
Landscape-Level Associations of Wintering Waterbird Diversity and Abundance from Remotely Sensed Wetland Characteristics of Poyang Lake


Abstract
:
1. Introduction
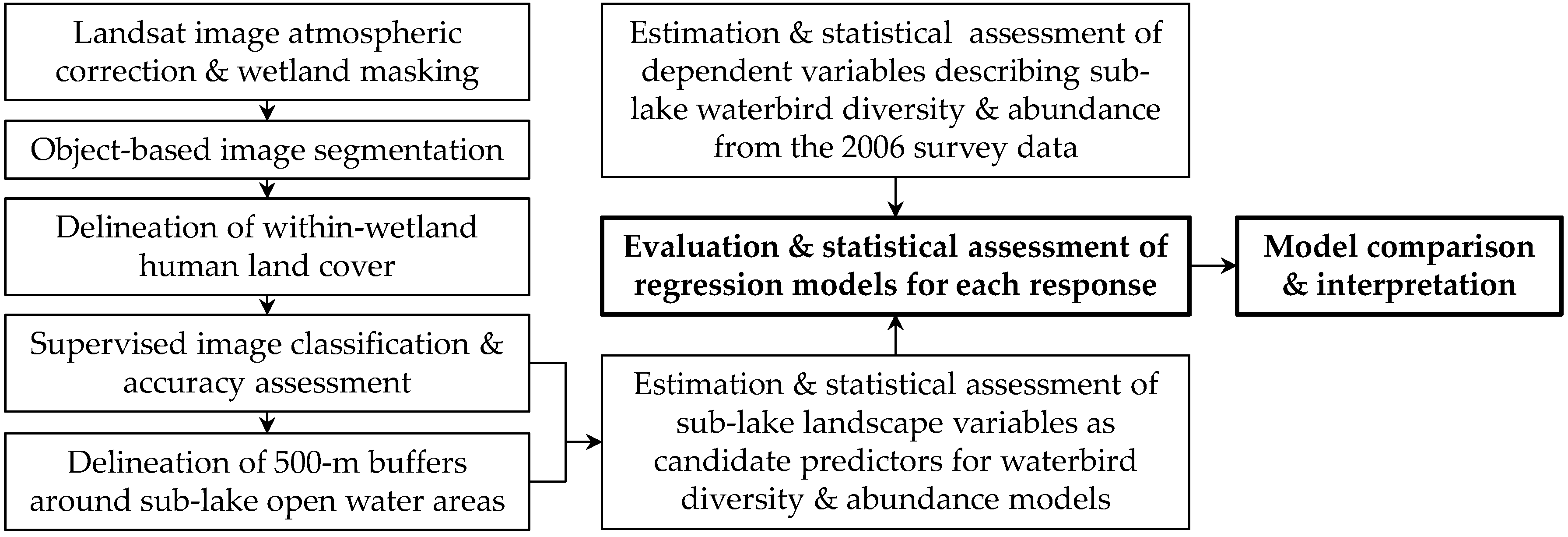
2. Materials and Methods
2.1. Study Area
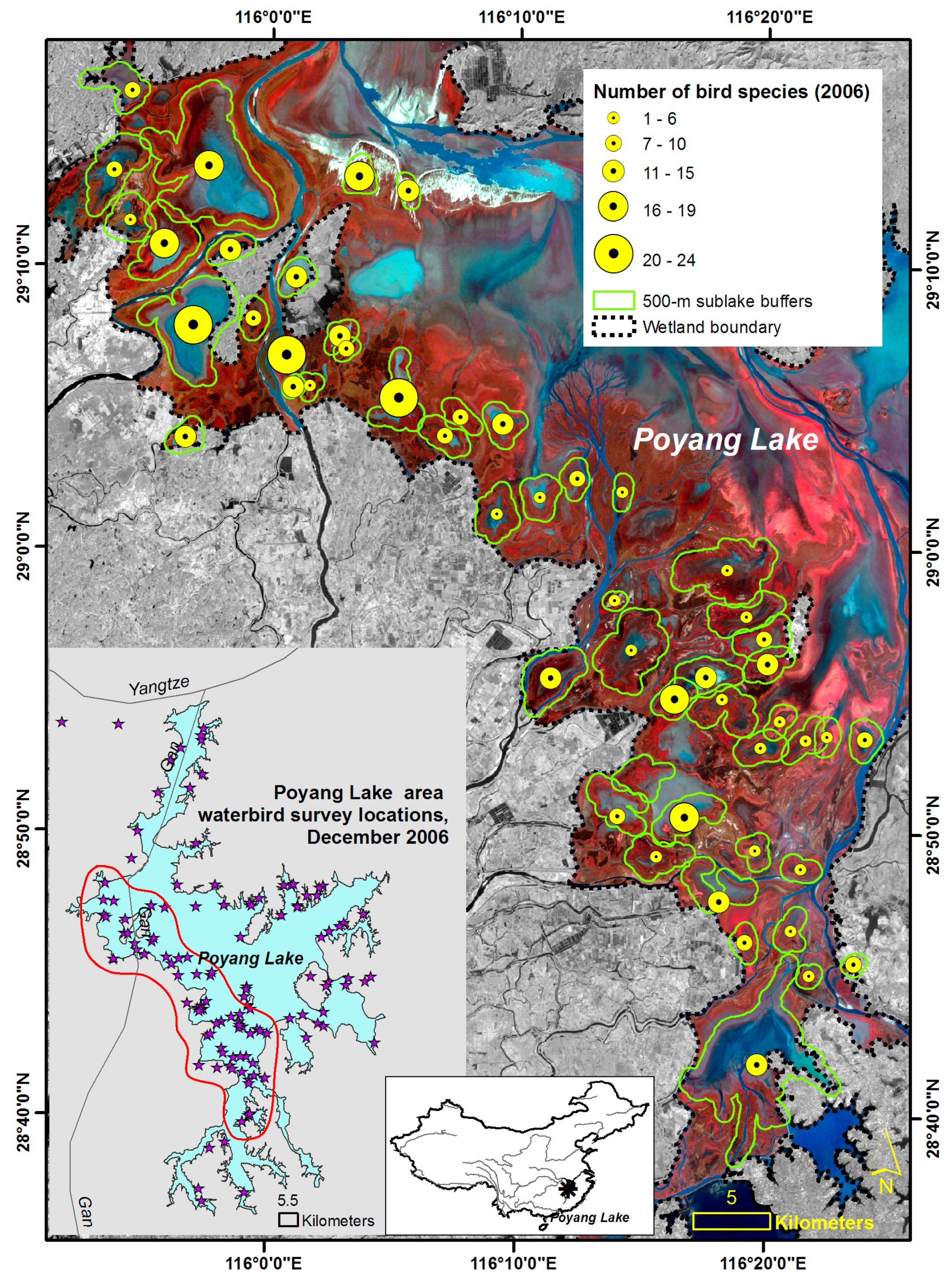
2.2. Waterbird Survey of 2006 and Dependent Variables
2.3. Independent Variables Based on Remote Sensing Data
2.4. Model Selection and Diagnostics for Spatial Autocorrelation
3. Results
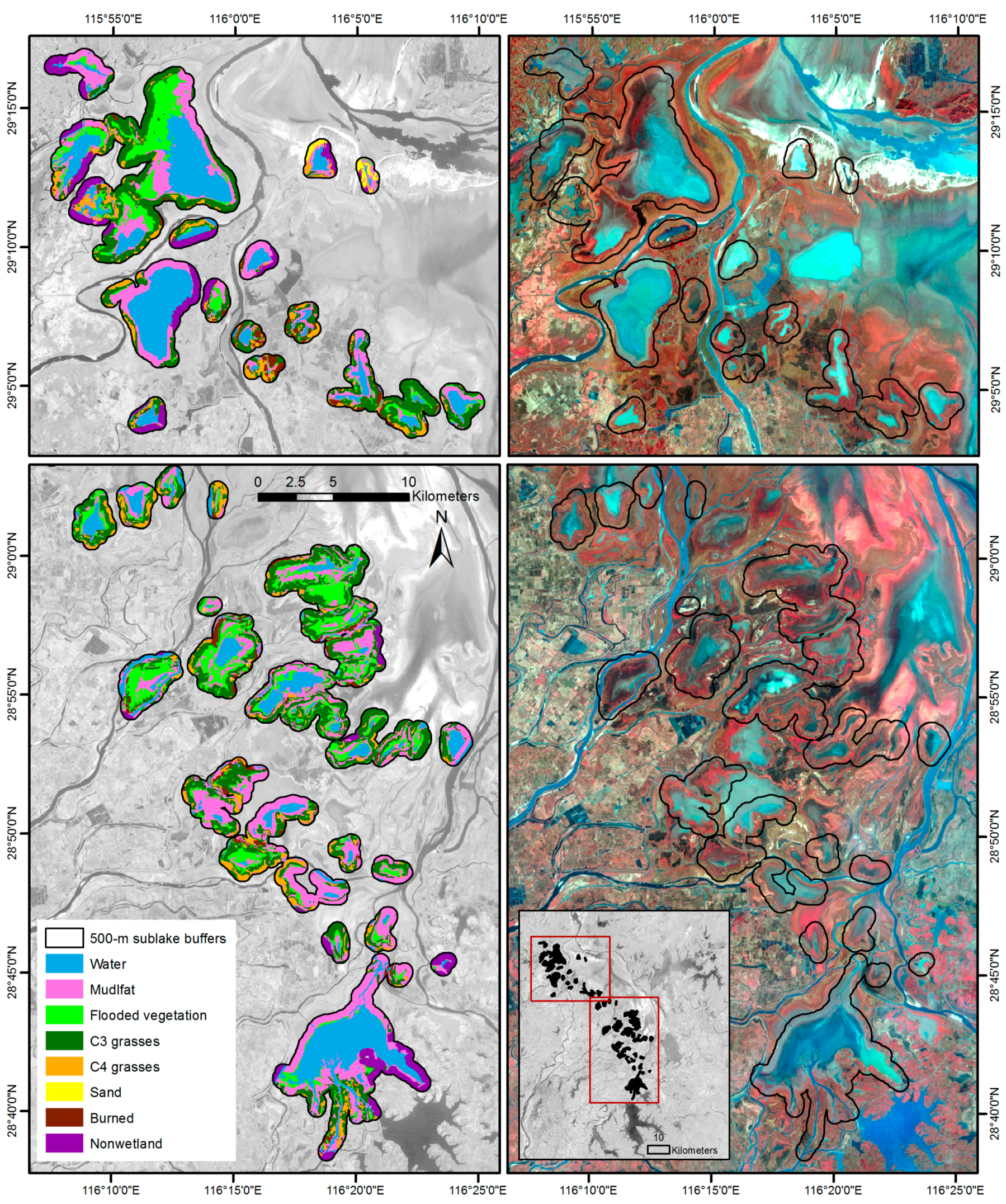
3.1. Image Classification Results
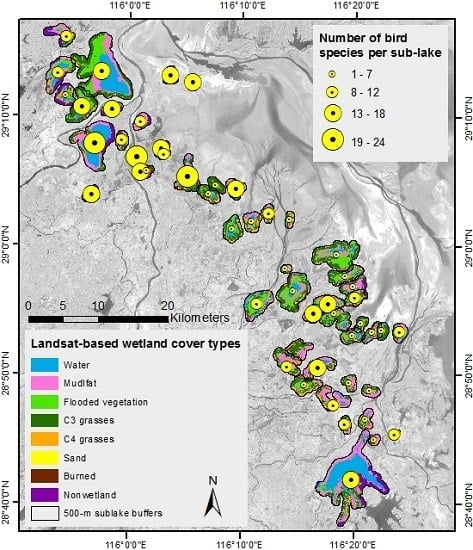
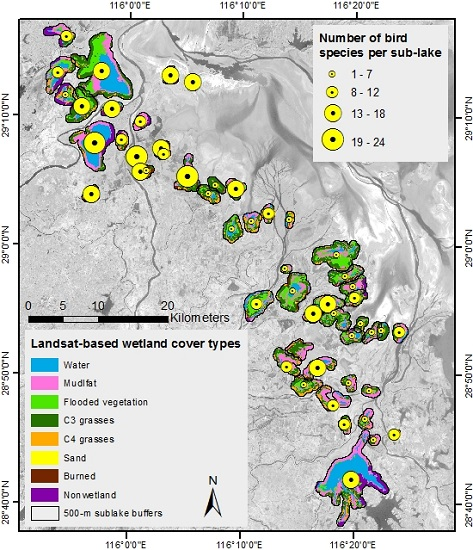
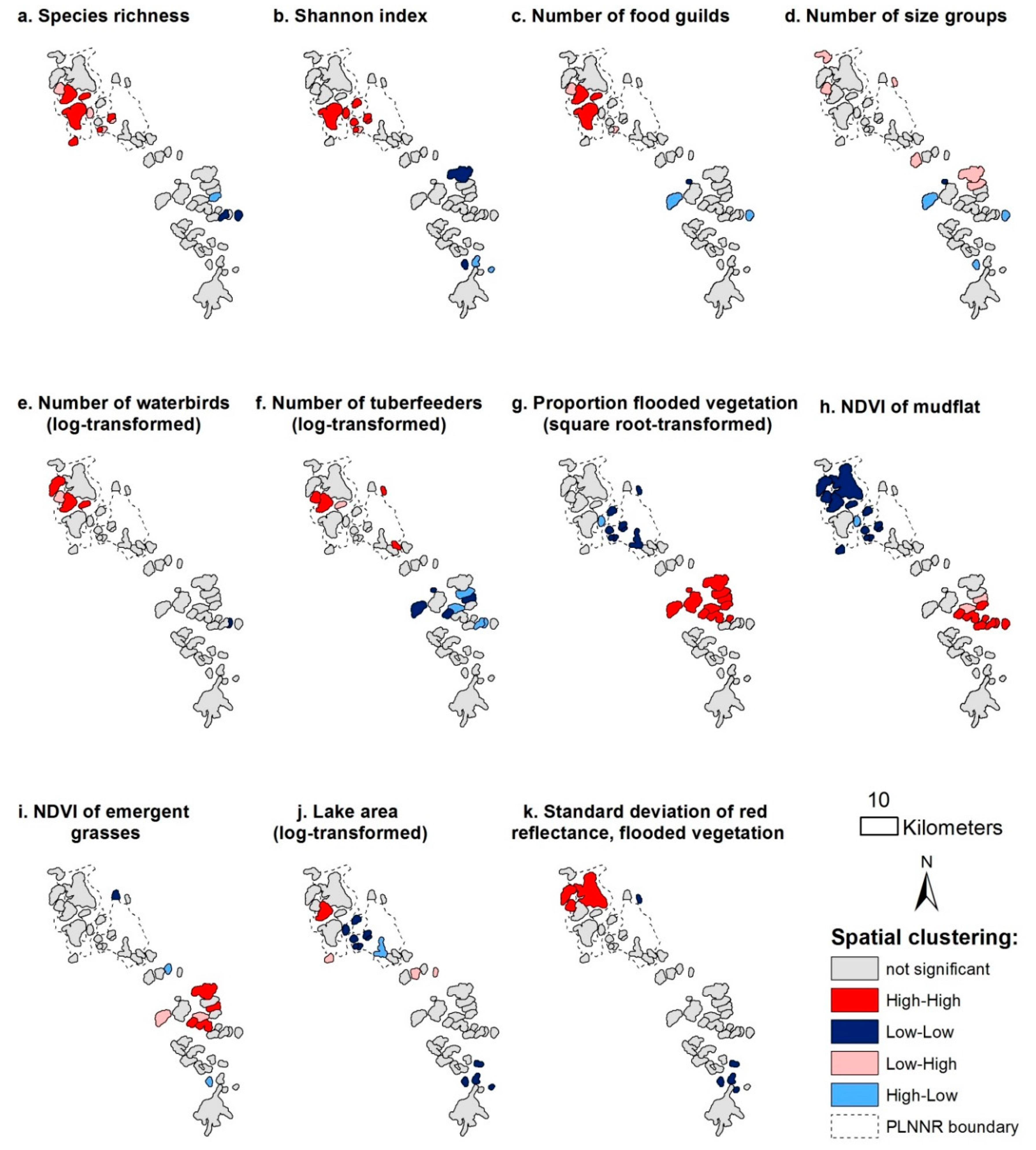
3.2. Spatial Patterns of Response and Predictor Variables
3.3. Variable Selection in Regression Models
3.4. Spatial Autocorrelation in Linear Regression Models
4. Discussion
4.1. Remotely Sensed Indicators of Poyang Lake Bird Diversity and Abundance
4.2. Spatial Autocorrelation in Diversity and Abundance Models
4.3. Uncertainties in Models and Study Limitations
4.4. Implications for Wetland Management and Conservation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
| NDVI | Normalized Difference Vegetation Index |
