1. Introduction
Northern peatlands have played a significant role in carbon (C) accumulation for millennia [
1]. Because of their large extent in northern regions (e.g., 12% of Canada, or 1 million km
2) [
2,
3,
4], it is important to evaluate their potential response to climate change [
5,
6]. Remote sensing studies of peatlands from both hyperspectral airborne and multispectral satellite sensors have shown potential in providing fundamental information about vegetation (e.g., leaf area index) and hydrology, especially with respect to the spatial patterns and temporal trends of these characteristics at large spatial scales (>100 ha) (e.g., [
7,
8,
9,
10,
11]). The advantage of airborne hyperspectral imagery in peatland research is that it can be collected at a fine spatial resolution (≤1 m) to carry out detailed mapping and modeling studies (e.g., [
8,
9,
12]). However, due to the inaccessible nature of many peatlands [
3] and the elevated cost of such data acquisition, especially for isolated northern peatlands, satellite imagery might be the most feasible long-term monitoring solution for these ecosystems. A novel aspect of peatland research using remotely sensed data is the integration of airborne hyperspectral imagery with multispectral satellite data for ecosystem modeling (e.g., hydrology, biogeochemistry).
Optical imagery in the VNIR (visible-near infrared) and SWIR (shortwave infrared) ranges (400–2500 nm) has been used to determine spectral differences between mosses and vascular plants in peatlands [
13]. Spectral indices incorporating both narrow and broadband sensors (e.g., [
14]) successfully assessed the relationship between surface moisture and water table position experimentally and at small spatial scales [
8,
15]. However, remote sensing studies at the ecosystem level are still needed to evaluate the full potential of such imagery, especially for peatlands where in situ data are available. Furthermore, a more thorough understanding of the effects of illumination and viewing geometries are needed for studies considering seasonal changes.
The difference between the carbon dioxide uptake through gross primary production (GPP) and ecosystem respiration represents the net ecosystem exchange (NEE) [
16]. In situ measurement of NEE via approaches such as flux chambers can be utilized to assess responses to diverse variables, including nutrient inputs (e.g., [
17]) and manipulations of water content [
18]. Eddy covariance towers are also commonly used in the assessment of NEE in peatland ecosystems, with the advantage that these provide a larger footprint than chambers and can provide near-continuous long-term data [
19,
20]. For large-scale studies, MODIS-derived products have been used to assess NEE [
21,
22], however, the large spatial resolution (e.g., 0.5–1 km) is inadequate to capture the small-scale heterogeneity of peatlands, along with other known limitations for peatlands such as the errors in the vapor pressure deficit determined from land cover (for GPP estimation) [
23,
24]. More recently, the potential of Landsat 7 ETM+ data (30 m resolution) was shown for determining peatland classes with high accuracy [
23], from which to estimate the C balance. Given the long history of the Landsat program, a less explored aspect is the development of C models (e.g., NEE) taking advantage of these multitemporal data. In addition, new satellites such as Sentinel-2 provide additional opportunities to evaluate Landsat products at finer spatial scales (e.g., 10–20 m spatial resolution), which may better capture the spatial heterogeneity of these ecosystems.
Biogeochemical processes such as gas exchange (e.g., CO
2, CH
4) and C accumulation in northern peatlands are closely related to the water table position [
25,
26,
27]. Given a lowering of the water table, an increase in CO
2 loss is expected [
25]. For example, the interannual variability of C exchange at the Mer Bleue peatland indicated that this ecosystem reduced its CO
2 sink capacity during summertime drought events [
20]. However, there is high variability in CO
2 exchange across peatland sites and seasonally within sites [
16]. Spatial variability of the vegetation composition [
28] and other variables, such as temperature [
29,
30], also impact the overall C budget of these ecosystems. Therefore, ongoing characterization of the spatial and temporal variability of the water table position and CO
2 uptake in peatlands is necessary.
In this study, we addressed two distinct but complementary objectives for estimating water table depth and NEE at the Mer Bleue Conservation Area using remotely sensed data. The first objective was to assess the relationship between a surface moisture index derived from SWIR hyperspectral imagery and water table depth. We hypothesized that changes in vegetation surface moisture throughout the growing season at Mer Bleue are closely related to water table depth [
8]; therefore, such an index could be used as a proxy. The second objective was to develop and test a NEE model based on a modified water index (MWI) calculated from multitemporal Landsat TM5 and Landsat 8 OLI data. As vegetation moisture content and phenology are closely related to CO
2 uptake in peatlands [
8], we predicted that the model could serve as a good estimator of NEE at large spatial scales. We applied the resultant NEE models to multitemporal Sentinel-2A and airborne hyperspectral VNIR imagery to determine spatial and temporal trends during the 2016 growing season and compare the results with observed NEE from an eddy covariance tower. This study builds upon results by [
31], mapping near surface water content in hollows and light-saturated gross photosynthesis for hummocks in the same peatland.
4. Discussion
In this study, we illustrate the potential of multitemporal airborne and satellite imagery for modeling water table depth and NEE for an ombrotrophic peatland. These remotely sensed data provide opportunities for ongoing monitoring of peatlands at a range of spatial and temporal scales. Physical models of peatland C balance require inputs from in situ measurements—such as precipitation, temperature, incoming radiation, and wind speed, among others—for successful parameterization [
54,
55]. While these variables are straightforward to collect for small spatial scales such as at individual eddy covariance towers, in situ measurements over large spatial extents are logistically unfeasible [
56]. Estimations of the parameters are also possible from regional or global climate models, but this may introduce additional uncertainties for predictions of hydrological behavior and gas exchange at the ecosystem level by not taking into account the spatial heterogeneity of the system. Remote sensing observations can help bridge the gap between small spatial scale, in situ measurements of peatland processes, and ecosystem scale models. Long-term satellite archives further provide the opportunity for historical assessment or estimations of processes at locations without in situ measurements. New generation satellite imagery (e.g., Sentinel-2, Landsat 8 OLI) and airborne hyperspectral imagery allow for highly detailed image acquisitions from which to monitor the response of these ecosystems to both allogenic (e.g., climate) and autogenic factors (e.g., change in hydrology).
