
Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel



Abstract
:
1. Introduction
2. Materials and Methods
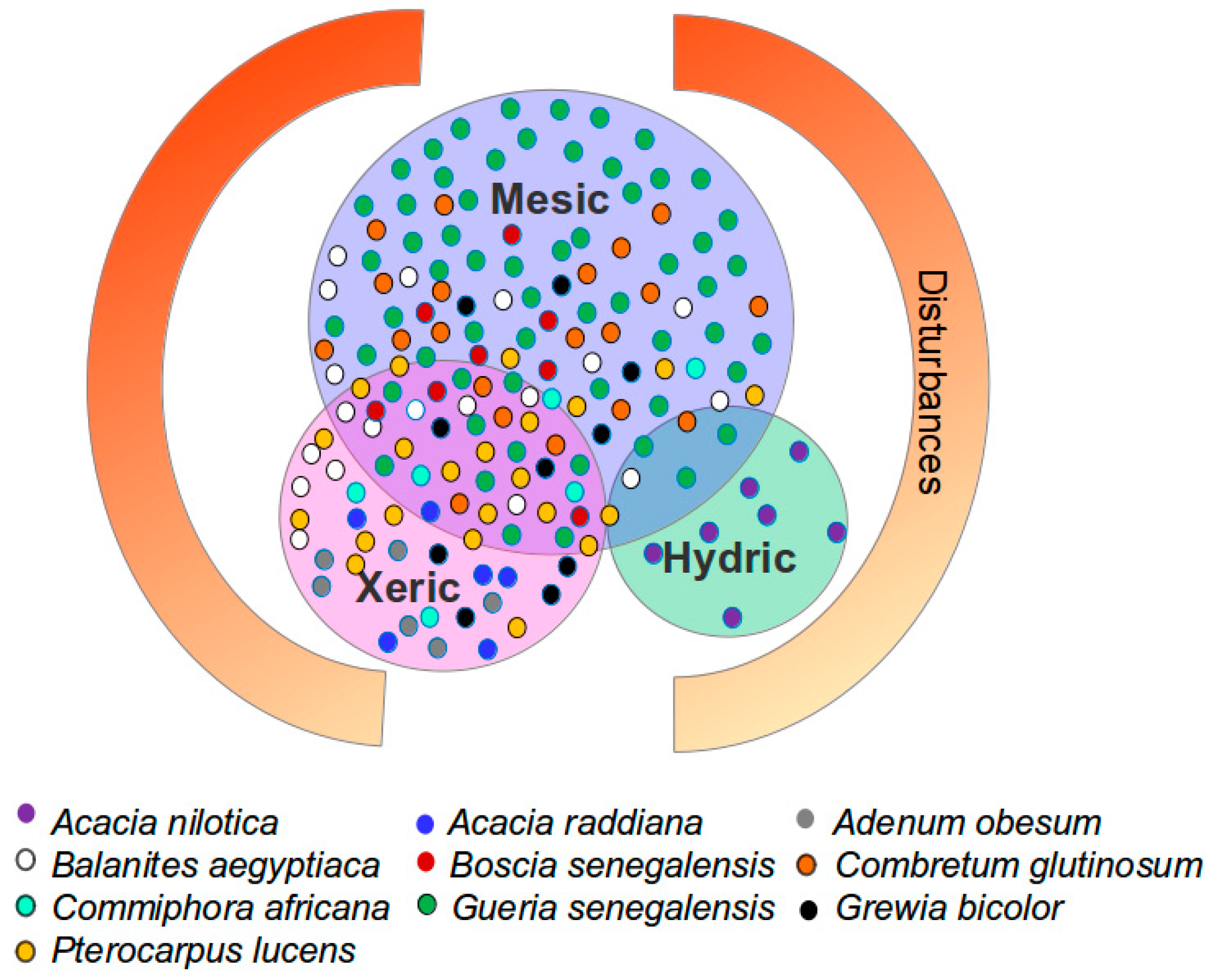
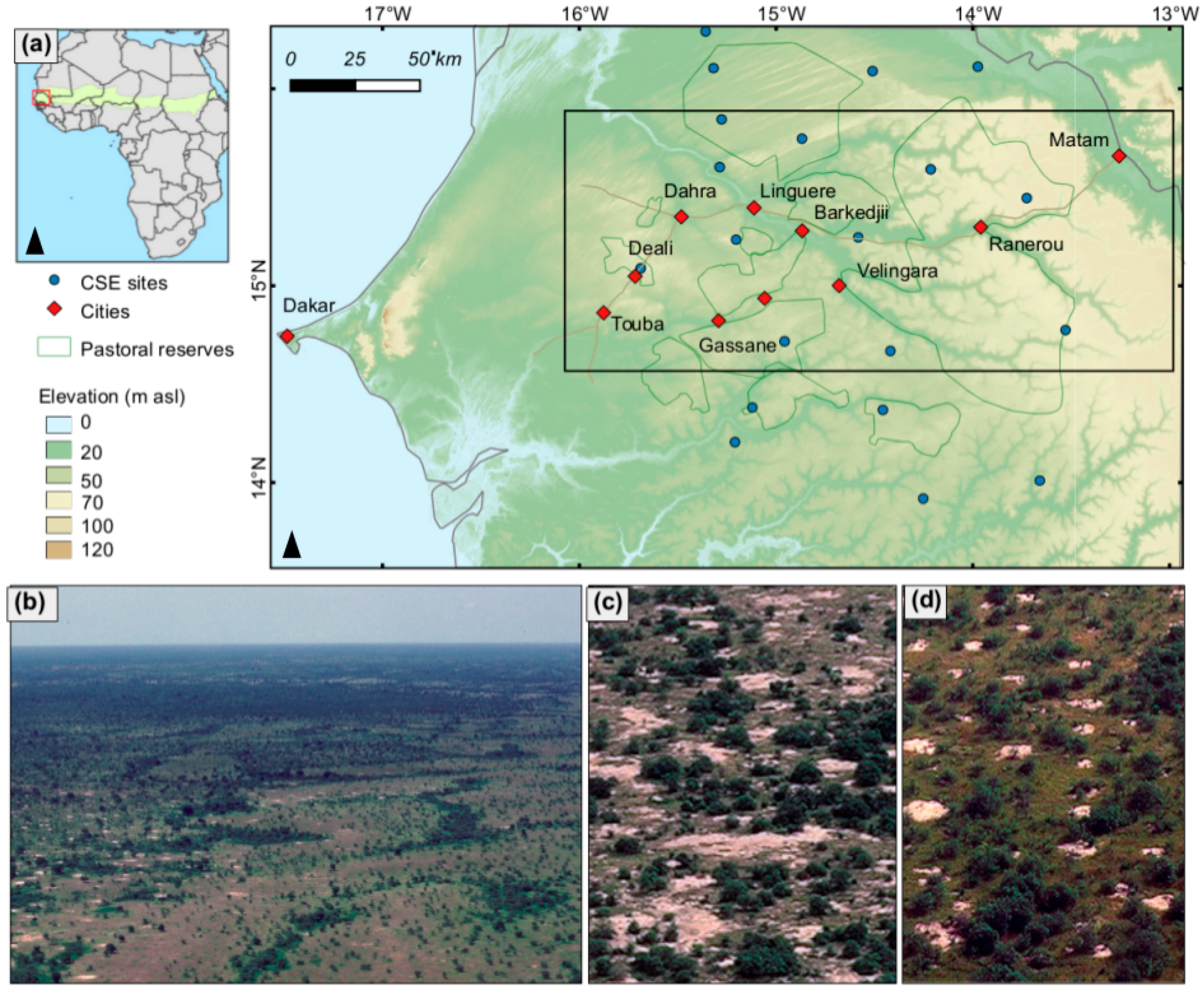
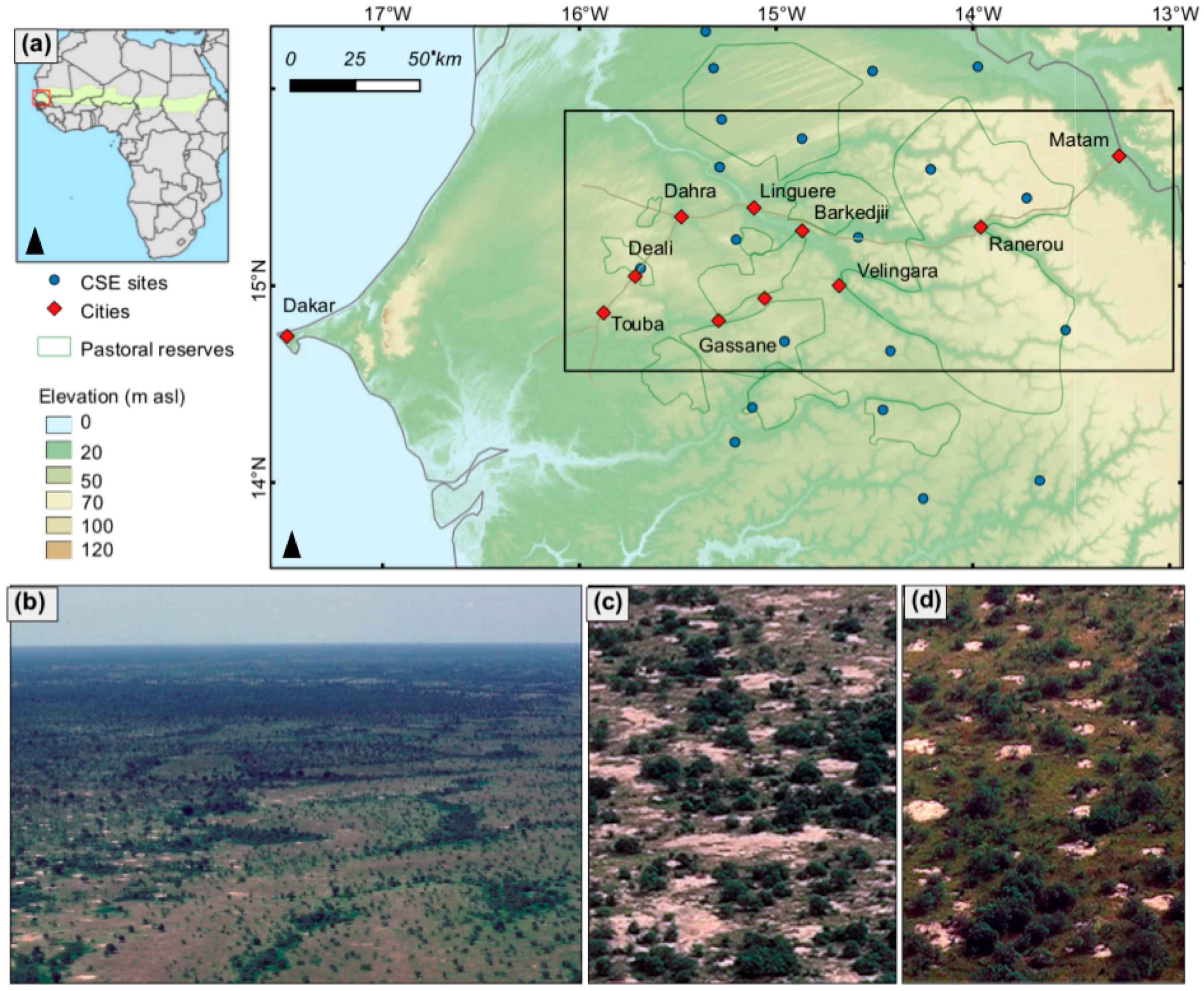
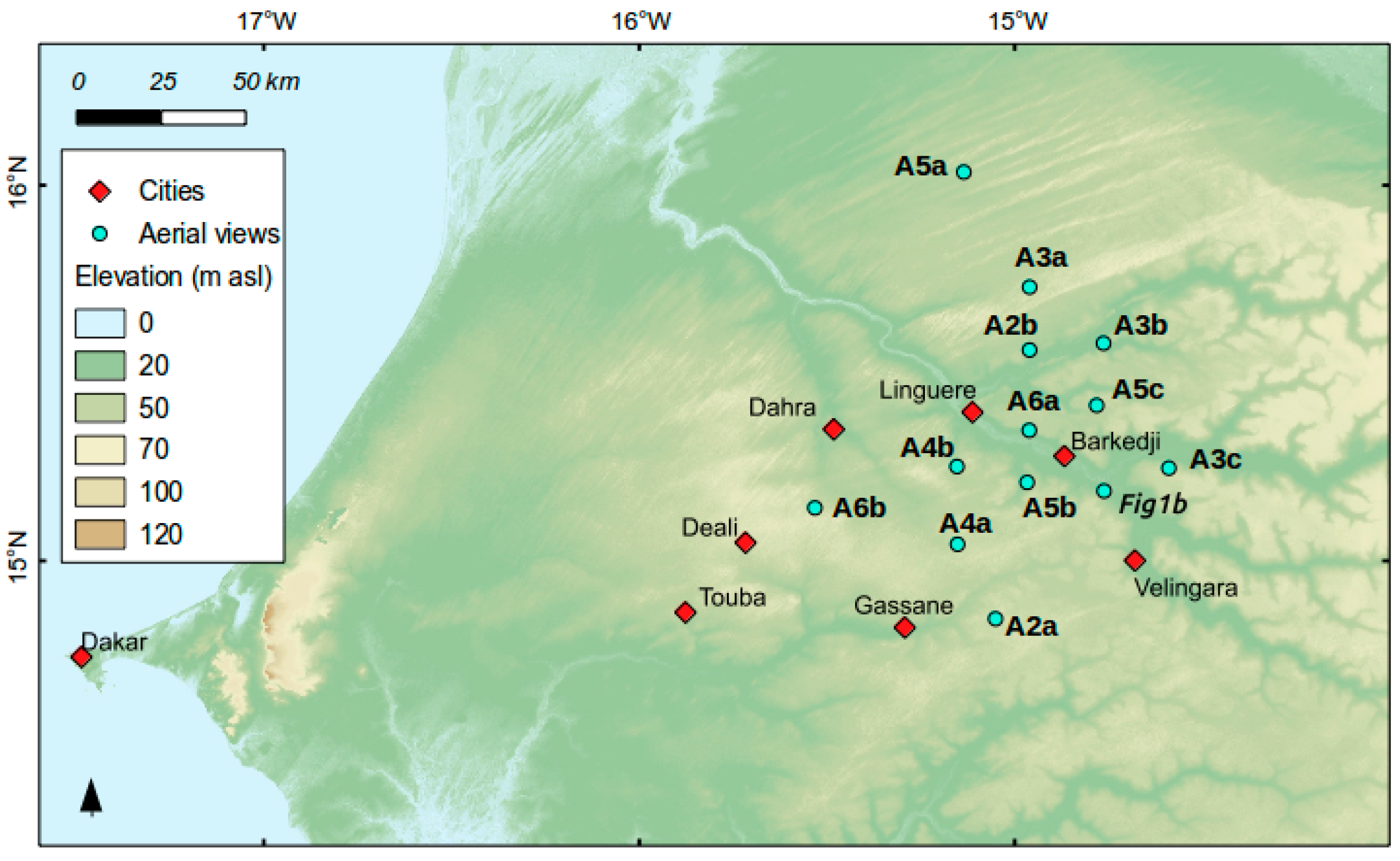
2.1. Study Area