| OBIA | Object-based image analysis |
| AIC | Akaike Information Criterion |
| PLNNR | Poyang Lake National Nature Reserve |
| ICF | International Crane Foundation |
| OLS | Ordinary least squares regression |
| LISA | Local indicators of spatial autocorrelation |
References
- Fang, J.; Wang, Z.; Zhao, S.; Li, Y.; Tang, Z.; Yu, D.; Ni, L.; Liu, H.; Xie, P.; Da, L.; et al. Biodiversity changes in the lakes of the Central Yangtze. Front. Ecol. Environ. 2006, 4, 369–377. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Leveque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Li, H.; Guan, B.; Zhou, C.; Wang, Z.; Deng, Z.; Zhi, Y.; Liu, Y.; Xu, C.; Fang, S.; et al. China’s natural wetlands: Past problems, current status, and future challenges. AMBIO 2007, 36, 335–342. [Google Scholar] [CrossRef]
- Kirby, J.S.; Stattersfield, A.J.; Butchart, S.H.M.; Evans, M.I.; Grimmett, R.F.A.; Jones, V.R.; O’Sullivan, J.; Tucker, G.M.; Newton, I. Key conservation issues for migratory land- and waterbird species on the world’s major flyways. Bird Conserv. Int. 2008, 18, S49–S73. [Google Scholar] [CrossRef]
