
Assimilation and Translocation of Dry Matter and Phosphorus in Rice Genotypes Affected by Salt-Alkaline Stress

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Treatment
2.3. Planting, Sampling and Plant Analysis
2.4. Calculation of Dry Matter and Phosphorus Indicators
2.5. Statistical Analysis
3. Results
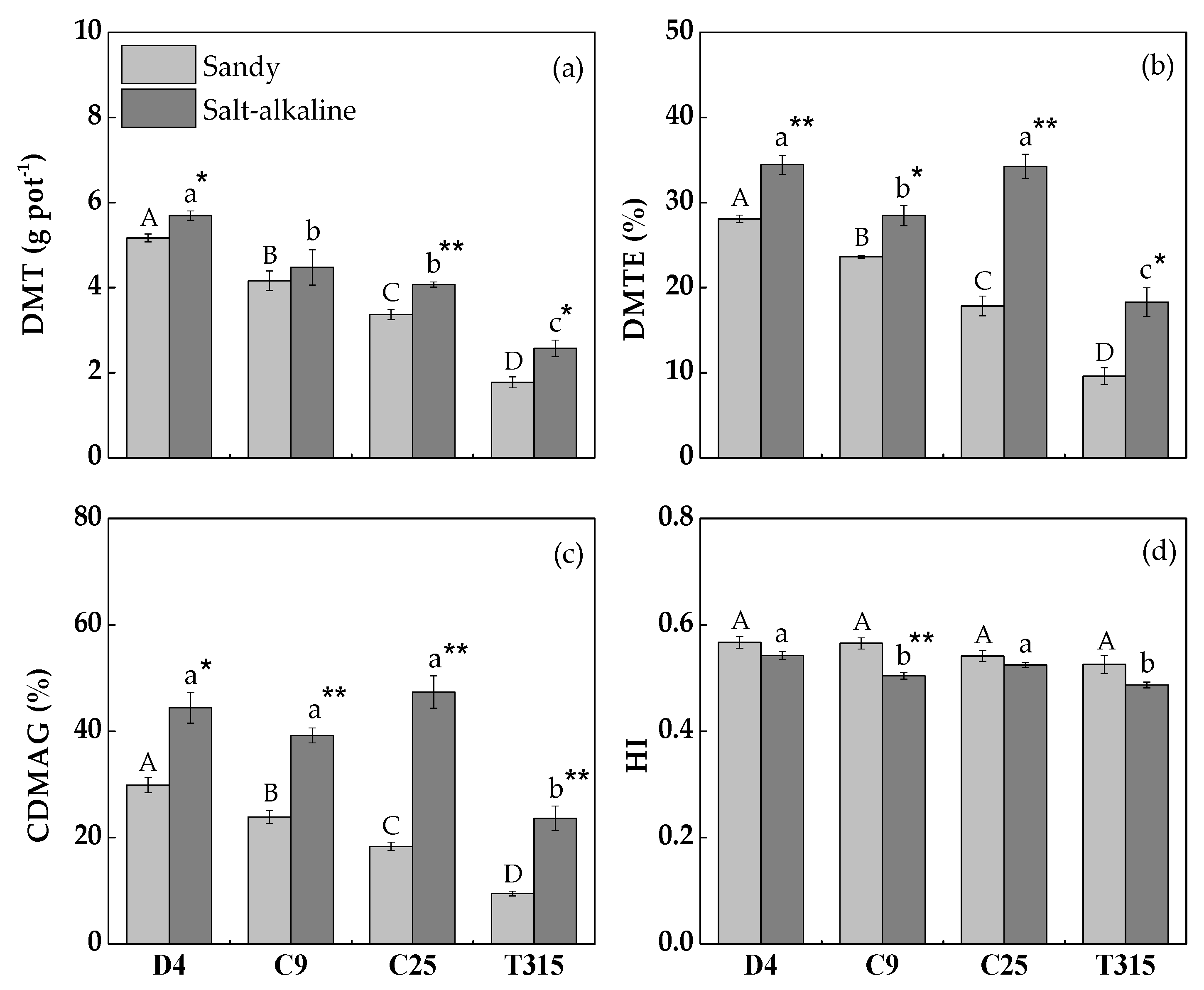
3.1. Dry Matter Accumulation
3.2. Phosphorus Concentration
3.3. Dry Matter Translocation
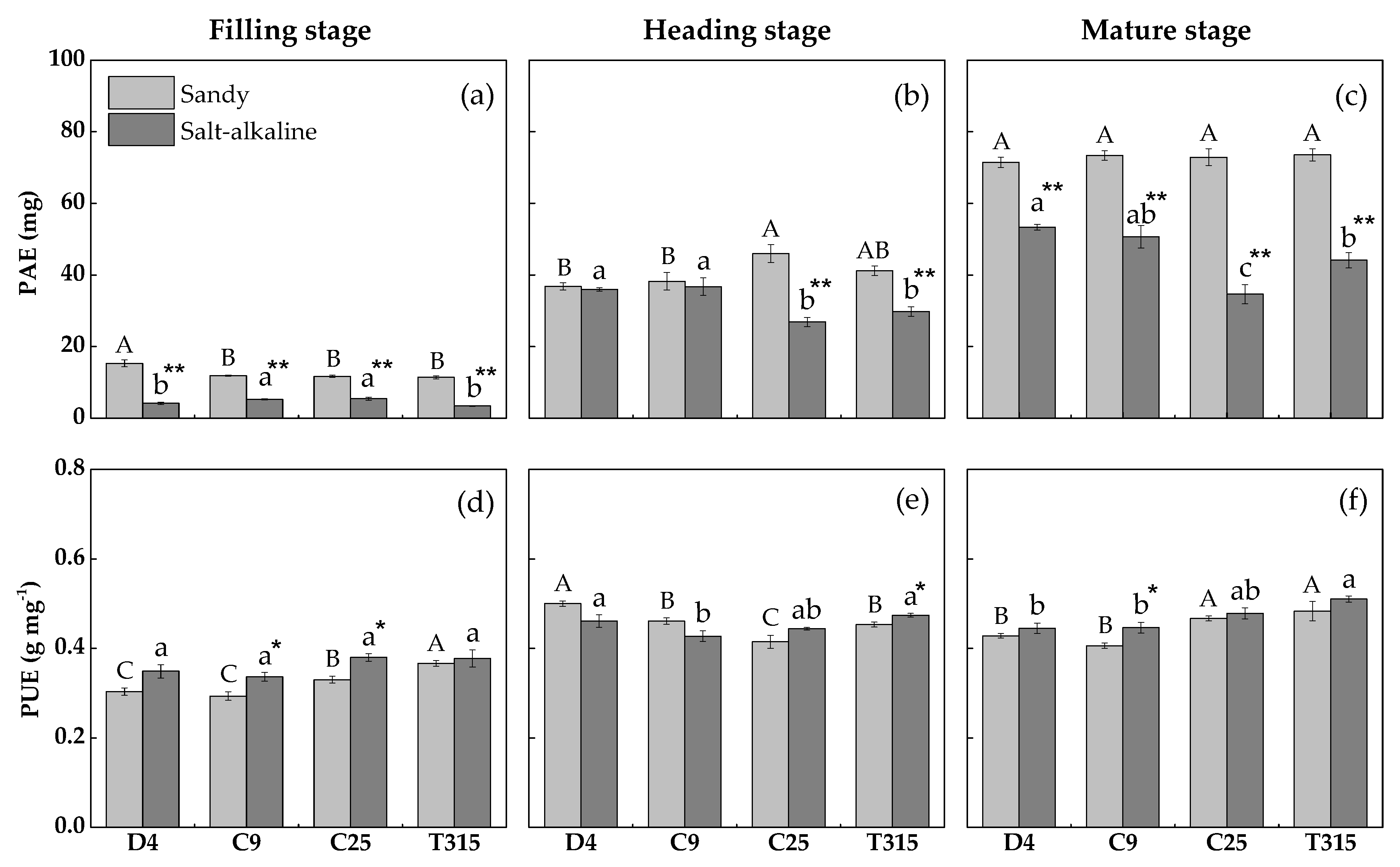
3.4. Phosphorus Acquisition and Utilization Efficiency
3.5. Phosphorus Translocation
4. Discussion
4.1. Dry Matter Accumulation and Translocation
4.2. Phosphate Accumulation and Translocation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Flowers, T.J.; Yeo, A.R. Breeding for salinity resistance in crop plants: Where next? Aust. J. Plant Physiol. 1995, 22, 875–884. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wu, Z.J.; Shi, Y.L.; Wang, R.Y. The resource of saline soil in the Northest China. Chin. J. Soil Sci. 2004, 5, 643–647. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liang, Z.W.; Wang, Z.C.; Chen, Y. Strategic reflections on rice development in saline and alkalineArea in west Jilin, China. Notrh Rice 2007, 6, 7–12. [Google Scholar]
- Eraslan, F.; Inal, A.; Gunes, A.; Alpaslan, M. Impact of exogenous salicylic acid on the growth, antioxidant activity and physiology of carrot plants subjected to combined salinity and boron toxicity. Sci. Hortic Amst. 2007, 113, 120–128. [Google Scholar] [CrossRef]
- Shabbir, G.; Hussain, N.; Bhatti, M.; Ahmad, A.; Javed, M.; Shakir, M.A. Salt tolerance potential of some selected fine rice cultivars. Field Stud. 2001, 1, 1175–1177. [Google Scholar]
- Jampeetong, A.; Brix, H. Effects of NaCl salinity on growth, morphology, photosynthesis and proline accumulation of Salvinia natans. Aquat. Bot. 2009, 91, 181–186. [Google Scholar] [CrossRef]
- Sairam, R.; Tyagi, A. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci. Bangalore 2004, 86, 407–421. [Google Scholar]
- Zhang, J.T.; Mu, C.S. Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius. Soil Sci. Plant Nutr. 2009, 55, 685–697. [Google Scholar] [CrossRef]
- Lv, B.S.; Li, X.W.; Ma, H.Y.; Sun, Y.; Wei, L.X.; Jiang, C.J.; Liang, Z.W. Differences in growth and physiology of rice in response to different saline-alkaline stress factors. Agron. J. 2013, 105, 1119–1128. [Google Scholar] [CrossRef]