2.2. Satellite Data
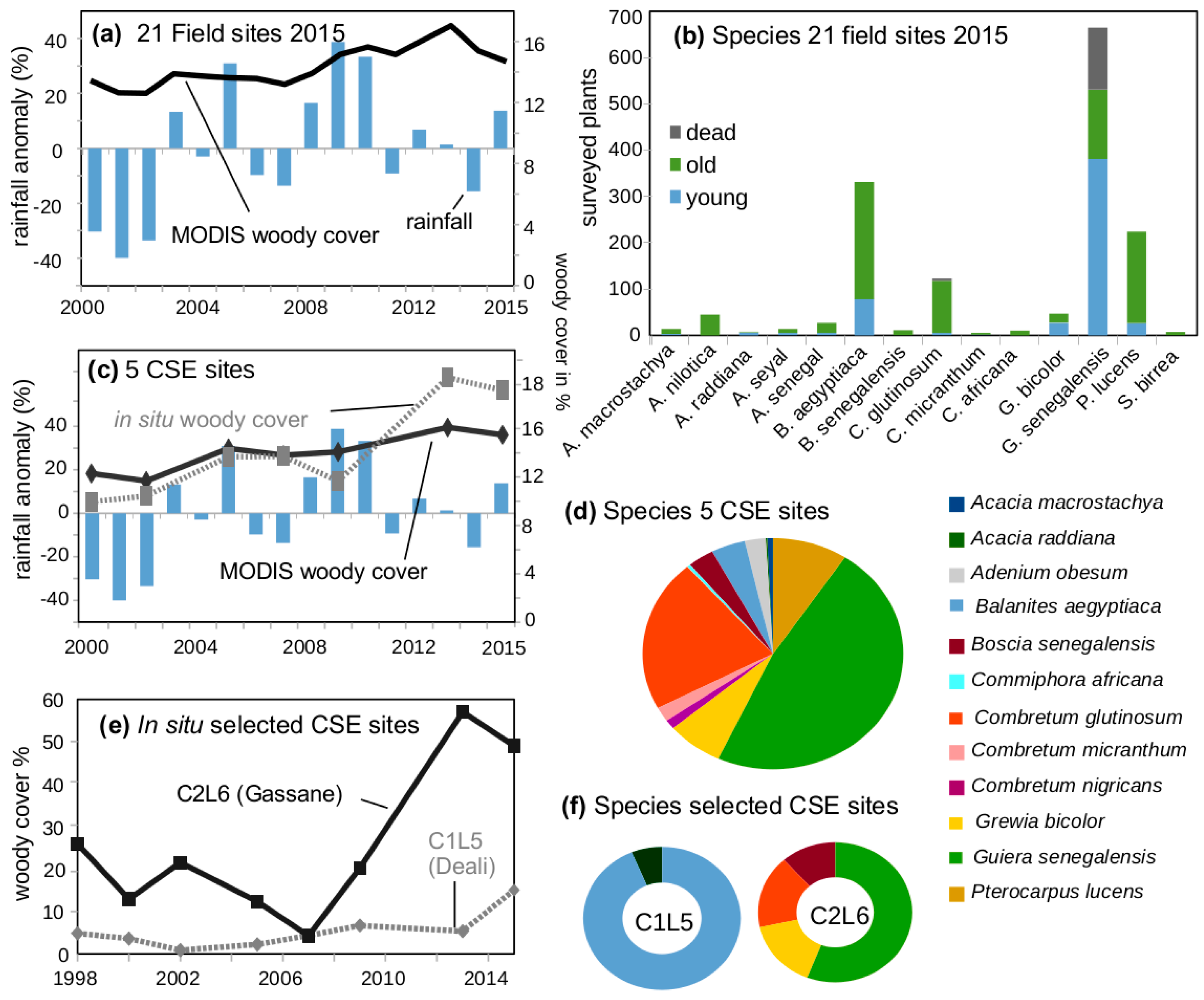
2.3. Estimating Woody Cover Dynamics from Sahel-Wide in Situ Data and Satellite Time Series
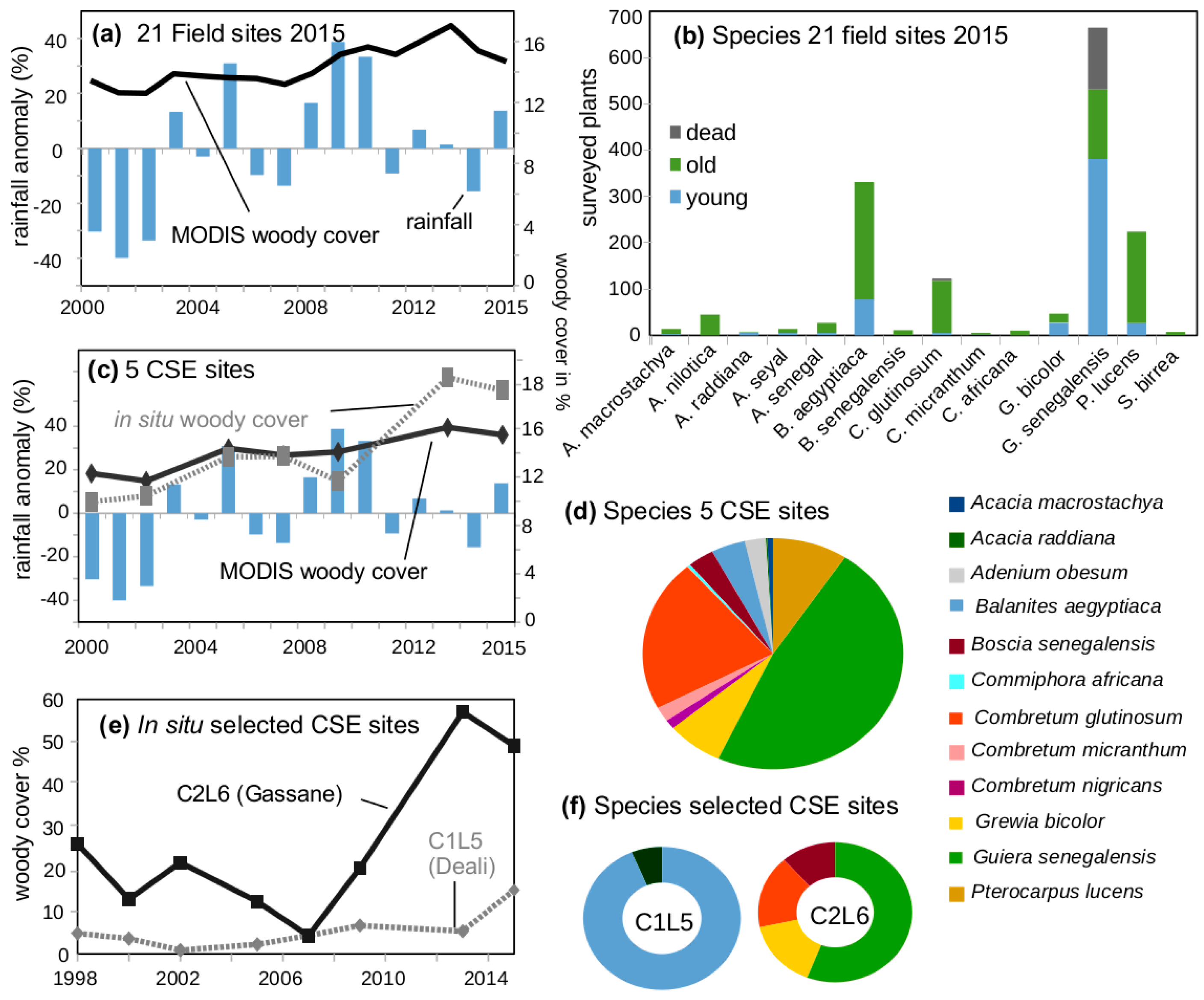
2.4. In Situ Woody Cover Measurements (5 CSE Sites)
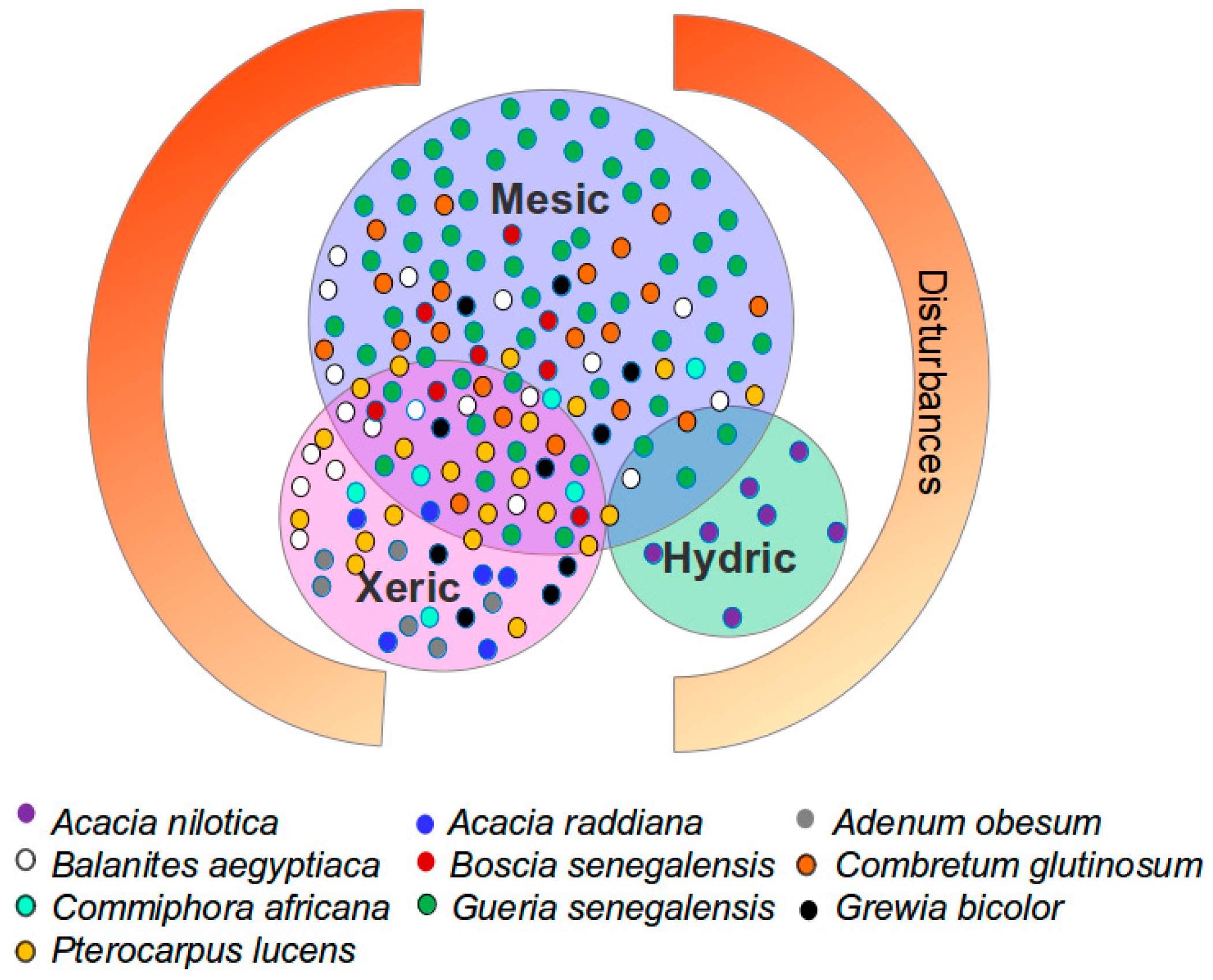
