Estimation of Leaf Area Index in a Mountain Forest of Central Japan with a 30-m Spatial Resolution Based on Landsat Operational Land Imager Imagery: An Application of a Simple Model for Seasonal Monitoring



Abstract
:
1. Introduction
2. Materials and Methods
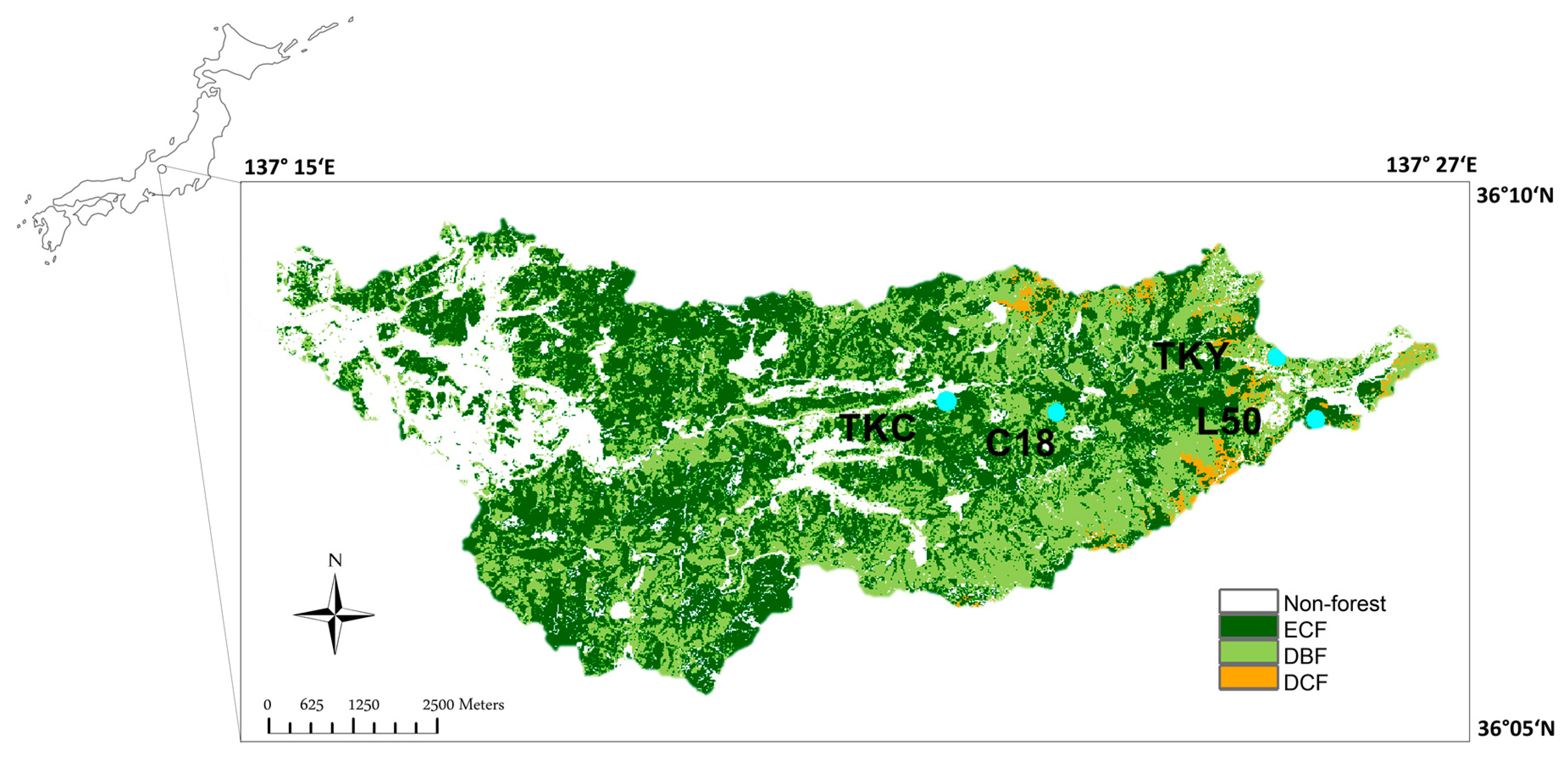
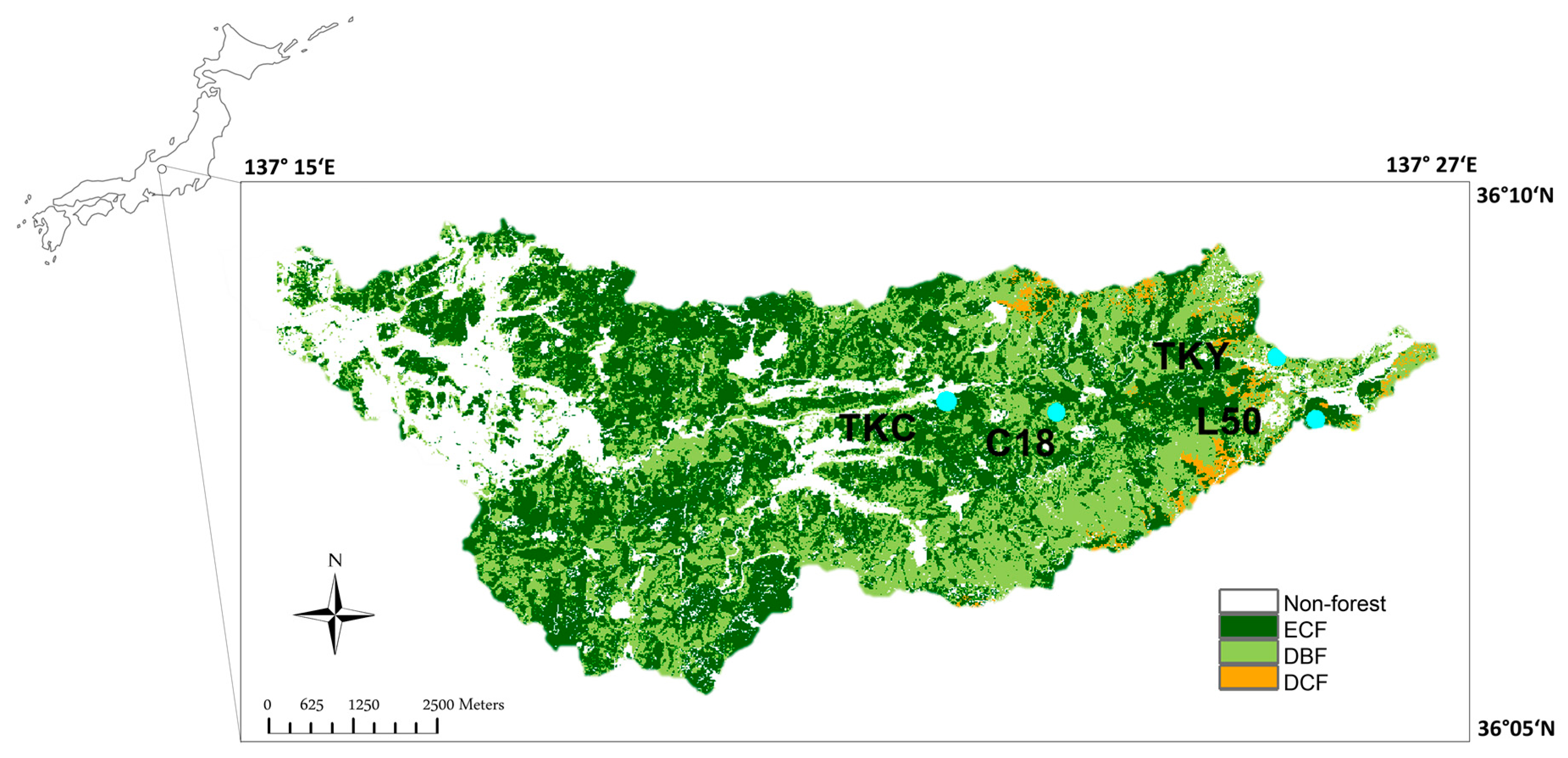
2.1. Study Area and In Situ Measurements
2.2. Pre-Processing of Satellite Imagery
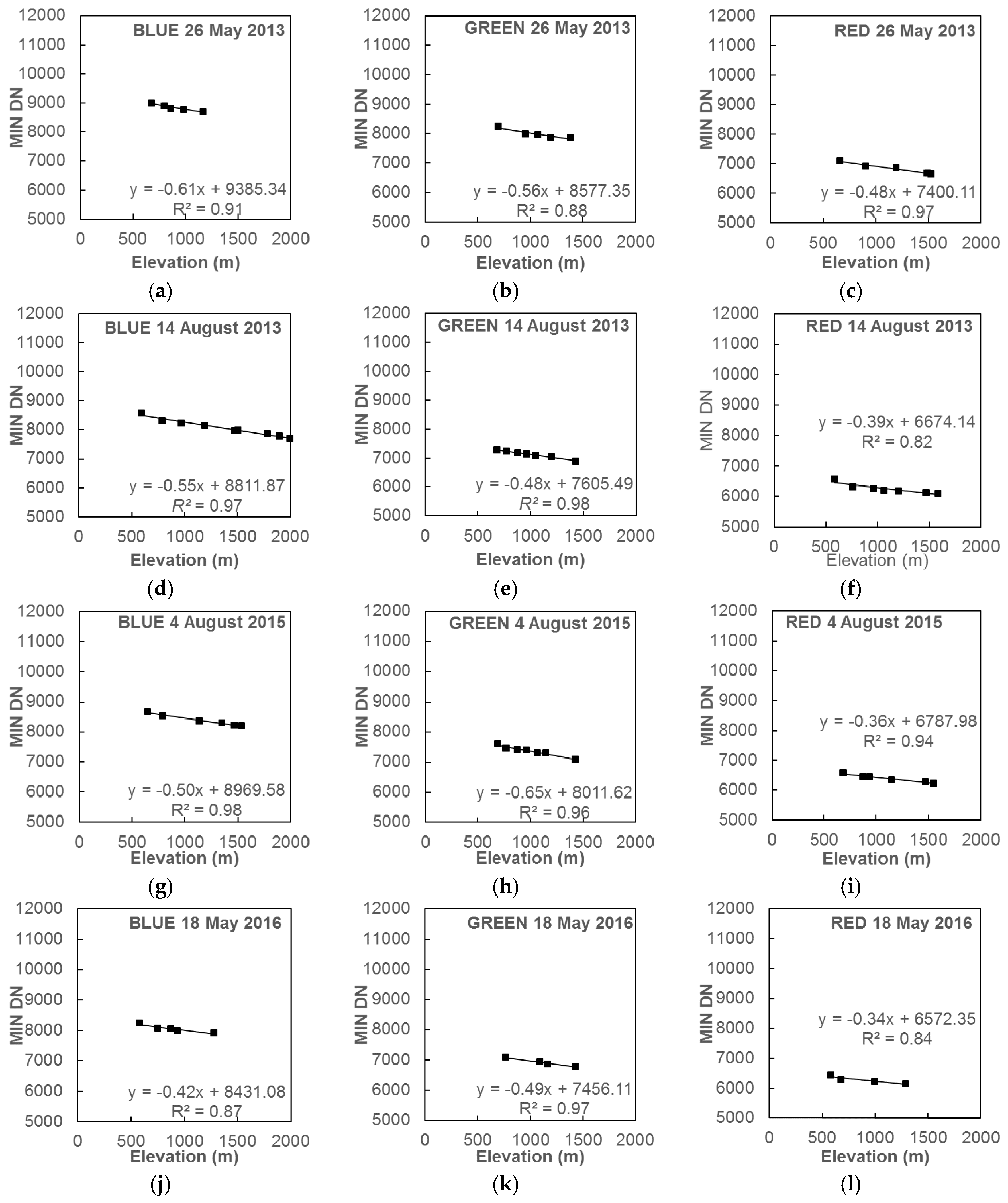
2.2.1. Atmospheric Correction
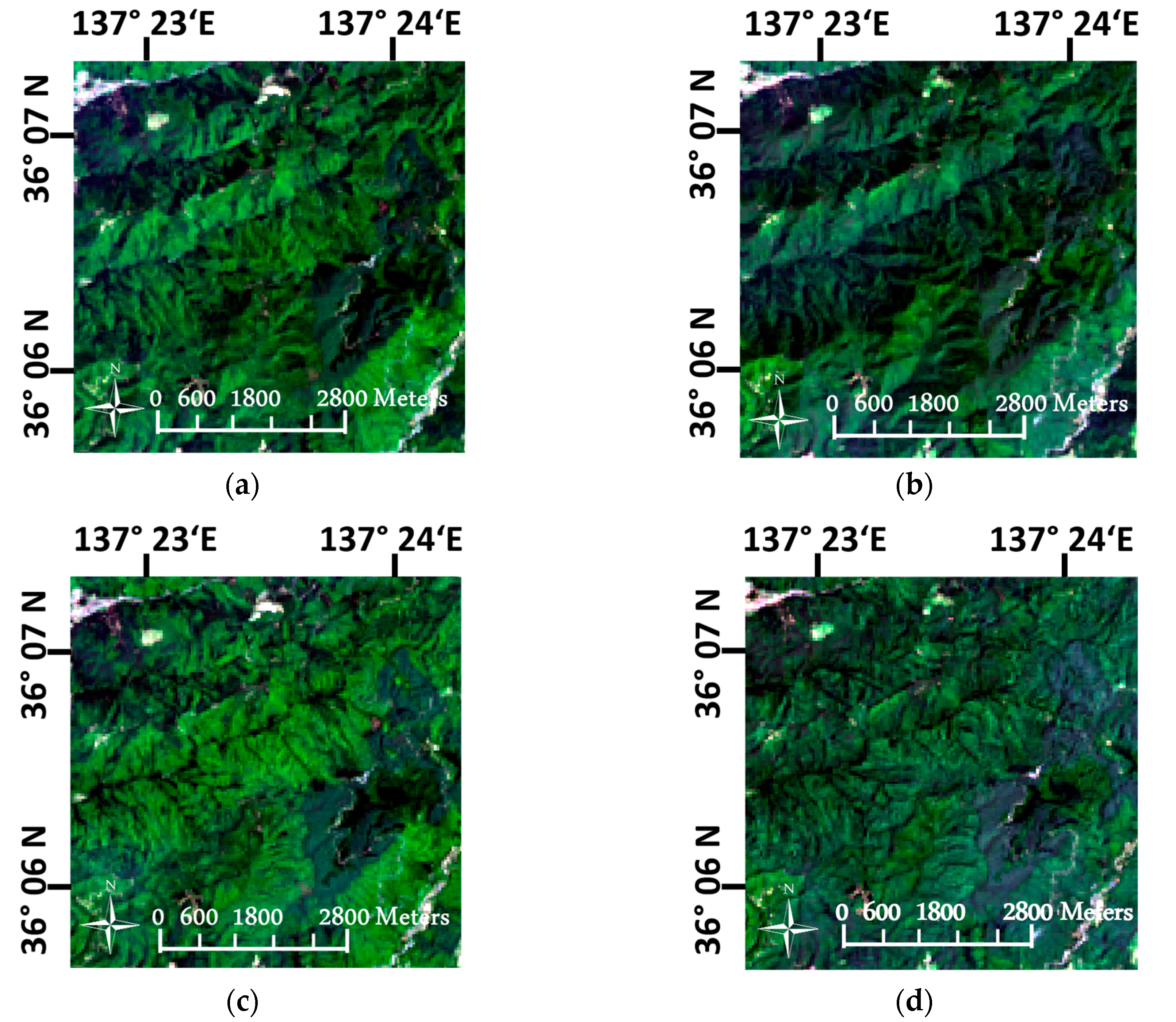
2.2.2. Topographic Correction
2.3. Leaf Area Index Model
3. Results
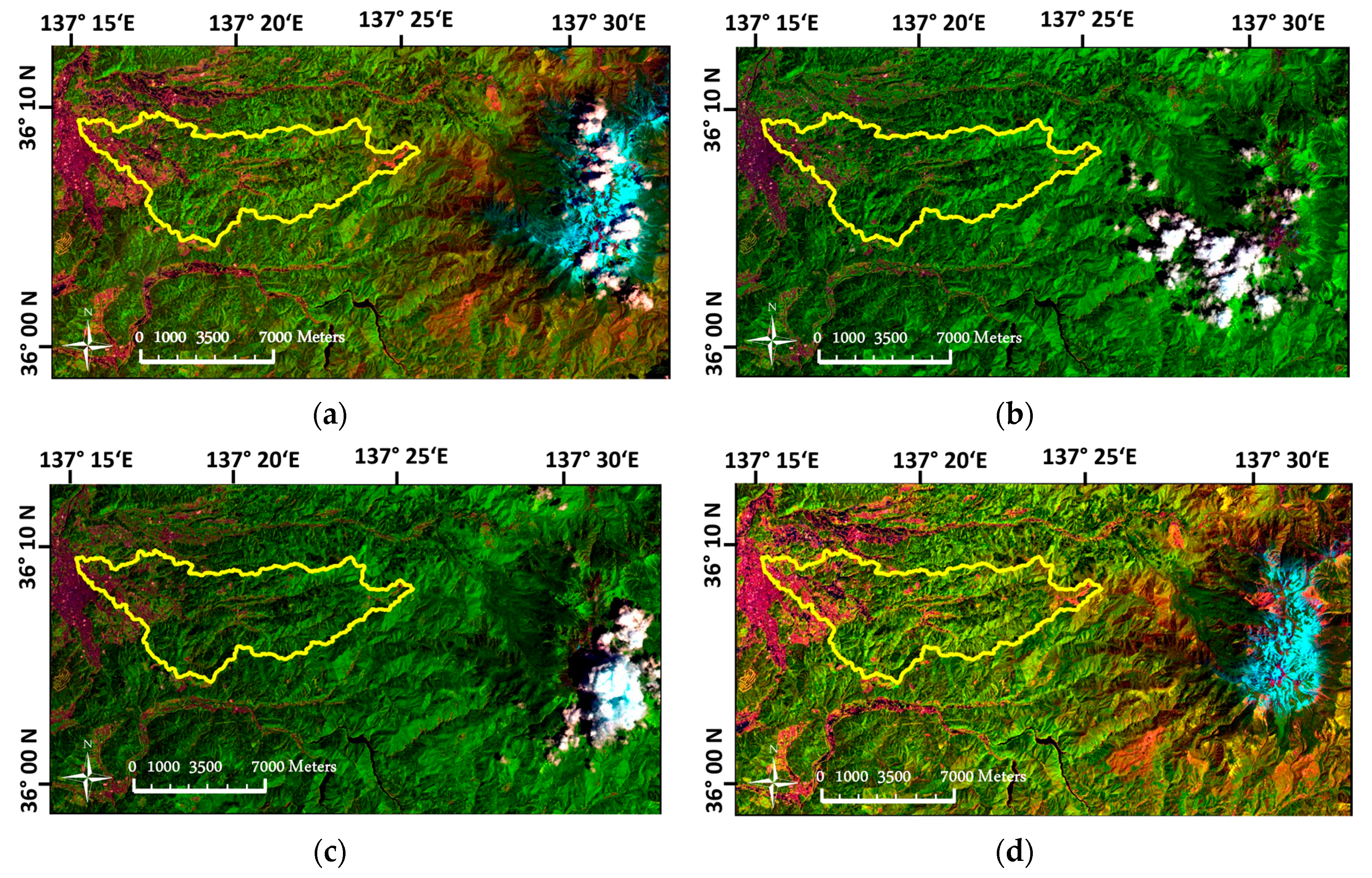
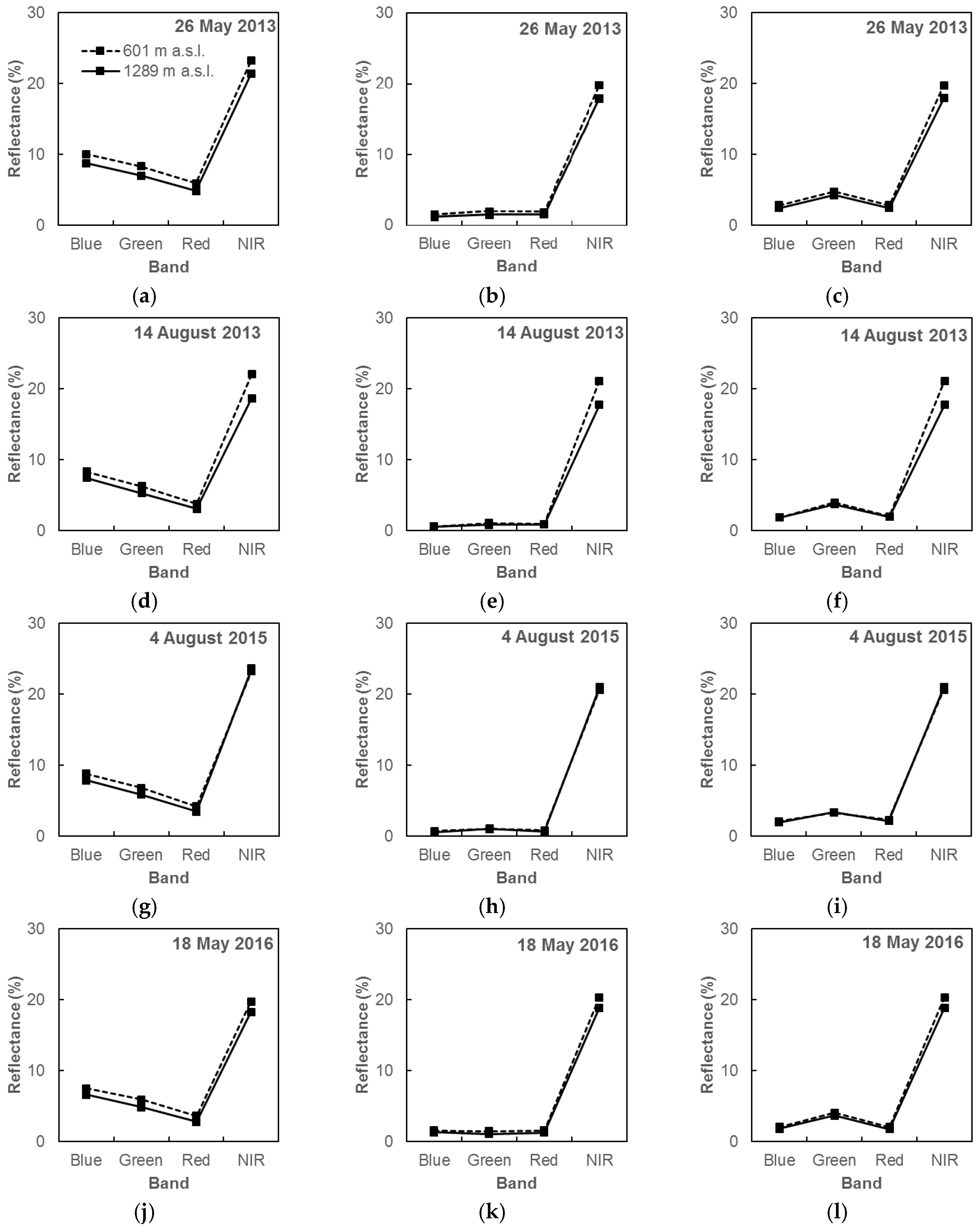
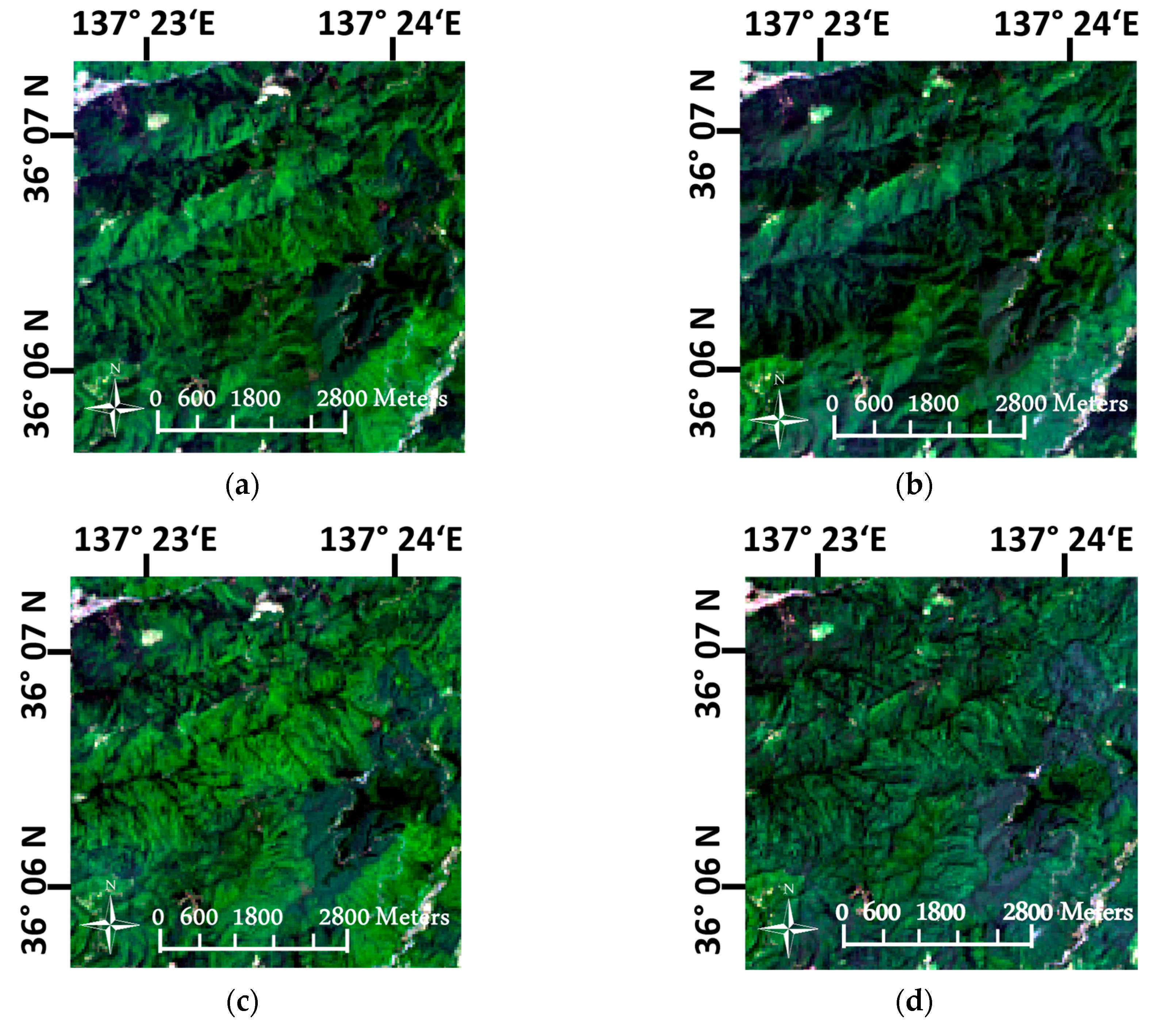
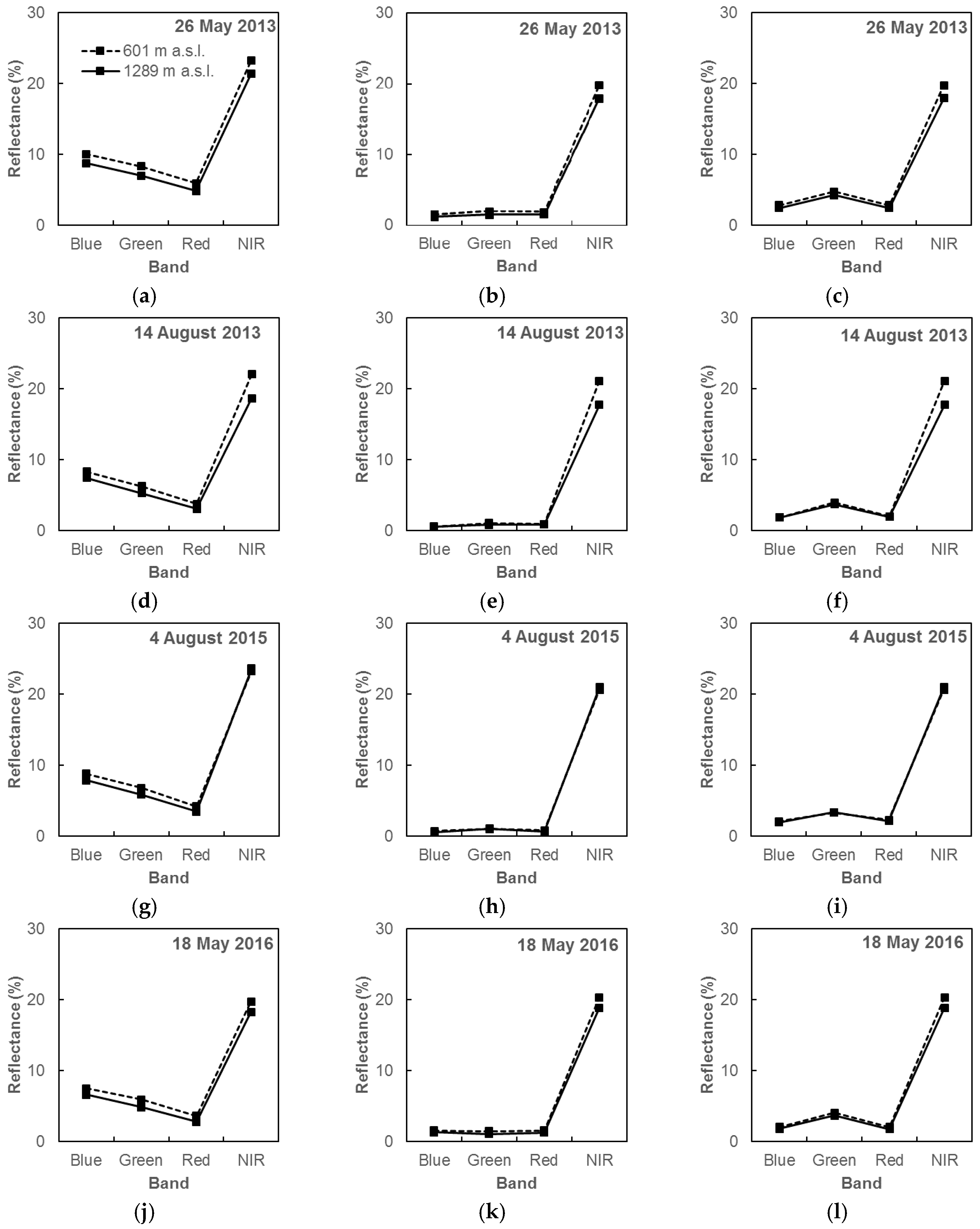
3.1. Pre-Processing of Landsat OLI Imagery
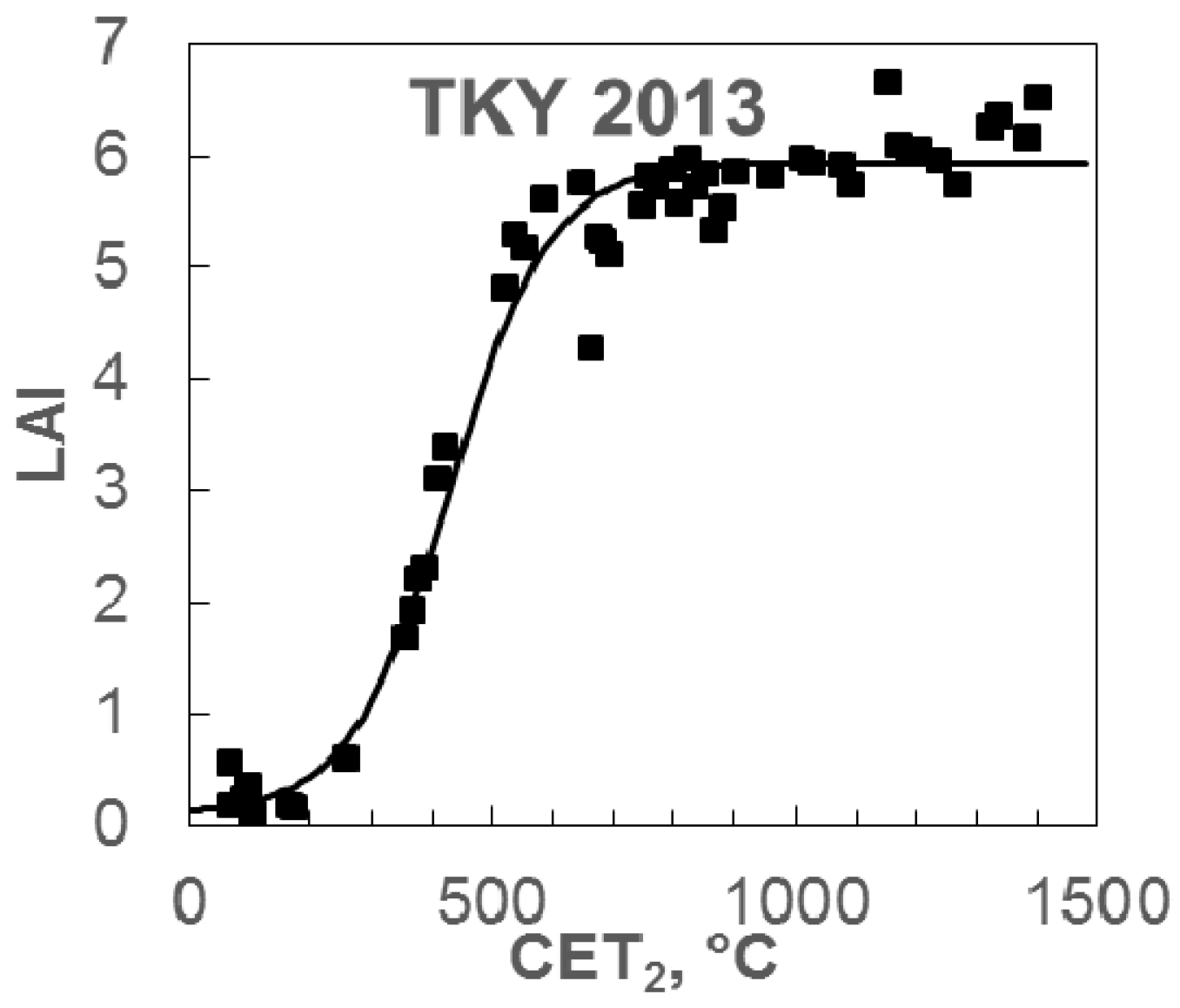
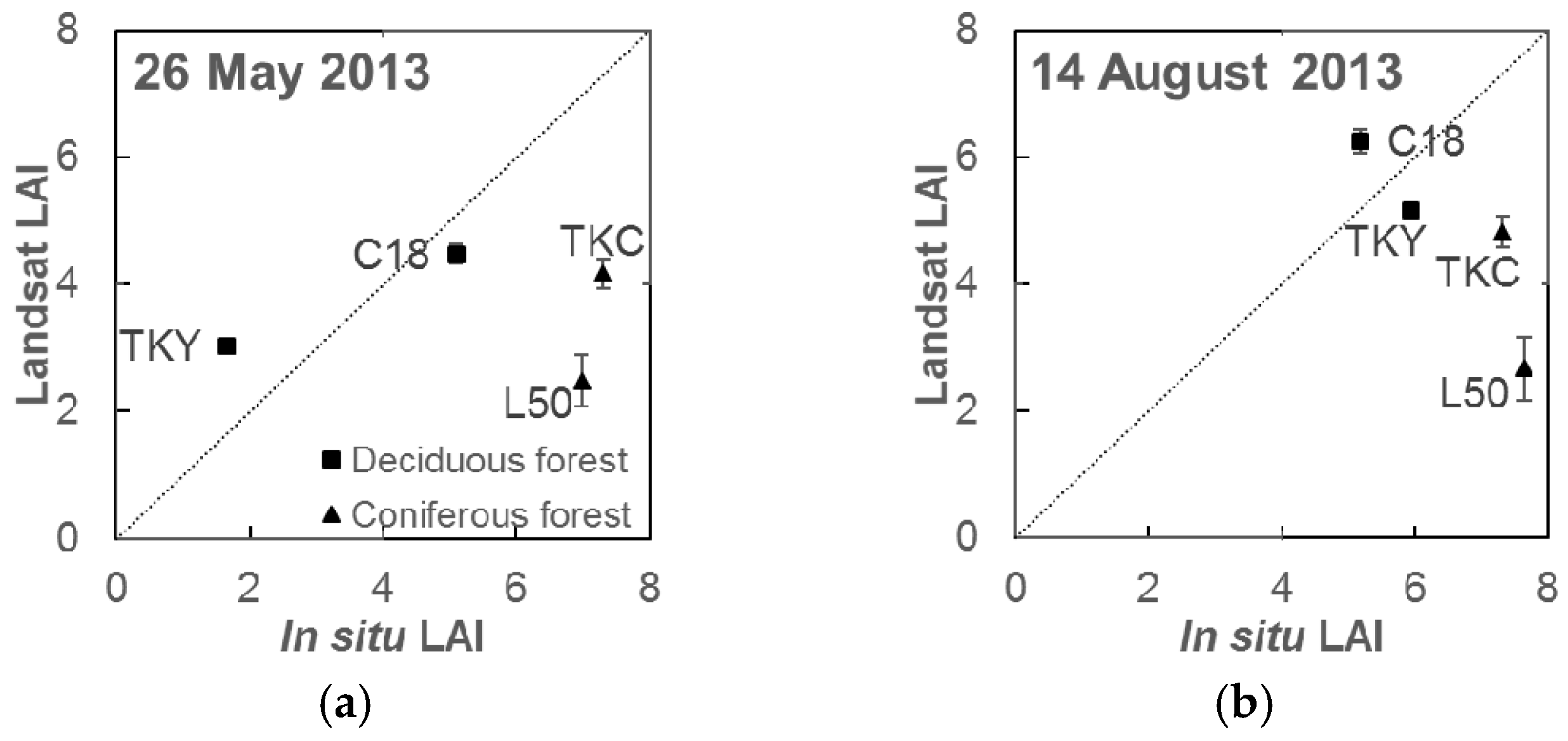
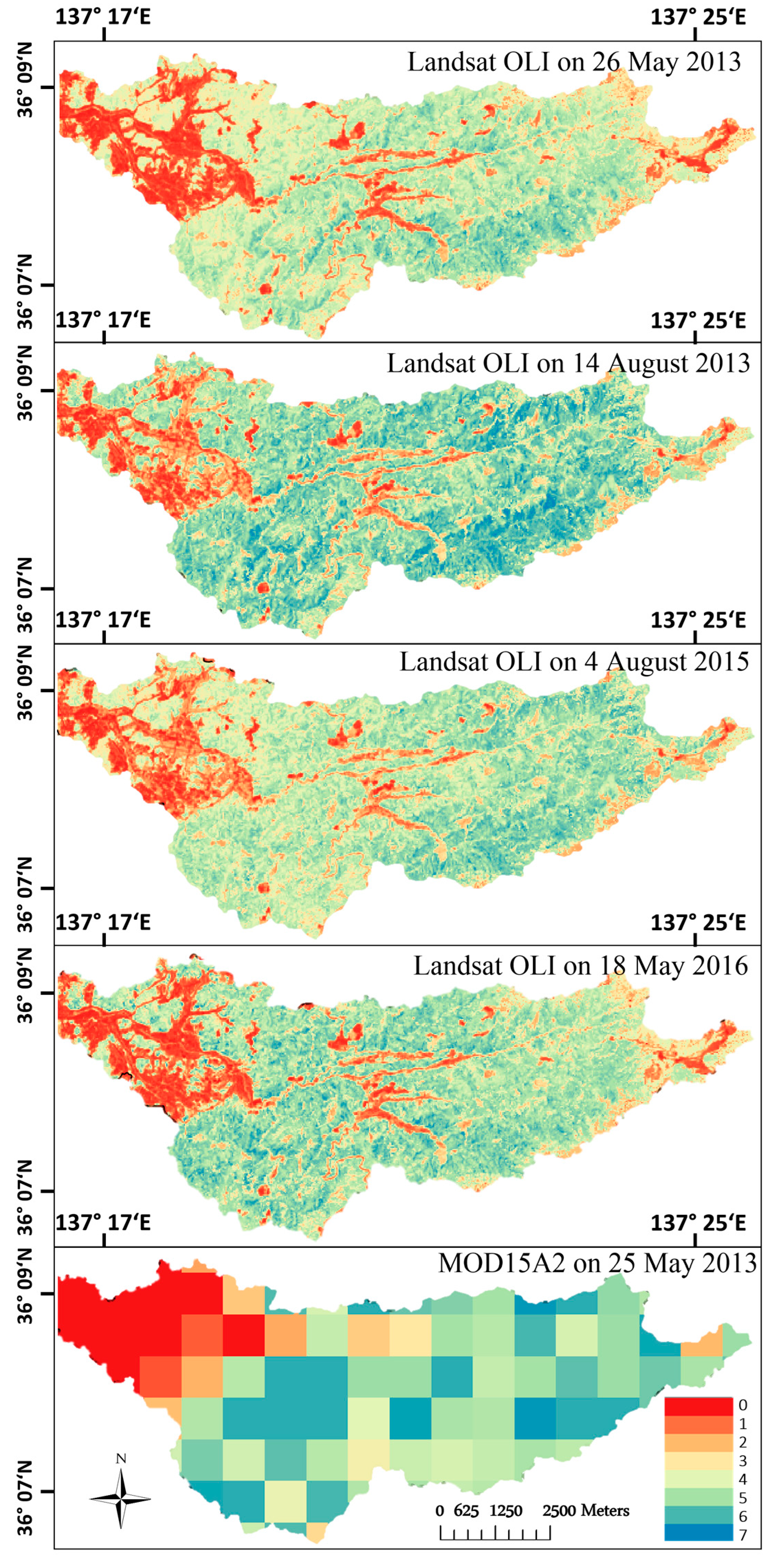
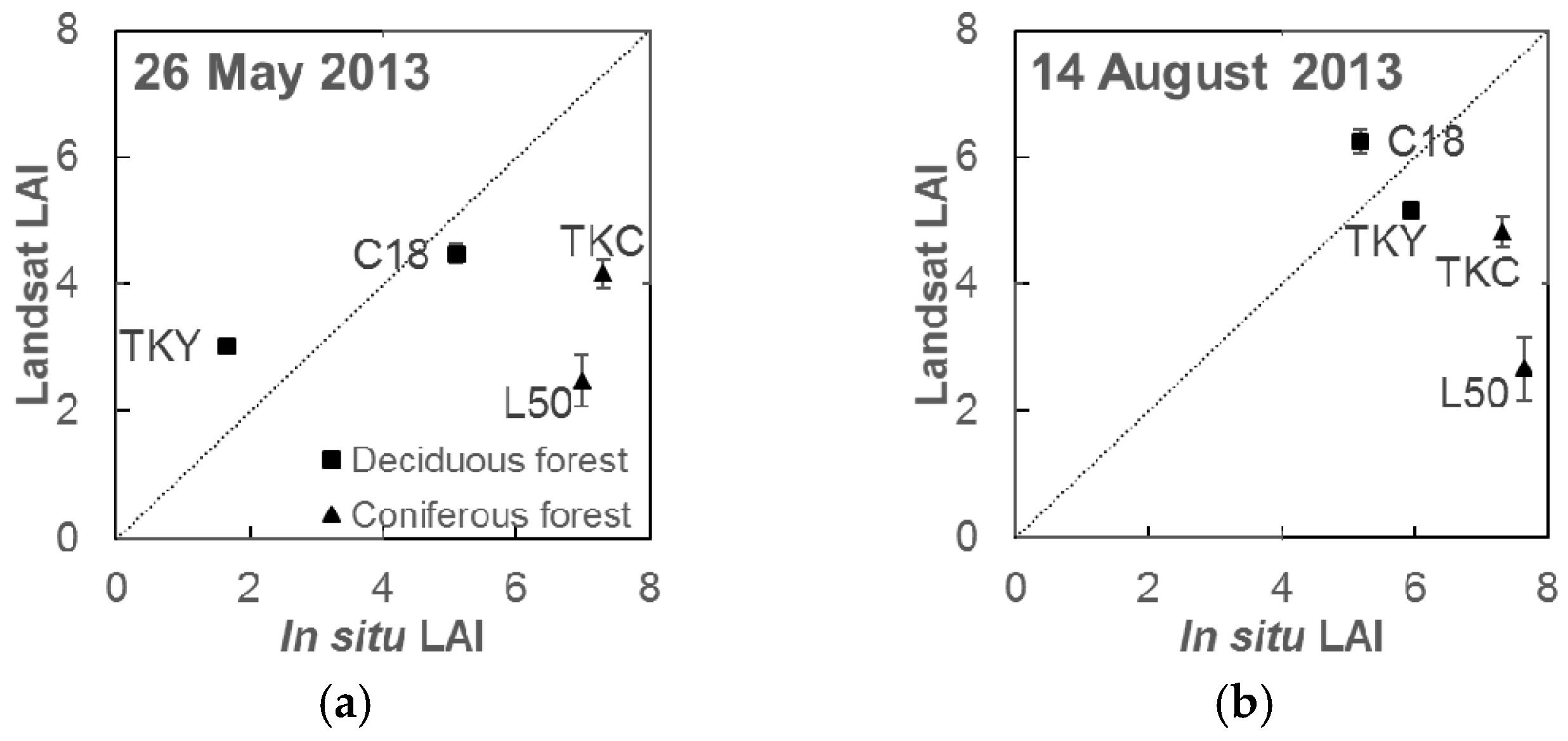
3.2. LAI Model Performance and Validation
4. Discussion
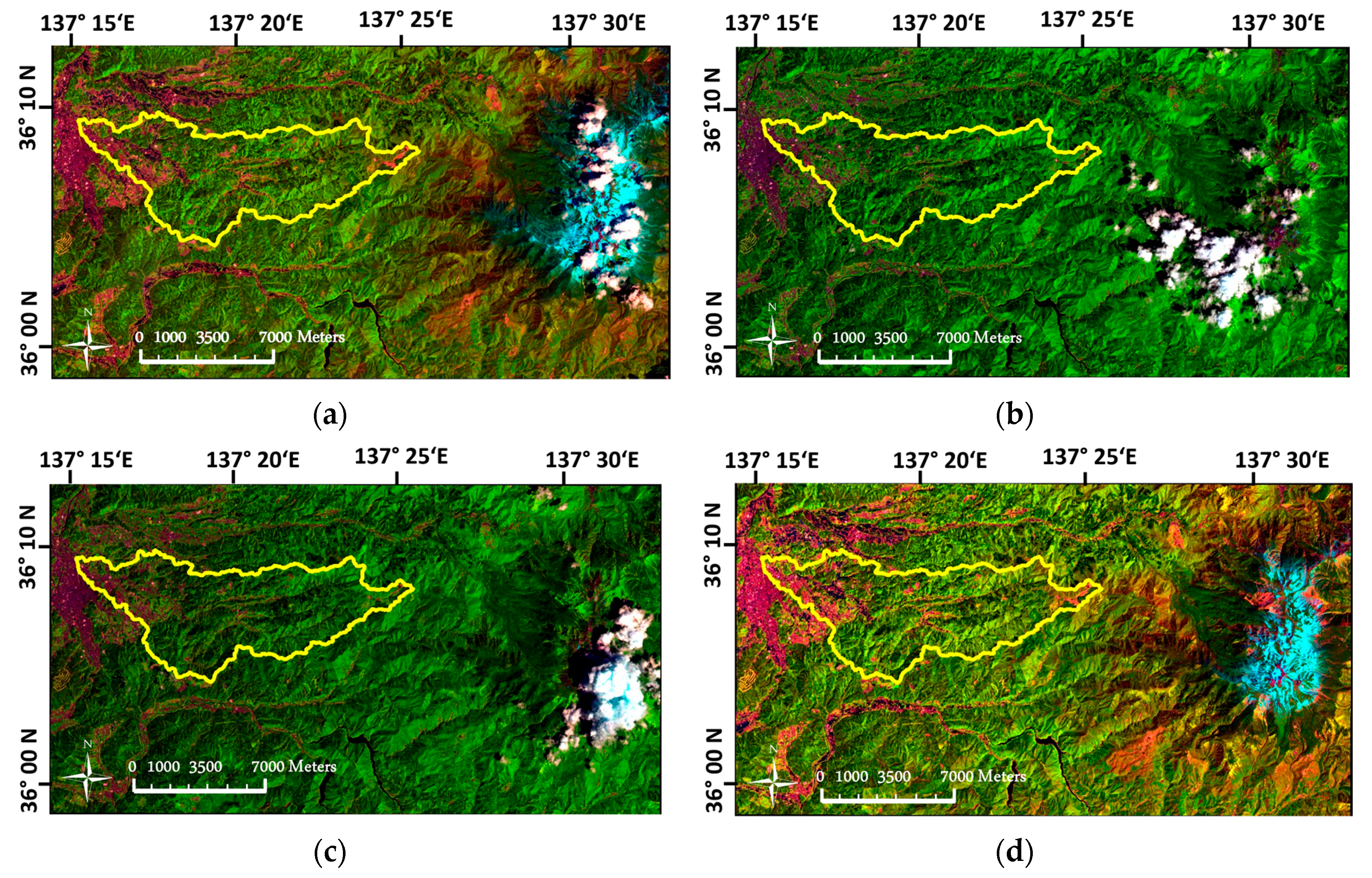
4.1. Imagery Pre-Processing in the Mountainous Landscape
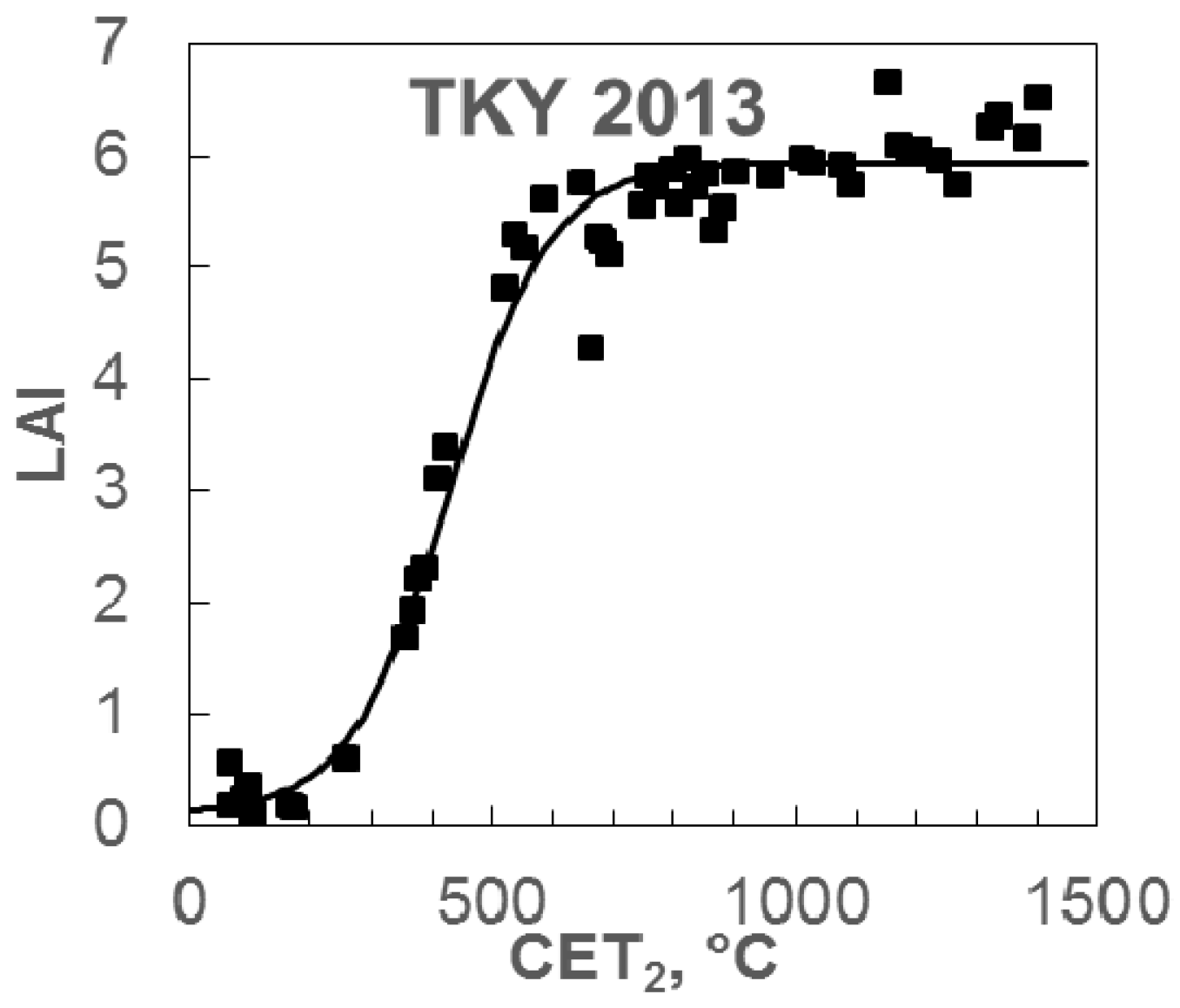
4.2. Accuracy of Leaf Area Index Estimation by the Simple LAI Model
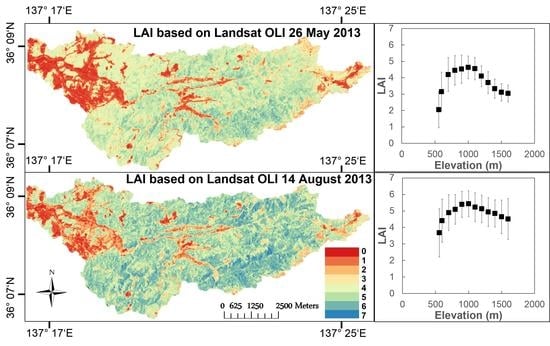
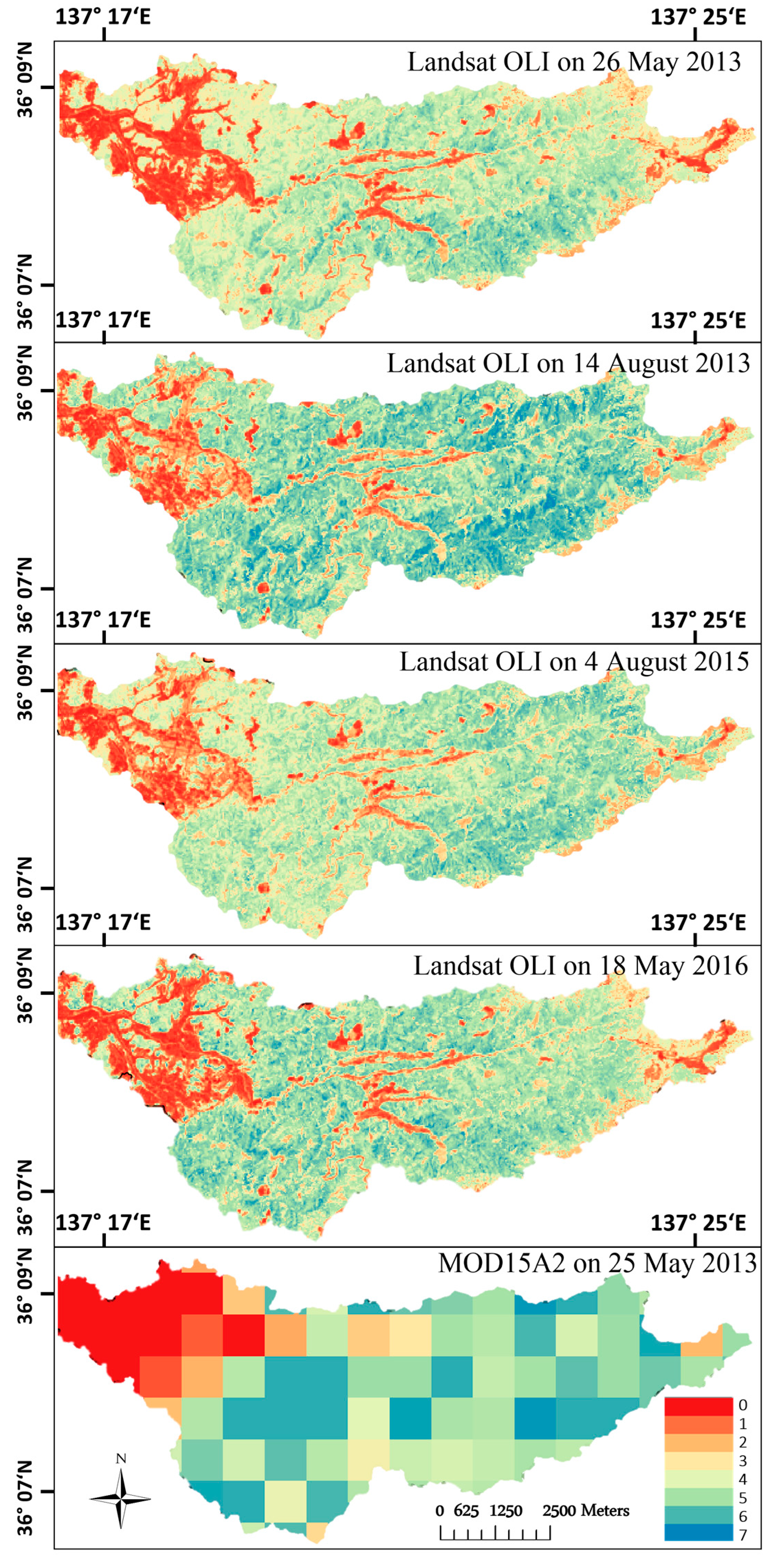
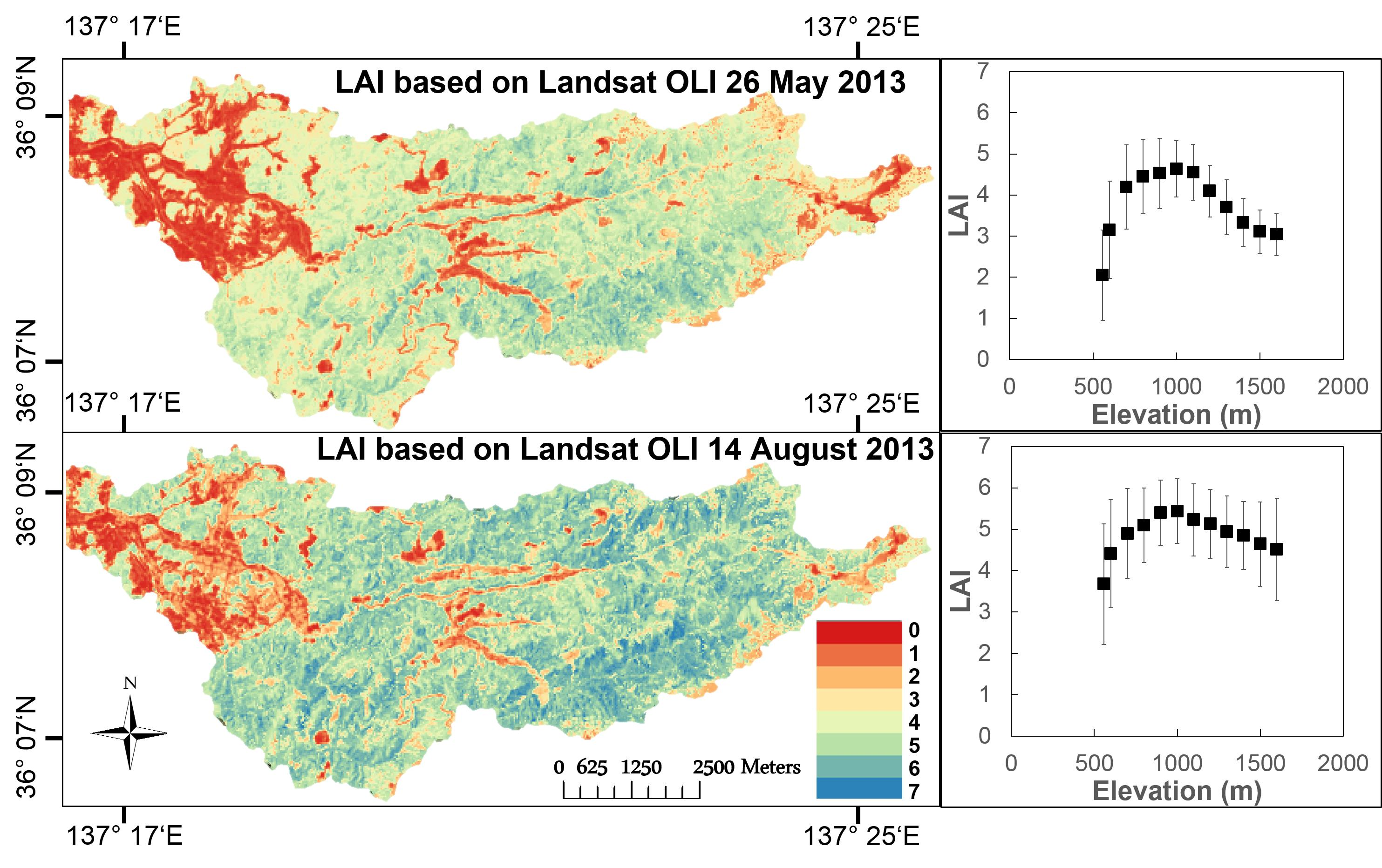
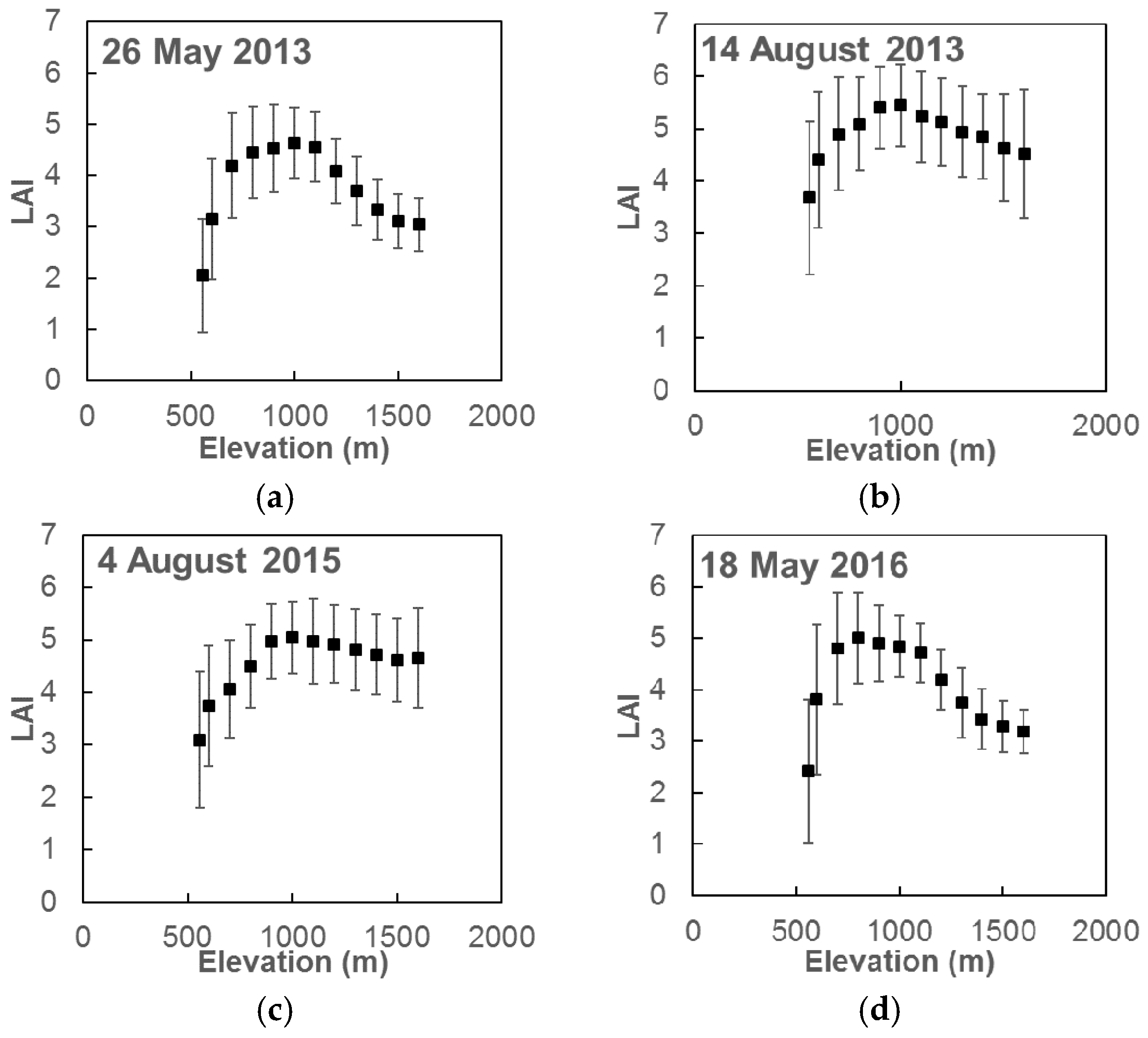
4.3. Spatial Leaf Area Index Variation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
Appendix B

References