- Wang, K.; Cui, K.H.; Liu, G.L.; Xie, W.B.; Yu, H.H.; Pan, J.F.; Huang, J.L.; Nie, L.X.; Shah, F.; Peng, S.B. Identification of quantitative trait loci for phosphorus use efficiency traits in rice using a high density SNP map. BMC Genet. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant. Nutr. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Dahiya, S.; Singh, M. Effect of salinity, alkalinity and iron application on the availability of iron, manganese, phosphorus and sodium in pea (Pisum sativum L.) crop. Plant Soil 1976, 44, 697–702. [Google Scholar] [CrossRef]
- Papakosta, D.K. Phosphorus accumulation and translocation in wheat as affected by cultivar and nitrogen-fertilization. J. Agron. Crop Sci. 1994, 173, 260–270. [Google Scholar] [CrossRef]
- Zribi, O.T.; Barhoumi, Z.; Kouas, S.; Ghandour, M.; Slama, I.; Abdelly, C. Insights into the physiological responses of the facultative halophyte Aeluropus littoralis to the combined effects of salinity and phosphorus availability. J. Plant Physiol. 2015, 189, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karlen, D.L.; Whitney, D.A. Dry-matter accumulation, mineral concentrations, and nutrient distribution in winter-wheat. Agron. J. 1980, 72, 281–288. [Google Scholar] [CrossRef]
- Uhde-Stone, C.; Gilbert, G.; Johnson, J.M.F.; Litjens, R.; Zinn, K.E.; Temple, S.J.; Vance, C.P.; Allan, D.L. Acclimation of white lupin to phosphorus deficiency involves enhanced expression of genes related to organic acid metabolism. Plant Soil 2003, 248, 99–116. [Google Scholar] [CrossRef]
- Liu, E.K.; Mei, X.R.; Yan, C.R.; Gong, D.Z.; Zhang, Y.Q. Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agr. Water Manag. 2016, 167, 75–85. [Google Scholar] [CrossRef]
- Richardson, A.E.; Simpson, R.J. Soil microorganisms mediating phosphorus availability. Plant. Physiol. 2011, 156, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Przulj, N.; Momcilovic, V. Genetic variation for dry matter and nitrogen accumulation and translocation in two-rowed spring barley II. Nitrogen translocation. Eur. J. Agron. 2001, 15, 255–265. [Google Scholar] [CrossRef]