- Cao, C.X.; Zhao, J.; Gong, P.; Ma, G.R.; Bao, D.M.; Tian, K.; Tian, R.; Niu, Z.G.; Zhang, H.; Xu, M.; et al. Wetland changes and droughts in southwestern China. Geomat. Nat. Hazards Risk 2012, 3, 79–95. [Google Scholar] [CrossRef]
- De Boer, W.F.; Cao, L.; Barter, M.; Wang, X.; Sun, M.; van Oeveren, H.; de Leeuw, J.; Barzen, J.; Prins, H.H.T. Comparing the Community Composition of European and Eastern Chinese Waterbirds and the Influence of Human Factors on the China Waterbird Community. AMBIO 2011, 40, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Jia, Y.; Saintilan, N.; Wang, Y.; Liu, G.; Lei, G.; Wen, L. Causality between abundance and diversity is weak for wintering migratory waterbirds. Freshw. Biol. 2016, 61, 206–218. [Google Scholar] [CrossRef]
- Wang, Y.; Jia, Y.; Guan, L.; Lu, C.; Lei, G.; Wen, L.; Liu, G. Optimising hydrological conditions to sustain wintering waterbird populations in Poyang Lake National Natural Reserve: Implications for dam operations. Freshw. Biol. 2013, 58, 2366–2379. [Google Scholar] [CrossRef]
- Ramsar Convention the List of Wetlands of International Importance, 2016. The Secretariat of the Convention on Wetlands (Ramsar, Iran, 1971). Available online: http://www.ramsar.org/sites/default/files/documents/library/sitelist.pdf (accessed on 1 February 2016).