- Bonan, G.B. Forests and climate change: forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, H.; Saigusa, N.; Nasahara, K.N.; Noda, H.; Yoshino, J.; Saitoh, T.M.; Nagai, S.; Murayama, S.; Koizumi, H. Effects of seasonal and interannual variations in leaf photosynthesis and canopy leaf area index on gross primary production of a cool-temperate deciduous broadleaf forest in Takayama, Japan. J. Plant Res. 2010, 123, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Muraoka, H.; Noda, H.M.; Nagai, S.; Motohka, T.; Saitoh, T.M.; Nasahara, K.N.; Saigusa, N. Spectral vegetation indices as the indicator of canopy photosynthetic productivity in a deciduous broadleaf forest. J. Plant Ecol. 2012, 6, 393–407. [Google Scholar] [CrossRef]
- Nasahara, K.N.; Muraoka, H.; Nagai, S.; Mikami, H. Vertical integration of leaf area index in a Japanese deciduous broad-leaved forest. Agric. For. Meteorol. 2008, 148, 1136–1146. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, H.; Eklundh, L.; Hall, K.; Lindroth, A. Estimating LAI in deciduous forest stands. Agric. For. Meteorol. 2005, 129, 27–37. [Google Scholar] [CrossRef]
- Biudes, M.S.; Machado, N.G.; Danelichen, V.H.; Souza, M.C.; Vourlitis, G.L.; Nogueira, J.D.S. Ground and remote sensing-based measurements of leaf area index in a transitional forest and seasonal flooded forest in Brazil. Int. J. Biometeorol. 2014, 58, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- Sprintsin, M.; Karnieli, A.; Berliner, P.; Rotenberg, E.; Yakir, D.; Cohen, S. The effect of spatial resolution on the accuracy of leaf area index estimation for a forest planted in the desert transition zone. Remote Sens. Environ. 2007, 109, 416–428. [Google Scholar] [CrossRef]
