Topography and Three-Dimensional Structure Can Estimate Tree Diversity along a Tropical Elevational Gradient in Costa Rica

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
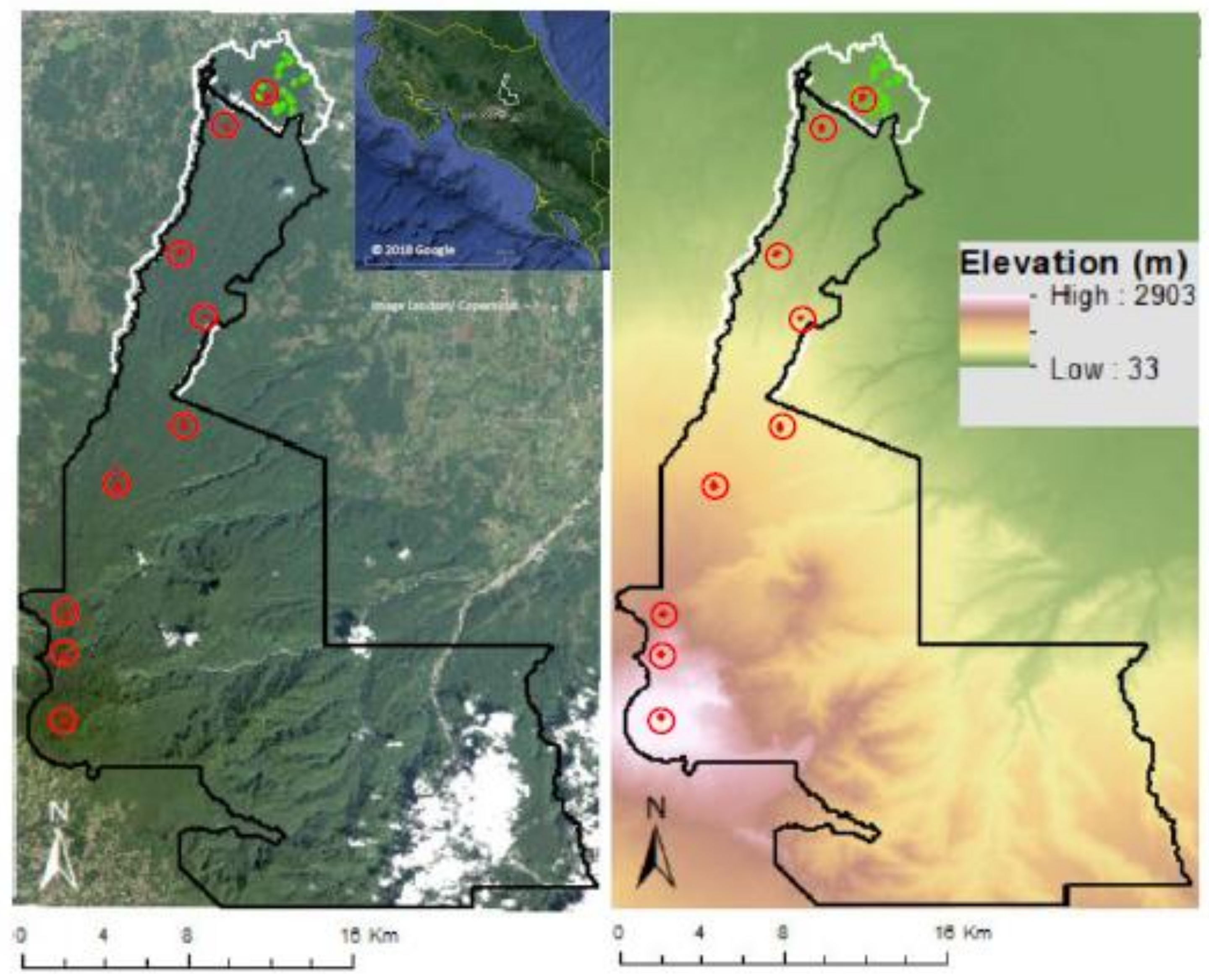
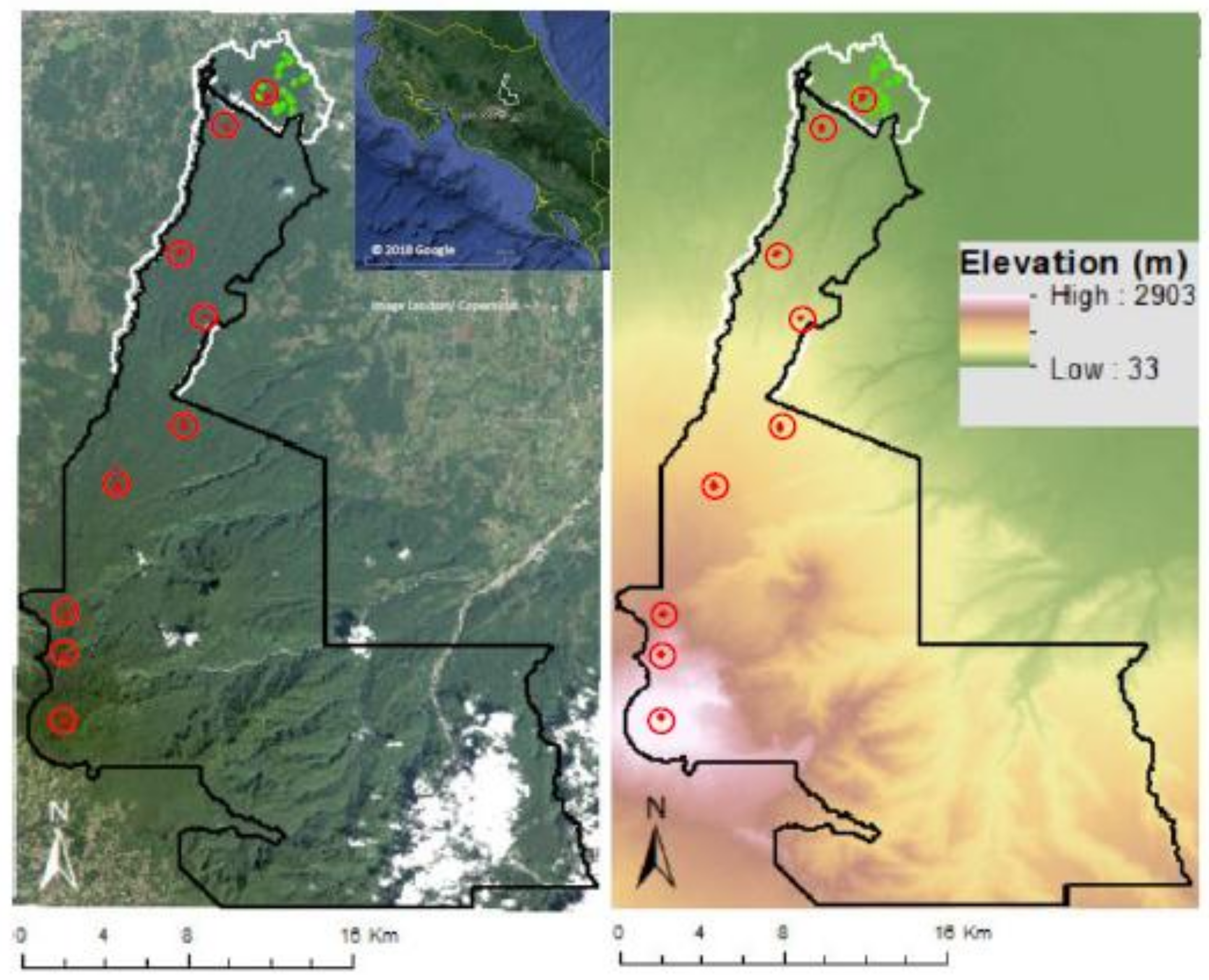
2.1. Study Area
2.2. Field Data
2.3. Lidar Remote Sensing
2.4. Data Analysis
3. Results
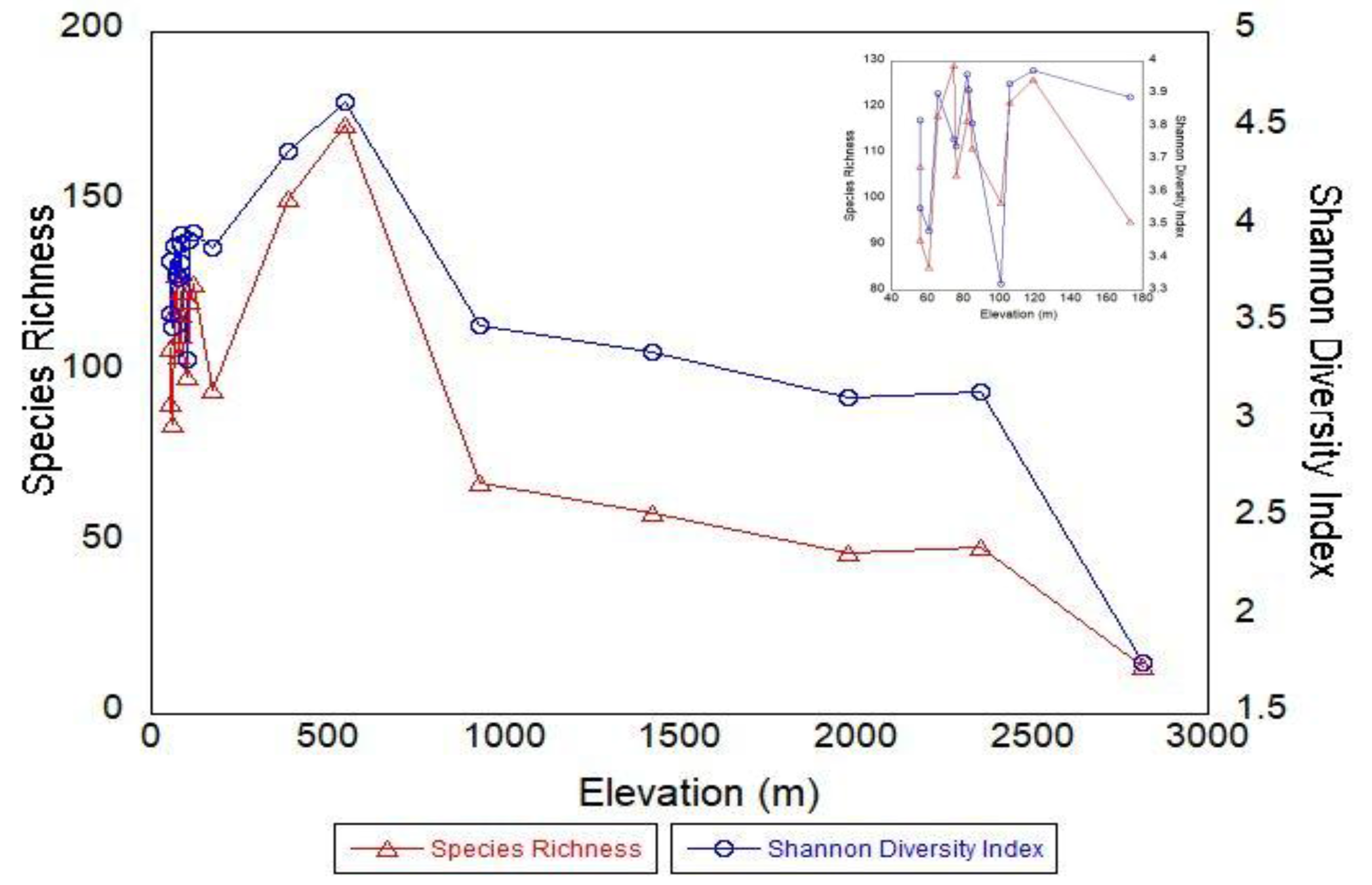
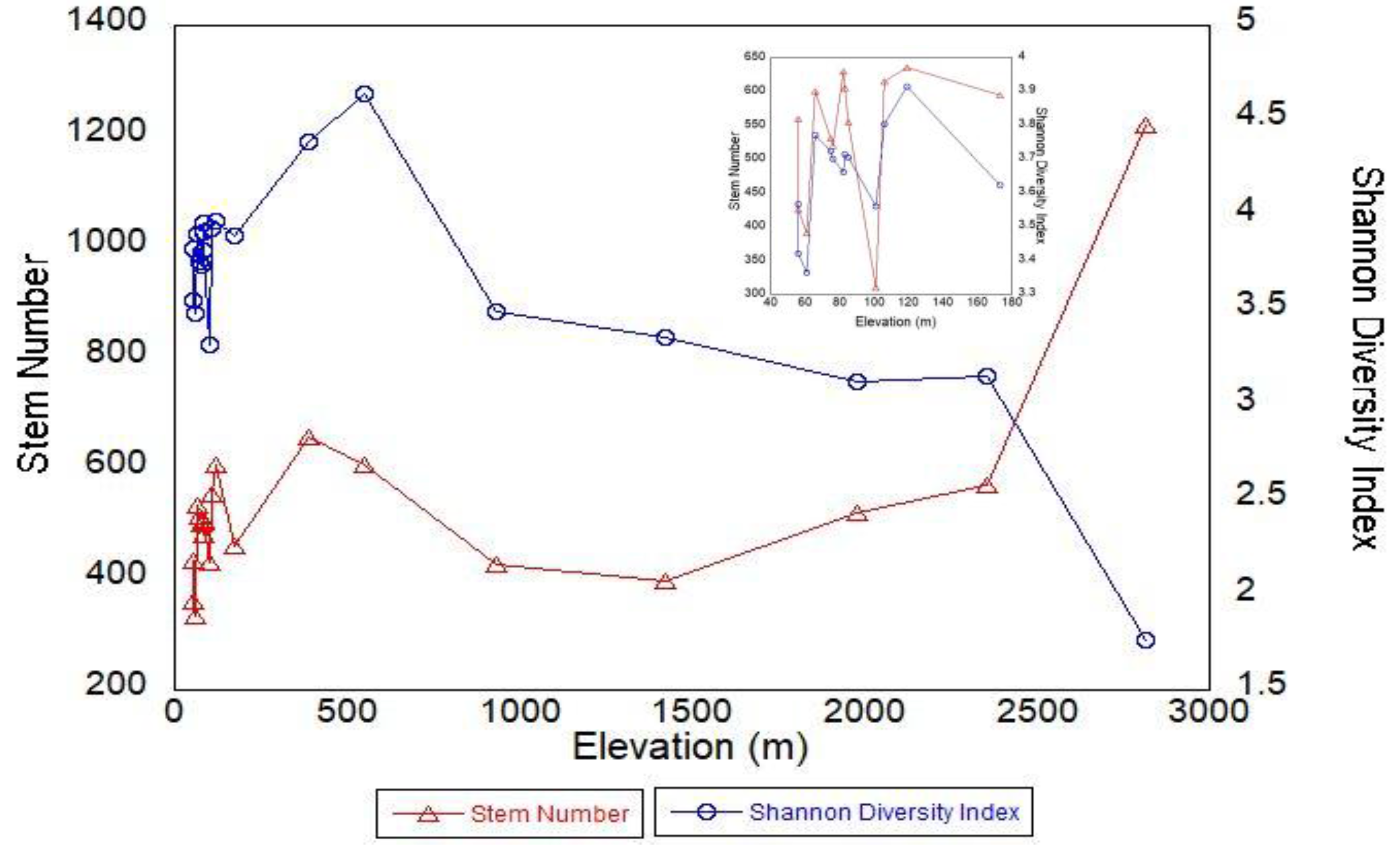
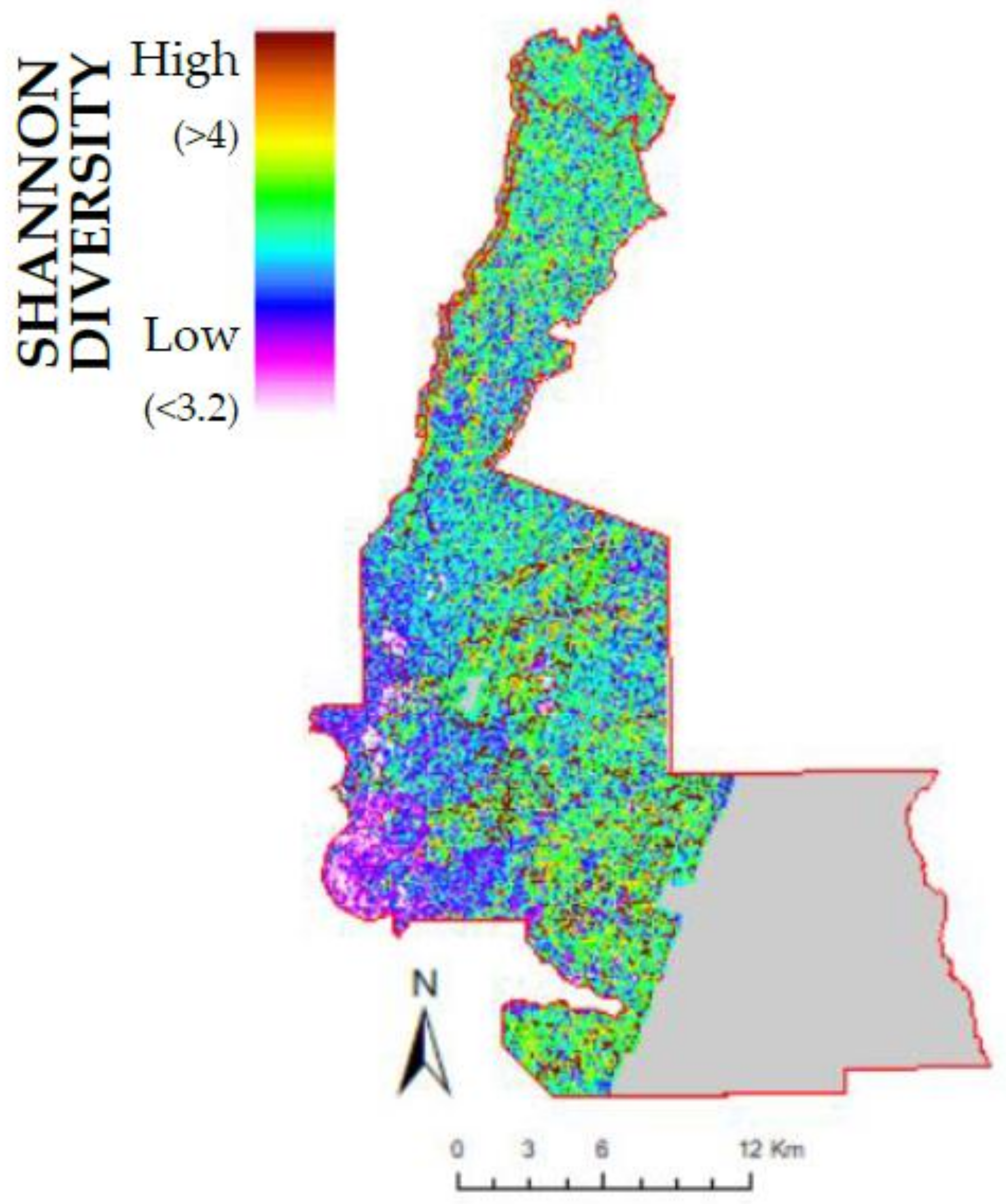
3.1. Species Richness, Diversity, and Forest Structure from Plots
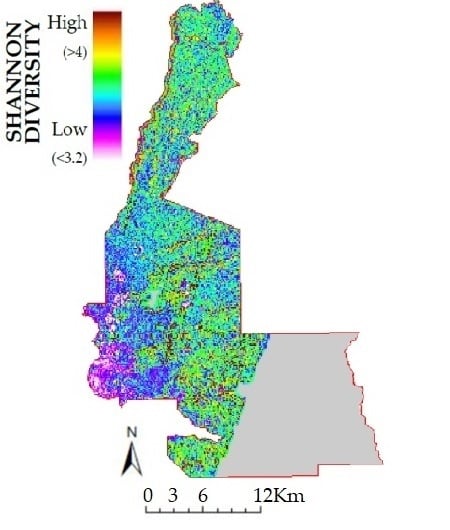
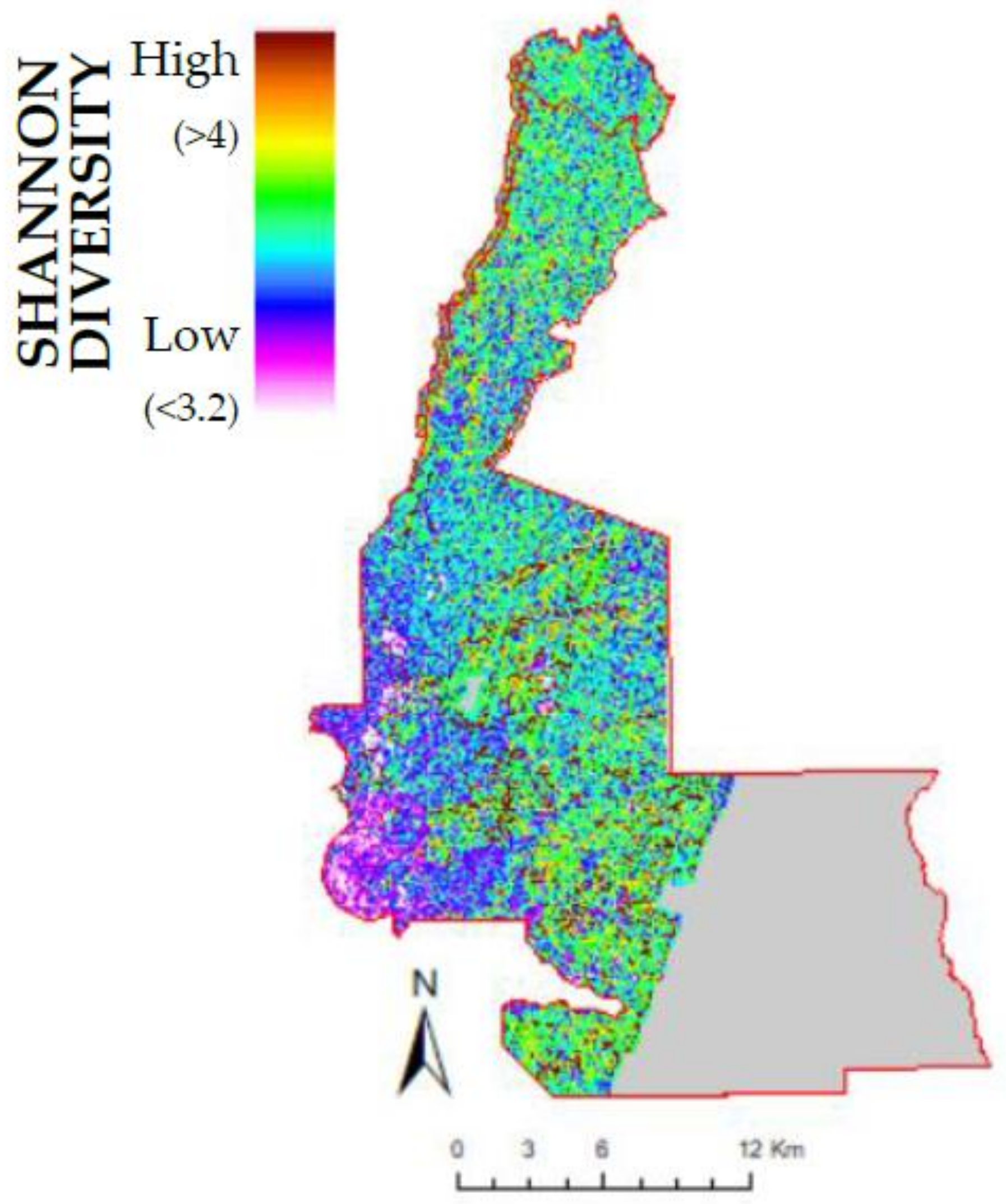
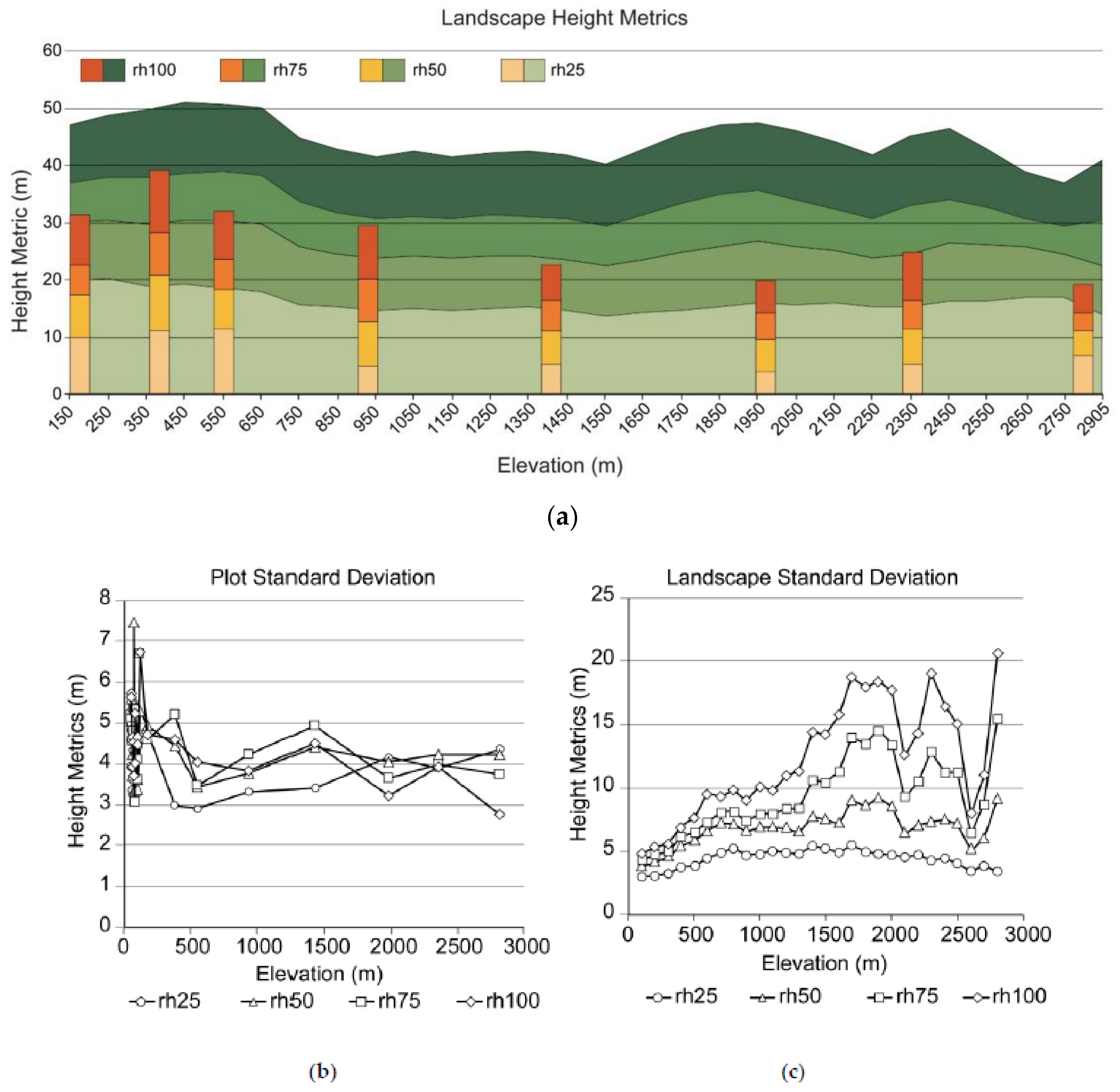
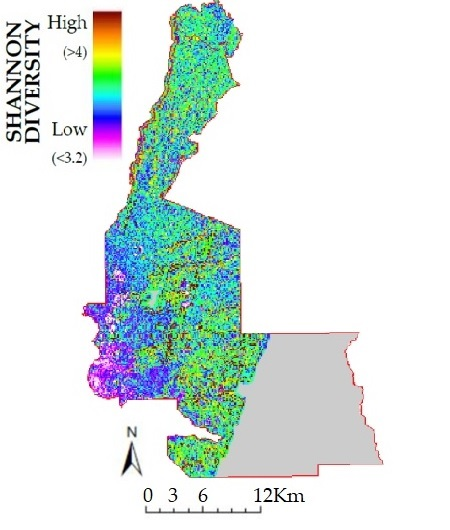
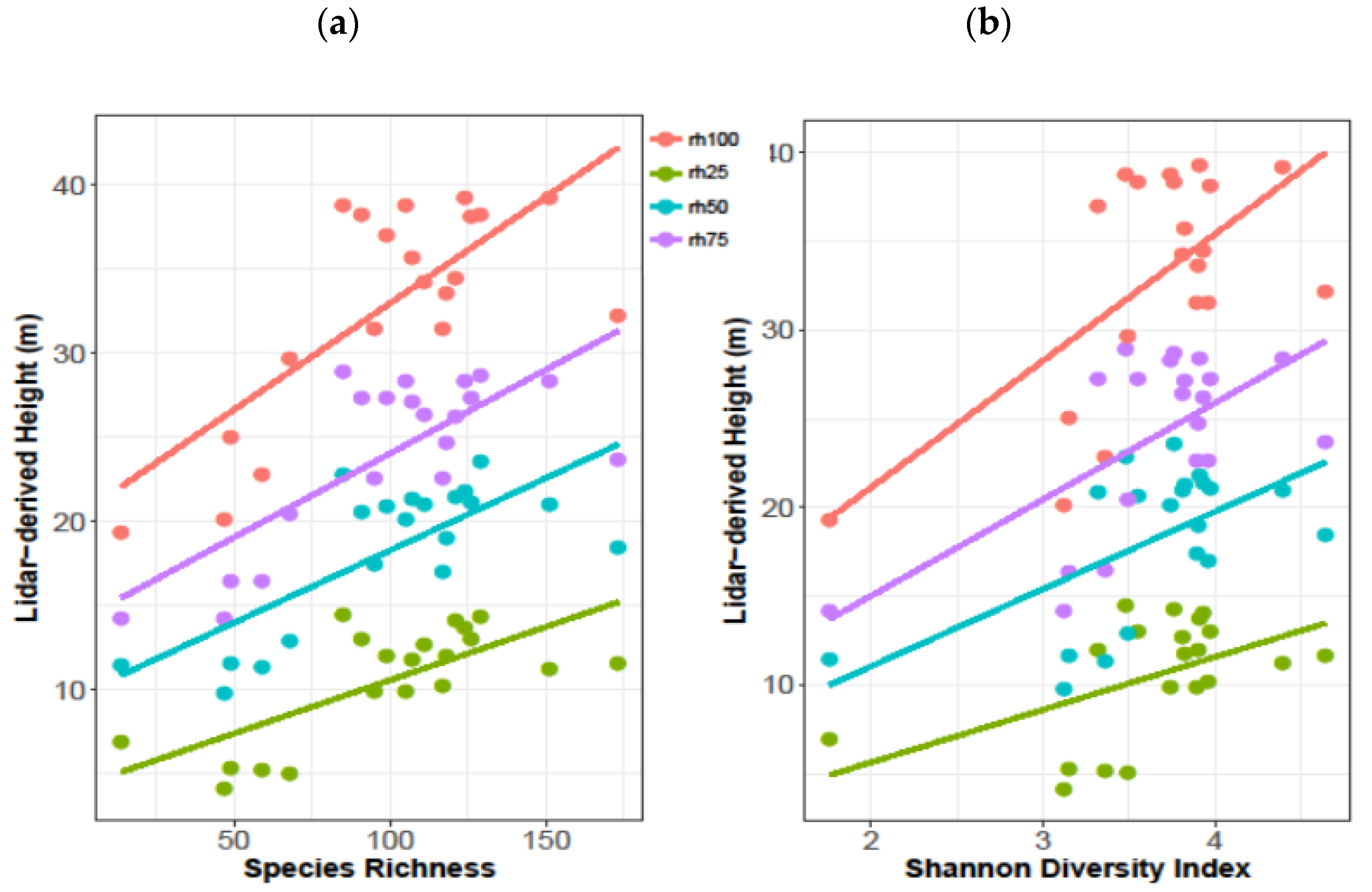
3.2. Lidar Metrics and Species Richness and Diversity
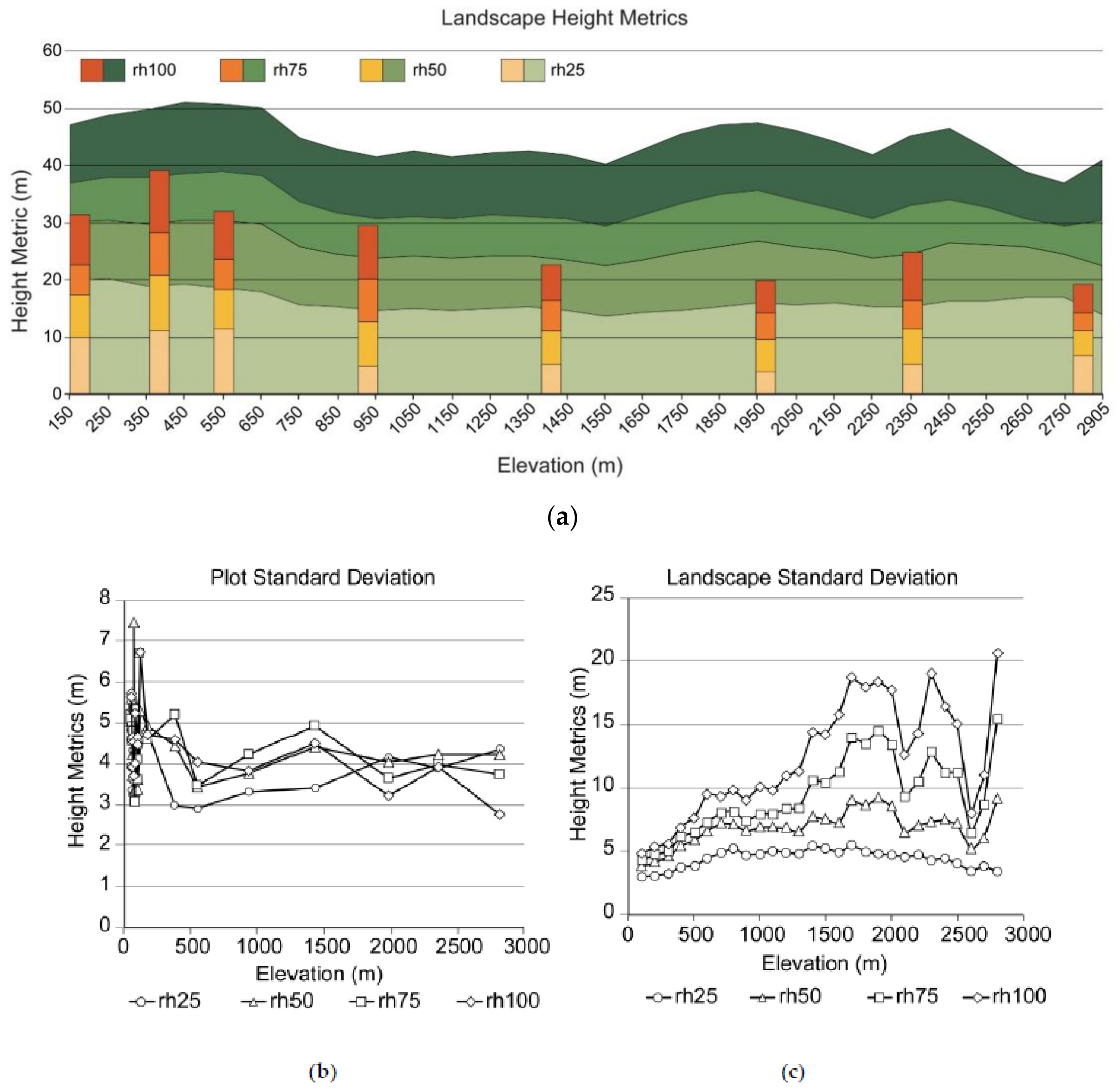
3.3. Plot and Landscape Data
4. Discussion
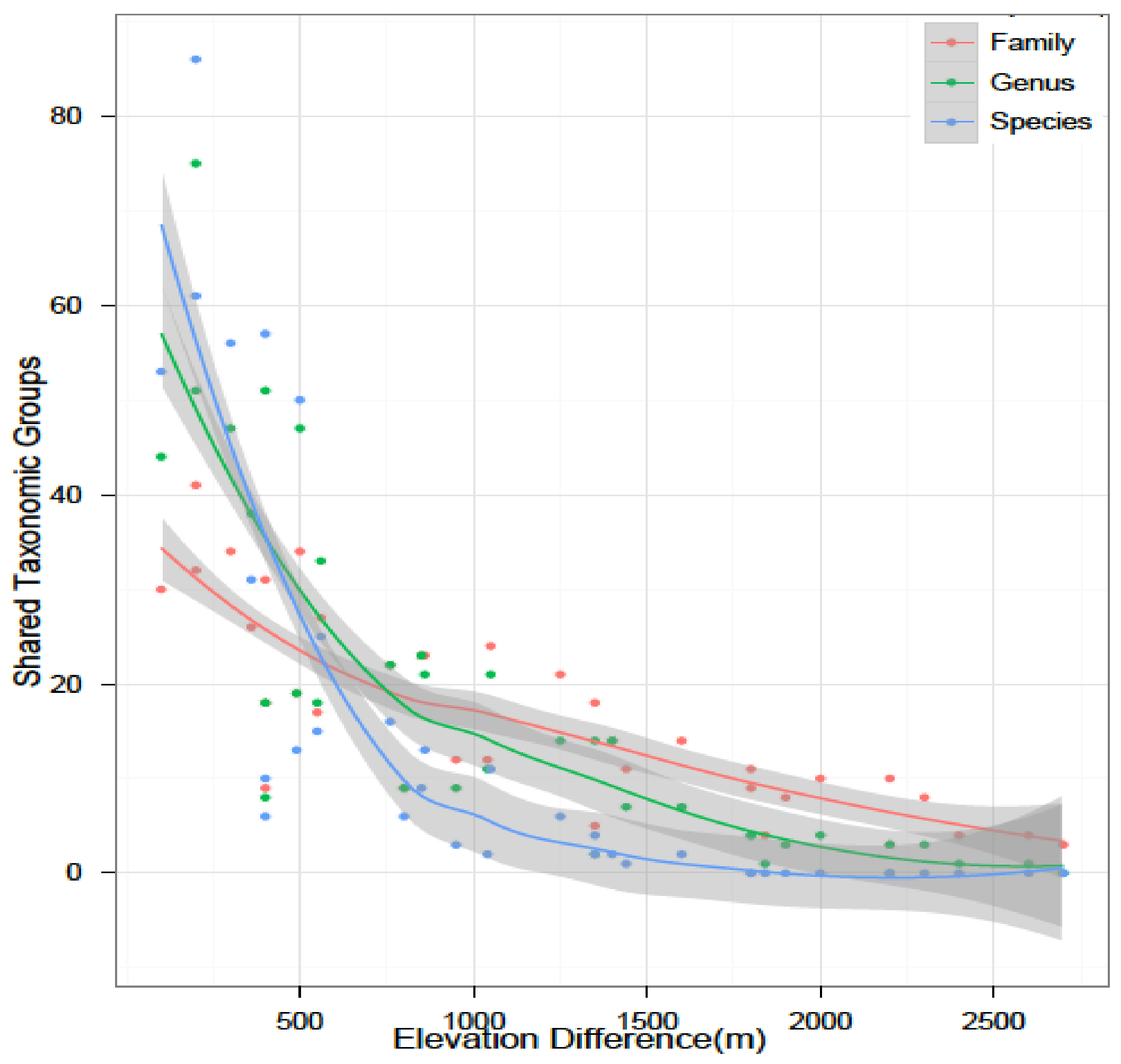
4.1. Species Richness and Diversity
4.2. Forest Structure from Field Measurements
4.3. Topography and Three Dimensional Structure from Lidar
4.4. Plots vs. Landscape
4.5. Modeling Diversity and Species Richness
4.6. Future Research
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chapin, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Jantz, S.M.; Barker, B.; Brooks, T.M.; Chini, L.P.; Huang, Q.; Moore, R.M.; Noel, J.; Hurtt, G.C. Future habitat loss and extinctions driven by land-use change in biodiversity hotspots under four scenarios of climate-change mitigation. Conserv. Biol. 2015, 29, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Olivares, I.; Svenning, J.-C.; van Bodegom, P.M.; Balslev, H. Effects of warming and drought on the vegetation and plant diversity in the Amazon basin. Bot. Rev. 2015, 81, 42–69. [Google Scholar] [CrossRef]
- Rose, R.A.; Byler, D.; Eastman, J.R.; Fleishman, E.; Geller, G.; Goetz, S.; Guild, L.; Hamilton, H.; Hansen, M.; Headley, R.; et al. Ten ways remote sensing can contribute to conservation. Conserv. Biol. 2015, 29, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina Mountains, Arizona. V. Biomass, production, and diversity along the elevation gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef]
- Lieberman, D.; Lieberman, M.; Peralta, R.; Hartshorn, G.S. Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. J. Ecol. 1996, 84, 137–152. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef]
- Guo, Q.; Kelt, D.A.; Sun, Z.; Liu, H.; Hu, L.; Ren, H.; Wen, J. Global variation in elevational diversity patterns. Sci. Rep. 2013, 3, 3007. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H. Geographical Ecology; Patterns in the Distribution of Species; Harper & Row: New York, NY, USA, 1972; p. 269. [Google Scholar]