- Arduini, I.; Masoni, A.; Ercoli, L.; Mariotti, M. Grain yield, and dry matter and nitrogen accumulation and remobilization in durum wheat as affected by variety and seeding rate. Eur. J. Agron. 2006, 25, 309–318. [Google Scholar] [CrossRef]
- Yang, J.-F.; He, L.-Y.; Zuo, D.-X.; Liu, Y.-F.; Wu, Z.-H.; Zhang, A.-Q.; Zhao, H.-E.; Liu, W.; Yan, C.; Men, Y.-Y. Phosphorous nutritional characteristics of rice in P-deficient soils with different pH values. Plant Nutr. Fertil. Sci. 2009, 15, 62–68. [Google Scholar]
- Gemenet, D.C.; Hash, C.T.; Sanogo, M.D.; Sy, O.; Zangre, R.G.; Leiser, W.L.; Haussmann, B.I. Phosphorus uptake and utilization efficiency in West African pearl millet inbred lines. Field Crops Res. 2015, 171, 54–66. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Wu, Z.H.; He, L.Y.; Zuo, D.X.; Yang, J.F.; Men, Y.Y. Characteristics of phosphorus nutrition of different rice genotypes under low-P stress at different growth stages. Chin. J. Rice Sci. 2008, 22, 71–76. [Google Scholar]
- Darwish, E.; Testerink, C.; Khalil, M.; El-Shihy, O.; Munnik, T. Phospholipid signaling responses in salt-stressed rice leaves. Plant Cell Physiol. 2009, 50, 986–997. [Google Scholar] [CrossRef] [PubMed]
- Borras, L.; Slafer, G.A.; Otegui, M.E. Seed dry weight response to source-sink manipulations in wheat, maize and soybean: A quantitative reappraisal. Field Crops Res. 2004, 86, 131–146. [Google Scholar] [CrossRef]
- Turkan, I.; Demiral, T. Recent developments in understanding salinity tolerance. Environ. Exp. Bot. 2009, 67, 2–9. [Google Scholar] [CrossRef]
- Flowers, T.J.; Flowers, S.A. Why does salinity pose such a difficult problem for plant breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, Z.; Khan, M.A.; Flowers, T. Causes of sterility in seed set of rice under salinity stress. J. Agron. Crop Sci. 2001, 187, 25–32. [Google Scholar] [CrossRef]
- Nemati, I.; Moradi, F.; Gholizadeh, S.; Esmaeili, M.; Bihamta, M. The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L.) seedlings. Plant Soil Environ. 2011, 57, 26–33. [Google Scholar]
- Naheed, G.; Shahbaz, M.; Akram, N.A. Interactive effect of rooting medium application of phosphorus and NaCl on plant biomass and mineral nutrients of rice (Oryza sativa L.). Pak J Bot 2008, 40, 1601–1608. [Google Scholar]
- Elgharably, A. Wheat response to combined application of nitrogen and phosphorus in a saline sandy loam soil. Soil Sci. Plant Nutr. 2011, 57, 396–402. [Google Scholar] [CrossRef]