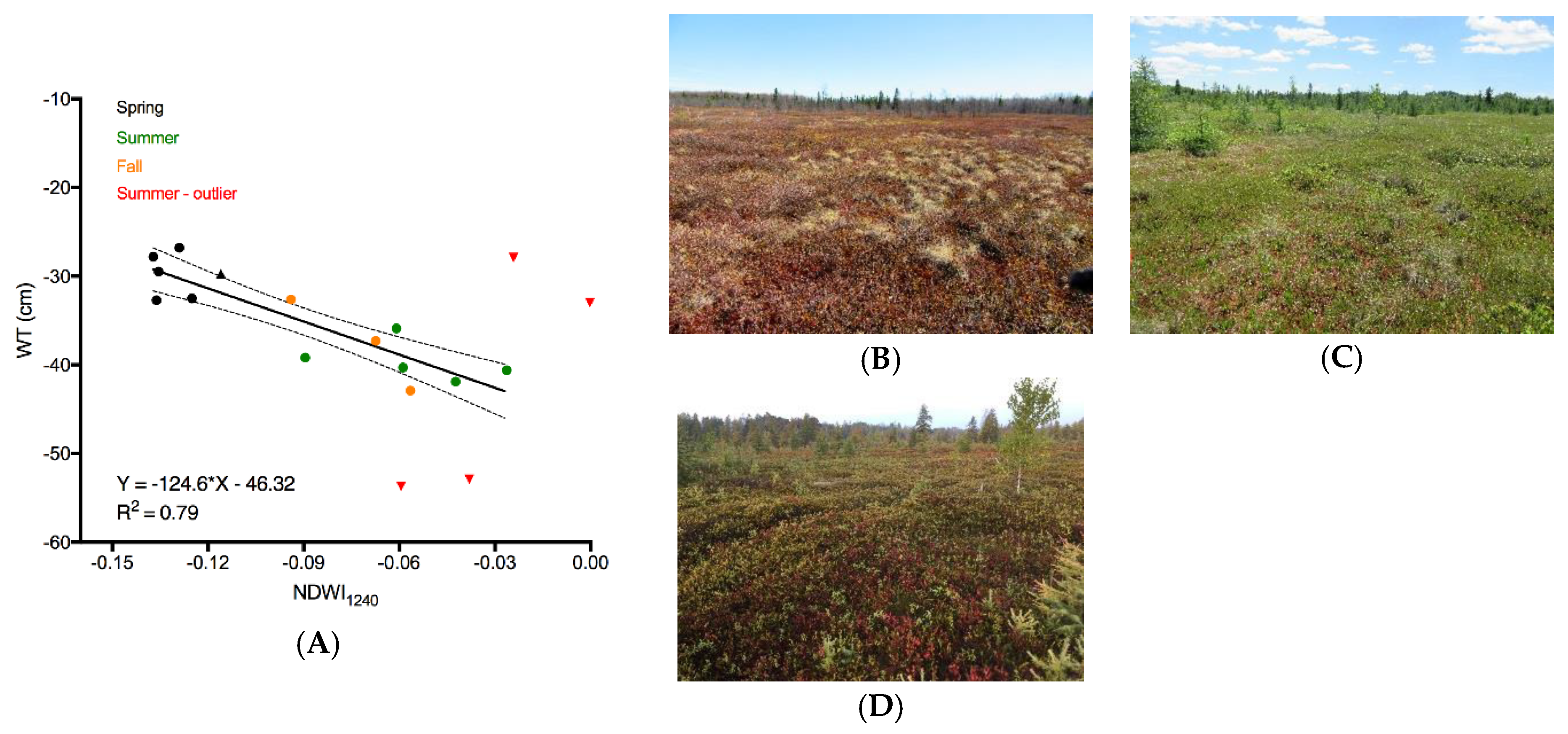
The multitemporal model relating NDWI
1240 to water table depth illustrates the sensitivity of the narrowband index to both liquid water content in the vegetation and phenology. Mer Bleue has both deciduous and evergreen vascular plants and there are substantial seasonal variations in nitrogen and chlorophyll concentration in the vascular plants, but not in the mosses [
9]. These changes can be seen in multitemporal imagery of the bog [
9,
51]. With the depth of the water table position having been greater than 20 cm from the surface on the days the SASI HSI were collected over the period of study (2011–2016), the NDWI
1240 does not directly observe the position of the water table. The reflectance at these wavelengths is determined solely by the top 3–5 cm of the
Sphagnum canopy [
57]. While other studies have used exposed surface water to infer the position of the water table across a peatland [
58], the relationship here shows NDWI
1240 as a proxy for water table position. This is important for Mer Bleue because, other than during exceptionally wet periods, the water table remains below the surface of the hollows [
20]. Water table depth is a subdued reflection of the surface microtopography [
35], with a strong association to the vegetation community [
1]. The magnitude of reflectance at 884.5 nm and 1238.6 nm are indicative of the moisture content in the superficial bog vegetation. As shown by [
56], the magnitude of
Sphagnum reflectance can vary by as much as 50% in the SWIR region from saturated to dry samples. In particular, the characteristic peaks in reflectance centered between 799–900 nm and 1250–1330 nm in wet
Sphagnum are lost as the amplitude of the reflectance in NIR–SWIR increases with drying. Extrapolating the model beyond the range of the WT table depth investigated here should be done with caution because it is unknown if the NDWI
1240:WT relationship would remain with very high (i.e., water at surface of hollows) or low (<−43 cm) WT positions. However, at Mer Bleue, a very high WT is rare; it mostly remains below the surface of hollows.
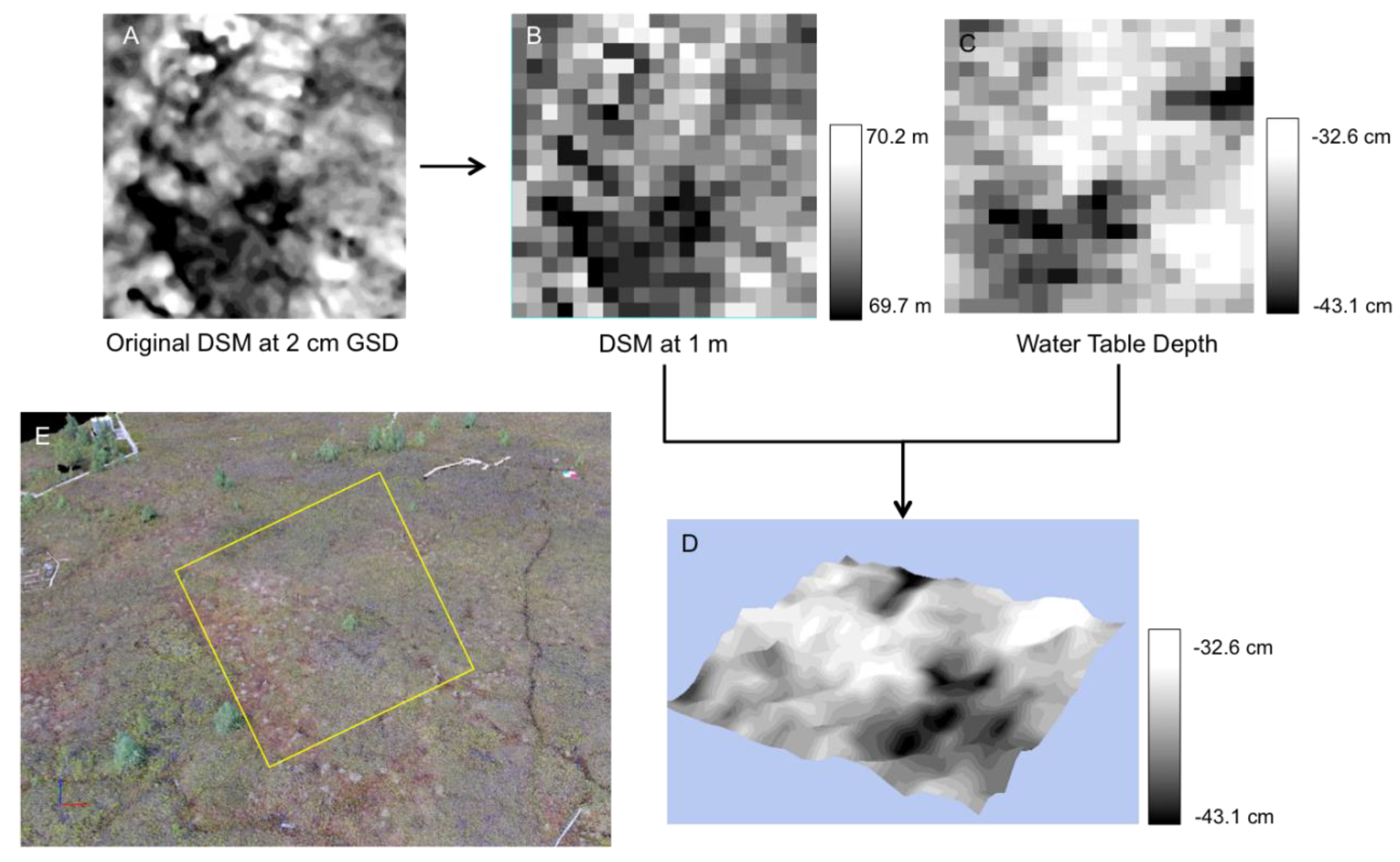
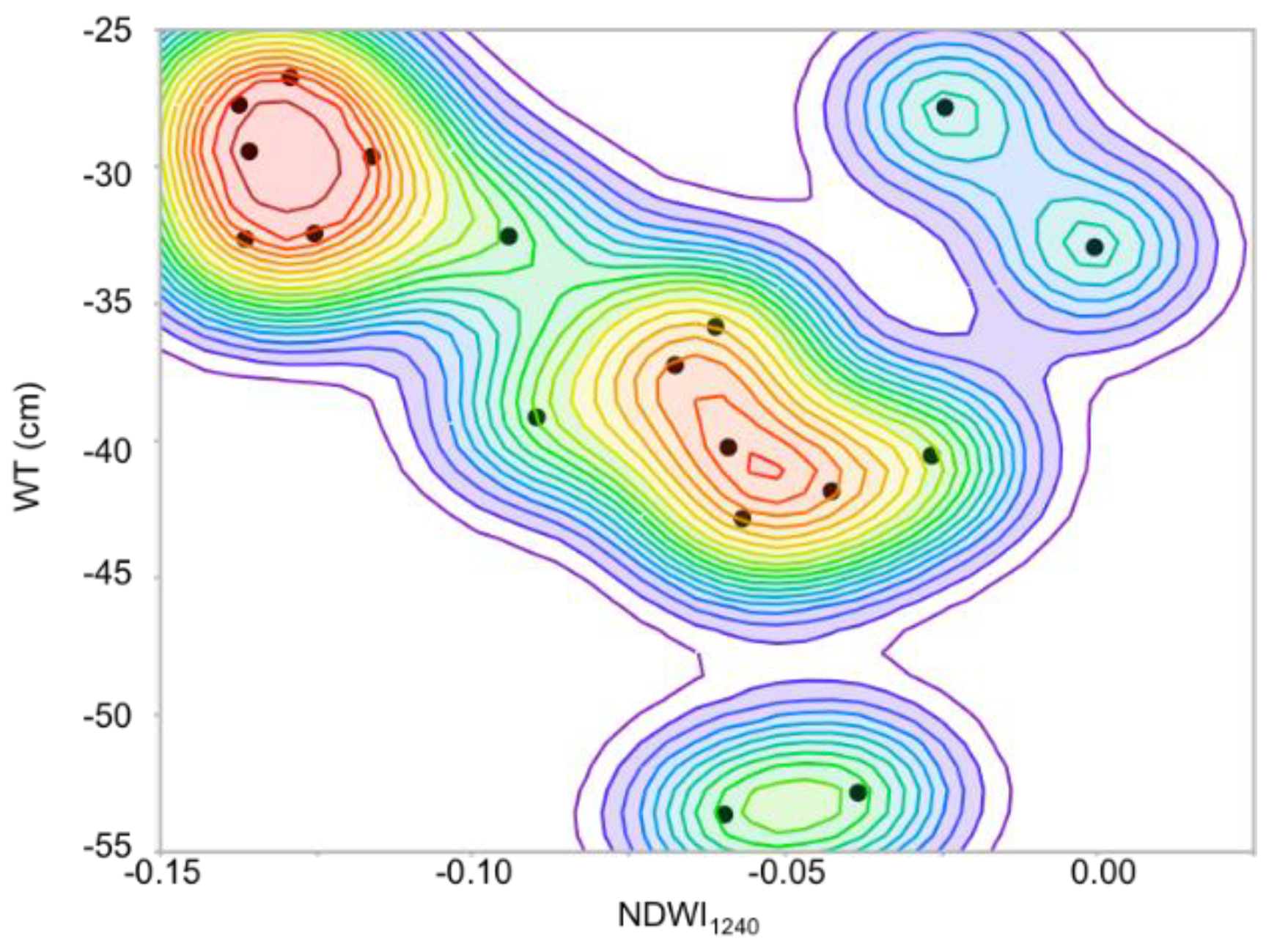
We found a lower correlation between microtopography and water table depth (