- Rohde, K. Latitudinal gradients in species-diversity—The search for the primary cause. Oikos 1992, 65, 514–527. [Google Scholar] [CrossRef]
- Rahbek, C. The elevational gradient of species richness - a uniform pattern. Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Nogues-Bravo, D.; Araujo, M.B.; Romdal, T.; Rahbek, C. Scale effects and human impact on the elevational species richness gradients. Nature 2008, 453, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Gentry, A.H. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann. Mo. Bot. Gard. 1988, 75, 1–34. [Google Scholar] [CrossRef]
- Vazquez, J.A.; Givnish, T.J. Altitudinal gradients in tropical forest composition, structure, and diversity in the sierra de manantlan. J. Ecol. 1998, 86, 999–1020. [Google Scholar]
- Wolf, J.H.D.; Alejandro, F. Patterns in species richness and distribution of vascular epiphytes in chiapas, mexico. J. Biogeogr. 2003, 30, 1689–1707. [Google Scholar] [CrossRef]
- McCain, C.M. The mid-domain effect applied to elevational gradients: Species richness of small mammals in costa rica. J. Biogeogr. 2004, 31, 19–31. [Google Scholar] [CrossRef]
- Carpenter, C. The environmental control of plant species density on a Himalayan elevation gradient. J. Biogeogr. 2005, 32, 999–1018. [Google Scholar] [CrossRef]
- Bhattarai, K.R.; Vetaas, O.R. Can rapoport’s rule explain tree species richness along the Himalayan elevation gradient, Nepal? Divers. Distrib. 2006, 12, 373–378. [Google Scholar] [CrossRef]
- Cardelus, C.L.; Colwell, R.K.; Watkins, J.E. Vascular epiphyte distribution patterns: Explaining the mid-elevation richness peak. J. Biogeogr. 2006, 94, 144–156. [Google Scholar] [CrossRef]
- Acharya, B.K.; Chettri, B.; Vijayan, L. Distribution pattern of trees along an elevation gradient of Eastern Himalaya, India. Acta Oecol. Int. J. Ecol. 2011, 37, 329–336. [Google Scholar] [CrossRef]
- Aynekulu, E.; Aerts, R.; Moonen, P.; Denich, M.; Gebrehiwot, K.; Vagen, T.-G.; Mekuria, W.; Boehmer, H.J. Altitudinal variation and conservation priorities of vegetation along the great rift valley escarpment, Northern Ethiopia. Biodivers. Conserv. 2012, 21, 2691–2707. [Google Scholar] [CrossRef]
- Feeley, K.J.; Hurtado, J.; Saatchi, S.; Silman, M.R.; Clark, D.B. Compositional shifts in Costa Rican forests due to climate-driven species migrations. Glob. Chang. Biol. 2013, 19, 3472–3480. [Google Scholar] [CrossRef] [PubMed]