- Dronova, I.; Gong, P.; Wang, L. Object-based analysis and change detection of major wetland cover types and their classification uncertainty during the low water period at Poyang Lake, China. Remote Sens. Environ. 2011, 115, 3220–3236. [Google Scholar] [CrossRef]
- Feng, L.; Hu, C.; Chen, X.; Cai, X.; Tian, L.; Gan, W. Assessment of inundation changes of Poyang Lake using MODIS observations between 2000 and 2010. Remote Sens. Environ. 2012, 121, 80–92. [Google Scholar] [CrossRef]
- Zhang, Y.; Jia, Q.; Prins, H.H.T.; Cao, L.; de Boer, W.F. Effect of conservation efforts and ecological variables on waterbird population sizes in wetlands of the Yangtze River. Sci. Rep. 2015, 5, 17136. [Google Scholar] [CrossRef] [PubMed]
- Canepuccia, A.D.; Isacch, J.P.; Gagliardini, D.A.; Escalante, A.H.; Iribarne, O.O. Waterbird response to changes in habitat area and diversity generated by rainfall in a SW Atlantic coastal lagoon. Waterbirds 2007, 30, 541–553. [Google Scholar] [CrossRef]
- Nohr, H.; Jorgensen, A.F. Mapping of biological diversity in Sahel by means of satellite image analyses and ornithological surveys. Biodivers. Conserv. 1997, 6, 545–566. [Google Scholar] [CrossRef]
- Lavers, C.P.; HainesYoung, R.H.; Avery, M.I. The habitat associations of dunlin (Calidris alpina) in the Flow Country of northern Scotland and an improved model for predicting habitat quality. J. Appl. Ecol. 1996, 33, 279–290. [Google Scholar] [CrossRef]
- Moffett, K.B.; Law, J.; Gorelick, S.M.; Nur, N.; Wood, J.K. Alameda Song Sparrow Abundance Related to Salt Marsh Vegetation Patch Size and Shape Metrics Quantified from Remote Sensing Imagery. San Franc. Estuary Watershed Sci. 2014, 12, 2. [Google Scholar] [CrossRef]
- Webb, E.B.; Smith, L.M.; Vrtiska, M.P.; Lagrange, T.G. Effects of Local and Landscape Variables on Wetland Bird Habitat Use during Migration through the Rainwater Basin. J. Wildl. Manag. 2010, 74, 109–119. [Google Scholar] [CrossRef]
- Pap, K.; Nagy, L.; Balogh, C.; G-Toth, L.; Liker, A. Environmental factors shaping the distribution of common wintering waterbirds in a lake ecosystem with developed shoreline. Hydrobiologia 2013, 716, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Fairbairn, S.E.; Dinsmore, J.J. Local and landscape-level influences on wetland bird communities of the prairie pothole region of Iowa, USA. Wetlands 2001, 21, 41–47. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Haskell, J.P. The effect of energy and seasonality on avian species richness and community composition. Am. Nat. 2003, 161, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Bergen, K.M.; Goetz, S.J.; Dubayah, R.O.; Henebry, G.M.; Hunsaker, C.T.; Imhoff, M.L.; Nelson, R.F.; Parker, G.G.; Radeloff, V.C. Remote sensing of vegetation 3-D structure for biodiversity and habitat: Review and implications for lidar and radar spaceborne missions. J. Geophys. Res. Biogeosci. 2009, 114, G00E06. [Google Scholar] [CrossRef]
- Goetz, S.J.; Fiske, G.J.; Bunn, A.G. Using satellite time-series data sets to analyze fire disturbance and forest recovery across Canada. Remote Sens. Environ. 2006, 101, 352–365. [Google Scholar] [CrossRef]
- Hawkins, B.A. Summer vegetation, deglaciation and the anomalous bird diversity gradient in eastern North America. Glob. Ecol. Biogeogr. 2004, 13, 321–325. [Google Scholar] [CrossRef]
- Bino, G.; Levin, N.; Darawshi, S.; Van Der Hal, N.; Reich-Solomon, A.; Kark, S. Accurate prediction of bird species richness patterns in an urban environment using Landsat-derived NDVI and spectral unmixing. Int. J. Remote Sens. 2008, 29, 3675–3700. [Google Scholar] [CrossRef]
- Wood, E.M.; Pidgeon, A.M.; Radeloff, V.C.; Keuler, N.S. Image Texture Predicts Avian Density and Species Richness. PLoS ONE 2013, 8, e63211. [Google Scholar] [CrossRef] [PubMed]
- Culbert, P.D.; Radeloff, V.C.; St-Louis, V.; Flather, C.H.; Rittenhouse, C.D.; Albright, T.P.; Pidgeon, A.M. Modeling broad-scale patterns of avian species richness across the Midwestern United States with measures of satellite image texture. Remote Sens. Environ. 2012, 118, 140–150. [Google Scholar] [CrossRef]
- Rundquist, D.; Narumalani, S.; Narayanan, R. A review of wetlands remote sensing and defining new considerations. Remote Sens. Rev. 2001, 20, 207–226. [Google Scholar] [CrossRef]
- Ozesmi, S.L.; Bauer, M.E. Satellite remote sensing of wetlands. Wetl. Ecol. Manag. 2002, 10, 381–402. [Google Scholar] [CrossRef]
- Tian, B.; Zhou, Y.; Zhang, L.; Yuan, L. Analyzing the habitat suitability for migratory birds at the Chongming Dongtan Nature Reserve in Shanghai, China. Estuar. Coast. Shelf Sci. 2008, 80, 296–302. [Google Scholar] [CrossRef]
- Richmond, O.M.W. Inferring Ecological Relationships from Occupancy Patterns for California Black Rails in the Sierra Nevada Foothills. Ph.D. Thesis, University of California Berkeley, Berkeley, CA, USA, December 2010. [Google Scholar]
- Dronova, I. Object-Based Image Analysis in Wetland Research: A Review. Remote Sens. 2015, 7, 6380–6413. [Google Scholar] [CrossRef]
- Blaschke, T. Object based image analysis for remote sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- Tobler, W. Computer movie simulating urban growth in Detroit region. Econ. Geogr. 1970, 46, 234–240. [Google Scholar] [CrossRef]
- Legendre, P. Spatial autocorrelation—Trouble or new paradigm? Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Anselin, L.; Bera, A.K.; Florax, R.; Yoon, M.J. Simple diagnostic tests for spatial dependence. Reg. Sci. Urban Econ. 1996, 26, 77–104. [Google Scholar] [CrossRef]
- Hoeting, J.A.; Davis, R.A.; Merton, A.A.; Thompson, S.E. Model selection for geostatistical models. Ecol. Appl. 2006, 16, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Bacaro, G.; Santi, E.; Rocchini, D.; Pezzo, F.; Puglisi, L.; Chiarucci, A. Geostatistical modelling of regional bird species richness: Exploring environmental proxies for conservation purpose. Biodivers. Conserv. 2011, 20, 1677–1694. [Google Scholar] [CrossRef]
- Dormann, C.F.; McPherson, J.M.; Araujo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D.; et al. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Miller, J.; Franklin, J.; Aspinall, R. Incorporating spatial dependence in predictive vegetation models. Ecol. Model. 2007, 202, 225–242. [Google Scholar] [CrossRef]
- Barzen, J. Phase 1 Report: How Development Projects May Impact Wintering Waterbirds at Poyang Lake; Unpublished report submitted to Hydro-ecology Institute of the Yangtze Water Resources Commission; International Crane Foundation: Baraboo, WI, USA, 2008; p. 14. Available online: https://www.savingcranes.org/wp-content/uploads/2008/05/Phase%201%20Report_English.pdf (accessed on 2 January 2016).