Figure 6) than [
1]. This is due to the spatial resolution (1 m) of the SWIR imagery and the degraded microtopography DSM (2 cm to 1 m). Due to the small spatial scale of the hummocks and hollows, 1 m does not retain their integrity [
31], resulting in a mixing of both the elevation and water table at the transition between hummocks and hollows. If these analyses were to be conducted at a finer spatial resolution (e.g., 5 cm), achievable from low-altitude UAV platforms, the correlation between microtopography and water table depth would likely be stronger. Further work investigating this relationship at fine spatial scales is important because UAV-based microtopography modeling is less expensive and more accessible than UAV based SWIR HSI.
Peatland hydrology is one of the most important factors influencing ecology and functioning, with WT depth an important predictor of vegetation structure and composition [
28]. In areas with a deeper WT, vascular vegetation in hummocks is taller, with the establishment of trees in areas with the lowest WT [
59]. Therefore, long-term changes in the WT depth could alter the spatial distribution and structure of the vascular plants. The position of the WT in the peat profile indicates the depth of the soil air in the pore space, while the vertical range between the surface and the maximum water table depth encompasses the thickness of the acrotelm in which most of the biogeochemical processes take place [
28]. Because the roots of many vascular plants require a minimum proportion of soil pore air, temporal monitoring of the WT depth (in a spatial context) could facilitate forecasting succession (i.e., changes in species composition or community structure). The vertical profile of
Sphagnum is a dense canopy with spaces and dead hyaline cells of the leaves and branches providing the mechanism for the retention of capillary water above the water table [
28]. Lowering of the WT decreases the soil water pressure [
60], increasing the possibility of desiccation of the mosses thus resulting in a loss of productivity and carbon sequestration [
61]. Substantial lowering of the WT through draining accelerates the decomposition process in the peat and releases of CO
2 to the atmosphere [
62].