2.5. In Situ Woody Species Measurements (21 2015 Field Sites)
2.6. Repeat Photography
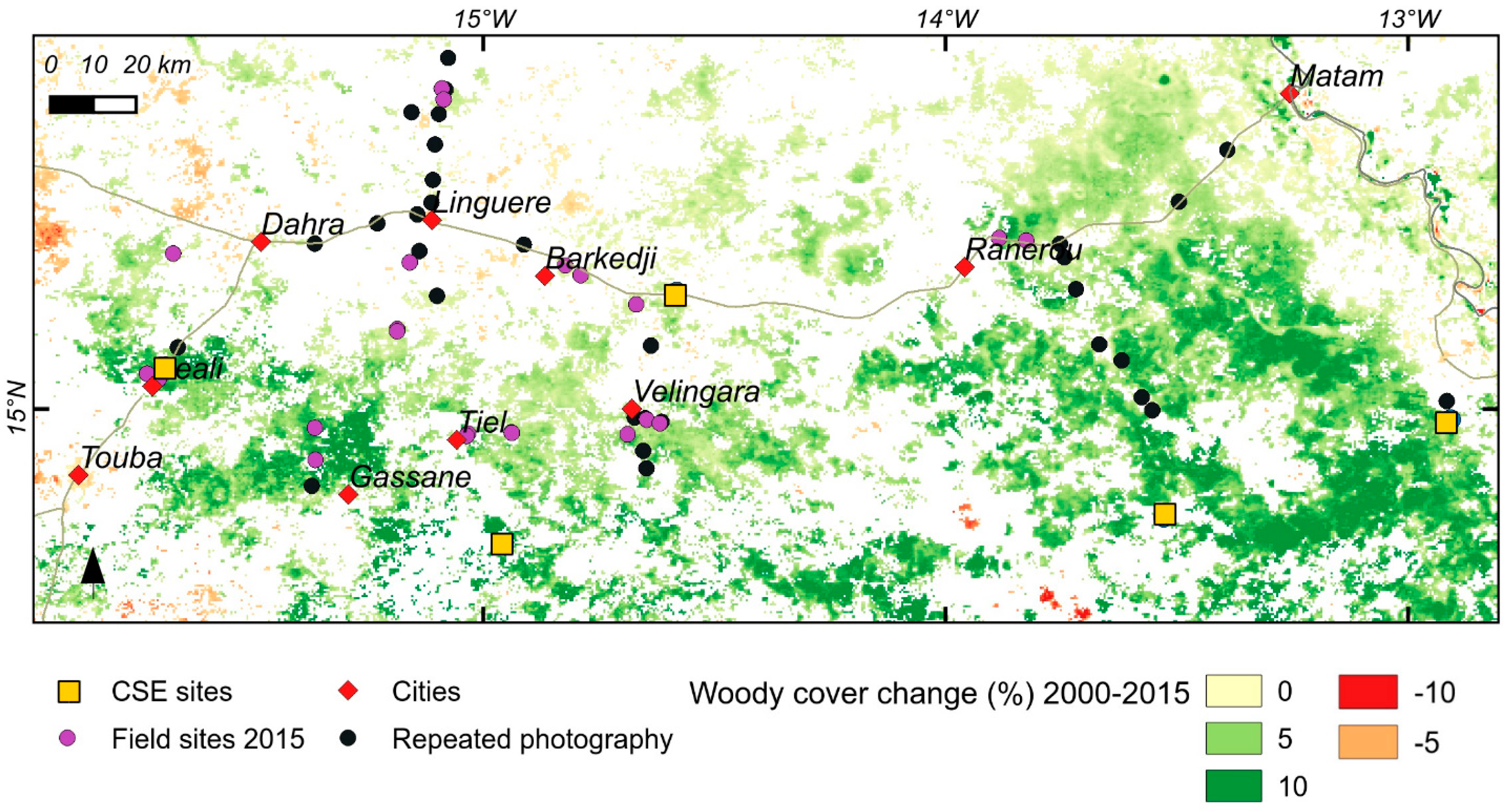
2.7. Mapping Negative Anomalies in Woody Cover
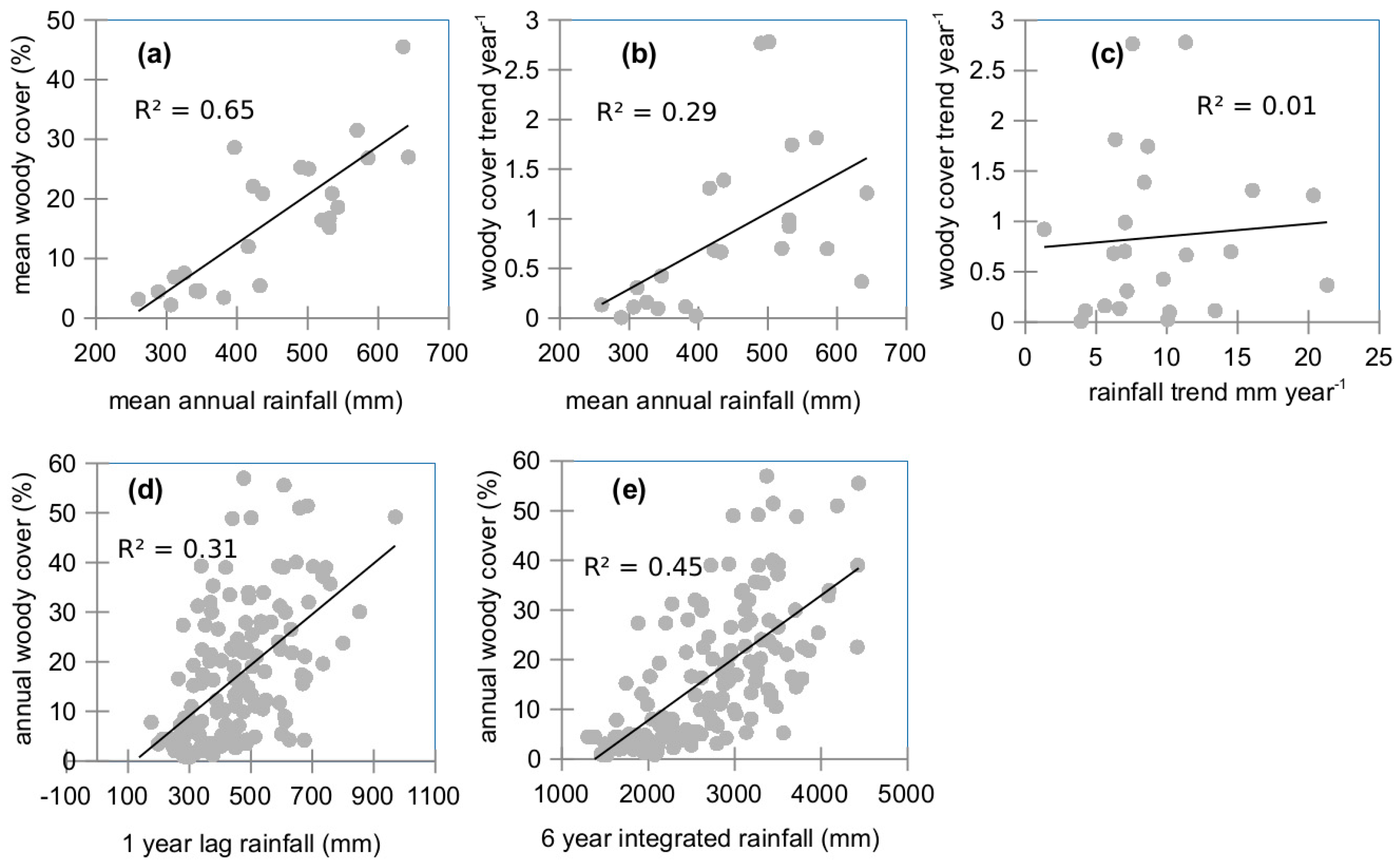
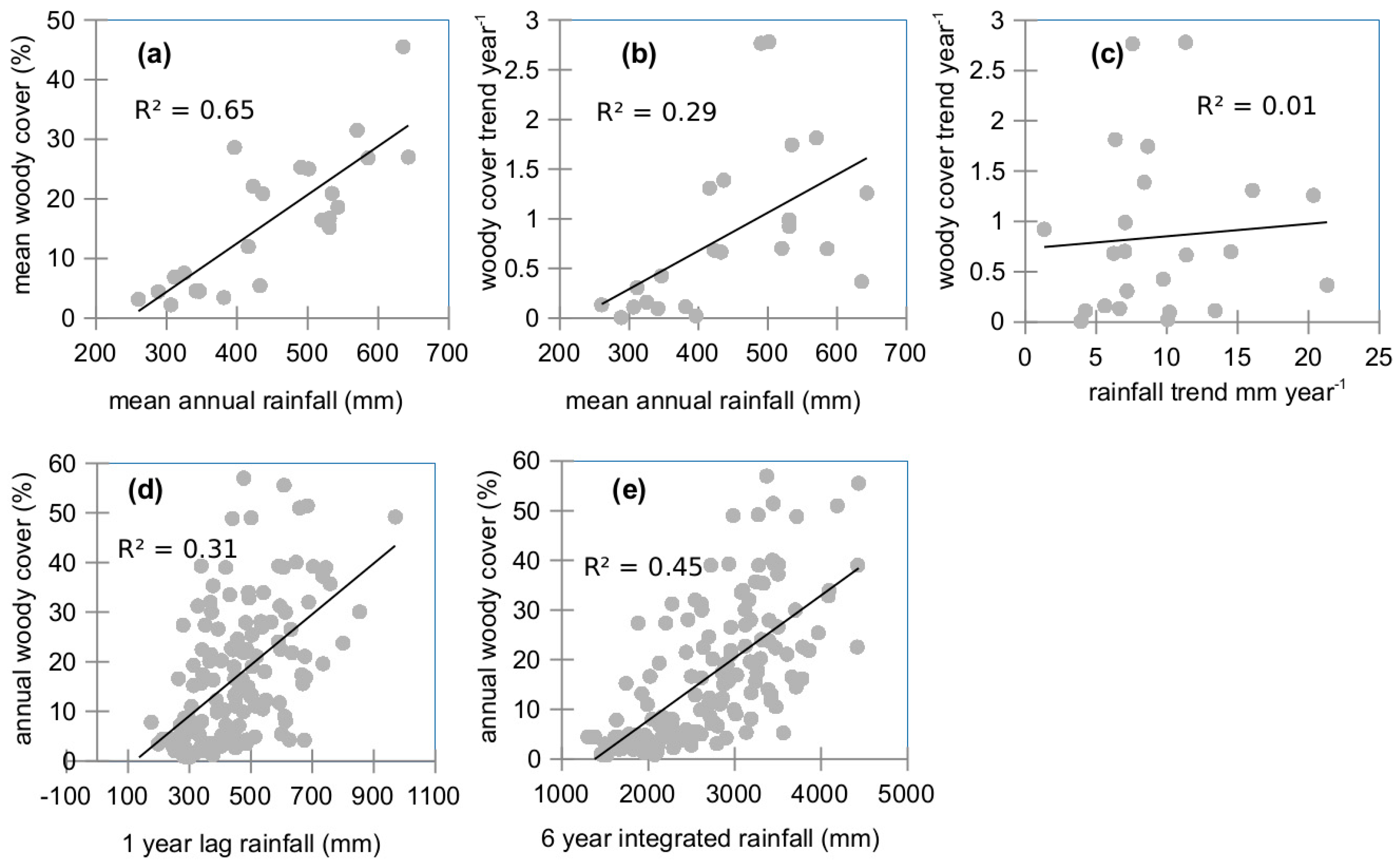
2.8. Relation between Woody Vegetation and Rainfall
3. Results