- The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/ (accessed on 2 January 2016).
- Qian, F.; Yu, C.; Jiang, H. Ground and aerial surveys of wintering waterbirds in Poyang Lake basin. In Proceedings of the UNEP/GEF Siberian Crane Wetland Project—Project Completion Workshop, Harbin, China, 14–15 October 2009; pp. 1–13.
- Burnham, J. Environmental Drivers of Siberian Crane (Grus Leucogeranus) Habitat Selection and Wetland Management and Conservation in China. Master’s Thesis, University of Wisconsin, Madison, WI, USA, December 2007. [Google Scholar]
- Ji, W.; Zeng, N.; Wang, Y.; Gong, P.; Bing, X.; Bao, S. Analysis on the Waterbirds Community Survey of Poyang Lake in Winter. Geogr. Inf. Sci. 2007, 13, 51–64. [Google Scholar] [CrossRef]
- Barzen, J. Phase 2 Report: Potential Impacts of a Water Control Structure on the Abundance and Distribution of Wintering Waterbirds at Poyang Lake; Unpublished Report Submitted to Hydro-Ecology Institute of the Yangtze Water Resources Commission; International Crane Foundation: Baraboo, WI, USA, 2009; p. 54. Available online: https://www.savingcranes.org/wp-content/uploads/2008/05/Phase%202%20Report_English.pdf (accessed on 2 January 2016).
- Wu, G. Impact of Human Activities on Water Level and Clarity and Underwater Light Climate of Vallisneria Spiralis L. in Poyang Lake, China. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, February 2008. [Google Scholar]
- Finlayson, M.; Harris, J.; McCartney, M.; Young, L.; Chen, Z. Report on Ramsar Visit to Poyang Lake Ramsar Site, P.R. China 12–17 April 2010; Report prepared on behalf of the Secretariat of the Ramsar Convention; The Ramsar Convention of Wetlands: Gland, Switzerland; Available online: http://archive.ramsar.org/pdf/Poyang_lake_report_v8.pdf (accessed on 15 January 2011).
- Guo, H.; Hu, Q.; Zhang, Q.; Feng, S. Effects of the Three Gorges Dam on Yangtze River flow and river interaction with Poyang Lake, China: 2003–2008. J. Hydrol. 2012, 416, 19–27. [Google Scholar] [CrossRef]
- Zhao, M.; Cong, P.; Barter, M.; Fox, A.D.; Cao, L. The changing abundance and distribution of Greater White-fronted Geese Anser albifrons in the Yangtze River floodplain: Impacts of recent hydrological changes. Bird Conserv. Int. 2012, 22, 135–143. [Google Scholar] [CrossRef]
- Sun, F.; Zhao, Y.; Gong, P.; Ma, R.; Dai, Y. Monitoring dynamic changes of global land cover types: Fluctuations of major lakes in China every 8 days during 2000–2010. Chin. Sci. Bull. 2014, 59, 171–189. [Google Scholar] [CrossRef]
- Dronova, I.; Gong, P.; Wang, L.; Zhong, L. Mapping dynamic cover types in a large seasonally flooded wetland using extended principal component analysis and object-based classification. Remote Sens. Environ. 2015, 158, 193–206. [Google Scholar] [CrossRef]
- Han, X.; Chen, X.; Feng, L. Four decades of winter wetland changes in Poyang Lake based on Landsat observations between 1973 and 2013. Remote Sens. Environ. 2015, 156, 426–437. [Google Scholar] [CrossRef]
- Markkola, J.; Iwabuchi, S.; Lei, G.; Aarvak, T.; Tolvanen, P.; Oien, I.J. Lesser white-fronted goose survey at the East Dongting and Poyang lakes in China, February 1999. In Fennoscandian Lesser White-fronted Goose Conservation Project. Annual Report 1999; Tolvanen, P., Øien, I.J., Ruokolainen, K., Eds.; WWF Finland Report No 12 and Norwegian Ornithological Society, NOF Rapportserie Report no. 1-2000: Helsinki, Finland, 2000; pp. 9–15. [Google Scholar]
- Shao, M.; Jiang, J.; Guo, H.; Zeng, B. Abundance, Distribution and Diversity Variations of Wintering Water Birds in Poyang Lake, Jiangxi Province, China. Pak. J. Zool. 2014, 46, 451–462. [Google Scholar]
- Shankman, D.; Keim, B.D.; Song, J. Flood frequency in China’s Poyang Lake region: Trends and teleconnections. Int. J. Climatol. 2006, 26, 1255–1266. [Google Scholar] [CrossRef]
- Qi, S.; Brown, D.G.; Tian, Q.; Jiang, L.; Zhao, T.; Bergen, K.A. Inundation Extent and Flood Frequency Mapping Using LANDSAT Imagery and Digital Elevation Models. Gisci. Remote Sens. 2009, 46, 101–127. [Google Scholar] [CrossRef]
- Dronova, I.; Gong, P.; Clinton, N.E.; Wang, L.; Fu, W.; Qi, S.; Liu, Y. Landscape analysis of wetland plant functional types: The effects of image segmentation scale, vegetation classes and classification methods. Remote Sens. Environ. 2012, 127, 357–369. [Google Scholar] [CrossRef]
- Wang, L.; Dronova, I.; Gong, P.; Yang, W.; Li, Y.; Liu, Q. A new time series vegetation-water index of phenological-hydrological trait across species and functional types for Poyang Lake wetland ecosystem. Remote Sens. Environ. 2012, 125, 49–63. [Google Scholar] [CrossRef]
- Fox, A.D.; Cao, L.; Zhang, Y.; Barter, M.; Zhao, M.J.; Meng, F.J.; Wang, S.L. Declines in the tuber-feeding waterbird guild at Shengjin Lake National Nature Reserve, China—A barometer of submerged macrophyte collapse. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 82–91. [Google Scholar] [CrossRef]
- Kwaiser, K. Accounting for Observation Uncertainty in Species-Habitat Models: A Case Study Using Bird Survey Data from Poyang Lake, China. Master’s Thesis, University of Michigan, Ann Arbor, MI, USA, August 2009. [Google Scholar]
- Barter, M.; Cao, L.; Chen, L.; Lei, G. Results of a survey for waterbirds in the lower Yangtze floodplain, China, in January–February 2004. Forktail 2005, 21, 1–7. [Google Scholar]
- Wu, X.; Lv, M.; Jin, Z.; Michishita, R.; Chen, J.; Tian, H.; Tu, X.; Zhao, H.; Niu, Z.; Chen, X.; et al. Normalized difference vegetation index dynamic and spatiotemporal distribution of migratory birds in the Poyang Lake wetland, China. Ecol. Indic. 2014, 47, 219–230. [Google Scholar] [CrossRef]