- Kuper, W.; Kreft, H.; Nieder, J.; Koster, N.; Barthlott, W. Large-scale diversity patterns of vascular epiphytes in neotropical montane rain forests. J. Biogeogr. 2004, 31, 1477–1487. [Google Scholar] [CrossRef]
- Homeier, J.; Breckle, S.-W.; Guenter, S.; Rollenbeck, R.T.; Leuschner, C. Tree diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich ecuadorian montane rain forest. Biotropica 2010, 42, 140–148. [Google Scholar] [CrossRef]
- Lefsky, M.A.; Cohen, W.B.; Parker, G.G.; Harding, D.J. Lidar remote sensing for ecosystem studies: Lidar, an emerging remote sensing technology that directly measures the three-dimensional distribution of plant canopies, can accurately estimate vegetation structural attributes and should be of particular interest to forest, landscape, and global ecologists. BioScience 2002, 52, 19–30. [Google Scholar]
- Hofton, M.A.; Rocchio, L.E.; Blair, J.B.; Dubayah, R. Validation of vegetation canopy lidar sub-canopy topography measurements for a dense tropical forest. J. Geodyn. 2002, 34, 491–502. [Google Scholar] [CrossRef]
- Blair, J.B.; Hofton, M.A. Modeling laser altimeter return waveforms over complex vegetation using high-resolution elevation data. Geophys. Res. Lett. 1999, 26, 2509–2512. [Google Scholar] [CrossRef]
- Clark, D.B.; Clark, D.A. Landscape-scale variation in forest structure and biomass in a tropical rain forest. For. Ecol. Manag. 2000, 137, 185–198. [Google Scholar] [CrossRef]
- Girardin, C.A.J.; Farfan-Rios, W.; Garcia, K.; Feeley, K.J.; Jorgensen, P.M.; Araujo Murakami, A.; Perez, L.C.; Seidel, R.; Paniagua, N.; Claros, A.F.F.; et al. Spatial patterns of above-ground structure, biomass and composition in a network of six andean elevation transects. Plant Ecol. Divers. 2014, 7, 161–171. [Google Scholar] [CrossRef]
- Condit, R.; Hubbell, S.P.; Lafrankie, J.V.; Sukumar, R.; Manokaran, N.; Foster, R.B.; Ashton, P.S. Species-area and species-individual relationships for tropical trees: A comparison of three 50-ha plots. J. Ecol. 1996, 84, 549–562. [Google Scholar] [CrossRef]
- Givnish, T.J. On the causes of gradients in tropical tree diversity. J. Ecol. 1999, 87, 193–210. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Quantifying biodiversity: Procedures and pitfalls in the measurement and comparison of species richness. Ecol. Lett. 2001, 4, 379–391. [Google Scholar] [CrossRef]
- Giriraj, A.; Murthy, M.S.R.; Ramesh, B.R. Vegetation composition, structure and patterns of diversity: A case study from the tropical wet evergreen forests of the western ghats, india. Edinburgh J. Bot. 2008, 65, 447–468. [Google Scholar] [CrossRef]
