
Spatiotemporal Pattern of Invasive Pedicularis in the Bayinbuluke Land, China, during 2019–2021: An Analysis Based on PlanetScope and Sentinel-2 Data

Abstract
:1. Introduction
- (1)
- Assessing the accuracy of PUL on predictions of the poisonous species Pedicularis;
- (2)
- Comparing the efficacy of Sentinel-2 and PlanetScope satellite imagery in the identification of Pedicularis;
- (3)
- Generating precise distribution maps of Pedicularis with time-series data for subsequent spatiotemporal analysis to support the conservation of the Bayinbuluke Grassland ecosystem through time-series remote sensing dynamic monitoring.
2. Materials and Methods
2.1. Study Area
2.2. Data Sources
2.2.1. UAV RGB Imagery
2.2.2. Sentinel-2 Imagery
2.2.3. PlanetScope Imagery
2.3. Datasets and Data Analysis
2.3.1. Generation of Additional Features
2.3.2. Construction of the Datasets
2.4. Methodology
2.4.1. Classification Method
2.4.2. Accuracy Assessment
2.4.3. Change Detection
3. Results
3.1. Comparison of Classification Accuracy on Sentinel-2 and PlanetScope
3.2. Changes in Dynamics of Pedicularis
4. Discussion
4.1. Influencing Factors of Classification Accuracy
4.2. Spatiotemporal Pattern of Pedicularis
4.3. A Case of Pedicularis Eradication
5. Conclusions
- (1)
- The proliferation of Pedicularis in the Bayinbuluke Grassland has resulted in significant ecological damage, necessitating the substantial expenditure of resources and efforts by the provincial government for rehabilitation efforts. In addressing this issue, change-detection methods utilizing remote sensing technology offer a practical approach for informed management and mitigation. Sentinel-2 images have the advantages of a large width, easily acquirable data, and high accuracy in extracting Pedicularis. The resolution of PlanetScope is higher than that of Sentinel-2, which is more advantageous when removing Pedicularis from small areas. The results of the study show that the PUL method is able to achieve a high recognition accuracy across different images.
- (2)
- Within the confines of the same sensor platform, the influence of feature count on improvements to the identification accuracy becomes obvious with an ample sample size, as evidenced by an increasing feature count coinciding with increased recognition accuracy. However, within an equivalent feature framework, the correlation between resolution elevation and accuracy enhancement does not invariably hold, implying that the resultant classification outcome is dependent on the inherent data quality obtained using the sensor apparatus.
- (3)
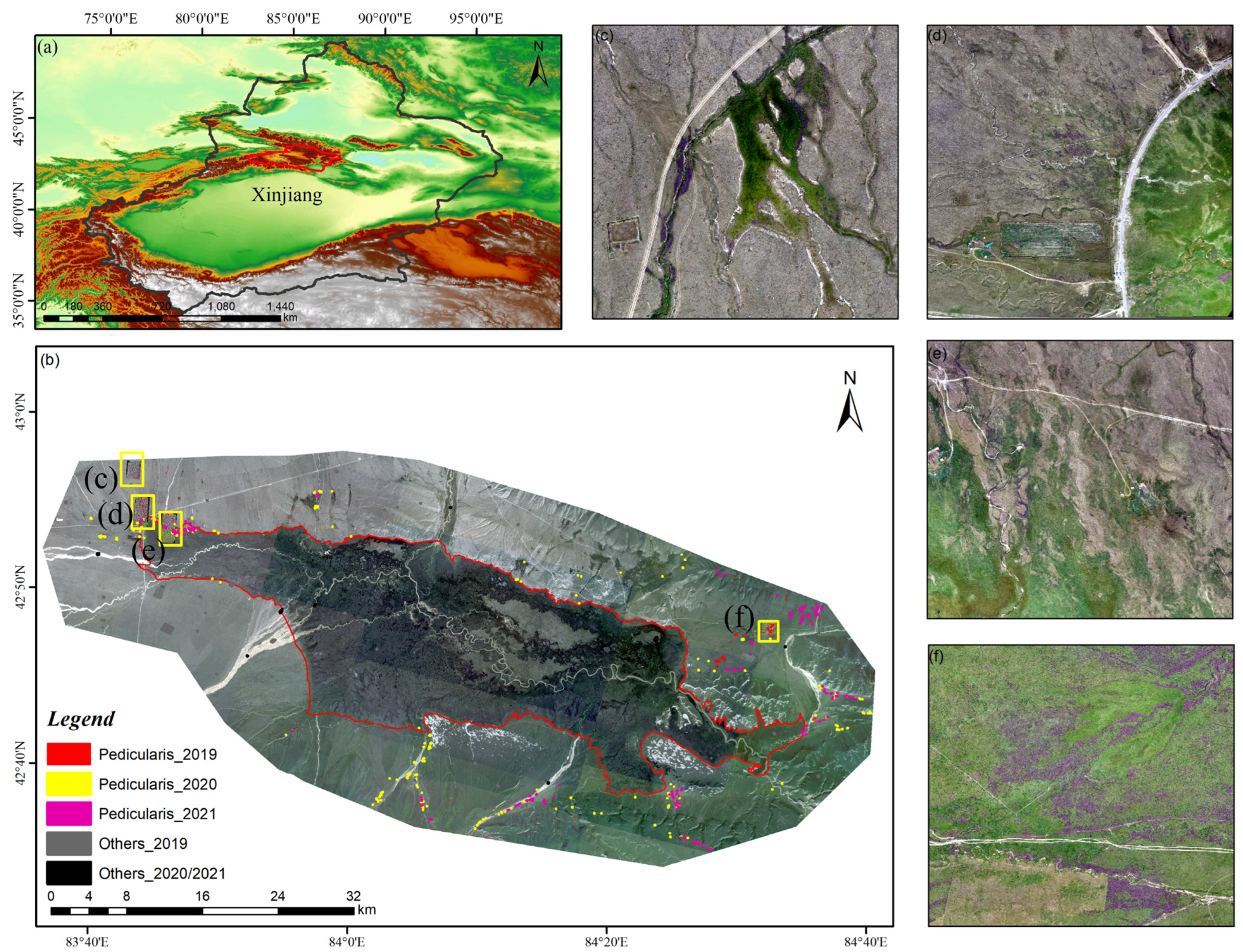