- Liordos, V. Foraging Guilds of Waterbirds Wintering in a Mediterranean Coastal Wetland. Zool. Stud. 2010, 49, 311–323. [Google Scholar]
- Chander, G.; Markham, B.L.; Helder, D.L. Summary of current radiometric calibration coefficients for Landsat MSS, TM, ETM+, and EO-1 ALI sensors. Remote Sens. Environ. 2009, 113, 893–903. [Google Scholar] [CrossRef]
- Vermote, E.F.; Tanre, D.; Deuze, J.L.; Herman, M.; Morcrette, J.J. Second Simulation of the Satellite Signal in the Solar Spectrum, 6S: An overview. IEEE Trans. Geosci. Remote Sens. 1997, 35, 675–686. [Google Scholar] [CrossRef]
- 6SV–MODIS Land Surface Reflectance Science Computing Facility. Available online: http://6s.ltdri.org/ (accessed on 10 February 2011).
- ESRI Data Basemaps. Available online: http://www.esri.com/data/basemaps (accessed on 5 April 2016).
- Trimble Documentation. eCognition Reference Book v. 8.8 2012; Trimble Germany GmbH: München, Germany, 2012. [Google Scholar]
- NASA/NGA Commercial Data Access. Available online: http://cad4nasa.gsfc.nasa.gov/ (accessed on 14 December 2011).
- Hall, M.; Frank, E.; Holmes, G.; Pfaringer, B.; Reutemann, P.; Witten, I. The WEKA Data Mining Software: An Update. ACM SIGKDD Explor. Newslett. 2009, 11, 10–18. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; p. 488. [Google Scholar]
- Jackman, S. pscl: Classes and Methods for R Developed in the Political Science Computational Laboratory, Stanford University. Department of Political Science, Stanford University. Stanford, California. R package Version 1.4.9. 2015. Available online: https://cran.r-project.org/web/packages/pscl/index.html (accessed on 10 April 2016).
- Zeileis, A.; Kleiber, C.; Jackman, S. Regression models for count data in R. J. Stat. Softw. 2008, 27, 1–25. [Google Scholar] [CrossRef]
- Anselin, L. Local indicators of spatial autocorrelation—LISA. Geogr. Anal. 1995, 27, 93–115. [Google Scholar] [CrossRef]
- Anselin, L.; Syabri, I.; Kho, Y. GeoDa: An introduction to spatial data analysis. Geogr. Anal. 2006, 38, 5–22. [Google Scholar] [CrossRef]
- Tuomisto, H. A diversity of beta diversities: Straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 2010, 33, 2–22. [Google Scholar] [CrossRef]
- Roshier, D.A.; Robertson, A.I.; Kingsford, R.T. Responses of waterbirds to flooding in an arid region of Australia and implications for conservation. Biol. Conserv. 2002, 106, 399–411. [Google Scholar] [CrossRef]
- Cerezo, A.; Cecilia Conde, M.; Poggio, S.L. Pasture area and landscape heterogeneity are key determinants of bird diversity in intensively managed farmland. Biodivers. Conserv. 2011, 20, 2649–2667. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Turner, D.P.; Cohen, W.B.; Kennedy, R.E.; Fassnacht, K.S.; Briggs, J.M. Relationships between leaf area index and Landsat TM spectral vegetation indices across three temperate zone sites. Remote Sens. Environ. 1999, 70, 52–68. [Google Scholar] [CrossRef]
- Goetz, S.; Steinberg, D.; Dubayah, R.; Blair, B. Laser remote sensing of canopy habitat heterogeneity as a predictor of bird species richness in an eastern temperate forest, USA. Remote Sens. Environ. 2007, 108, 254–263. [Google Scholar] [CrossRef]
- Liu, X.P.; Kelin, W.; Geli, Z. Perspectives and policies: Ecological industry substitutes in wetland restoration of the Middle Yangtze. Wetlands 2004, 24, 633–641. [Google Scholar]
- Xing, Y.; Xie, P.; Yang, H.; Wu, A.; Ni, L. The change of gaseous carbon fluxes following the switch of dominant producers from macrophytes to algae in a shallow subtropical lake of China. Atmos. Environ. 2006, 40, 8034–8043. [Google Scholar] [CrossRef]
- Legendre, P.; Dale, M.R.T.; Fortin, M.J.; Gurevitch, J.; Hohn, M.; Myers, D. The consequences of spatial structure for the design and analysis of ecological field surveys. Ecography 2002, 25, 601–615. [Google Scholar] [CrossRef]
- Laba, M.; Blair, B.; Downs, R.; Monger, B.; Philpot, W.; Smith, S.; Sullivan, P.; Baveye, P.C. Use of textural measurements to map invasive wetland plants in the Hudson River National Estuarine Research Reserve with IKONOS satellite imagery. Remote Sens. Environ. 2010, 114, 876–886. [Google Scholar] [CrossRef]
- Taft, O.W.; Colwell, M.A.; Isola, C.R.; Safran, R.J. Waterbird responses to experimental drawdown: Implications for the multispecies management of wetland mosaics. J. Appl. Ecol. 2002, 39, 987–1001. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Min–Max | Mean (st. dev.) |
|---|---|---|---|
| Species richness | Number of waterbird species per sub-lake | 2–23 | 9.1 (5.44) |
| Shannon index | Diversity index which accounts for both the number of species and evenness of their abundance calculated as , where pi is the proportion of individuals from species i in the whole dataset and R is the total number of species in the dataset | 0.2–2.03 | 1.1 (0.44) |
| Number of food guilds | The number of foraging guilds represented per sub-lake, out of 6 groups after [45]: tuber-feeding, sedge/grass-eating, seed eating/dabbling, benthic insect/larvae eating, fish eating and zooplankton eating birds | 1–6 | 3.9 (1.38) |
| Number of size groups | The number of bird groups defined by foraging habit (wading versus floating/diving birds) and size (average body mass greater or less than 2 kg for floating birds, average body length greater or less than 0.8 m for waders) | 1–4 | 3.2 (0.98) |
| Total waterbird abundance | Total number of waterbirds per sub-lake | 17–94,658 | 7201 (16,262) |