- Watson, D. Comparative physiological studies on the growth of field crops: II. The effect of varying nutrient supply on net assimilation rate and leaf area. Ann. Bot. 1947, 11, 375–407. [Google Scholar] [CrossRef]
- Chen, J.M.; Black, T. Defining leaf area index for non-flat leaves. Plant Cell Environ. 1992, 15, 421–429. [Google Scholar] [CrossRef]
- Myneni, R.B.; Ramakrishna, R.; Nemani, R.; Running, S.W. Estimation of global leaf area index and absorbed PAR using radiative transfer models. IEEE Trans. Geosci. Remote Sens. 1997, 35, 1380–1393. [Google Scholar] [CrossRef]
- Olivas, P.C.; Oberbauer, S.F.; Clark, D.B.; Clark, D.A.; Ryan, M.G.; O’Brien, J.J.; Ordonez, H. Comparison of direct and indirect methods for assessing leaf area index across a tropical rain forest landscape. Agric. For. Meteorol. 2013, 177, 110–116. [Google Scholar] [CrossRef]
- White, J.D.; Running, S.W.; Nemani, R.; Keane, R.E.; Ryan, K.C. Measurement and remote sensing of LAI in Rocky Mountain montane ecosystems. Can. J. For. Res. 1997, 27, 1714–1727. [Google Scholar] [CrossRef]
- Manninen, T.; Korhonen, L.; Voipio, P.; Lahtinen, P.; Stenberg, P. Leaf Area Index (LAI) Estimation of Boreal Forest Using Wide Optics Airborne Winter Photos. Remote Sens. 2009, 1, 1380–1394. [Google Scholar] [CrossRef]
- Bolstad, P.V.; Vose, J.M.; McNulty, S.G. Forest productivity, leaf area, and terrain in southern Appalachian deciduous forests. For. Sci. 2001, 47, 419–427. [Google Scholar]