- The post-classification comparison algorithm avoids spectral differences in remote sensing images, especially long-time-series images from different sensors. It enables the rapid monitoring of regional variations in the distribution of different land types. However, it is highly dependent on the stability of the model, and a transferred, high-accuracy classification model needs to be further developed. The distribution of Pedicularis is concentrated in the northwestern and southwestern parts of Bayinbuluke Swan Lake. From 2019 to 2021, the distribution area of Pedicularis exhibited a fluctuating trend, initially increasing and then subsequently decreasing, with the 2021 area measuring 157.2063 km2. Despite better eradication efforts in the northeast region, the distribution area of Pedicularis did not exhibit significant changes, indicating that grassland managers may not have done enough to control the growth of Pedicularis.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fu, P.; Yao, J.; Hu, J.; Guo, X. Capital Endowments, Policy Perceptions and Herdsmen’s Willingness to Reduce Livestock:A Case Study from the World Natural Heritage Site of Bayinbuluke. Acta Agrestia Sin. 2021, 29, 780–787. [Google Scholar]
- Hameed, A.; Zafar, M.; Ahmad, M.; Sultana, S.; Bahadur, S.; Anjum, F.; Shuaib, M.; Taj, S.; Irm, M.; Altaf, M.A. Chemo-taxonomic and biological potential of highly therapeutic plant Pedicularis groenlandica Retz. using multiple microscopic techniques. Microsc. Res. Tech. 2021, 84, 2890–2905. [Google Scholar] [CrossRef] [PubMed]
- Elkind, K.; Sankey, T.T.; Munson, S.M.; Aslan, C.E. Invasive buffelgrass detection using high-resolution satellite and UAV imagery on Google Earth Engine. Remote Sens. Ecol. Conserv. 2019, 5, 318–331. [Google Scholar] [CrossRef]
- Sui, X.; Li, A.; Guan, K. Impacts of climatic changes as well as seed germination characteristics on the population expansion of Pedicularis verticillata. Ecol. Environ. Sci. 2013, 22, 1099–1104. [Google Scholar]
- Yanyan, L.I.U.; Yukun, H.U.; Jianmei, Y.U.; Kaihui, L.I.; Guogang, G.A.O.; Xin, W. Study on Harmfulness of Pedicularis myriophylla and Its Control Measures. Arid Zone Res. 2008, 25, 778–782. [Google Scholar]
- Hongtao, J.I.A.; Pingan, J.; Luming, C.; Chengyi, Z.; Yukun, H.U. Estimation of Organic Carbon Storage of Bayinbuluke Alpine Grassland Ecosystem. Xinjiang Agric. Sci. 2006, 43, 480–483. [Google Scholar]
- Pingan, J.; Hui, L.I.; Hongtao, J.I.A.; Zihong, D. Impacts of fencing on soil animals diversity beneath mountainous lawn vegetation in Bayinbuluke. J. Northwest Sci-Tech Univ. Agric. For. 2007, 35, 69–74. [Google Scholar]
- Choudhary, K.; Boori, M.S.; Kupriyanov, A. Landscape Analysis through Remote Sensing and GIS Techniques: A Case Study of Astrakhan, Russia. In Proceedings of the 8th International Conference on Graphic and Image Processing (ICGIP), Tokyo, Japan, 29–31 October 2016. [Google Scholar]
