
Influence of Different Mowing Systems on Community Characteristics and the Compensatory Growth of Important Species of the Stipa grandis Steppe in Inner Mongolia

Abstract
:1. Introduction
2. Study Area
3. Materials and Methods
3.1. Experimental Setup
3.2. Investigation of Community Characteristics
- Individual biomass = biomassi/numberi
- Importance value index (Pi) = (Relative height + Relative density + Relative biomass)/3
- Shannon–Wiener index (H’) = −∑PilnPi
- Pielou’s evenness index (Jsw) = H’/lnS
- Richness index = (S − 1) logN
3.3. Assessment of Compensatory Growth of Dominant Species in the Community
3.4. Calculation of Compensatory Growth Index of Biomass (G/C)
3.5. Statistical Analyses
4. Results
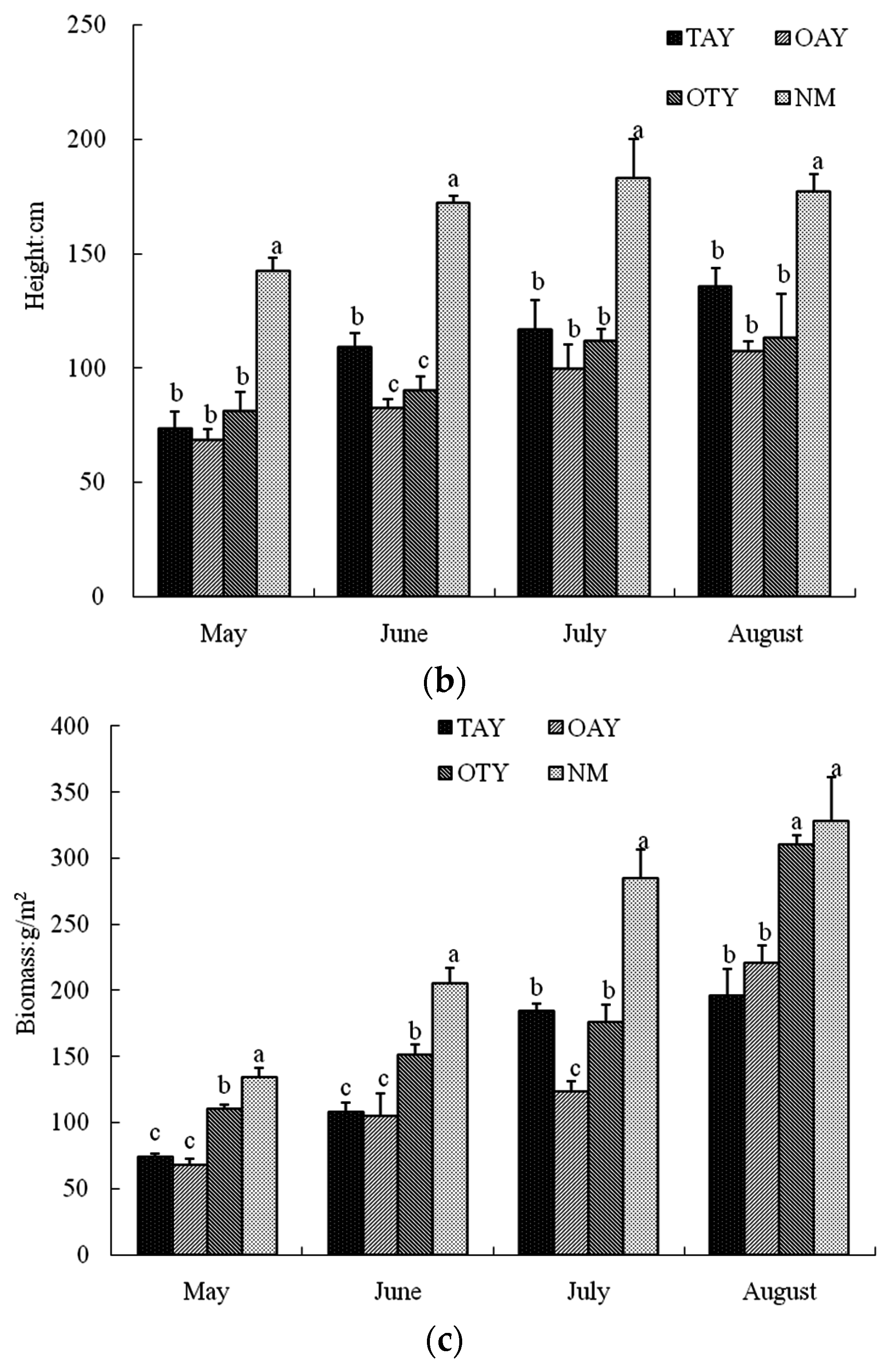
4.1. Community Characteristics
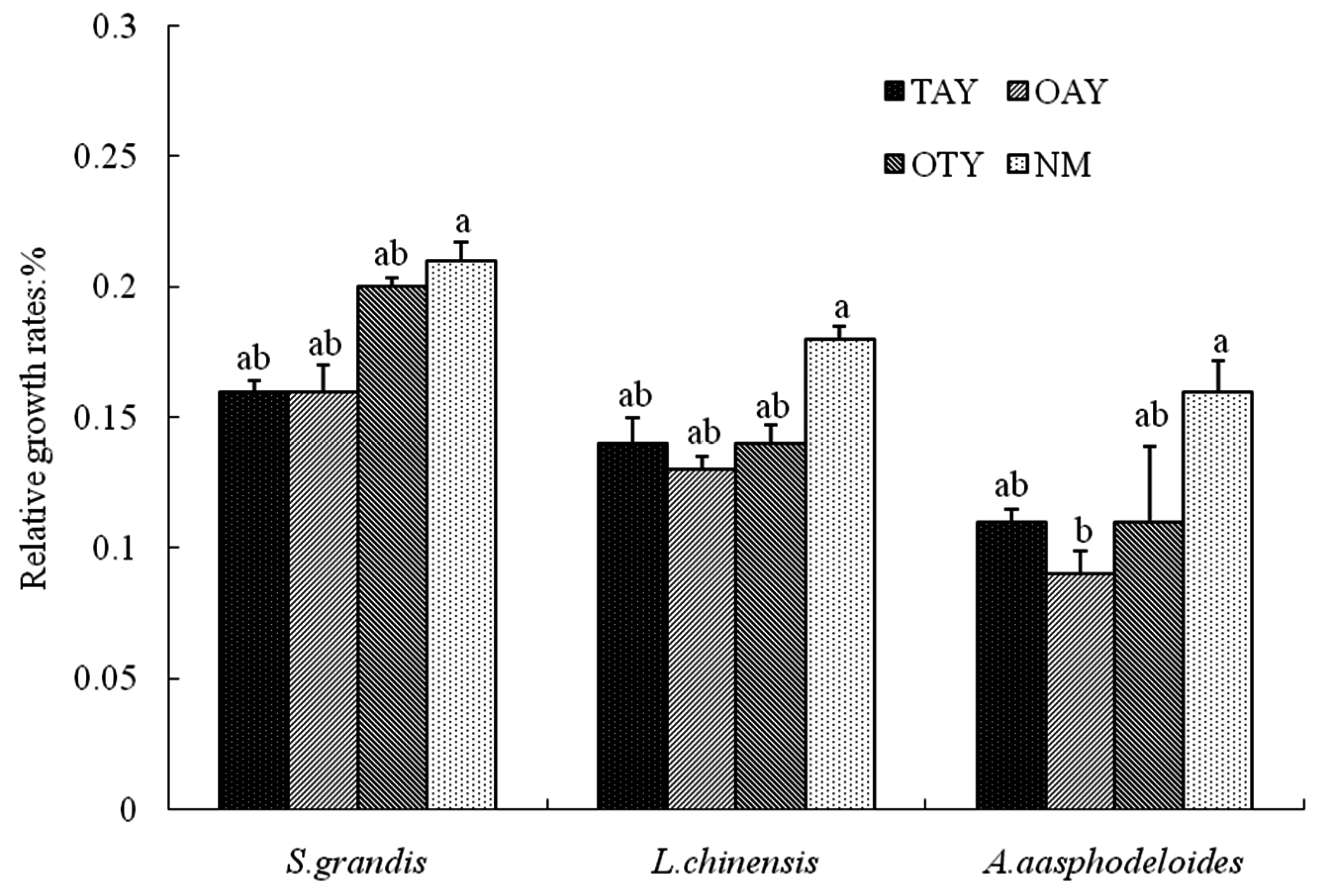
4.2. Compensatory Growth
5. Discussion
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sun, D.Z.; Li, F.S.; Yang, H.S.; An, R.J.; Liu, J. Effects of Cutting Times on Growth and Yield of Alfalfa in the Next Year. Grassl. Chin. 2005, 5, 33–37. (In Chinese) [Google Scholar]
- Rong, Y.P.; Han, J.G.; Wang, P.; Mao, P.S. Effects of Cutting Rate on Carbohydrate Reserves and Nitrogen Content of RussianWildryegrass (Psathyrosta chysperennis). Grassl. Chin. 2000, 8, 279–288. (In Chinese) [Google Scholar]
- Zhong, X.X.; Jiang, H.D.; Cao, W.X.; Gu, H.R. Effect of N fertilization level and defoliation date on Ca, P and Mg mineral concentration in Hybrid Pennisetum and their relationship to ruminant requirements. Acta Pratacult. Sin. 2005, 5, 87–91. (In Chinese) [Google Scholar]
- Hassink, J. Effects of soil texture and grassland management on soil organic C and N and rates of C and N mineralization. Soil Biol. Biochem. 1994, 26, 1221–1231. [Google Scholar] [CrossRef]
- Joseph, C.B.; Douglas, S.C.; David, P.B. Nitrogen and defoliation management: Effects on yield and nutritive of flaccid grass. Agron. J. 1998, 90, 85–92. [Google Scholar]
- Li, B. Effects of Long-Term Fencing Closure on Soil and Vegetation in Mountain Pasture of Bayanbulak. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2009. [Google Scholar]