| Abundance of tuber feeding birds | The number of birds from the tuber-feeding foraging guild including Leucogeranus leucogeranus, Grus monarcha, Grus vipio, Anser cygnoides and Cygnus columbianus | 0–46,395 | 2750 (8185) |
| Cover Type Name | Description |
|---|---|
| Water | Inundated areas with water coverage above the ground or vegetation surface: sub-lakes, channels, pools, rivers etc. |
| Mudflat | Exposed lake bottomland directly adjacent to the water body with sparse (<30%) or no plant cover |
| Emergent grassland | Green photosynthetically active emergent wetland vegetation dominated by C3 grasses and forbs |
| Flooded vegetation | Green photosynthetically active vegetation with “wet” spectral signal (significantly lower near- and short-wave-infrared range than emergent grasses); includes inundated emergent, floating and submerged aquatic macrophytes and their mixtures |
| Senescent grasses | Perennial vegetation that maintains senescent biomass during the winter, typically dominated by mixed warm-season C4-grasses and reeds that grow in higher-elevation sub-lake and channel periphery |
| Burned vegetation | Recently burned grassland with distinct dark soil/ash and little or no vegetation regrowth |
| Human land use | Areas of active human land use adjacent to Poyang Lake wetlands (residential, agriculture, extraction, etc.) |
| Category | Name (Model Code) | Definition |
|---|---|---|
| Area | Total sub-lake unit area (area) | Total area of the sub-lake water body and its 500-m buffer neighborhood, m2 |
| Prevalence of habitat cover types within sub-lake neighborhood | Percent mudflat (%mudflat) | Proportion of the area classified as mudflat within the sub-lake neighborhood |
| Percent emergent vegetation (%emgrass) | Proportion of the area classified as emergent grassland within the sub-lake neighborhood | |
| Percent flooded vegetation (%floodveg) | Proportion of the area classified as flooded vegetation within the sub-lake neighborhood | |
| Spectral greenness | Normalized Difference Vegetation Index (NDVI) of emergent grassland (ndvi emgrass) | Spectral index of vegetation greenness, here calculated as mean object-level NDVI from Landsat TM bands 3 (red) and 4 (near-infrared) within green emergent grass class for each sub-lake: NDVI= (Band 4 − Band 3)/(Band 4 + Band 3) |
| Normalized Difference Vegetation Index (NDVI) of mudflat (ndvi mudflat) | Calculated using the same formula as above as mean of the objects within the mudflat class | |
| Spectral heterogeneity of habitat cover types | Spectral heterogeneity of mudflat (stdev Red mud) | Standard deviation of the object-level mean values for Landsat TM band 3 (red) among the mudflat image objects |
| Spectral heterogeneity of flooded vegetation (stdev Red floodveg) | Standard deviation of the object-level mean values for Landsat TM band 3 (red) among the flooded vegetation image objects | |
| Spectral heterogeneity of emergent vegetation (stdev Red emgrass) | Standard deviation of the object-level mean values for Landsat TM band 3 (red) among the emergent C3 grass image objects | |
| Heterogeneity of primitive patch shapes within habitat cover types | Heterogeneity of shape index for mudflat (stdev SI mud) | Standard deviation of the shape index (perimeter of the image object divided by four times the square root of its area [66]) for primitive image objects classified as mudflat within the sub-lake neighborhood |
| Heterogeneity of shape index for emergent vegetation (stdev SI emgrass) | Standard deviation of the shape index (perimeter of the image object divided by four times the square root of its area [66]) for primitive image objects classified as emergent grassland within the sub-lake neighborhood | |
| Potential human disturbance within sub-lake neighborhood | Percent of burned vegetation area (%burnveg) | Proportion of the area classified as burnt vegetation within the sub-lake neighborhood |
| Percent of human land use (%human LU) | Proportion of the area representing active human land use (residential or agriculture) within the sub-lake neighborhood |
| Assigned Class: | Reference Class | User’s Accuracy % | ||||||
|---|---|---|---|---|---|---|---|---|
| Emergent Grassland | Mudflat | Senescent Grassland | Flooded Vegetation | Water | Burned Vegetation | Sand | ||
| Overall Accuracy 94.3%, Kappa 0.934 | ||||||||
| Emergent grassland | 48 | 3 | 2 | 90.6 | ||||
| Mudflat | 47 | 1 | 2 | 94.0 | ||||
| Senescent grassland | 2 | 36 | 94.7 | |||||
| Flooded vegetation | 1 | 37 | 3 | 90.2 | ||||
| Water | 2 | 1 | 40 | 93.0 | ||||
| Burned vegetation | 35 | 100 | ||||||
| Sand | 40 | 100 | ||||||
| Producer’s accuracy,% | 96.0 | 94.0 | 90.0 | 92.5 | 100 | 87.5 | 100 | |
| Dependent Variable: | Species Richness | Shannon Index | Number of Food Guilds | |||||
| Independent Variable | R2 | AICC | Independent Variable | R2 | AICc | Independent Variable | R2 | AICc |
| ndvi mudflat | 0.35 | 303.2 | ndvi mudflat | 0.15 | 64.1 | ndvi mudflat | 0.21 | 178.5 |
| Sqrt (%floodveg) | 0.21 | 313.4 | Sqrt (%floodveg) | 0.12 | 65.5 | Sqrt (%floodveg) | 0.12 | 184.1 |
| Stdev Red mud | 0.11 | 319.3 | Stdev Red floodveg | 0.08 | 67.9 | Ln(area) | 0.08 | 186.1 |
| Intercept-only | 0.00 | 322.7 | Intercept-only | 0.00 | 69.81 | Intercept-only | 0.00 | 188.3 |
| Dependent Variable: | Number of Size Groups | Ln (Number of Birds) | Ln (Number of Tuber Feeding Birds + 1) | |||||
| Independent Variable | R2 | AICc | Independent Variable | R2 | AICc | Independent Variable | R2 | AICc |