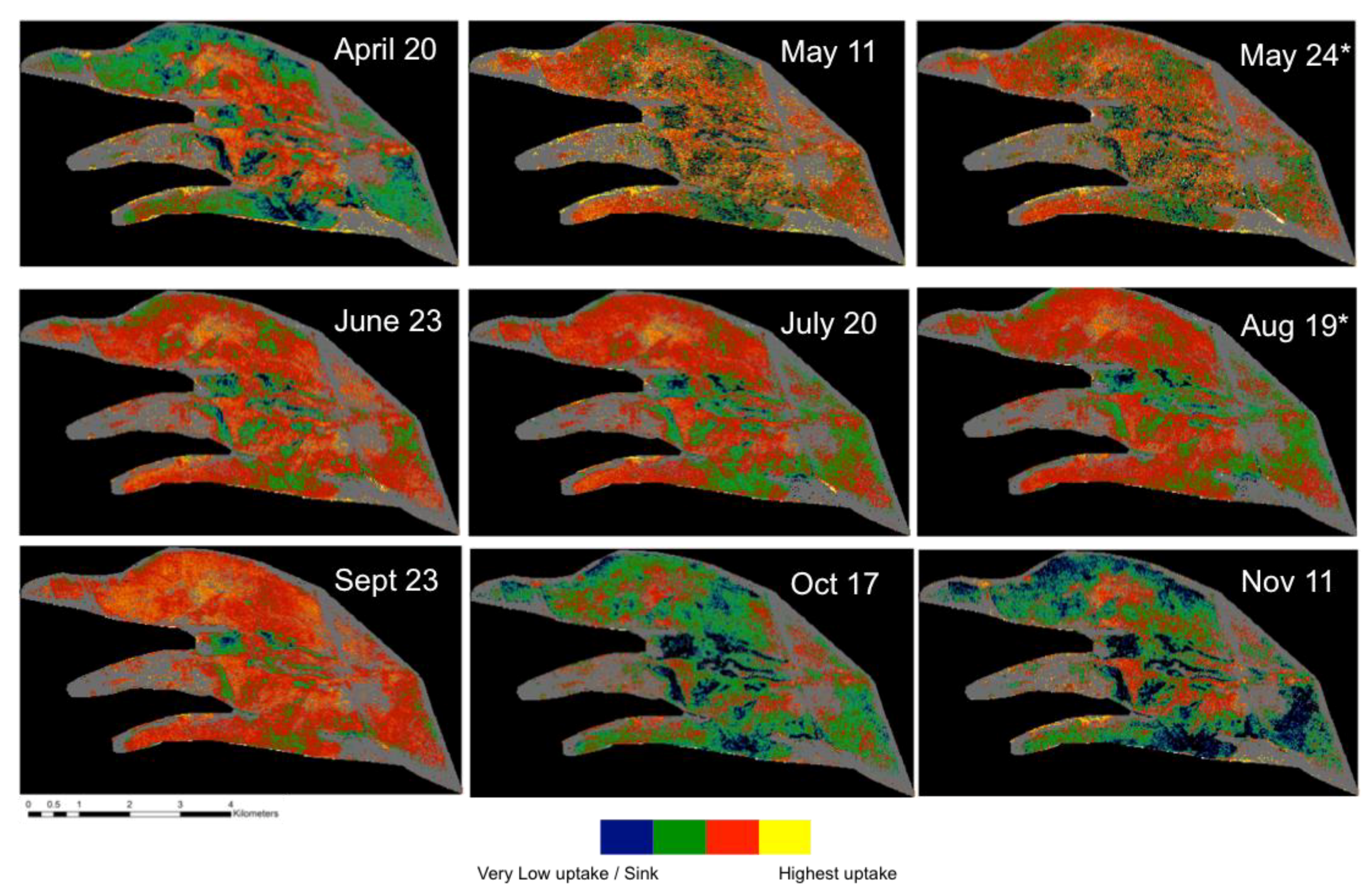
In order to estimate WT with NDWI
1240 from airborne HSI, the illumination and acquisition geometries should be carefully considered. As shown in
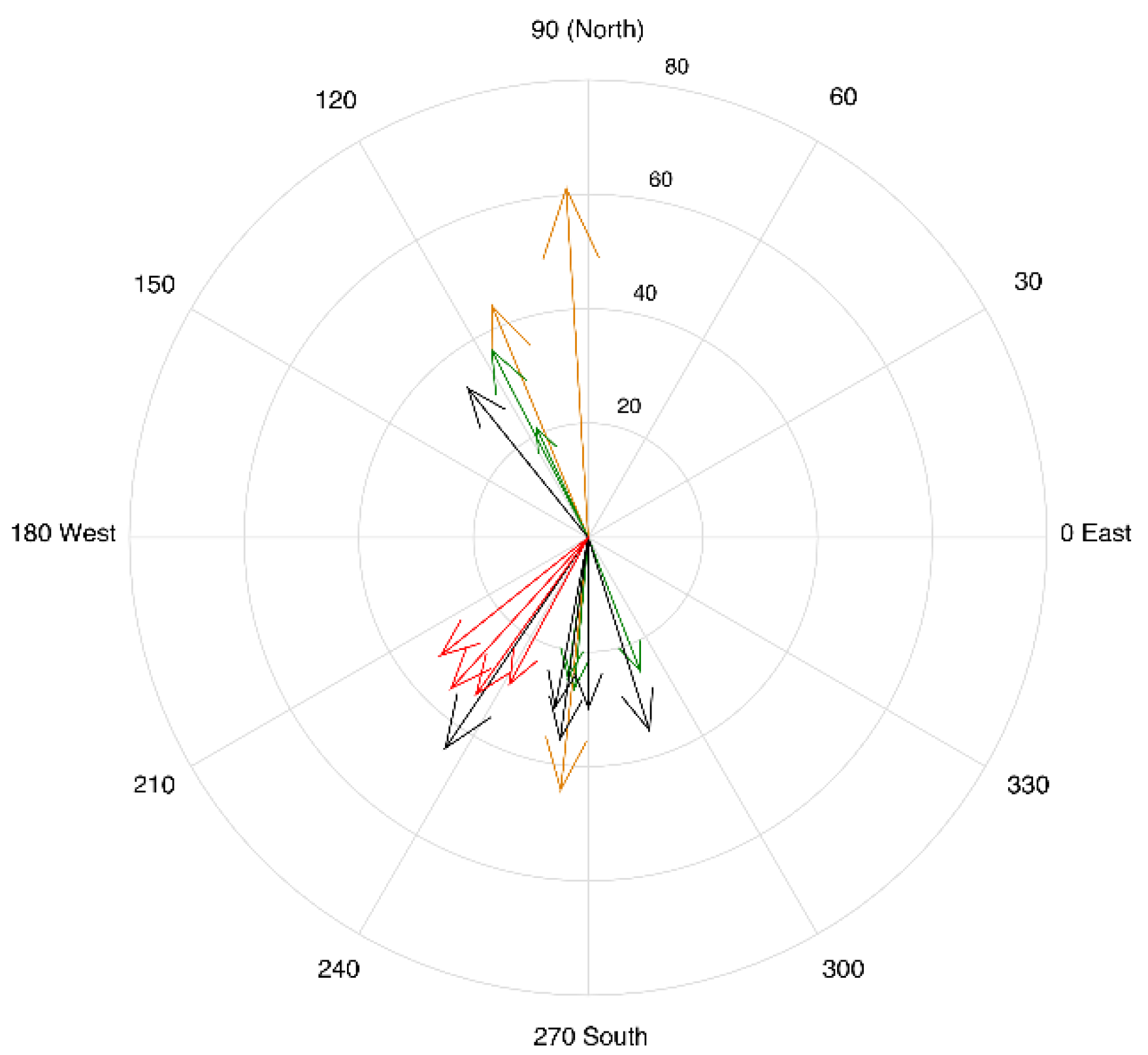
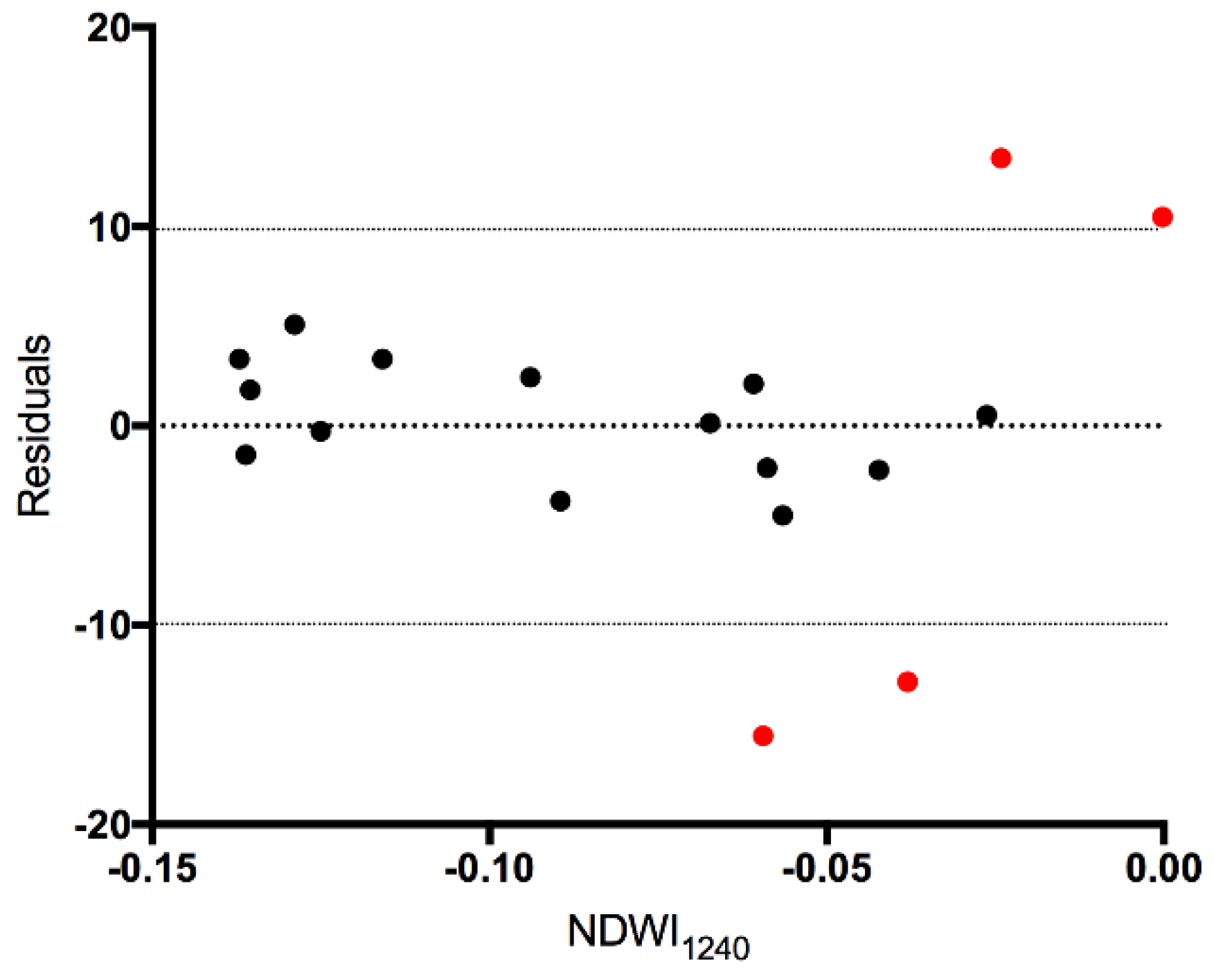
Figure 4, during the summer, with the vascular plants in full foliage, the illumination geometry affects the utility of the image for WT position estimation. The residuals (
Figure A1) indicate that the flight lines in the summer (leaf on) with the RAA diagonal to the field of view (128.7–151.8°) were greater than ±10 cm. A non-parametric quantile density plot (
Figure A2) further illustrates the dissimilarity between the NDWI
1240:WT relationship in these HSI and the rest of the sampling dates. As a result, these four images were not used in the estimation of the WT depth. In contrast, the spring (leaf off) his, with an RAA of 145.9° (black triangle in
Figure 3A), was not an outlier in the NDWI
1240:WT relationship due to lack of leaves on the vascular plants (
Figure 3D and
Figure 4). While additional data collection (e.g., HSI collected over a broader range of solar zenith angle (SZA) and SAA throughout the growing season) and analysis are planned to fully understand the reason for this, we believe it is potentially related to the anisotropy of the bog. When reflectance properties of a surface are not perfectly diffuse, they have some degree of anisotropy. This directional characteristic of the surface reflectance when accounted for from all angles is referred to as the bidirectional reflectance distribution function (BRDF) [
63]. Vegetation, in general, has long been accepted as having anisotropic reflectance properties [
64], however, the majority of studies have focused on modeling and quantifying other ecosystems with minimal studies examining peatlands [
65].
Understanding the effects of the anisotropic properties of peatlands on remotely sensed data is important for biogeochemical modeling because transformations applied to reflectance (such as vegetation indices) are strongly affected by BRDF [