- Wang, W.; Liang, C.Z.; Liu, Z.L.; Hao, D.Y. Analysis of the Plant Individual Behavior during the Degradation and Restoring Succession in Steppe Community. Chin. J. Plant Ecol. 2000, 24, 268–274. (In Chinese) [Google Scholar]
- Liu, M.L.; Bao, Y.; Yang, C.; Zhang, X.S. Effect of Mowing Systems on Composition of Stipa Grandis Community in Inner Mongolia Steppe. J. Beijing Norm. Univ. (Natl. Sci.) 2007, 43, 83–87. (In Chinese) [Google Scholar]
- Liu, J.L. Study on Impact of Mowing and Grazing on Vegetation and Soil of Typical Steppe in Xilingol, Inner Mongolia. Ph.D. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2013. [Google Scholar]
- Wang, M.J.; Li, Q.F.; Qing, X.L.; Li, J.H. Effects of Cutting Treatments on Plant Growth and Reproductive Status in Stipa baicalensis Steppe I. Effects in the Treating Year and Next Spring. J. Inn. Mong. Agric. Univ. 2002, 23, 20–28. (In Chinese) [Google Scholar]
- Belsky, A.J.; Carson, W.P.; Jenson, C.L. Overcompensation by Plants: Herbivory optimization or red herring. Evolut. Ecol. 1993, 7, 109–121. [Google Scholar] [CrossRef]
- Vail, S.G. Selection for over compensatory plant responses to herbivory: A mechanism for the evolution of plant-herbivore mutualism. Am. Nat. 1992, 139, 1–8. [Google Scholar] [CrossRef]
- Zhong, Y.K.; Bao, Q.H.; Sun, W. A study on reasonable mowing system of natural mowing steppe in Baiyinxile farm, Inner Mongolia. Acta Ecol. Sin. 1991, 13, 242–249. [Google Scholar]
- Van Staalduinen, M.A.; Dobarro, I.; Peco, B. Interactive effects of clipping and nutrient availability on the compensatory growth of a grass species. Plant Ecol. 2010, 208, 55–64. [Google Scholar] [CrossRef]
- Bernhardt-Romermann, M.; Romermann, C.; Sperlich, S.; Schmidt, W. Explaining grassland biomass—The contribution of climate, species and functional diversity depends on fertilization and mowing frequency. J. Appl. Ecol. 2011, 48, 1088–1097. [Google Scholar] [CrossRef]