- Gao, S.; Zheng, J.; Ma, T.; Wu, J.; Nasongcaoketu; Maidi, K. Research on the Applicability of Remote Sensing Monitoring of Inedible Grass Pedicularis sp. by GF-1 WFV Satellite in Bayanbulak Grassland. Xinjiang Agric. Sci. 2017, 54, 1949–1956. [Google Scholar]
- Huber, N.; Ginzler, C.; Pazur, R.; Descombes, P.; Baltensweiler, A.; Ecker, K.; Meier, E.; Price, B. Countrywide classification of permanent grassland habitats at high spatial resolution. Remote Sens. Ecol. Conserv. 2023, 9, 133–151. [Google Scholar] [CrossRef]
- Lopatin, J.; Dolos, K.; Kattenborn, T.; Fassnacht, F.E. How canopy shadow affects invasive plant species classification in high spatial resolution remote sensing. Remote Sens. Ecol. Conserv. 2019, 5, 302–317. [Google Scholar] [CrossRef]
- Kakembo, V.; Smith, J.; Kerley, G. A Temporal Analysis of Elephant-Induced Thicket Degradation in Addo Elephant National Park, Eastern Cape, South Africa. Rangel. Ecol. Manag. 2015, 68, 461–469. [Google Scholar] [CrossRef]
- Singh, P.S.; Singh, V.P.; Pandey, M.K.; Karthikeyan, S.; IEEE. One-class Classifier Ensemble based Enhanced Semisupervised Classification of Hyperspectral Remote Sensing Images. In Proceedings of the 2nd IEEE International Conference on Emerging Smart Computing and Informatics (ESCI), All India Shri Shivaji Memorial Soc, Inst Informat Technol, Pune, India, 12–14 March 2020; pp. 22–27. [Google Scholar]
- Hossain, M.A.; Jia, X.; Benediktsson, J.A. One-Class Oriented Feature Selection and Classification of Heterogeneous Remote Sensing Images. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2016, 9, 1606–1612. [Google Scholar] [CrossRef]
- Li, W.; Guo, Q.; Elkan, C. A Positive and Unlabeled Learning Algorithm for One-Class Classification of Remote-Sensing Data. IEEE Trans. Geosci. Remote Sens. 2011, 49, 717–725. [Google Scholar] [CrossRef]
- Mack, B.; Roscher, R.; Waske, B. Can I Trust My One-Class Classification? Remote Sens. 2014, 6, 8779–8802. [Google Scholar] [CrossRef]
- Li, W.K.; Guo, Q.H. A maximum entropy approach to one-class classification of remote sensing imagery. Int. J. Remote Sens. 2010, 31, 2227–2235. [Google Scholar] [CrossRef]
- Braun, A.C. Evaluation of One-Class Svm for Pixel-Based and Segment-Based Classification in Remote Sensing. In Proceedings of the ISPRS-Technical-Commission III Symposium on Photogrammetric Computer Vision and Image Analysis (PCV), Saint Mande, France, 1–3 September 2010; pp. 160–165. [Google Scholar]
- Chang, S.; Du, B.; Zhang, L. A Subspace Selection-Based Discriminative Forest Method for Hyperspectral Anomaly Detection. IEEE Trans. Geosci. Remote Sens. 2020, 58, 4033–4046. [Google Scholar] [CrossRef]