66]. Goniometer measurements of moss BRDF have indicated that the infrared region showed a greater degree of variability with change in azimuth angle than the visible wavelengths [
67]. For moss samples, there was no pronounced hot spot [
67]. Instead, the BRDF effects constitute a higher reflectivity perpendicular to the illumination angle at low view angles (similar to illumination conditions found at high latitudes). The BRDF properties of vascular plant canopies are primarily characterized by a hotspot, a peak in reflectance when the sun is directly behind the sensor [
61]. At the ecosystem scale, BRDF is a complex process, influenced not only by crown size, density, and spacing between crowns, but also the background soil BRDF, which for peatlands is the moss canopy. Vascular vegetation phenology has also been observed to strongly influence BRDF properties [
63,
68]. As such, their influence on the overall surface reflectance of the peatland must also be considered.
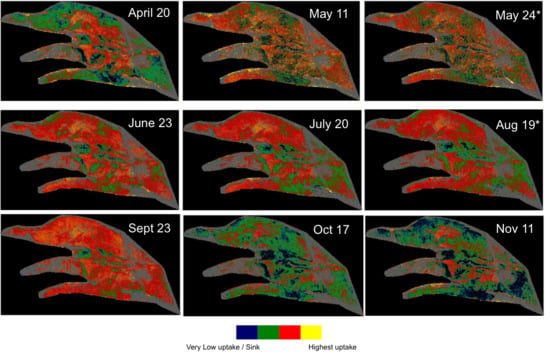
The broader implication of these observations relates to the mission planning of airborne imagery coincidental with satellite overpasses. At the latitude of Mer Bleue, coincidental image acquisition with Sentinel-2 and Landsat 8 OLI with mission planning aiming to minimize cross-track illumination effects results in suboptimal illumination conditions in the summer (i.e., vascular plants in full foliage) for retrieving biogeochemical properties of the peatland such as near surface water content [
31] or WT position relying on SWIR wavelengths. While wavelengths in the VNIR range should be less impacted, allowing for the retrieval of other characteristics such as pigments contents [
9], analyses requiring the SWIR region need additional considerations. Application of similar analyses to satellite based SWIR bands (e.g., Sentinel-2 bands 11 and 12) could face comparable challenges without a comprehensive BRDF correction [
69].