- Knyazikhin, Y.; Martonchik, J.; Diner, D.; Myneni, R.; Verstraete, M.; Pinty, B.; Gobron, N. Estimation of vegetation canopy leaf area index and fraction of absorbed photosynthetically active radiation from atmosphere-corrected MISR data. J. Geophys. Res. Atmos. 1998, 103, 32239–32256. [Google Scholar] [CrossRef]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI) 3g and Fraction of Photosynthetically Active Radiation (FPAR) 3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Spanner, M.A.; Pierce, L.L.; Peterson, D.L.; Running, S.W. Remote sensing of temperate coniferous forest leaf area index. The influence of canopy closure, understory vegetation and background reflectance. Int. J. Remote Sens. 1990, 11, 95–111. [Google Scholar] [CrossRef]
- Peterson, D.L.; Spanner, M.A.; Running, S.W.; Teuber, K.B. Relationship of thematic mapper simulator data to leaf area index of temperate coniferous forests. Remote Sens. Environ. 1987, 22, 323–341. [Google Scholar] [CrossRef]
- Nagai, S.; Inoue, T.; Ohtsuka, T.; Kobayashi, H.; Kurumado, K.; Muraoka, H.; Nasahara, K.N. Relationship between spatio-temporal characteristics of leaf-fall phenology and seasonal variations in near surface-and satellite-observed vegetation indices in a cool-temperate deciduous broad-leaved forest in Japan. Int. J. Remote Sens. 2014, 35, 3520–3536. [Google Scholar] [CrossRef]
- Jonckheere, I.; Fleck, S.; Nackaerts, K.; Muys, B.; Coppin, P.; Weiss, M.; Baret, F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agric. For. Meteorol. 2004, 121, 19–35. [Google Scholar] [CrossRef]