- Unger, M.; Homeier, J.; Leuschner, C. Relationships among leaf area index, below-canopy light availability and tree diversity along a transect from tropical lowland to montane forests in NE Ecuador. Trop. Ecol. 2013, 54, 33–45. [Google Scholar]
- Bhuyan, P.; Khan, M.; Tripathi, R. Tree diversity and population structure in undisturbed and human-impacted stands of tropical wet evergreen forest in arunachal pradesh, eastern himalayas, India. Biodivers. Conserv. 2003, 12, 1753–1773. [Google Scholar] [CrossRef]
- Liang, J.; Buongiorno, J.; Monserud, R.A.; Kruger, E.L.; Zhou, M. Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality. For. Ecol. Manag. 2007, 243, 116–127. [Google Scholar] [CrossRef]
- Wolf, J.A.; Fricker, G.A.; Meyer, V.; Hubbell, S.P.; Gillespie, T.W.; Saatchi, S.S. Plant species richness is associated with canopy height and topography in a neotropical forest. Remote Sens. 2012, 4, 4010–4021. [Google Scholar] [CrossRef]
- Slik, J.W.F.; Paoli, G.; McGuire, K.; Amaral, I.; Barroso, J.; Bastian, M.; Blanc, L.; Bongers, F.; Boundja, P.; Clark, C.; et al. Large trees drive forest aboveground biomass variation in moist lowland forests across the tropics. Glob. Ecol. Biogeogr. 2013, 22, 1261–1271. [Google Scholar] [CrossRef]
- Fricker, G.A.; Wolf, J.A.; Saatchi, S.S.; Gillespie, T.W. Predicting spatial variations of tree species richness in tropical forests from high-resolution remote sensing. Ecol. Appl. 2015, 25, 1776–1789. [Google Scholar] [CrossRef] [PubMed]
- Drake, J.B.; Dubayah, R.O.; Knox, R.G.; Clark, D.B.; Blair, J.B. Sensitivity of large-footprint lidar to canopy structure and biomass in a neotropical rainforest. Remote Sens. Environ. 2002, 81, 378–392. [Google Scholar] [CrossRef]
- Tang, H.; Dubayah, R.; Swatantran, A.; Hofton, M.; Sheldon, S.; Clark, D.B.; Blair, B. Retrieval of vertical lai profiles over tropical rain forests using waveform lidar at la selva, Costa Rica. Remote Sens. Environ. 2012, 124, 242–250. [Google Scholar] [CrossRef]
- Dubayah, R.O.; Drake, J.B. Lidar remote sensing for forestry. J. For. 2000, 98, 44–46. [Google Scholar]
- Dubayah, R.O.; Sheldon, S.L.; Clark, D.B.; Hofton, M.A.; Blair, J.B.; Hurtt, G.C.; Chazdon, R.L. Estimation of tropical forest height and biomass dynamics using lidar remote sensing at la selva, Costa Rica. J. Geophys. Res. Biogeosci. 2010, 115. [Google Scholar] [CrossRef]
- Drake, J.B.; Dubayah, R.O.; Clark, D.B.; Knox, R.G.; Blair, J.B.; Hofton, M.A.; Chazdon, R.L.; Weishampel, J.F.; Prince, S. Estimation of tropical forest structural characteristics using large-footprint lidar. Remote Sens. Environ. 2002, 79, 305–319. [Google Scholar] [CrossRef]