- Tilman, D.; Lehman, C.L.; Thomson, K.T. Plant diversity and ecosystem productivity: Theoretical considerations. Proc. Natl. Acad. Sci. USA 1997, 94, 1857–1861. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, A.; Yamamura, N. Herbivory promotes plant production and reproduction in nutrient poor conditions effects of plant adaptive phenology. Am. Nat. 2004, 163, 138–153. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.U.; Chapin, F.S., III; Ewel, J.J.; Hector, A.; Inchausti, P. Effects ofbiodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Kassen, R.; Buckling, A.; Bell, G.; Rainey, P.B. Diversity peaks at inter mediate productivity in a laboratory microcosm. Nature 2000, 406, 508–512. [Google Scholar] [CrossRef] [PubMed]
- Rajaniemi, T.K. Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses. J. Ecol. 2002, 90, 316–329. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia 1993, 96, 169–178. [Google Scholar] [CrossRef]
- Nowak, R.S.; Caldwell, M.M. A test of compensatory photosynthesis in the field: Implications for herbivory tolerance. Oecologia 1984, 61, 311–318. [Google Scholar] [CrossRef]
- Rosenthal, J.P.; Kotanen, P.M. Terrestrial plant tolerance to herbivory. Trends Ecol. Evol. 1994, 9, 145–148. [Google Scholar] [CrossRef]
- Paige, K.N.; Whitham, T.G. Overcompensation in response to mammalian herbivory: The advantage of being eaten. Am. Nat. 1987, 129, 407–416. [Google Scholar] [CrossRef]
- Richards, J.; Mueller, R.; Mott, J. Tillering in tussock grasses in relation to defoliation and apical bud removal. Ann. Bot. 1988, 62, 173–179. [Google Scholar]
- Fan, J.; Zhong, H.; Liang, B.; Du, Z. A study on competition among perennial ryegrass and six other species in different conditions of stress and disturbance. Acta Phytoecol. Sin. 2003, 27, 522–530. [Google Scholar] [CrossRef]
- Connell, J. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Kelemen, A.; Török, P.; Valkó, O.; Deák, B.; Miglécz, T.; Tóth, K. Sustaining recovered grasslands is not likely without proper management: Vegetation changes after cessation of mowing. Biodivers. Conserv. 2014, 23, 1–11. [Google Scholar] [CrossRef]
- Valkó, O.; Török, P.; Matus, G.; Tóthmérész, B. Is regular mowing the most appropriate and cost-effective management maintaining diversity and biomass of target forbs in mountain hay meadows? Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 303–309. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | Time of Mowing | |
|---|---|---|