- Dambros, C.S.; Morais, J.W.; Azevedo, R.A.; Gotelli, N.J. Isolation by distance, not rivers, control the distribution of termite species in the Amazonian rain forest. Ecography 2017, 40, 1242–1250. [Google Scholar] [CrossRef]
- Wang, W.; Tang, J.; Zhang, N.; Xu, X.; Zhang, A.; Wang, Y. Automated Detection Method to Extract Pedicularis Based on UAV Images. Drones 2022, 6, 399. [Google Scholar] [CrossRef]
- Morshed, N.; Yorke, C.; Zhang, Q. Urban Expansion Pattern and Land Use Dynamics in Dhaka, 1989–2014. Prof. Geogr. 2017, 69, 396–411. [Google Scholar] [CrossRef]
- Franklin, S.E.; Ahmed, O.S.; Wulder, M.A.; White, J.C.; Hermosilla, T.; Coops, N.C. Large Area Mapping of Annual Land Cover Dynamics Using Multitemporal Change Detection and Classification of Landsat Time Series Data. Can. J. Remote Sens. 2015, 41, 293–314. [Google Scholar] [CrossRef]
- Angulo, D.; Angulo, F.; Olivar, G. Dynamics and Forecast in a Simple Model of Sustainable Development for Rural Populations. Bull. Math. Biol. 2015, 77, 368–389. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.J.; Luo, J.C.; Zhou, Y.N.; Wang, C.P.; Xi, J.B.; Fang, J.W. Geo-Object-Based Land Cover Map Update for High-Spatial-Resolution Remote Sensing Images via Change Detection and Label Transfer. Remote Sens. 2020, 12, 174. [Google Scholar] [CrossRef]
- Yu, W.J.; Zhou, W.Q.; Qian, Y.G.; Yan, J.L. A new approach for land cover classification and change analysis: Integrating backdating and an object-based method. Remote Sens. Environ. 2016, 177, 37–47. [Google Scholar] [CrossRef]
- Chen, X.H.; Chen, J.; Shi, Y.S.; Yamaguchi, Y. An automated approach for updating land cover maps based on integrated change detection and classification methods. ISPRS-J. Photogramm. Remote Sens. 2012, 71, 86–95. [Google Scholar] [CrossRef]
- Qian, Y.G.; Zhou, W.Q.; Yu, W.J.; Han, L.J.; Li, W.F.; Zhao, W.H. Integrating Backdating and Transfer Learning in an Object-Based Framework for High Resolution Image Classification and Change Analysis. Remote Sens. 2020, 12, 4094. [Google Scholar] [CrossRef]
- Xu, Y.D.; Yu, L.; Zhao, F.R.; Cai, X.L.; Zhao, J.Y.; Lu, H.; Gong, P. Tracking annual cropland changes from 1984 to 2016 using time-series Landsat images with a change-detection and post-classification approach: Experiments from three sites in Africa. Remote Sens. Environ. 2018, 218, 13–31. [Google Scholar] [CrossRef]
- Feng, X.; Li, P.; Cheng, T. Detection of Urban Built-Up Area Change From Sentinel-2 Images Using Multiband Temporal Texture and One-Class Random Forest. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2021, 14, 6974–6986. [Google Scholar] [CrossRef]
- Kempeneers, P.; Sedano, F.; Strobl, P.; McInerney, D.O.; San-Miguel-Ayanz, J. Increasing Robustness of Postclassification Change Detection Using Time Series of Land Cover Maps. IEEE Trans. Geosci. Remote Sens. 2012, 50, 3327–3339. [Google Scholar] [CrossRef]