- Cui, H.; Takeoka, Y.; Wada, T. Effect of sodium chloride on the panicle and spikelet morphogenesis in rice [Oryza sativa], 2: Developmental morphology of the panicle. Jpn. J. Crop Sci. 1995, 64, 593–600. [Google Scholar] [CrossRef]
- Rao, P.S.; Mishra, B.; Gupta, S.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar] [CrossRef]
- Nieman, R.H.; Clark, R.A. Interactive effects of salinity and phosphorus nutrition on concentrations of phosphate and phosphate esters in mature photosynthesizing corn leaves. Plant Physiol. 1976, 57, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Hakim, M.; Juraimi, A.S.; Begum, M.; Hanafi, M.; Ismail, M.R.; Selamat, A. Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). Afr. J. Biotechnol. 2010, 9, 1911–1918. [Google Scholar]
- Khan, M.A.; Abdullah, Z. Salinity-sodicity induced changes in reproductive physiology of rice (Oryza sativa) under dense soil conditions. Environ. Exp. Bot. 2003, 49, 145–157. [Google Scholar] [CrossRef]
- Asch, F.; Dingkuhn, M.; Wittstock, C.; Doerffling, K. Sodium and potassium uptake of rice panicles as affected by salinity and season in relation to yield and yield components. Plant Soil 1999, 207, 133–145. [Google Scholar] [CrossRef]
- Fabre, D.; Siband, P.; Dingkuhn, M. Characterizing stress effects on rice grain development and filling using grain weight and size distribution. Field Crops Res. 2005, 92, 11–16. [Google Scholar] [CrossRef]
- Masoni, A.; Ercoli, L.; Mariotti, M.; Arduini, I. Post-anthesis accumulation and remobilization of dry matter, nitrogen and phosphorus in durum wheat as affected by soil type. Eur. J. Agron. 2007, 26, 179–186. [Google Scholar] [CrossRef]
- Van Herwaarden, A.F.; Angus, J.F.; Richards, R.A.; Farquhar, G.D. “Haying-off”, the negative grain yield response of dryland wheat to nitrogen fertiliser—II. Carbohydrate and protein dynamics. Aust. J. Agric. Res. 1998, 49, 1083–1093. [Google Scholar] [CrossRef]
- Qadir, M.; Qureshi, R.H.; Ahmad, N. Amelioration of calcareous saline sodic soils through phytoremediation and chemical strategies. Soil Use Manag. 2002, 18, 381–385. [Google Scholar] [CrossRef]
- Grattan, S.R.; Grieve, C.M. Salinity mineral nutrient relations in horticultural crops. Sci Hortic Amst. 1999, 78, 127–157. [Google Scholar] [CrossRef]