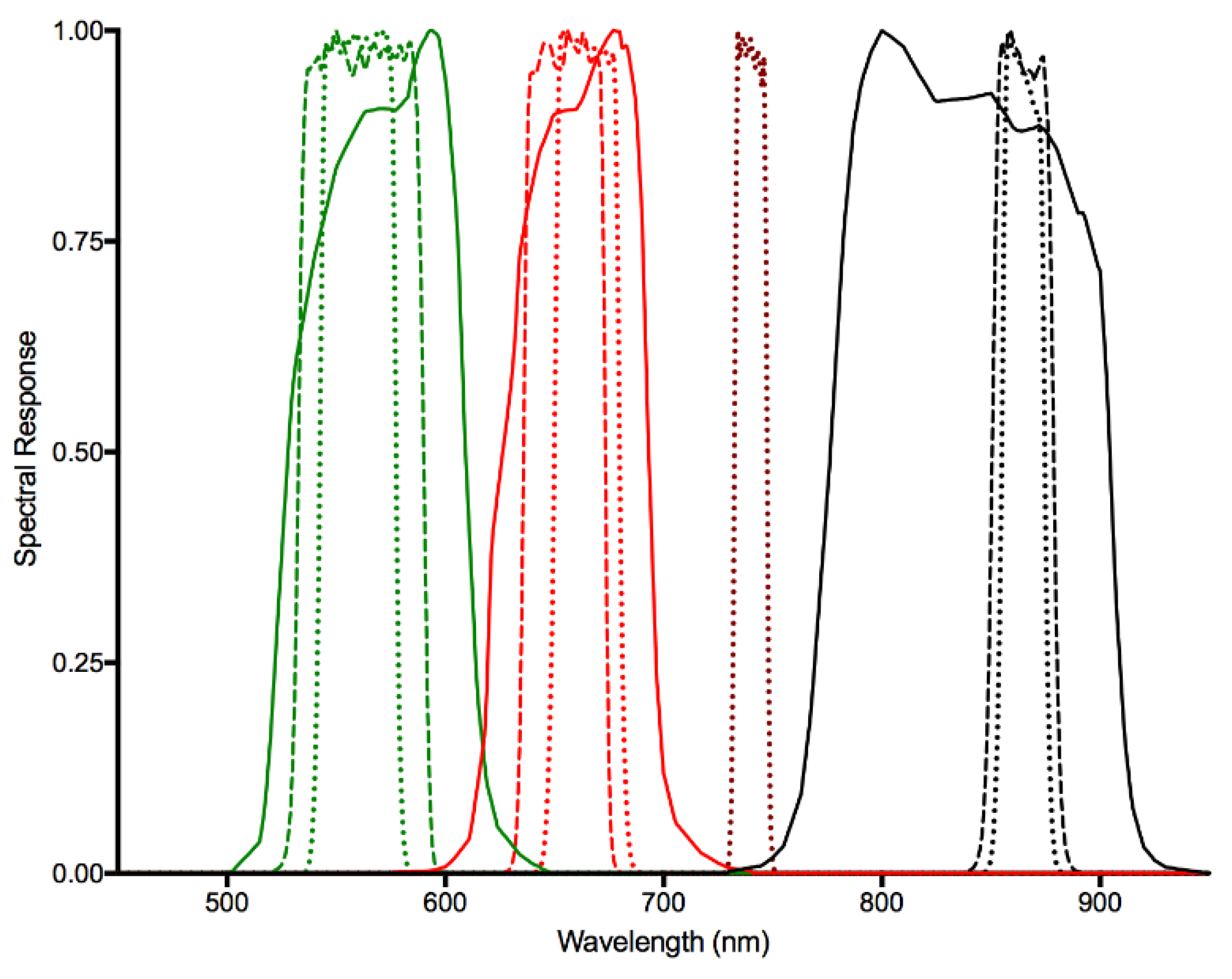
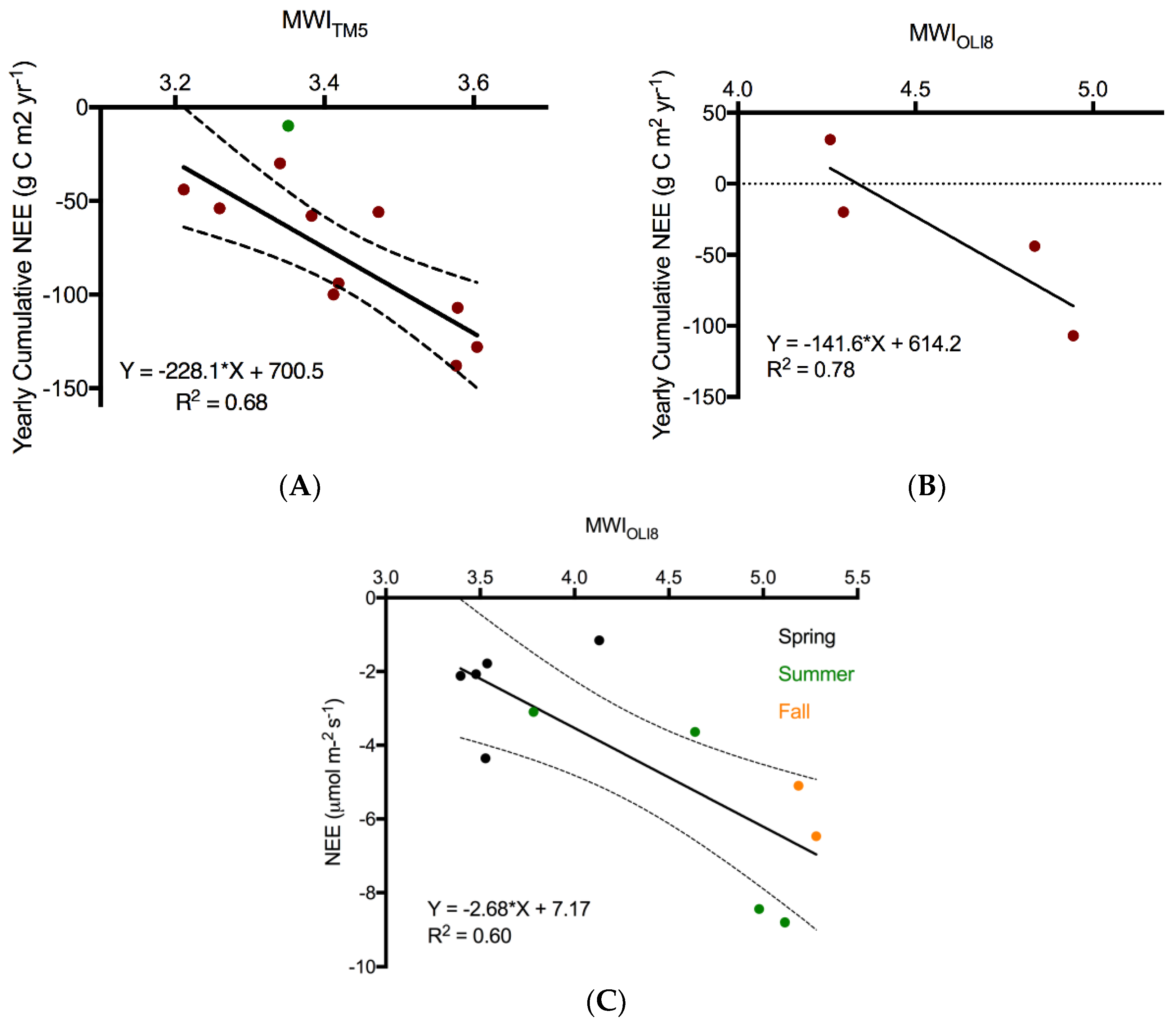
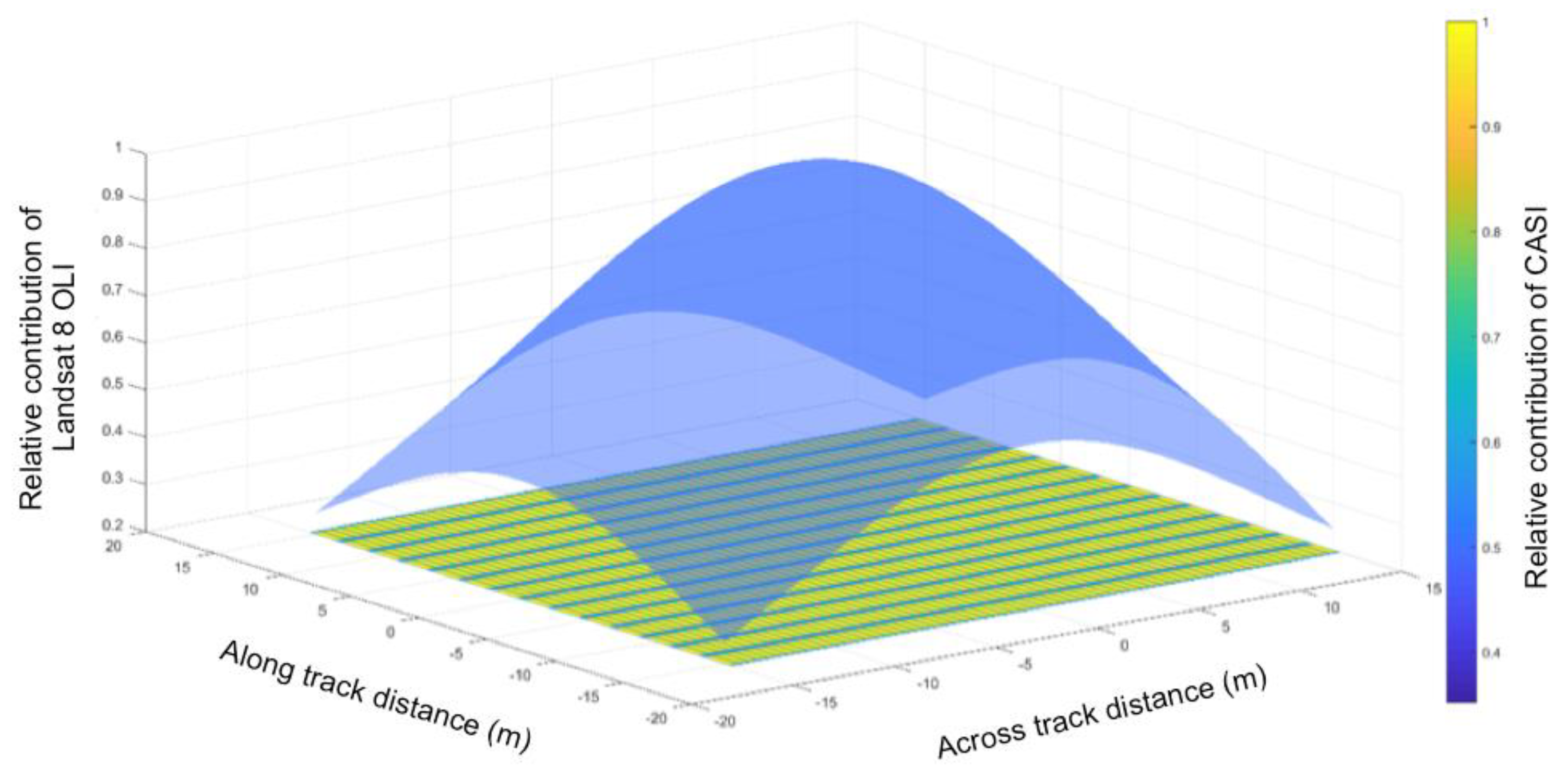
Despite our utilization of the new Collection 1, Tier 1 surface reflectance imagery for the Landsat TM5 and OLI 8 multitemporal models predicting NEE, it was not possible to combine data from the two sensors under our current framework. The fundamental differences in spectral response (
Figure 2), spatial response (i.e., line spread functions), radiometry (e.g., SNR, and radiometric resolution), and geometry (location on Earth contributing to the signal) [
70] are likely the primary factors in the differences between the models derived from TM5 and OLI8 imagery. Several important changes have been made to the Landsat sensors over the years, resulting in improvements in data quality from Landsat 8 OLI in comparison to its predecessors as described in detail by [