| Twice a year | TAY | June and August of 2011, 2012 |
| Once a year | OAY | End of August in 2010, 2011, 2012 |
| Once every other year | OTY | End of August of 2010 and 2012 |
| No mowing/Enclosure | NM | Starting from early 2010 |
| Species Name | Water Ecotype | Importance Values | |||
|---|---|---|---|---|---|
| TAY | OAY | OTY | NM | ||
| Stipa grandis | Xerophyte | 0.35 | 0.36 | 0.4 | 0.41 |
| Anemarrhena asphodeloides | Mid-Xerophyte | 0.34 | 0.17 | 0.21 | 0.23 |
| Leymus chinensis | Mid-Xerophyte | 0.26 | 0.31 | 0.26 | 0.13 |
| Cleistogenes squarrosa | Xerophyte | 0.015 | 0.025 | ||
| Allium bidentatum | Xerophyte | 0.014 | 0.029 | 0.018 | 0.065 |
| Allium plurifoliatum | Xerophyte | 0.047 | 0.023 | ||
| Carex korshinskii | Mid-Xerophyte | 0.055 | 0.085 | ||
| Allium tuberosum | Mid-Xerophyte | 0.018 | 0.021 | ||
| Allium condensatum | Mid-Xerophyte | 0.056 | |||
| Artemisia frigida | Xerophyte | 0.058 | |||
| Oxytropis myriophylla | Mid-Xerophyte | 0.014 | 0.018 | ||
| summation | 6 | 7 | 8 | 7 | |
| Diversity Index | TAY | OAY | OTY | NM | |
|---|---|---|---|---|---|
| May | Shannon-Wiener | 1.33 ± 0.28 b | 1.30 ± 0.26 b | 1.36 ± 0.13 b | 1.62 ± 0.11 a |
| Pielou | 0.80 ± 0.11 a | 0.84 ± 0.31 a | 0.80 ± 0.12 a | 0.83 ± 0.05 a | |
| Richness | 16.13 ± 1.97 ab | 13.80 ± 1.67 b | 18.50 ± 1.21 a | 11.23 ± 1.22 c | |
| June | Shannon-Wiener | 1.41 ± 0.15 a | 1.32 ± 0.38 b | 1.44 ± 0.07 a | 1.47 ± 0.06 a |
| Pielou | 0.79 ± 0.14 ab | 0.82 ± 0.19 ab | 0.94 ± 0.11 a | 0.75 ± 0.09 ab | |
| Richness | 15.97 ± 1.68 b | 10.69 ± 1.05 c | 24.29 ± 0.57 a | 25.55 ± 3.32 a | |
| July | Shannon-Wiener | 1.42 ± 0.22 ab | 1.29 ± 0.22 b | 1.56 ± 0.15 ab | 1.64 ± 0.18 a |
| Pielou | 0.86 ± 0.16 a | 0.85 ± 0.17 a | 0.92 ± 0.10 a | 0.81 ± 0.07 a | |
| Richness | 19.41 ± 3.65 a | 14.20 ± 1.65 b | 21.81 ± 0.65 a | 15.80 ± 1.78 b | |
| August | Shannon-Wiener | 1.52 ± 0.34 ab | 1.36 ± 0.11 b | 1.78 ± 0.22 a | 1.63 ± 0.15 a |
| Pielou | 0.8 ± 0.18 a | 0.79 ± 0.09 ab | 0.88 ± 0.09 a | 0.81 ± 0.07 a | |
| Richness | 20.5 ± 3.62 c | 12.32 ± 1.03 d | 24.26 ± 0.33 b | 30.63 ± 3.36 a |
| Date | The Individual Biomass (g/Plant) | ||||
|---|---|---|---|---|---|
| TAY | OAY | OTY | NM | ||