- Pires, J.M.; Prance, G.T. The Vegetation Types of the Brazilian Amazon; Pergamon Press: New York, NY, USA, 1985; p. 144. [Google Scholar]
- Austin, M.P.; Pausas, J.G.; Nicholls, A.O. Patterns of tree species richness in relation to environment in Southeastern New South Wales, Australia. Aust. J. Ecol. 1996, 21, 154–164. [Google Scholar] [CrossRef]
- Burnett, M.R.; August, P.V.; Brown, J.H.; Killingbeck, K.T. The influence of geomorphological heterogeneity on biodiversity I. A patch-scale perspective. Conserv. Biol. 1998, 12, 363–370. [Google Scholar] [CrossRef]
- Macarthur, R.; Klopfer, P.H. Population effects of natural selection. Am. Nat. 1961, 95, 195–199. [Google Scholar] [CrossRef]
- Goetz, S.; Steinberg, D.; Dubayah, R.; Blair, B. Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens. Environ. 2007, 108, 254–263. [Google Scholar] [CrossRef]
- Bergen, K.M.; Goetz, S.J.; Dubayah, R.O.; Henebry, G.M.; Hunsaker, C.T.; Imhoff, M.L.; Nelson, R.F.; Parker, G.G.; Radeloff, V.C. Remote sensing of vegetation 3-d structure for biodiversity and habitat: Review and implications for lidar and radar spaceborne missions. J. Geophys. Res. Biogeosci. 2009, 114, 13. [Google Scholar] [CrossRef]
- Clark, D.B.; Hurtado, J.; Saatchi, S.S. Tropical rain forest structure, tree growth and dynamics along a 2700-m elevational transect in costa rica. PLoS ONE 2015, 10, e0122905. [Google Scholar] [CrossRef] [PubMed]
- Holdridge, L. The life zone system. Adansonia 1966, 6, 199–203. [Google Scholar]
- Grieve, I.C.; Proctor, J.; Cousins, S.A. Soil variation with altitude on Volcan Barva, Costa-Rica. Catena 1990, 17, 525–534. [Google Scholar] [CrossRef]
- Team Network. Tropical Ecology Assessment & Monitoring Network: Early Warning System for Nature. Available online: http://www.teamnetwork.org/site/volcan-barva (accessed on 1 January 2013).
- Blair, J.B.; Hofton, M.; Rabine, D.L. Processing of NASA Lvis Elevation and Canopy (LGE, LCE and LGW) Data Products; Center, C.N.G.S.F., Ed.; 2006. Available online: https://lvis.gsfc.nasa.gov (accessed on 1 January 2013).
- NASA’s Land, Vegetation, and Ice Sensor (NASA). Available online: http://lvis.gsfc.nasa.gov (accessed on 1 January 2013).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2011. [Google Scholar]
- Hurlbert, S.H. The nonconcept of species diversity: A critique and alternative parameters. Ecology 1971, 52, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Evolution and measurement of species diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef]