70]. However, aside from the obvious spectral resolution differences (
Figure 2), we believe two other aspects influencing the specificity of the data acquired are important to consider in this context: instrument design (whiskbroom TM5 vs. pushbroom OLI 8) and radiometric resolution (8 bit TM5 vs. 12 bit OLI 8). The increased dynamic range (from the noise floor to the maximum radiance levels for each band) also leads to an improved noise and quantization performance [
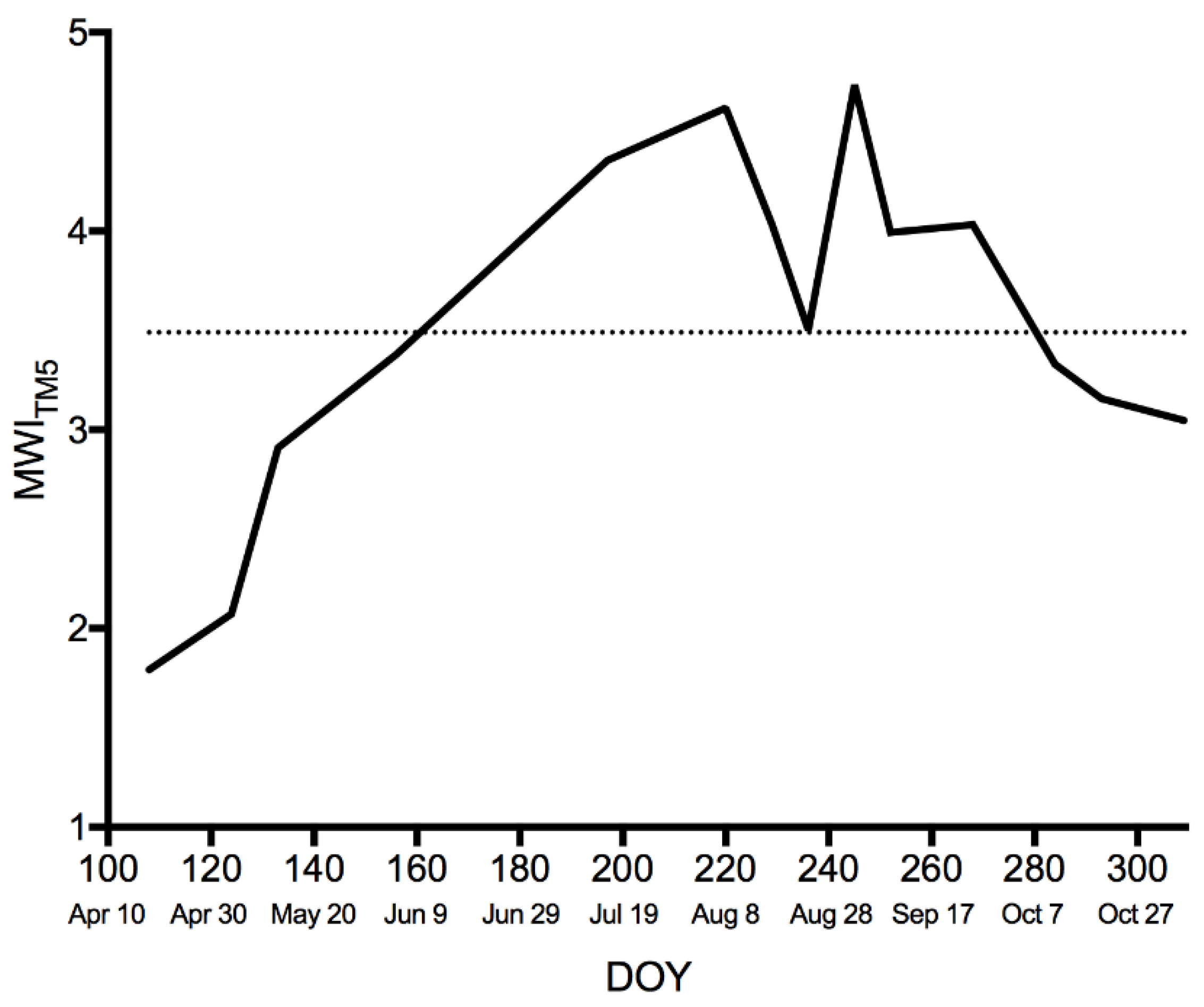
71]. The increased radiometric resolution and higher SNR are likely largely responsible for the greater range of MWI from Landsat 8 OLI over a similar range in observed NEE (
Figure 8). Experimental degradation of the radiometric resolution of Landsat 8 OLI from 12 to 8 bits illustrated a loss of information at 9 bits, where the distribution of the original 12-bit data was no longer preserved (analysis not shown).