| S. grandis | May 2013 | 2.82 ± 0.70 b | 2.26 ± 0.34 b | 6.29 ± 0.44 a | 6.70 ± 0.67 a |
| June 2013 | 5.71 ± 0.47 ab | 3.52 ± 0.41 b | 8.23 ± 0.80 a | 7.77 ± 2.04 a | |
| July 2013 | 5.27 ± 0.63 ab | 3.27 ± 0.39 ab | 5.44 ± 0.46 ab | 13.83 ± 1.07 a | |
| August 2013 | 6.88 ± 1.20 a | 9.06 ± 1.25 a | 12.13 ± 0.92 a | 9.96 ± 2.82 a | |
| L. chinensis | May 2013 | 0.19 ± 0.022 bc | 0.11 ± 0.003 c | 0.21 ± 0.041 b | 0.35 ± 0.009 a |
| June 2013 | 0.31 ± 0.030 a | 0.15 ± 0.017 a | 0.31 ± 0.104 a | 0.48 ± 0.23 a | |
| July 2013 | 0.40 ± 0.062 a | 0.20 ± 0.018 a | 0.30 ± 0.15 a | 0.24 ± 0.017 a | |
| August 2013 | 0.39 ± 0.046 bc | 0.29 ± 0.023 c | 0.48 ± 0.048 b | 0.81 ± 0.022 a | |
| A. asphodeloides | May 2013 | 0.15 ± 0.006 b | 0.10 ± 0.009 c | 0.12 ± 0.012 bc | 0.33 ± 0.017 a |
| June 2013 | 0.25 ± 0.030 ab | 0.14 ± 0.025 b | 0.19 ± 0.014 ab | 0.30 ± 0.056 a | |
| July 2013 | 0.34 ± 0.038 a | 0.17 ± 0.020 b | 0.23 ± 0.046 ab | 0.28 ± 0.047 ab | |
| August 2013 | 0.27 ± 0.050 a | 0.22 ± 0.007 a | 0.24 ± 0.083 a | 0.28 ± 0.12 a | |
| Compensation Index | TAY | OAY | OTY |
|---|---|---|---|
| Stipa grandis | 0.46 c | 0.68 b | 0.99 a |
| Leymus chinensis | 0.88 b | 0.66 c | 1.10 a |
| Anemarrhena asphodeloides | 1.92 b | 1.34 c | 2.69 a |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Z.; Yang, J.; Gu, R.; Liang, Y.; Yan, Y.; Gao, Q.; Yang, J. Influence of Different Mowing Systems on Community Characteristics and the Compensatory Growth of Important Species of the Stipa grandis Steppe in Inner Mongolia. Sustainability 2016, 8, 1121. https://doi.org/10.3390/su8111121
Wan Z, Yang J, Gu R, Liang Y, Yan Y, Gao Q, Yang J. Influence of Different Mowing Systems on Community Characteristics and the Compensatory Growth of Important Species of the Stipa grandis Steppe in Inner Mongolia. Sustainability. 2016; 8(11):1121. https://doi.org/10.3390/su8111121
Chicago/Turabian StyleWan, Zhiqiang, Jiuyan Yang, Rui Gu, Yan Liang, Yulong Yan, Qingzhu Gao, and Jie Yang. 2016. "Influence of Different Mowing Systems on Community Characteristics and the Compensatory Growth of Important Species of the Stipa grandis Steppe in Inner Mongolia" Sustainability 8, no. 11: 1121. https://doi.org/10.3390/su8111121