- Monsi, M.; Saeki, T. On the factor light in plant communities and its importance for matter production. Ann. Bot. 2005, 95, 549–567. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Li, X.; Wang, J. Advanced Remote Sensing: Terrestrial Information Extraction and Applications, 1st ed.; Academic Press, Elsevier: Oxford, UK, 2012; pp. 347–380. [Google Scholar]
- Propastin, P.; Kappas, M. Retrieval of coarse-resolution leaf area index over the Republic of Kazakhstan using NOAA AVHRR satellite data and ground measurements. Remote Sens. 2012, 4, 220–246. [Google Scholar] [CrossRef]
- Gray, J.; Song, C. Mapping leaf area index using spatial, spectral, and temporal information from multiple sensors. Remote Sens. Environ. 2012, 119, 173–183. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Susaki, J.; Hara, K.; JongGeol, P.; Yasuda, Y.; Kajiwara, K.; Honda, Y. Validation of temporal BRDFs of paddy fields estimated from MODIS reflectance data. IEEE Trans. Geosci. Remote Sens. 2004, 42, 1262–1270. [Google Scholar] [CrossRef]
- Sasai, T.; Saigusa, N.; Nasahara, K.N.; Ito, A.; Hashimoto, H.; Nemani, R.; Hirata, R.; Ichii, K.; Takagi, K.; Saitoh, T.M. Satellite-driven estimation of terrestrial carbon flux over Far East Asia with 1-km grid resolution. Remote Sens. Environ. 2011, 115, 1758–1771. [Google Scholar] [CrossRef]
- Roy, D.P.; Wulder, M.A.; Loveland, T.R.; C.E, W.; Allen, R.G.; Anderson, M.C.; Helder, D.; Irons, J.R.; Johnson, D.M.; Kennedy, R.; et al. Landsat-8: Science and product vision for terrestrial global change research. Remote Sens. Environ. 2014, 145, 154–172. [Google Scholar] [CrossRef]
- Ke, Y.; Im, J.; Lee, J.; Gong, H.; Ryu, Y. Characteristics of Landsat 8 OLI-derived NDVI by comparison with multiple satellite sensors and in-situ observations. Remote Sens. Environ. 2015, 164, 298–313. [Google Scholar] [CrossRef]