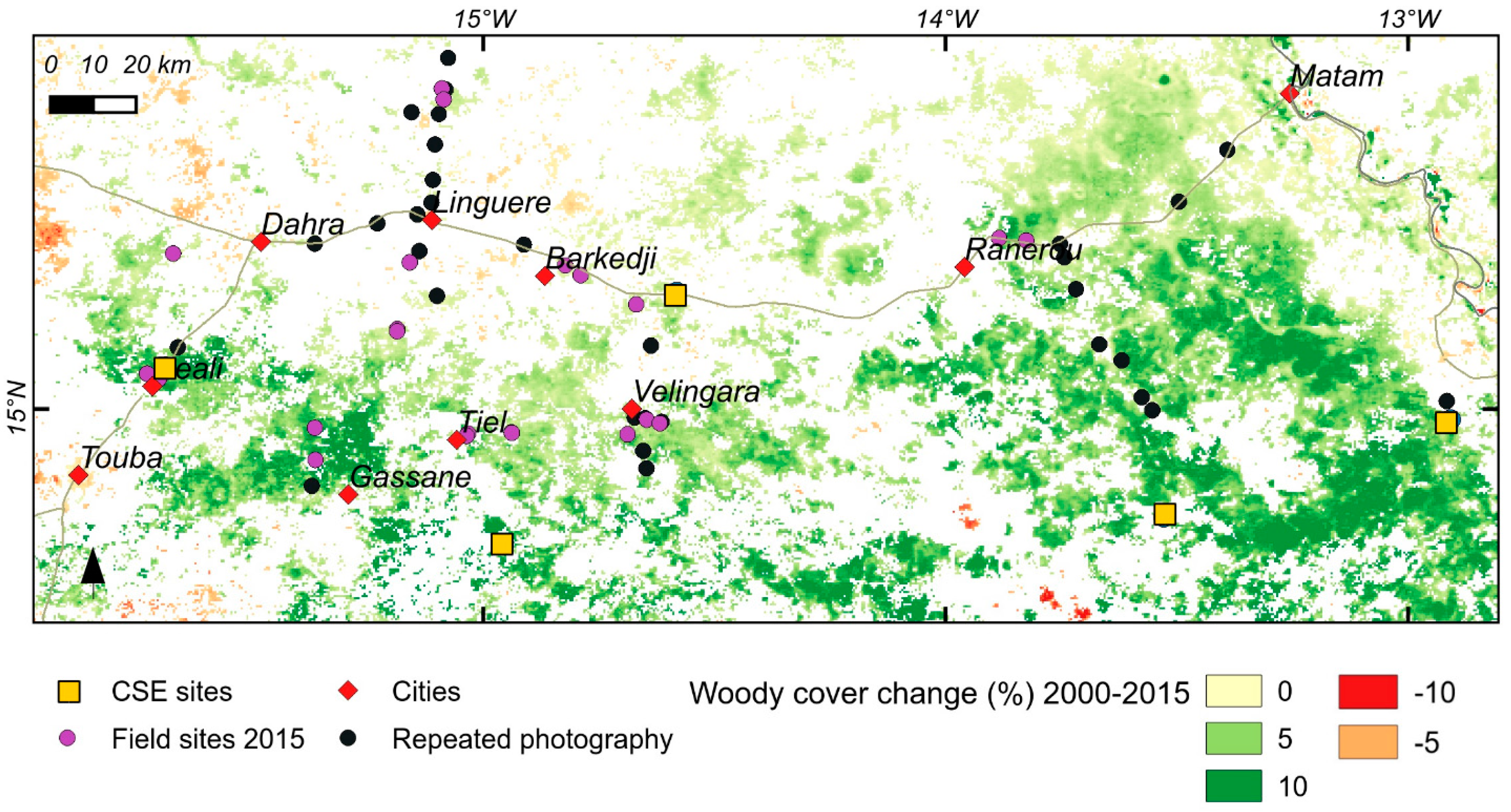
3.1. Woody Cover Changes in 2000–2015
3.2. Species Composition at “Woody Greening” Sites
3.3. Repeat Photography
3.4. Earth Observation Based Mapping of Woody Mortality Rate
3.5. Mass Dying of Guiera senegalensis
3.6. Rainfall Dynamics Impact on Woody Cover
