced mutagenesis have been used for a while to obtain new varieties improved for several agronomic and food quality traits. From 1930 to 2014 more than 3200 genotypes have been obtained by mutation breeding and released. Paradoxically, the deployment of new biotechnology-based plant breeding techniques offers great opportunities for highly precise plant breeding, enabling predictable alteration of the genetic make-up of the plant, but their products are not accepted as they are the result of a genetic modification. With site-directed nucleases, the modification is even more precise, because the insertion of DNA can be targeted to a predefined genomic region. Therefore, if mutagenic varieties are compatible with the principles of organic farming, cisgenic and genome edited crops—legally excluded from organic agriculture—actually sound like the most reasonable solutions for organic production.
Another issue to point out is whether genetic engineering is actually far not only from classical breeding approaches, but even from those natural mechanisms that drive evolution. In fact, genetic engineering has been brought into an evolutionary perspective of natural mutations by authorities such as the Nobel Prize winner Werner Arber; according to his view, transgenesis and naturally occurring molecular evolution (that is the spontaneous generation of genetic variants) follow exactly the same strategies [
68]. “Therefore, conjectural risks of genetic engineering must be of the same order as those for natural biological evolution and for conventional breeding methods. These risks are known to be quite low. There is no scientific reason to assume special long-term risks for GM crops”. This conclusion is further supported by the quite recent discovery that Agrobacterium T-DNAs carrying expressed genes are naturally integrated in the genome of cultivated sweet potato, one of the world’s most consumed foods, resulting in an example of a natural transgenic crop. This finding could influence the public’s current perception that transgenic crops are “unnatural” [
69].
In contrast to the common belief that the cultivation of GM plants is not sustainable, cisgenic potatoes resistant to late blight can certainly assure less treatment requirements than most of the commercial varieties that are susceptible. Of all potato farmers in EU, 99% spray up to 20 fungicides per year against
P. infestans, with a cost estimated at about €5.2 billion worldwide for both control and yield losses [
48]. Besides economic costs, frequent chemical applications are risky for farmers’ health and have undesirable environmental side effects [
70].
Another arguable issue in the debate between opponents and proponents of genetic engineering is the plant genetic resources management. Organic farming promotes germplasm conservation in order to preserve and use the genetic diversity as an integral part of the organic farming activity. The loss of genetic diversity, or genetic erosion, is a serious threat to biodiversity, but it was already present before the appearance of the first GM plants, and this is mainly due to the choice of most farmers worldwide to leave their multiple local varieties and landraces for genetically uniform, high-yielding (not GM) varieties. As genetic diversity is the basis for breeding, recovery of genetic resources has become a priority and new breeding techniques represent a great possibility to re-introduce traits that have been lost or have never been present in the cultivated taxa, a process called “re-wilding”. The collaboration between local farming communities in the centers of origin of cultivated potato in the Andes of South America, precious holders of traditional knowledge, and potato breeders could allow retrieval of a plethora of interesting traits from landraces, heirloom varieties, and wild crop relatives, that show interesting features in terms of resilience and nutritional value. The potato heritage is actually mainly in the hands of the International Potato Center (CIP, member of CGIAR consortium, [
71]), engaged in discovering and preserving by in situ conservation ecotypes, native and old potato varieties, carrying biotic and abiotic stress-defense traits and/or enriched in functional molecules precious for nutritional and health properties. This is the case of traits such as high anthocyanins content, typical of wild red to purple flesh colored potatoes or in zeaxanthin, responsible for the orange flesh of some Andean potato clones. These traits are not present in tetraploid cultivated potatoes that have a white to yellow flesh, according to the amount and composition in carotenoids (
Figure 3).
Through cisgenesis and genome editing, it is now possible to re-introduce these traits in cultivated species. The improvement of sequencing technologies has led to the knowledge of the complete genome sequence of many agricultural species, including potato species [
72,
73]. By whole or partial re-sequencing of genetic resources, NPBTs can support the identification of major genes and the mining of superior alleles. Once this information is gained, potato genetic resources can be used as source of information to correct relevant genes in elite crops carrying some defects, or as sources of genes to transfer to elite crops, replacing alleles or introducing new genes in the cultivated gene pool [
74].