- Crowson, M.; Hagensieker, R.; Waske, B. Mapping land cover change in northern Brazil with limited training data. Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 202–214. [Google Scholar] [CrossRef]
- Colditz, R.R.; Acosta-Velazquez, J.; Diaz Gallegos, J.R.; Vazquez Lule, A.D.; Teresa Rodriguez-Zuniga, M.; Maeda, P.; Cruz Lopez, M.I.; Ressl, R. Potential effects in multi-resolution post-classification change detection. Int. J. Remote Sens. 2012, 33, 6426–6445. [Google Scholar] [CrossRef]
- Bao, A.; Cao, X.; Chen, X.; Xia, Y. Study on Models for Monitoring of Aboveground Biomass about Bayinbuluke grassland Assisted by Remote Sensing. In Proceedings of the Conference on Remote Sensing and Modeling of Ecosystems for Sustainability, San Diego, CA, USA, 13 August 2008. [Google Scholar]
- Chen, X.; Yang, Z.; Wang, T.; Han, F. Landscape Ecological Risk and Ecological Security Pattern Construction in World Natural Heritage Sites: A Case Study of Bayinbuluke, Xinjiang, China. ISPRS Int. J. Geo-Inf. 2022, 11, 328. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; Zhu, X.; Jia, H.; Han, D. Landscape Pattern Changes in AlpineWetland of Bayanbulak Swan Lake during 1996–2015. J. Nat. Resour. 2018, 33, 1897–1911. [Google Scholar]
- Liu, Q.; Yang, Z.P.; Han, F.; Shi, H.; Wang, Z.; Chen, X.D. Ecological Environment Assessment in World Natural Heritage Site Based on Remote-Sensing Data. A Case Study from the Bayinbuluke. Sustainability 2019, 11, 6385. [Google Scholar] [CrossRef]
- Grabska, E.; Frantz, D.; Ostapowicz, K. Evaluation of machine learning algorithms for forest stand species mapping using Sentinel-2 imagery and environmental data in the Polish Carpathians. Remote Sens. Environ. 2020, 251, 112103. [Google Scholar] [CrossRef]
- De Vroey, M.; de Vendictis, L.; Zavagli, M.; Bontemps, S.; Heymans, D.; Radoux, J.; Koetz, B.; Defourny, P. Mowing detection using Sentinel-1 and Sentinel-2 time series for large scale grassland monitoring. Remote Sens. Environ. 2022, 280, 113145. [Google Scholar] [CrossRef]
- Kimm, H.; Guan, K.Y.; Jiang, C.Y.; Peng, B.; Gentry, L.F.; Wilkin, S.C.; Wang, S.B.; Cai, Y.P.; Bernacchi, C.J.; Peng, J.; et al. Deriving high-spatiotemporal-resolution leaf area index for agroecosystems in the US Corn Belt using Planet Labs CubeSat and STAIR fusion data. Remote Sens. Environ. 2020, 239, 111615. [Google Scholar] [CrossRef]
- Holloway-Brown, J.; Helmstedt, K.J.; Mengersen, K.L. Interpolating missing land cover data using stochastic spatial random forests for improved change detection. Remote Sens. Ecol. Conserv. 2021, 7, 649–665. [Google Scholar] [CrossRef]
- Saber, A.; El-Sayed, I.; Rabah, M.; Selim, M. Evaluating change detection techniques using remote sensing data: Case study New Administrative Capital Egypt. Egypt. J. Remote Sens. Space Sci. 2021, 24, 635–648. [Google Scholar] [CrossRef]