- Awaya, Y. Forest type classification over Gujo city using NDVI based on phenology (in Japanese). In Proceedings of the 61st Autumn Conference of the Remote Sensing Society of Japan, Tokyo, Japan, 1–2 November 2016; pp. 251–252. [Google Scholar]
- Awaya, Y.; Takahashi, T. Evaluating the Differences in Modeling Biophysical Attributes between Deciduous Broadleaved and Evergreen Conifer Forests Using Low-Density Small-Footprint LiDAR Data. Remote Sens. 2017, 9, 572. [Google Scholar] [CrossRef]
- Saitoh, T.M.; Tamagawa, I.; Muraoka, H.; Lee, N.-Y.M.; Yashiro, Y.; Koizumi, H. Carbon dioxide exchange in a cool-temperate evergreen coniferous forest over complex topography in Japan during two years with contrasting climates. J. Plant Res. 2010, 123, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, N.; Awaya, Y.; Kojima, T. Classification of forest vegetation types using LiDAR data and Quickbird images—Case study of the Daihachiga river basin in Takayama. J. Jpn. Agric. Syst. Soc. 2012, 28, 115–122. [Google Scholar]
- SATECO. Gifu University 21st Century COE Program “Satellite Ecology”. Available online: http://www.green.gifu-u.ac.jp/sateco/eng/index.html (accessed on 17 January 2018).
- Ohtsuka, T.; Shizu, Y.; Nishiwaki, A.; Yashiro, Y.; Koizumi, H. Carbon cycling and net ecosystem production at an early stage of secondary succession in an abandoned coppice forest. J. Plant Res. 2010, 123, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, T.; Akiyama, T.; Hashimoto, Y.; Inatomi, M.; Sakai, T.; Jia, S.; Mo, W.; Tsuda, S.; Koizumi, H. Biometric based estimates of net primary production (NPP) in a cool-temperate deciduous forest stand beneath a flux tower. Agric. For. Meteorol. 2005, 134, 27–38. [Google Scholar] [CrossRef]