| ndvi mudflat | 0.15 | 149.2 | Ln(area) | 0.2 | 216.0 | ndvi mudflat | 0.09 | 267.3 |
| ndvi emgrass | 0.09 | 152.4 | ndvi mudflat | 0.13 | 220.4 | Sqrt (%floodveg) | 0.08 | 267.4 |
| Ln (area) | 0.08 | 153.2 | Intercept-only | 0.00 | 225.2 | Intercept-only | 0.00 | 269.6 |
| Intercept-only | 0.00 | 155.2 | ||||||
| Models | R2 OLS | AICc OLS | AICc Lag | AICc Error | Akaike Weight |
|---|---|---|---|---|---|
| Dependent Variable: Species Richness | |||||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − ndvi mudflat − stdev SI emgrass | 0.54 | 294.38 | 297.17 | 291.54 | 0.60 |
| −Sqrt(%floodveg) + ndvi emgrass – ndvi mudflat − stdev SI emgrass | 0.50 | 295.94 | 298.60 | 293.92 | |
| −Sqrt(%floodveg) + ndvi emgrass − ndvi mudflat − stdev SI emgrass + stdev Red emgrass | 0.52 | 296.15 | 298.83 | 292.34 | 0.40 |
| Dependent Variable: Shannon Index | |||||
| −Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg + stdev Red emgrass | 0.40 | 52.69 * | 0.27 | ||
| −Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg | 0.37 | 52.76 * | 0.26 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg | 0.39 | 53.01 * | 0.23 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg + stdev Red emgrass | 0.42 | 53.90 * | 0.15 | ||
| −Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg + stdev Red emgrass − ndvi mudflat | 0.41 | 54.58 * | 0.10 | ||
| Dependent Variable: Number Of Food Guilds | |||||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass | 0.38 | 169.73 | 170.45 | 172.24 | 0.35 |
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − ndvi mudflat | 0.41 | 170.23 * | 0.4 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − %emgrass | 0.41 | 170.29 | 171.18 | 172.96 | 0.25 |
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass + %human LU | 0.39 | 171.55 | 172.72 | 174.24 | |
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − stdev SI emgrass | 0.39 | 171.73 | 172.55 | 174.42 | |
| Dependent Variable: Number of Size Groups | |||||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass | 0.33 | 140.55 * | 0.25 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass + %human LU | 0.35 | 141.59 * | 0.15 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − ndvi mudflat | 0.35 | 141.71 * | 0.14 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − %emgrass | 0.35 | 141.72 * | 0.14 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − %burnveg | 0.35 | 141.89 * | 0.13 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass − %mudflat | 0.34 | 142.41 * | 0.10 | ||
| Ln(area) − Sqrt(%floodveg) + ndvi emgrass + stdev Red floodveg | 0.34 | 142.45 * | 0.10 | ||
| Dependent Variable: Natural Log of the Total Number of Birds | |||||
| Ln(area) + ndvi emgrass − ndvi mudflat − stdev SI emgrass | 0.4 | 207.90 | 209.62 | 210.56 | 0.23 |
| Ln(area) − ndvi mudflat − stdev SI emgrass | 0.35 | 209.19 * | 0.44 | ||
| Ln(area) + ndvi emgrass − ndvi mudflat − stdev SI emgrass − %burnveg | 0.41 | 209.24 | 211.47 | 211.59 | |
| Ln(area) − ndvi mudflat − stdev SI emgrass − %burnveg | 0.38 | 209.83 * | 0.33 | ||
| Ln(area) + ndvi emgrass − ndvi mudflat − stdev SI emgrass − Sqrt(%floodveg) | 0.41 | 209.84 | 211.55 | 212.63 | |
| Dependent Variable: (Natural Log + 1) of the Number of Tuber Feeding Birds | |||||
| Ln(area) − Sqrt(%floodveg) | 0.16 | 264.41 | 263.87 | 265.17 | 0.36 |
| Ln(area) − Sqrt(%floodveg) − stdev SI emgrass | 0.19 | 264.79 | 264.56 | 266.11 | 0.22 |
| − ndvi mudflat − stdev SI emgrass | 0.14 | 265.19 | 264.54 | 265.61 | 0.28 |
| Ln(area) − Sqrt(%floodveg) − %burnveg | 0.17 | 266.15 | 266.03 | 267.45 | |
| Ln(area) − ndvi mudflat − stdev SI emgrass | 0.17 | 266.19 | 265.51 | 266.97 | 0.14 |
| − ndvi mudflat | 0.09 | 266.25 * | |||
| %emgrass − ndvi mudflat − stdev SI emgrass | 0.17 | 266.31 * | |||
| − Sqrt(%floodveg) | 0.08 | 266.39 * | |||
| Count Model | Zero-Inflation Model | AICc | Akaike Weight |
|---|---|---|---|
| Ln(area) − stdev SI emgrass | ndvi mudflat + Sqrt(%floodveg) | 661.4 | 0.28 |
| Ln(area) − stdev SI emgrass | Sqrt(%floodveg) | 661.5 | 0.27 |
| Ln(area) − stdev SI emgrass − Sqrt(%floodveg) | Sqrt(%floodveg) | 663.0 | 0.13 |
| Ln(area) − stdev SI emgrass − Sqrt(%floodveg) | ndvi mudflat + Sqrt(%floodveg) | 663.3 | 0.11 |
| Ln(area) – stdev SI emgrass − ndvi mudflat | Sqrt(%floodveg) | 663.3 | 0.11 |
| ndvi emgrass − ndvi mudflat − %burnveg | Sqrt(%floodveg) | 663.3 | 0.11 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dronova, I.; Beissinger, S.R.; Burnham, J.W.; Gong, P. Landscape-Level Associations of Wintering Waterbird Diversity and Abundance from Remotely Sensed Wetland Characteristics of Poyang Lake. Remote Sens. 2016, 8, 462. https://doi.org/10.3390/rs8060462
Dronova I, Beissinger SR, Burnham JW, Gong P. Landscape-Level Associations of Wintering Waterbird Diversity and Abundance from Remotely Sensed Wetland Characteristics of Poyang Lake. Remote Sensing. 2016; 8(6):462. https://doi.org/10.3390/rs8060462
Chicago/Turabian StyleDronova, Iryna, Steven R. Beissinger, James W. Burnham, and Peng Gong. 2016. "Landscape-Level Associations of Wintering Waterbird Diversity and Abundance from Remotely Sensed Wetland Characteristics of Poyang Lake" Remote Sensing 8, no. 6: 462. https://doi.org/10.3390/rs8060462
APA StyleDronova, I., Beissinger, S. R., Burnham, J. W., & Gong, P. (2016). Landscape-Level Associations of Wintering Waterbird Diversity and Abundance from Remotely Sensed Wetland Characteristics of Poyang Lake. Remote Sensing, 8(6), 462. https://doi.org/10.3390/rs8060462