- Gao, S.; Lin, J.; Ma, T.; Wu, J.; Zheng, J. Extraction and Analysis of Hyperspectral Data and Characteristics fromPedicularis on Bayanbulak Grassland in Xinjiang. Remote Sens. Technol. Appl. 2018, 33, 908–914. [Google Scholar]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Gnyp, M.L.; Miao, Y.X.; Yuan, F.; Ustin, S.L.; Yu, K.; Yao, Y.K.; Huang, S.Y.; Bareth, G. Hyperspectral canopy sensing of paddy rice aboveground biomass at different growth stages. Field Crops Res. 2014, 155, 42–55. [Google Scholar] [CrossRef]
- Wang, D.; Cui, B.C.; Duan, S.S.; Chen, J.J.; Fan, H.; Lu, B.B.; Zheng, J.H. Moving north in China: The habitat of Pedicularis kansuensis in the context of climate change. Sci. Total Environ. 2019, 697, 133979. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.; Jiang, H.; Crow, W.T.; Liu, S.; Zhao, S.; Liu, J.; Yang, S. Estimating Corn Canopy Water Content From Normalized Difference Water Index (NDWI): An Optimized NDWI-Based Scheme and Its Feasibility for Retrieving Corn VWC. IEEE Trans. Geosci. Remote Sens. 2021, 59, 8168–8181. [Google Scholar] [CrossRef]
- Francini, S.; McRoberts, R.E.; Giannetti, F.; Mencucci, M.; Marchetti, M.; Mugnozza, G.S.; Chirici, G. Near-real time forest change detection using PlanetScope imagery. Eur. J. Remote Sens. 2020, 53, 233–244. [Google Scholar] [CrossRef]
- Sui, Y.; Shao, F.; Wang, C.; Sun, R.; Ji, J. Complex network modeling of spectral remotely sensed imagery: A case study of massive green algae blooms detection based on MODIS data. Phys. A-Stat. Mech. Its Appl. 2016, 464, 138–148. [Google Scholar] [CrossRef]
- Zhao, B.; Yang, F.; Zhang, R.; Shen, J.; Pilz, J.; Zhang, D. Application of unsupervised learning of finite mixture models in ASTER VNIR data-driven land use classification. J. Spat. Sci. 2021, 66, 89–112. [Google Scholar] [CrossRef]
- Zhou, X.-X.; Li, Y.-Y.; Luo, Y.-K.; Sun, Y.-W.; Su, Y.-J.; Tan, C.-W.; Liu, Y.-J. Research on remote sensing classification of fruit trees based on Sentinel-2 multi-temporal imageries. Sci. Rep. 2022, 12, 11549. [Google Scholar] [CrossRef]
- Mordelet, F.; Vert, J.P. A bagging SVM to learn from positive and unlabeled examples. Pattern Recognit. Lett. 2014, 37, 201–209. [Google Scholar] [CrossRef]
- Halligan, S.; Altman, D.G.; Mallett, S. Disadvantages of using the area under the receiver operating characteristic curve to assess imaging tests: A discussion and proposal for an alternative approach. Eur. Radiol. 2015, 25, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zheng, S.; Wang, X. The Spatiotemporal Changes and the Impacts of Climate Factors on Grassland in the Northern Songnen Plain (China). Sustainability 2021, 13, 6568. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, X.; Xie, F. Macro analysis of spatiotemporal variations in ecosystems from 1996 to 2016 in Xishuangbanna in Southwest China. Environ. Sci. Pollut. Res. 2021, 28, 40192–40202. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Kong, L.; Ouyang, Z. Characteristics and Driving Mechanism of Regional Ecosystem Assets Change in the Process of Rapid Urbanization—A Case Study of the Beijing–Tianjin–Hebei Urban Agglomeration. Remote Sens. 2022, 14, 5747. [Google Scholar] [CrossRef]
- Fan, C.; Chen, X.; Zhong, L.; Zhou, M.; Shi, Y.; Duan, Y. Improved Wallis Dodging Algorithm for Large-Scale Super-Resolution Reconstruction Remote Sensing Images. Sensors 2017, 17, 623. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, Q.; Zhang, Y.; Wang, H.; Du, X. Normalized NDVI valley area index (NNVAI)-based framework for quantitative and timely monitoring of winter wheat frost damage on the Huang-Huai-Hai Plain, China. Agric. Ecosyst. Environ. 2020, 292, 106793. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, D. A robust and adaptive spatial-spectral fusion model for PlanetScope and Sentinel-2 imagery. Gisci. Remote Sens. 2022, 59, 520–546. [Google Scholar] [CrossRef]
- Ye, N.; Morgenroth, J.; Xu, C.; Chen, N. Indigenous forest classification in New Zealand-A comparison of classifiers and sensors. Int. J. Appl. Earth Obs. Geoinf. 2021, 102, 102395. [Google Scholar] [CrossRef]
- Wang, Q.; Zhai, P.-M.; Qin, D.-H. New perspectives on ‘warming–wetting’trend in Xinjiang, China. Adv. Clim. Chang. Res. 2020, 11, 252–260. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band Name | Sentinel-2A/Sentinel-2B Central Wavelength (nm) | Resolution (Meters) |
|---|---|---|
| Band 1—Coastal aerosol | 443.9/442.2 | 60 |
| Band 2—Blue | 496.6/492.1 | 10 |
| Band 3—Green | 560.0/559.0 | 10 |
| Band 4—Red | 664.5/664.9 | 10 |
| Band 5—Vegetation red edge | 703.9/703.8 | 20 |
| Band 6—Vegetation red edge | 740.2/739.1 | 20 |
| Band 7—Vegetation red edge | 782.5/779.7 | 20 |
| Band 8—NIR | 835.1/832.9 | 10 |
| Band 8A—Narrow NIR | 864.8/864.0 | 20 |
| Band 9—Water Vapor | 945.0/943.2 | 60 |
| Band 10—SWIR–Cirrus | 1373.5/1376.9 | 60 |
| Band 11—SWIR | 1613.7/1610.4 | 20 |
| Band 12—SWIR | 2202.4/2185.7 | 20 |
| Band Name | Spatial Resolution (m) | Spectral Wavelength (nm) |
|---|---|---|
| Blue | 3.0 | 464–517 |
| Green | 547–585 | |
| Red | 650–682 | |
| NIR | 846–888 |
| Training Samples (Pixels) | Test Samples (Pixels) | ||||
|---|---|---|---|---|---|
| Pedicularis | Others | Pedicularis | Others | ||
| 2019 | Sentinel-2 data (7/13 features) | 7690 | 7690 | 1923 | 5598 |
| PlanetScope data (7 features) | 62,063 | 62,063 | 15,515 | 455,615 | |
| 2020 | Sentinel-2 data (7/13 features) | 1943 | 6477 | 486 | 1900 |