- Lumley, T. R Package: Leaps. Regression Subset Selection. Thomas Lumley Based on Fortran Code by Alan Miller. Available online: http://CRAN.R-project.org/package=leaps (accessed on 18 March 2018).
- Giraud, C. Introduction to High-Dimensional Statistics; Chapman & Hall/CRC: Boca Raton, FL, USA, 2015. [Google Scholar]
- Dijkstra, T.K. Ridge regression and its degrees of freedom. Qual. Quant. 2014, 48, 3185–3193. [Google Scholar] [CrossRef]
- Friedman, J.; Hastie, T.; Tibshirani, R. Regularization paths for generalized linear models via coordinate descent. J. Stat. Softw. 2010, 33, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Muenchow, J.; von Wehrden, H.; Frank Rodriguez, E.; Arismendiz Rodriguez, R.; Bayer, F.; Richter, M. Woody vegetation of a peruvian tropical dry forest along a climatic gradient depends more on soil than annual precipitation. Erdkunde 2013, 67, 241–248. [Google Scholar] [CrossRef]
- Feeley, K.J.; Gillespie, T.W.; Terborgh, J.W. The utility of spectral indices from landsat etm+ for measuring the structure and composition of tropical dry forests. Biotropica 2005, 37, 508–519. [Google Scholar] [CrossRef]
- Cayuela, L.; Benayas, J.M.; Justel, A.; Salas-Rey, J. Modelling tree diversity in a highly fragmented tropical montane landscape. Glob. Ecol. Biogeogr. 2006, 15, 602–613. [Google Scholar] [CrossRef]
- Pau, S.; Gillespie, T.W.; Wolkovich, E.M. Dissecting NDVI-species richness relationships in Hawaiian dry forests. J. Biogeogr. 2012, 39, 1678–1686. [Google Scholar] [CrossRef]
- Gillespie, T.W.; Saatchi, S.; Pau, S.; Bohlman, S.; Giorgi, A.P.; Lewis, S. Towards quantifying tropical tree species richness in tropical forests. Int. J. Remote Sens. 2009, 30, 1629–1634. [Google Scholar] [CrossRef]
- Chambers, J.Q.; Asner, G.P.; Morton, D.C.; Anderson, L.O.; Saatchi, S.S.; Espírito-Santo, F.D.B.; Palace, M.; Souza, C. Regional ecosystem structure and function: Ecological insights from remote sensing of tropical forests. Trends Ecol. Evolut. 2007, 22, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ordonez, Y.; Leblon, B.; Soria-Ruiz, J. Forest Inventory Using Optical and Radar Remote Sensing; INTECH Open Access Publisher: London, UK, 2009. [Google Scholar]