- Inoue, T.; Nagai, S.; Inoue, S.; Ozaki, M.; Sakai, S.; Muraoka, H.; Koizumi, H. Seasonal variability of soil respiration in multiple ecosystems under the same physical–geographical environmental conditions in central Japan. For. Sci. Technol. 2012, 8, 52–60. [Google Scholar] [CrossRef]
- Noda, H.M.; Muraoka, H.; Nasahara, K.N.; Saigusa, N.; Murayama, S.; Koizumi, H. Phenology of leaf morphological, photosynthetic, and nitrogen use characteristics of canopy trees in a cool-temperate deciduous broadleaf forest at Takayama, central Japan. Ecol. Res. 2015, 30, 247–266. [Google Scholar] [CrossRef]
- Saitoh, T.M.; Nagai, S.; Yoshino, J.; Kondo, H.; Tamagawa, I.; Muraoka, H. Effects of canopy phenology on deciduous overstory and evergreen understory carbon budgets in a cool-temperate forest ecosystem under ongoing climate change. Ecol. Res. 2015, 30, 267–277. [Google Scholar] [CrossRef]
- Kira, T. Forest ecosystems of east and southeast Asia in a global perspective. Ecol. Res. 1991, 6, 185–200. [Google Scholar] [CrossRef]
- Chavez, P.S. An improved dark-object subtraction technique for atmospheric scattering correction of multispectral data. Remote Sens. Environ. 1988, 24, 459–479. [Google Scholar] [CrossRef]
- USGS. Using the USGS Landsat 8 Product. Available online: https://landsat.usgs.gov/using-usgs-landsat-8-product (accessed on 17 January 2018).