- Martinez, V.; Lächli, A. Phosphorus translocation in salt-stressed cotton. Physiol. Plant. 1991, 83, 627–632. [Google Scholar] [CrossRef]
- Bauer, A.; Frank, A.B.; Black, A.L. Aerial parts of hard red spring wheat 1. Dry-matter distribution by plant development stage. Agron. J. 1987, 79, 845–852. [Google Scholar] [CrossRef]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source-sink relations. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Awad, A.; Edwards, D.; Campbell, L. Phosphorus enhancement of salt tolerance of tomato. Crop Sci. 1990, 30, 123–128. [Google Scholar] [CrossRef]
- Zribi, O.T.; Houmani, H.; Kouas, S.; Slama, I.; Ksouri, R.; Abdelly, C. Comparative study of the interactive effects of salinity and phosphorus availability in wild (Hordeum maritimum) and cultivated barley (H-vulgare). J. Plant Growth Regul. 2014, 33, 860–870. [Google Scholar] [CrossRef]
- Tahir, I.S.A.; Nakata, N. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop Sci. 2005, 191, 106–115. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soils | EC 1:5 μS·cm−1 | pH | Total N·g·kg−1 | Total P g·kg−1 | Available P mg·kg−1 | Organic Matter % | CEC cmol·kg−1 | ESP | Na+ mg·kg−1 | HCO3− mg·kg−1 |
|---|---|---|---|---|---|---|---|---|---|---|
| SS | 86.6 | 7.59 | 461.8 | 163.59 | 2.68 | 0.99 | 9.74 | 1.15 | 2.74 | 292.8 |
| SAS | 358 | 8.81 | 535.9 | 254.63 | 9.39 | 1.24 | 17.33 | 7.59 | 347 | 770.4 |
| Genotypes | Dry Matter (g) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Stem | Leaf | Grain | |||||||
| (TS) | SS | SAS | rDMR | SS | SAS | rDMR | SS | SAS | rDMR |
| D4 | 2.46 | 0.79 | 67.73a ** | 2.17 | 0.65 | 69.98a ** | |||
| C9 | 1.96 | 1.01 | 48.55b ** | 1.43 | 0.76 | 46.90c ** | |||
| C25 | 1.88 | 1.16 | 38.18c ** | 1.99 | 0.86 | 56.58b ** | |||
| T315 | 2.35 | 0.71 | 69.89a ** | 1.83 | 0.57 | 68.83a ** | |||
| (HS) | |||||||||
| D4 | 11.77 | 10.48 | 10.88a | 4.09 | 4.05 | 1.01c | 2.55 | 2.02 | 20.33b * |
| C9 | 11.96 | 10.68 | 10.64c | 3.61 | 3.41 | 5.60c | 2.02 | 1.56 | 22.89b |
| C25 | 12.33 | 7.92 | 35.78c ** | 4.32 | 2.79 | 35.57a ** | 2.35 | 1.23 | 47.60a ** |
| T315 | 12.20 | 9.04 | 25.87a ** | 4.19 | 3.48 | 16.94b ** | 2.23 | 1.59 | 28.11b |
| (MS) | |||||||||
| D4 | 9.61 | 8.03 | 16.40c * | 3.63 | 2.83 | 22.13d | 17.34 | 12.89 | 25.67c ** |