GM crops are regulated differently in different countries worldwide. GMOs regulation in the US is relatively favorable to GMO development and this makes the US the world’s leading producers of genetically modified crops. In 2012, of the 170.3 million hectares of biotech crops globally, the United States accounted for 69.5 million, over 40% of the total. The United States does not have any federal legislation that is specific to GMOs. Rather, GMOs are considered pursuant to health, safety, and environmental legislation governing conventional products. The US approach to GMO regulation is focused on the nature of the products, rather than the process in which they were produced [
75]. At the opposite side, the European Union has established a legal framework to ensure that the development of modern biotechnology, and more specifically of GMOs, takes place in safe conditions. The legal framework aims to protect human and animal health and the environment by introducing a safety assessment of the highest possible standards at the EU level before any GMO is placed on the market; to put in place harmonized procedures for risk assessment and authorization of GMOs that are efficient, time-limited, and transparent; to ensure clear labeling of GMOs placed on the market in order to enable consumers as well as professionals (e.g., farmers, and food feed chain operators) to make an informed choice; and to ensure the traceability of GMOs placed on the market. The main building blocks of EU GMO legislation are Directive 2001/18/EC [
76] on the deliberate release of GMOs into the environment and Regulation (EC) 1829/2003 [
77] on genetically modified food and feed. In order to take into account scientific and technical development in plant biotechnology, in 2007 the European commission established a New Techniques Working Group to evaluate a number of NPBTs in respect to the current European legislation. The European Food Safety Authority (EFSA) and the Joint Research Center (JRC) were asked to issue a science-based opinion on the potential of these technologies and the classification of their products. Products of NPBTs are still regarded and regulated as GMOs. However, already 10 years ago Schouten and colleagues [
27,
78] proposed to exclude cisgenesis from EU regulation of GM products because “cisgenic plants are fundamentally different from transgenic plants”, and they should be “handled at the regulatory level like traditionally bred plants”. Moreover, “similar hazards can be associated with cisgenesis and conventional bred plants, while novel hazards can be associated with intragenesis and transgenesis” (EFSA Opinion 2012, [
79]). Whether identified hazards translate into risks to human and animal health and the environment depend from factors such as the extent to which the plant is cultivated or its product consumed. According to the GMO panel of EFSA, the current guidance for risk assessment of transgenic plants can be applied also to those developed through cisgenesis, intragenesis, and site-directed nuclease-3, but with more flexibility and a lower amount of data required, determined on a case-by-case basis.
GM potatoes must be accepted also by consumers to become a market success. Although no scientific evidence has been related so far to a safer use of cisgenesis instead of transgenesis, a more positive consumers’ perception has been reported towards the use of genes from a compatible gene pool [
80,
81,
82]. Clearly, it adds a new dimension to the debate on GMO [
83]. However, consumers’ resistance to GM products is still perceivable, especially in the EU. The story of biotech potatoes highlights how much consumers’ opinion can be affected by unfounded concerns that are cyclically proposed by environmental groups and organic farming supporters, stopping public research and halting innovation and economics. However, if correctly informed both on positive and negative aspects of the use of biotechnology, consumers become more condescending in GM food consumption. This underlines the importance to educate consumers on the use of biotechnology to produce healthy food.
The last but not the least important point, is that consumers’ acceptance greatly depends on the classification as GM or not GM of the products of NPBTs. So we close the cycle, returning to government acceptance and the need to streamline the assessment procedures and reduce regulatory costs. If cisgenesis and genome editing will be deregulated in EU, hopefully there will also be major acceptance by consumers.
4. Discussion and Conclusions
The restricted use or even the prohibition of chemical fertilizers, pesticides, and other agents is generally a guarantee for a lower environmental impact of organic farming, but it is also linked to a lower yield and higher land use, unsuitable to sustainable food production. Food supply is a great concern for human kind and takes place in a complex global scenario. There is an increasing demand for food, due to human population growth; but food production is seriously threated by climate change and the constant pressure of pests and pathogens. From one side, sustainable agriculture is what agricultural systems should aim for in order to face future; from the other, it is necessary to guarantee secure and nutritious food for everyone, though the question is how. Is it really necessary to increase production or is it mostly a question of different allocation and more efficient exploitation of the sources we already have? ONU stated that by 2050, farmers must produce 40% more food to feed an estimated nine billion people on the planet, mainly concentrated in the large urban centers, i.e., far from areas dedicated to the food production. The consequence is that more land will be sacrificed to agriculture unless the yield gap would be closed. In agreement with other authors [
84,
85,
86], we consider that organic farming as it is conceived today is not the way to sustainably feed the world, particularly in a scenario where high food productivity is required, with extreme care of the natural resources. An approach based on integrated farming (organic and conventional), the use of low-input agronomic practices and the use of genetically improved varieties requiring less inputs can be essential for enabling food security in a sustainable way. In order to face the major challenges of organic production, smart technologies could be used toward organic genotypes with enhanced productivity and resource-use efficiency, with low impact on the environment. If organic supporters will recognize the fact that, today, organic production is mainly (95%) based on varieties bred for conventional farming and, therefore, are unsuitable for low-input agriculture, with all the negative consequences associated with that, they probably would accept the possibility of improving organic farming with the cultivation of crops bred by NPBTS. Many desirable characteristics and naturally occurring variations already exist within the gene pool of potato species (i.e., in wild relatives) and this further makes NPBTs suitable for this species without the necessity to introduce genes from not-crossable genotypes. Among new breeding tools, cisgenesis and precise genome editing allow the creation of varieties that represent a solution to actual problems of potato cultivation and/or with improved nutritional quality, and that can be totally indistinguishable from the unengineered one. Easy transformation and fast regeneration allow the production of cisgenic potato plants also without the use of selectable marker genes, like those encoding for antibiotic or herbicide resistance. In that line, we argue that modern techniques can fit within the four principles of organic farming (health, ecology, fairness, and care), while some authors already suggested that cisgenesis and reverse breeding based germplasm should be banned from organic farming and organic breeding programs [
87,
88]. However, if we look at these processes as some of the ones that have created variability and drove evolution, there is the possibility that these processes, when applied wisely, will be accepted also in a holistic farming vision.