The multiple focal plane array module (FPA) pushbroom design of OLI 8 (and Sentinel-2) replaces the mirror scanning mechanism from Landsat TM5 resulting in, among other characteristics, improved geometric accuracy. However, the stability of the band-to-band registration from the whiskbroom design is more challenging to achieve with the modern sensor approach making use of the multiple FPA pushbroom design [
70]. Because the detectors for the different bands are separated in the along-track direction, there is a time delay between the different bands as they image the same location on Earth. For OLI 8, this is estimated to be 1.1 s [
70], which results in terrain parallax creating challenges in the removal of high altitude cirrus (i.e., band-to-band registration between the cirrus and optical channels) and changes in satellite position and attitude that must be accounted for to establish the line of sight [
44,
70,
71,
72,
73,
74,
75]. Despite efforts to standardize the continuity of land surface products between subsequent generations of satellite sensors [
76], our results suggest caution in combining data from sensors with greatly different spectral and radiometric properties for peatland biogeochemical modeling. There is a further consideration of the sensor design related to the offset between the two rows of FPAs in both OLI 8 and Sentinel-2; one set has a slightly forward view of the Earth while the other set has a slightly rearward view [
71,
72,
75]. This is an important consideration for peatlands in the summer, where the vascular plant component has stronger BRDF properties. The difference in the radiance collected by the two sets of FPAs for targets with substantial BRDF properties may be uncorrectable [
73], but this effect is mitigated by updated band-specific gain values.
The challenge of removing high-altitude cirrus is seen in our results with the Sentinel-2 imagery from 24 May to 19 August. Full removal of high-altitude cirrus from satellite imagery is not a trivial problem [
77,
78,
79,
80,
81]. On Landsat 8 OLI, the cirrus band is affected by spectral cross-talk with contamination from the neighboring SWIR band [
71] and both Sentinel-2 and Landsat 8 OLI have various magnitudes of band spatial misregistration. This leads to a potential problem for the cirrus removal if the channels have different parallax angles [
80,
81].
A further consideration requiring more in-depth analyses is that data from all sensors underwent atmospheric correction through different models. Landsat TM5 Collection 1, Tier 1 imagery was atmospherically corrected using the Landsat Ecosystem Disturbance Adaptive Processing System (LEDAPS) [
82]. The Landsat 8 OLI Collection 1, Tier 1 imagery was atmospherically corrected using the Landsat Surface Reflectance Code (LaSRC) optimized for Landsat 8 [
83]. Sentinel-2A was atmospherically corrected with Sen2cor [
45] and the SASI and CASI HSI were atmospherically corrected with ENVI FLAASH 5.4.1 or ATCOR 4.7.0. While there is no comprehensive comparison between these five approaches to atmospheric correction, differences are expected [
84]; ATCOR and FLAASH are based on MODTRAN, while LEDAPS and LaRSC implement the MODIS/6S and Vectorial 6S (6SV) models, respectively. Sen2Cor implements a custom algorithm (Sentinel-2 Atmospheric Correction—S2AC) based on the libRadtran4 code [
85]. Differences have been shown between the approaches for the two Landsat sensors, where LaSRC processing provides improvements in the surface reflectance over LEDAPS [
83]. Similarly, differences have been observed between ATCOR and FLAASH for airborne HSI [
86]. However, more extensive comparisons are needed between the HSI and satellite approaches to determine the uncertainty introduced by the different implementations of atmospheric correction and their effect on peatland biogeochemical models.
Finally, our results contribute to the growing body of literature illustrating hyperspectral imagery improves the retrieval of fundamental peatland characteristics over multispectral data at all spatial scales. The HSI used here both have a 14-bit dynamic range. From approximately 1000 m AGL flight altitude, neither the CASI nor SASI were close to saturation; over the peatland only approximately one-third of the dynamic range was utilized. While this may appear to be a cause for concern for low overall SNR, it is not a problem because summation (on- or off-chip) can be used to improve SNR, if needed [
38]. The multiple finer spatial pixels (
Figure A3) from airborne HSI in comparison to satellite imagery provide a more uniform contribution of the ground elements to the imagery. This is an important consideration for peatlands with fine (≤1 m) spatial structures (e.g., hummocks and hollows). The underutilized dynamic range of the CASI and SASI also suggests that miniaturized versions of the HSI with similar spectral and radiometric properties could be successfully deployed on UAV platforms, where the sensor would be considerably closer to the peatland surface (e.g., <150 m AGL). The higher spatial resolution from UAV-based imagery would benefit WT position models. Planned hyperspectral satellite missions (e.g., EnMap, WaterSat, HyspIRI) could further improve the ongoing remote monitoring of peatlands globally in light of increased pressures from various drivers of change.
5. Conclusions
We successfully modeled WT depth and NEE from airborne hyperspectral and satellite multispectral imagery, respectively. To the best of our knowledge, this is the first time these parameters were retrieved from optical imagery for a peatland at such a fine spatial resolution (1 m for WT depth and 20–30 m for NEE). The narrow band index NDWI1240 was shown to be sensitive to both liquid water content in the vegetation as well as phenology and, therefore, can be used as a proxy for estimating WT depth.
For the retrieval of NEE, we propose separate models, one for historical estimations from Landsat TM5 and a separate model going forward with Landsat 8 OLI or Sentinel-2. We found it was not possible to combine data from sensors with very different spectral and radiometric properties into one model. Despite the successful retrieval of NEE from satellite imagery, the use of airborne HSI improved the relationship with eddy covariance tower-measured NEE. We believe the higher radiometric resolution of the airborne HSI and much finer spatial and spectral resolutions are the reasons for this difference.
The anisotropic properties of the peatland should be considered for the collection of SWIR airborne HSI in the summer when the vascular plants are in full foliage, however, this is less of a concern in the spring prior to green-up. Multitemporal HSI should be collected with consistent illumination and viewing geometries in these types of studies, with the RAA in line with the heading as much as possible. For satellite-based estimations of peatland biogeochemical properties, care must be taken with regards to the detection and removal of high-altitude cirrus. Despite the transparent nature of these clouds, they impact the surface reflectance products, and scenes with cirrus contamination cannot be used to reliably retrieve NEE.
Despite their importance, uncertainties about the total global peatland spatial extent persist, due in part to difficulties in mapping peatlands from coarse resolution remotely sensed data [
4,
10]. Recent estimates of global peatland extent surpass 4.5 million km
2 [
4], with over 1 million km
2 in both Europe [
87] and Canada [
3], respectively, based on compilations of national datasets. With the increasing global archives of Landsat and Sentinel-2 imagery, and planetary scale computing platforms, such as Google Earth Engine [
42], the methodologies developed here for NEE could be applied to other northern peatlands without tree cover. Ground measurements of NEE from the eddy covariance method should be incorporated to validate the models if applied elsewhere. Global networks such as FLUXNET [
88] collect multitemporal data of CO
2 exchange that could be used as model training and validation. Protocols being established for satellite-based Land Data Product Validation [
89] will allow for greater intercomparability between sites. In the absence of airborne or UAV-based hyperspectral imagery, data from planned spaceborne hyperspectral missions could be investigated to determine their utility for predicting WT depth, provided sufficient
in situ data are available to validate the results, especially for WT depth outside the range examined here.
Climate change affects peatlands directly [
90,
91]. One of the most important drivers of change in the functioning of northern peatlands is the WT depth. Even modest decreases in water table depth coupled with increased air temperature have been shown to lead to profound changes in the proportional contributions of mosses and vascular plants to biomass production [
91]. In addition, drought conditions with a lowering of the WT have recently been shown to result in an increase in higher ecosystem respiration, due in part to changes in vegetation composition where vascular plants replaced
Sphagnum mosses [
90]. Remotely sensed data offer a reliable means to monitor such changes on an ongoing basis.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