| PlanetScope data (7 features) | 15,568 | 51,893 | 15,515 | 15,434 | |
| 2021 | Sentinel-2 data (7/13 features) | 2395 | 7983 | 599 | 1900 |
| PlanetScope data (7 features) | 19,520 | 65,066 | 4880 | 15,434 | |
| Year | Datasets | Types | Metrics | |||
|---|---|---|---|---|---|---|
| Recall | Precision | Accuracy | F1-Score | |||
| 2019 | Sentinel-2 data (7 features) | Pedicularis | 0.9212 | 0.9286 | 0.9617 | 0.9248 |
| Others | 0.9757 | 0.9730 | ||||
| Sentinel-2 data (13 features) | Pedicularis | 0.9278 | 0.9536 | 0.9700 | 0.9405 | |
| Others | 0.9845 | 0.9754 | ||||
| PlanetScope data (7 features) | Pedicularis | 0.8458 | 0.6042 | 0.8678 | 0.7049 | |
| Others | 0.8728 | 0.9610 | ||||
| 2020 | Sentinel-2 data (7 features) | Pedicularis | 0.8861 | 0.7307 | 0.9169 | 0.8009 |
| Others | 0.9241 | 0.9721 | ||||
| Sentinel-2 data (13 features) | Pedicularis | 0.8710 | 0.9045 | 0.9583 | 0.8874 | |
| Others | 0.9786 | 0.9702 | ||||
| PlanetScope data (7 features) | Pedicularis | 0.8340 | 0.6132 | 0.8708 | 0.7067 | |
| Others | 0.8793 | 0.9584 | ||||
| 2021 | Sentinel-2 data (7 features) | Pedicularis | 0.8971 | 0.8629 | 0.9454 | 0.8796 |
| Others | 0.9591 | 0.9703 | ||||
| Sentinel-2 data (13 features) | Pedicularis | 0.8864 | 0.9277 | 0.9593 | 0.9065 | |
| Others | 0.9802 | 0.9678 | ||||
| PlanetScope data (7 features) | Pedicularis | 0.8985 | 0.7399 | 0.9313 | 0.8115 | |
| Others | 0.9377 | 0.9791 | ||||
| Year | Area (km2) | Area Ratio (%) |
|---|---|---|
| 2019 | 195.7803 | 5.55% |
| 2020 | 124.9584 | 3.54% |
| 2021 | 157.2063 | 4.46% |
| 2019–2020 | 2020–2021 | 2019–2021 | ||||
|---|---|---|---|---|---|---|
| Pedicularis | Others | Pedicularis | Others | Pedicularis | Others | |
| Pedicularis | 21.8880 | 173.8923 | 18.9476 | 106.0108 | 33.6437 | 162.1330 |
| Others | 103.0704 | 3225.0944 | 138.2587 | 3260.7280 | 123.5590 | 3204.6058 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Tang, J.; Zhang, N.; Wang, Y.; Xu, X.; Zhang, A. Spatiotemporal Pattern of Invasive Pedicularis in the Bayinbuluke Land, China, during 2019–2021: An Analysis Based on PlanetScope and Sentinel-2 Data. Remote Sens. 2023, 15, 4383. https://doi.org/10.3390/rs15184383
Wang W, Tang J, Zhang N, Wang Y, Xu X, Zhang A. Spatiotemporal Pattern of Invasive Pedicularis in the Bayinbuluke Land, China, during 2019–2021: An Analysis Based on PlanetScope and Sentinel-2 Data. Remote Sensing. 2023; 15(18):4383. https://doi.org/10.3390/rs15184383
Chicago/Turabian StyleWang, Wuhua, Jiakui Tang, Na Zhang, Yanjiao Wang, Xuefeng Xu, and Anan Zhang. 2023. "Spatiotemporal Pattern of Invasive Pedicularis in the Bayinbuluke Land, China, during 2019–2021: An Analysis Based on PlanetScope and Sentinel-2 Data" Remote Sensing 15, no. 18: 4383. https://doi.org/10.3390/rs15184383