- Simonson, W.D.; Allen, H.D.; Coomes, D.A. Use of an airborne lidar system to model plant species composition and diversity of mediterranean oak forests. Conserv. Biol. 2012, 26, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Ranson, K.J.; Kimes, D.S.; Blair, J.B.; Kovacs, K. Forest vertical structure from glas: An evaluation using LVIS and SRTM data. Remote Sens. Environ. 2008, 112, 107–117. [Google Scholar] [CrossRef]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Elevation (m) | Species Richness | ShannonDiversity Index | Stem Number(#) | Basal Area (m2) | rh25 (m) | rh50 (m) | rh75 (m) | rh100 (m) | Slope (%) | Aspect (°) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A3 | 56 | 107 | 3.82 | 433 | 26.7 | 11.8 | 21.3 | 27.1 | 35.7 | 6.9 | 204.2 |
| A6 | 56 | 91 | 3.55 | 360 | 24.1 | 13 | 20.6 | 27.3 | 38.3 | 6.3 | 213.2 |
| A1 | 61 | 85 | 3.48 | 332 | 27.9 | 14.5 | 22.8 | 28.9 | 38.8 | 6.4 | 187.6 |
| L4 | 66 | 118 | 3.90 | 535 | 22.5 | 12 | 19 | 24.7 | 33.6 | 9 | 152.7 |
| L5 | 75 | 129 | 3.76 | 512 | 22.0 | 14.3 | 23.6 | 28.7 | 38.3 | 15.9 | 282.9 |
| P4 | 76 | 105 | 3.74 | 500 | 25.1 | 9.9 | 20.1 | 28.3 | 38.8 | 10.8 | 221 |
| L3 | 82 | 117 | 3.96 | 481 | 27.0 | 10.2 | 17 | 22.6 | 31.5 | 10.5 | 154.3 |
| P6 | 83 | 124 | 3.91 | 507 | 23.8 | 13.7 | 21.8 | 28.4 | 39.3 | 18.8 | 181.4 |
| P3 | 85 | 111 | 3.81 | 502 | 25.5 | 12.7 | 21 | 26.4 | 34.3 | 14.3 | 190.8 |
| VB1 | 101 | 99 | 3.32 | 430 | 22.9 | 12 | 20.9 | 27.3 | 37 | 6.9 | 183 |
| L6 | 106 | 121 | 3.93 | 552 | 28.0 | 14.1 | 21.4 | 26.2 | 34.5 | 10.2 | 106.9 |
| L2 | 119 | 126 | 3.97 | 607 | 23.6 | 13 | 21.1 | 27.3 | 38.1 | 19.9 | 215 |
| VB2 | 173 | 95 | 3.89 | 461 | 25.2 | 9.9 | 17.4 | 22.6 | 31.5 | 7.6 | 184.5 |
| VB4 | 386 | 151 | 4.39 | 657 | 27.1 | 11.2 | 21 | 28.4 | 39.2 | 20.6 | 165.4 |
| VB3 | 552 | 173 | 4.64 | 606 | 27.5 | 11.6 | 18.5 | 23.7 | 32.2 | 10.8 | 42.2 |
| VB7 | 933 | 68 | 3.49 | 426 | 20.4 | 5 | 12.9 | 20.4 | 29.7 | 14.2 | 164.2 |
| VB8 | 1425 | 59 | 3.36 | 397 | 24 | 5.2 | 11.3 | 16.5 | 22.8 | 12.8 | 229.5 |
| VB5 | 1976 | 47 | 3.12 | 521 | 26.9 | 4.1 | 9.8 | 14.2 | 20.1 | 9.5 | 135.7 |
| VB10 | 2355 | 49 | 3.15 | 569 | 35.8 | 5.3 | 11.6 | 16.4 | 25 | 25.9 | 81.6 |
| VB6 | 2814 | 14 | 1.76 | 1221 | 66.2 | 6.9 | 11.4 | 14.2 | 19.3 | 9.7 | 126.4 |
| Species Richness | Shannon | |
|---|---|---|
| Diversity | ||
| Field data | ||
| Stems | −0.272 | −0.517 * |
| Basal Area | −0.550* | −0.739 ** |
| Topography | ||
| Elevation | −0.771 ** | −0.743 ** |
| Slope | 0.098 | 0.131 |
| Aspect | −0.027 | −0.332 |
| 3-D structure | ||
| rh100 | 0.741 ** | 0.644 ** |
| rh75 | 0.751 ** | 0.630 ** |
| rh50 | 0.745 ** | 0.581 ** |
| rh25 | 0.707 ** | 0.510 * |
| rh100 sd | 0.304 | 0.287 |
| rh75 sd | 0.313 | 0.267 |
| rh50 sd | 0.0389 | 0.063 |
| rh25 sd | −0.228 | −0.288 |
| Predictors | Selected Predictors in Model | r2 | BIC |
|---|---|---|---|
| 1 | Elev. | 0.589 | 0.71 |