The effect of organic farming systems on nutraceutical aspects of the tubers has not been studied exhaustively so far, and only a few scientific studies are currently available to provide if organic practices can really result in a surplus nutritional value in order to justify the feeling that “organic is better” or possibly to reconsider the use of these claims for organic plant foods. For sure, potato genetic improvement can instead benefit from straightforward methodologies for precisely targeted genome editing in order to improve the characteristics related to consumer-health. Gene knockouts achieved by TALEN represent the first applications in potato and GM lines with a significant reduction in acrylamide levels in chips, successfully obtained, without the insertion of any foreign DNA. Knockout of endogenous genes can allow the elimination from elite genotypes or from wild relatives of undesired traits such as accumulation of toxic compounds. Application of the Cas9 nuclease system in potatoes is still at the beginning and due to a high degree of specificity and flexibility is intended to become a preferred technology for potato improvement, not only for gene knockout but also for knock in, gene replacement, or gene insertion. Site specific cleavage coupled with the co-addition of a homologous repair sequence can create new alleles via homology-directed repair. This approach can be used, for example, to introduce allelic variants from wild relatives to elite cultivars, avoiding linkage drag. Interestingly, after Cas9 has cleaved a defined target site, a gene (a cisgene, for example) can be incorporated in this locus by homologous recombination (HDR); in this way a gene integration site will also be predictable. Up to now, the unpredictability of integration and expression is still a limitation for public acceptance, regulatory issues, and of course acceptance by organic farmers. Therefore, genome editing technologies could help in reducing these concerns especially if assessments of the products and not of procedures would be adopted, with evaluations conducted case by case, excluding products obtained with new technologies form the regulations for traditional GM plants.
The European Union supports the need for continuous progress in plant breeding techniques, therefore recognizing that they can help to overcome limitations of traditional breeding and enlarge the portfolio of products developed in the EU, but on the other hand imposed severe restrictions to ensure a safety assessment at the highest possible standards. EFSA affirmed that cisgenic plants pose risks similar to those of plants obtained with conventional breeding, but de facto, cisgenesis is still considered as transgenesis in the EU. While United States, Canada, and Australia are open oriented to distinguish cisgenic and intragenic from traditional GMO, Europe and other countries still keep a more cautious approach. While the US Ministry of Agriculture declared that products derived from genome editing technologies will be evaluated case by case, the European Commission has not delivered any opinion yet.
The public’s fears of “Frankenfoods” still persist and even though cisgenic crops increase their share in public opinion, cisgenesis will not have any clout if the public rejects GMO. The terms GM and GMO have been always used with a negative connotation and in our opinion they should be replaced to remove the public negative idea linked to them. We think that classifying NPBTs as GMOs means again to lose competitiveness and technological advancement for breeders and researchers. The consumer is the king of market and if people will trust false myths, there will be no more innovation and competitiveness. The different positions of the parties should converge in compliance with the principles of coexistence and preservation of food and food safety. In the past, great effort was made to evaluate the coexistence and the recommendation was to consider it case by case. In particular, potato, due to the vegetative mode of propagation, always proved suitable for the co-existence of organic, conventional, and GM crops in a cultivation system, because of very low levels of contamination and no significant changes of practices being needed. For cisgenic or genome edited crops, not containing foreign genes, the co-existence with unengineered crops should not to be considered at all as a questionable issue. Evidence-based communication is the only way not to mislead consumers and rebuild trust in science. Agree or not, science must leave aside social, ethical, and political aspects.
In conclusion, the identification of organic varieties will play a key role in improving the management system for organically produced potatoes, still far from an optimization. Research effort and policies should converge to develop a farming system that can benefit from varieties able to express organoleptic, nutritional, and agronomic potential under low-input farming conditions, with adequate pyramiding resistance and nutrient use. Mainly for the species requiring long-term breeding to release new varieties such as potato, this activity could be achieved only through a responsible use of NPBT and correct communication with the public.








{kind=link}
{kind=link}
{kind=link}