| C9 | 10.39 | 8.45 | 18.52c * | 3.04 | 2.73 | 10.32c | 17.42 | 11.38 | 34.74bc ** |
| C25 | 11.98 | 5.78 | 51.87a ** | 3.65 | 2.09 | 42.85a ** | 18.39 | 8.68 | 52.94a ** |
| T315 | 12.85 | 8.74 | 31.91b ** | 4.00 | 2.80 | 30.01b ** | 18.63 | 10.95 | 40.98b ** |
| Genotypes | Phosphorus Concentration (mg·g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Stem | Leaf | Grain | |||||||
| (TS) | SS | SAS | rPC | SS | SAS | rPC | SS | SAS | rPC |
| D4 | 3.78 | 3.14 | 0.83b | 2.76 | 2.57 | 0.93a | |||
| C9 | 3.63 | 3.25 | 0.89ab | 3.13 | 2.61 | 0.84a ** | |||
| C25 | 3.45 | 2.82 | 0.82b ** | 2.64 | 2.41 | 0.92a | |||
| T315 | 3.01 | 3.00 | 1.00a | 2.38 | 2.25 | 0.95a | |||
| (HS) | |||||||||
| D4 | 2.04 | 2.16 | 1.06ab | 2.08 | 2.29 | 1.11a * | 1.68 | 2.03 | 1.21a * |
| C9 | 2.15 | 2.48 | 1.15a | 2.46 | 2.18 | 0.89b | 1.76 | 1.76 | 1.01b |
| C25 | 2.62 | 2.33 | 0.89c | 2.17 | 2.16 | 1.00ab | 1.79 | 1.99 | 1.12ab |
| T315 | 2.25 | 2.10 | 0.94bc * | 2.33 | 2.07 | 0.89b | 1.77 | 2.23 | 1.26a ** |
| (MS) | |||||||||
| D4 | 0.81 | 0.84 | 1.04a | 1.19 | 1.36 | 1.15a * | 3.43 | 3.33 | 0.97a |
| C9 | 0.94 | 0.90 | 0.96a | 1.23 | 1.05 | 0.86b | 3.66 | 3.53 | 0.96a |
| C25 | 0.80 | 0.84 | 1.05a | 0.92 | 1.04 | 1.13a * | 3.26 | 3.19 | 0.98a |
| T315 | 0.62 | 0.68 | 1.11a | 0.72 | 0.91 | 1.26a ** | 3.38 | 3.25 | 0.97a |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Li, J.; Jia, X.; Yang, F.; Wang, Z. Assimilation and Translocation of Dry Matter and Phosphorus in Rice Genotypes Affected by Salt-Alkaline Stress. Sustainability 2016, 8, 568. https://doi.org/10.3390/su8060568
Tian Z, Li J, Jia X, Yang F, Wang Z. Assimilation and Translocation of Dry Matter and Phosphorus in Rice Genotypes Affected by Salt-Alkaline Stress. Sustainability. 2016; 8(6):568. https://doi.org/10.3390/su8060568
Chicago/Turabian StyleTian, Zhijie, Jingpeng Li, Xueying Jia, Fu Yang, and Zhichun Wang. 2016. "Assimilation and Translocation of Dry Matter and Phosphorus in Rice Genotypes Affected by Salt-Alkaline Stress" Sustainability 8, no. 6: 568. https://doi.org/10.3390/su8060568
APA StyleTian, Z., Li, J., Jia, X., Yang, F., & Wang, Z. (2016). Assimilation and Translocation of Dry Matter and Phosphorus in Rice Genotypes Affected by Salt-Alkaline Stress. Sustainability, 8(6), 568. https://doi.org/10.3390/su8060568