| 2 | rh100 + rh25.SD | 0.584 | −4.637 |
| 3 | Aspect + rh100 + rh25.SD | 0.599 | −4.341 |
| 4 | Aspect + rh25 + rh50 + rh25.SD | 0.579 | −3.56 |
| 5 | Aspect + Aspect.SD + rh25 + rh50 + rh25.SD | 0.654 | −3.269 |
| 6 | Aspect + Aspect.SD + rh25 + rh50 + rh25.SD + rh100.SD | 0.689 | −2.429 |
| 7 | Aspect + Slope.SD + Aspect.SD + rh25 + rh50 + rh25.SD + rh100.SD | 0.711 | −0.865 |
| 8 | Elev. + Aspect + Slope.SD + Aspect.SD + rh25 + rh50 + rh25.SD + rh100.SD | 0.721 | 1.363 |
| 9 | Elev. + Aspect + Aspect.SD + rh25 + rh50 + rh75 + rh100 + rh25.SD + rh50.SD | 0.745 | 2.574 |
| 10 | Aspect + Slope.SD + Aspect.SD + rh25 + rh50 + rh75 + rh100 + Elev.SD + rh25.SD + rh50.SD | 0.79 | 1.698 |
| Predictors | Selected Predictors in Model | r2 | BIC |
|---|---|---|---|
| 1 | Elev. | 0.547 | −5.958 |
| 2 | Elev. + rh25.SD | 0.541 | −10.202 |
| 3 | Elev. + Slope + Aspect | 0.737 | −11.921 |
| 4 | Elev. + Slope + Aspect + rh25 | 0.787 | −15.231 |
| 5 | Elev. + Slope + Aspect + Aspect.SD + rh25 | 0.824 | −16.82 |
| 6 | Elev. + Slope + Aspect + Aspect.SD + rh25 + rh25.SD | 0.838 | −15.455 |
| 7 | Elev. + Slope + Aspect + Aspect.SD + rh25 + Elev.SD + rh75.SD | 0.872 | −17.257 |
| 8 | Elev. + Slope + Aspect + Aspect.SD + rh25 + rh50 + Elev.SD + rh75.SD | 0.878 | −15.145 |
| 9 | Elev. + Slope + Aspect + Aspect.SD + rh25 + rh50 + rh75 + Elev.SD + rh75.SD | 0.914 | −19.152 |
| 10 | Elev. + Aspect + Slope.SD + Aspect.SD + rh25 + rh50 + rh75 + rh100 + rh25.SD + rh50.SD | 0.941 | −23.79 |
| Model Method | Shannon Diversity Model | Species Richness Model |
|---|---|---|
| Best Subsets Multiple Linear Regression | 0.018 | 572.172 |
| Ridge Regression | 0.387 | 2628.755 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, C.; Saatchi, S.; Clark, D.; Hurtado Astaiza, J.; Hubel, A.F.; Gillespie, T.W. Topography and Three-Dimensional Structure Can Estimate Tree Diversity along a Tropical Elevational Gradient in Costa Rica. Remote Sens. 2018, 10, 629. https://doi.org/10.3390/rs10040629
Robinson C, Saatchi S, Clark D, Hurtado Astaiza J, Hubel AF, Gillespie TW. Topography and Three-Dimensional Structure Can Estimate Tree Diversity along a Tropical Elevational Gradient in Costa Rica. Remote Sensing. 2018; 10(4):629. https://doi.org/10.3390/rs10040629
Chicago/Turabian StyleRobinson, Chelsea, Sassan Saatchi, David Clark, Johanna Hurtado Astaiza, Anna F. Hubel, and Thomas W. Gillespie. 2018. "Topography and Three-Dimensional Structure Can Estimate Tree Diversity along a Tropical Elevational Gradient in Costa Rica" Remote Sensing 10, no. 4: 629. https://doi.org/10.3390/rs10040629