- Liang, S.; Fang, H.; Chen, M. Atmospheric Correction of Landsat ETM+ Land Surface Imagery. Part I: Methods. IEEE Trans. Geosci. Remote Sens. 2001, 39, 2490–2498. [Google Scholar] [CrossRef]
- Williams, D.L. A comparison of spectral reflectance properties at the needle, branch, and canopy level for selected conifer species. Remote Sens. Environ. 1991, 35, 79–93. [Google Scholar] [CrossRef]
- Kaufman, Y.J. The atmospheric effect on remote sensing and its correction. In Theory and Applications of Optical Remote Sensing; Asrar, G., Ed.; Wiley: New York, NY, USA, 1989; pp. 336–428. [Google Scholar]
- Ge, H.; Lu, D.; He, S.; Xu, A.; Zhou, G.; Du, H. Pixel-based Minnaert correction method for reducing topographic effects on a Landsat 7 ETM+ image. Photogramm. Eng. Remote Sens. 2008, 74, 1343–1350. [Google Scholar] [CrossRef]
- Smith, J.; Lin, T.L.; Ranson, K. The Lambertian assumption and Landsat data. Photogramm. Eng. Remote Sens. 1980, 46, 1183–1189. [Google Scholar]
- Tokola, T.; Sarkeala, J.; Van der Linden, M. Use of topographic correction in Landsat TM-based forest interpretation in Nepal. Int. J. Remote Sens. 2001, 22, 551–563. [Google Scholar] [CrossRef]
- Awaya, T.; Kodani, E.; Iehara, T.; Hosoda, K.; Takahashi, T.; Sakai, T. Mapping leaf area index over Japanese Archipelago—Comparison of the results by plot data and MODIS data. In Proceedings of the 16th Seiken forum on earth environmental monitoring, University of Tokyo, Tokyo, Japan, 26–27 March 2007; pp. 88–91. (In Japanese). [Google Scholar]
- Chen, J.M.; Rich, P.M.; Gower, S.T.; Norman, J.M.; Plummer, S. Leaf area index of boreal forests: Theory, techniques, and measurements. J. Geophys. Res. Atmos. 1997, 102, 29429–29443. [Google Scholar] [CrossRef]
- Ross, J.; Sulev, M. Sources of errors in measurements of PAR. Agric. For. Meteorol. 2000, 100, 103–125. [Google Scholar] [CrossRef]
- Ridao, E.; Conde, J.R.; Mínguez, M.I. Estimating ƒAPAR from nine vegetation indices for irrigated and nonirrigated faba bean and semileafless pea canopies. Remote Sens. Environ. 1998, 66, 87–100. [Google Scholar] [CrossRef]
- Inoue, Y.; Olioso, A. Methods of Estimating Plant Productivity and CO 2 Flux in Agro-Ecosystems–Liking Measurements, Process Models, and Remotely Sensed Information. Elsevier Oceanogr. Ser. 2007, 73, 295–502. [Google Scholar]
- Jenkins, J.; Richardson, A.D.; Braswell, B.; Ollinger, S.V.; Hollinger, D.Y.; Smith, M.-L. Refining light-use efficiency calculations for a deciduous forest canopy using simultaneous tower-based carbon flux and radiometric measurements. Agric. For. Meteorol. 2007, 143, 64–79. [Google Scholar] [CrossRef]
- Saitoh, T.M.; Nagai, S.; Noda, H.M.; Muraoka, H.; Nasahara, K.N. Examination of the extinction coefficient in the Beer–Lambert law for an accurate estimation of the forest canopy leaf area index. For. Sci. Technol. 2012, 8, 67–76. [Google Scholar] [CrossRef]
- Hirata, R.; Hirano, T.; Saigusa, N.; Fujinuma, Y.; Inukai, K.; Kitamori, Y.; Takahashi, Y.; Yamamoto, S. Seasonal and interannual variations in carbon dioxide exchange of a temperate larch forest. Agric. For. Meteorol. 2007, 147, 110–124. [Google Scholar] [CrossRef]
- Koike, T.; Mao, Q.; Inada, N.; Kawaguchi, K.; Hoshika, Y.; Kita, K.; Watanabe, M. Growth and photosynthetic responses of cuttings of a hybrid larch (Larix gmelinii var. japonica x L. kaempferi) to elevated ozone and/or carbon dioxide. Asian J. Atmos. Environ. 2012, 6, 104–110. [Google Scholar] [CrossRef]
- Teillet, P.; Fedosejevs, G. On the dark target approach to atmospheric correction of remotely sensed data. Can. J. Remote Sens. 1995, 21, 374–387. [Google Scholar] [CrossRef]
- Kaufman, Y.J.; Sendra, C. Algorithm for automatic atmospheric corrections to visible and near-IR satellite imagery. Int. J. Remote Sens. 1988, 9, 1357–1381. [Google Scholar] [CrossRef]
- Chen, J.M.; Cihlar, J. Retrieving leaf area index of boreal conifer forests using Landsat TM images. Remote Sens. Environ. 1996, 55, 153–162. [Google Scholar] [CrossRef]
- Staelens, J.; Nachtergale, L.; Luyssaert, S.; Lust, N. A model of wind-influenced leaf litterfall in a mixed hardwood forest. Can. J. For. Res. 2003, 33, 201–209. [Google Scholar] [CrossRef]
- Smith, F.W.; Sampson, D.A.; Long, J.N. Notes: Comparison of leaf area index estimates from tree allometrics and measured light interception. For. Sci. 1991, 37, 1682–1688. [Google Scholar]
- Baldocchi, D.D.; Matt, D.R.; Hutchison, B.A.; McMillen, R.T. Solar radiation within an oak—Hickory forest: An evaluation of the extinction coefficients for several radiation components during fully-leafed and leafless periods. Agric. For. Meteorol. 1984, 32, 307–322. [Google Scholar] [CrossRef]
- Lee, B.; Kwon, H.; Miyata, A.; Lindner, S.; Tenhunen, J. Evaluation of a Phenology-Dependent Response Method for Estimating Leaf Area Index of Rice Across Climate Gradients. Remote Sens. 2017, 9, 20. [Google Scholar] [CrossRef]
- Li, C.; Song, J.; Wang, J. Modifying Geometric-Optical Bidirectional Reflectance Model for Direct Inversion of Forest Canopy Leaf Area Index. Remote Sens. 2015, 7, 11083–11104. [Google Scholar] [CrossRef]
- Setoyama, Y.; Sasai, T. Analyzing decadal net ecosystem production control factors and the effects of recent climate events in Japan. J. Geophys. Res. Biogeosci. 2013, 118, 337–351. [Google Scholar] [CrossRef]
- Parton, W.; Scurlock, J.; Ojima, D.; Gilmanov, T.; Scholes, R.; Schimel, D.; Kirchner, T.; Menaut, J.C.; Seastedt, T.; Garcia Moya, E. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide. Glob. Biogeochem. Cycles 1993, 7, 785–809. [Google Scholar] [CrossRef]
- Watanabe, H.; Mogi, Y. Soil carbon stocks in mature Sugi and Hinoki stands. Bull. Gifu Prefect. Res. Inst. For. 2005, 34, 11–18. (In Japanese) [Google Scholar]
- Schulze, E.-D.; Schulze, W.; Koch, H.; Arneth, A.; Bauer, G.; Kelliher, F.; Hollinger, D.; Vygodskaya, N.; Kusnetsova, W.; Sogatchev, A. Aboveground biomass and nitrogen nutrition in a chronosequence of pristine Dahurian Larix stands in eastern Siberia. Can. J. For. Res. 1995, 25, 943–960. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot Code | Location | Altitude (m a.s.l.) | Forest Type | Dominant Species | Forest Age (years) | Plot Size (m2) | Annual Mean Air Temperature (°C) | Annual Precipitation (mm) | References |
|---|---|---|---|---|---|---|---|---|---|
| TKY | 36°08′46″N, 137°25′23″E | 1420 | DBF | Quercus crispula, Betula ermanii, Betula platyphylla var. japonica | 50–60 | 100 × 100 | 6.6 | 2400 | [36] |
| TKC | 36°08′24″N, 137°22′15″E | 800 | ECF | Cryptomeria japonica | 40–50 | 30 × 50 | 11.2 | 1723 | [3,32] |
| C18 | 36°08′22″N, 137°23′17″E | 1160 | DBF | Betula platyphylla var. japonica, Castanea crenata, Quercus crispula | 30 | 20 × 30 | 7.2 | NA* | [35] |
| L50 | 36°08′04″N, 137°25′41″E | 1330 | DCF | Larix kaempferi | 50–60 | 30 × 30 | 7.2 | 2075 | [37] |
| Image Date | Sun Elevation Angle (°) | Cloud Coverage (%) |
|---|---|---|
| 26 May 2013 | 67.36 | 6.49 |
| 14 August 2013 | 61.07 | 1.82 |
| 4 August 2015 | 62.68 | 9.90 |
| 18 May 2016 | 65.87 | 0.35 |
| Image Date | OLI Band | Regression * | R2 |
|---|---|---|---|
| 26 May 2013 | Blue | y = −0.61x + 9385.34 | 0.91 |
| Green | y = −0.56x + 8577.35 | 0.88 | |
| Red | y = −0.48x + 7400.11 | 0.97 | |
| 14 August 2013 | Blue | y = −0.55x + 8811.87 | 0.97 |
| Green | y = −0.48x + 7605.49 | 0.98 | |
| Red | y = −0.39x + 6674.14 | 0.82 | |
| 4 August 2015 | Blue | y = −0.50x + 8969.58 | 0.98 |
| Green | y = −0.65x + 8011.62 | 0.96 | |
| Red | y = −0.36x + 6787.98 | 0.94 | |
| 18 May 2016 | Blue | y = −0.42x + 8431.08 | 0.87 |
| Green | y = −0.49x + 7456.11 | 0.97 | |
| Red | y = −0.34x + 6572.35 | 0.84 |
| Image Date | Image Reflectance of Dark ECF Pixels | Offset RF | ||||
|---|---|---|---|---|---|---|
| Blue (%) | Green (%) | Red (%) | Blue (%) | Green (%) | Red (%) | |
| 26 May 2013 | 0.6 ± 0.2 * | 1.1 ± 0.1 | 0.9 ± 0.1 | 1.3 | 2.8 | 1.0 |
| 14 August 2013 | 0.6 ± 0.0 | 1.0 ± 0.1 | 0.8 ± 0.1 | 1.3 | 2.9 | 1.1 |
| 4 August 2015 | 0.5 ± 0.1 | 0.6 ± 0.1 | 0.4 ± 0.2 | 1.4 | 2.3 | 1.5 |
| 18 May 2016 | 1.4 ± 0.1 | 1.3 ± 0.1 | 1.4 ± 0.1 | 0.5 | 2.6 | 0.5 |
| Norway spruce [44] | 1.9 ± 0.2 | 3.9 ± 0.4 | 1.9 ± 0.2 | - | - | - |
| Plot Code | Estimation Method | Maximum LAI | Spring LAI * |
|---|---|---|---|
| TKY | PAR transmittance | 5.9 ± 0.4 | 1.7 |
| TKY | Litter-fall collection | 5.0 ± 0.5 | - |
| TKC | Allometry | 7.3 | 7.3 |
| C18 | Litter-fall collection | 5.2 | 5.1 |
| L50 | Allometry | 7.6 | 7.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnikova, I.; Awaya, Y.; Saitoh, T.M.; Muraoka, H.; Sasai, T. Estimation of Leaf Area Index in a Mountain Forest of Central Japan with a 30-m Spatial Resolution Based on Landsat Operational Land Imager Imagery: An Application of a Simple Model for Seasonal Monitoring. Remote Sens. 2018, 10, 179. https://doi.org/10.3390/rs10020179
Melnikova I, Awaya Y, Saitoh TM, Muraoka H, Sasai T. Estimation of Leaf Area Index in a Mountain Forest of Central Japan with a 30-m Spatial Resolution Based on Landsat Operational Land Imager Imagery: An Application of a Simple Model for Seasonal Monitoring. Remote Sensing. 2018; 10(2):179. https://doi.org/10.3390/rs10020179
Chicago/Turabian StyleMelnikova, Irina, Yoshio Awaya, Taku M. Saitoh, Hiroyuki Muraoka, and Takahiro Sasai. 2018. "Estimation of Leaf Area Index in a Mountain Forest of Central Japan with a 30-m Spatial Resolution Based on Landsat Operational Land Imager Imagery: An Application of a Simple Model for Seasonal Monitoring" Remote Sensing 10, no. 2: 179. https://doi.org/10.3390/rs10020179