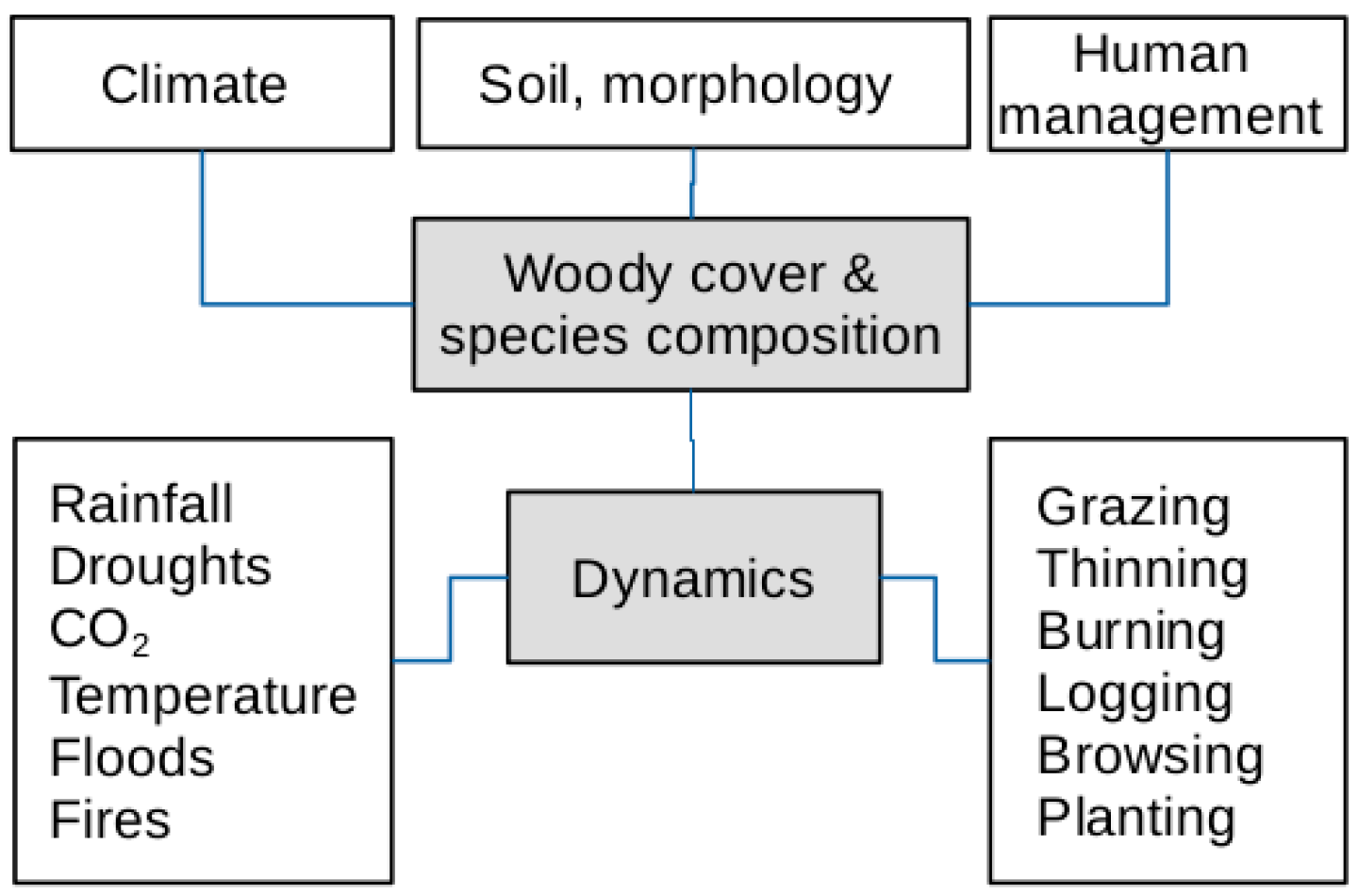
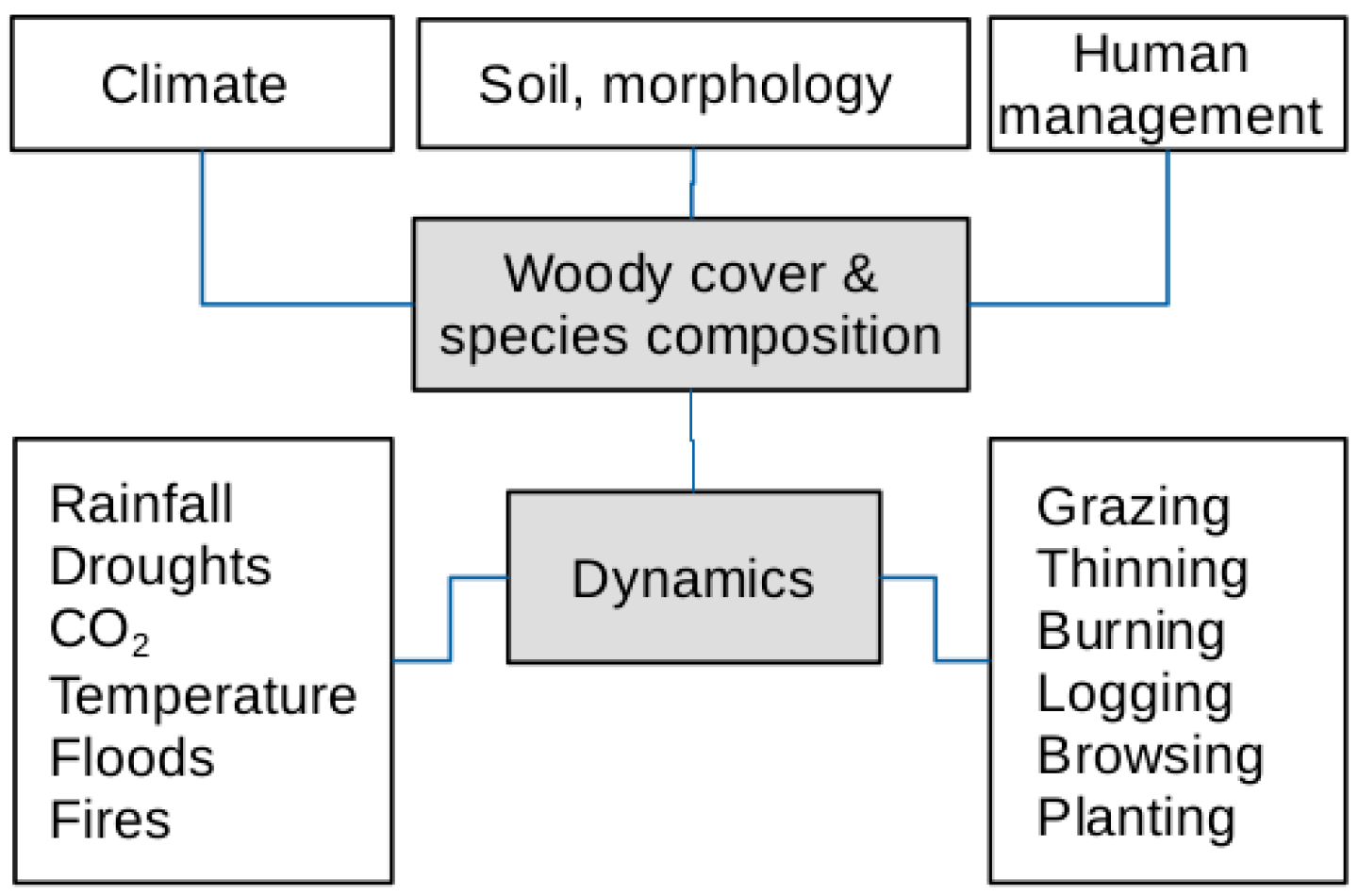
4. Discussion
5. Conclusions and Outlook
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A







References
- Karlson, M.; Ostwald, M. Remote sensing of vegetation in the Sudano-Sahelian zone: A literature review from 1975 to 2014. J. Arid Environ. 2016, 124, 257–269. [Google Scholar] [CrossRef]
- Mbow, C.; Brandt, M.; Ouedraogo, I.; de Leeuw, J.; Marshall, M. What four decades of Earth Observation tell us about land degradation in the Sahel? Remote Sens. 2015, 7, 4048–4067. [Google Scholar] [CrossRef]
- Knauer, K.; Gessner, U.; Dech, S.; Kuenzer, C. Remote sensing of vegetation dynamics in West Africa. Int. J. Remote Sens. 2014, 35, 6357–6396. [Google Scholar] [CrossRef]
- Rasmussen, K.; D’haen, S.; Fensholt, R.; Fog, B.; Horion, S.; Nielsen, J.O.; Rasmussen, L.V.; Reenberg, A. Environmental change in the Sahel: Reconciling contrasting evidence and interpretations. Reg. Environ. Chang. 2016, 16, 673–680. [Google Scholar] [CrossRef]
- Kaptué, A.T.; Prihodko, L.; Hanan, N.P. On regreening and degradation in Sahelian watersheds. Proc. Natl. Acad. Sci. USA 2015, 112, 12133–12138. [Google Scholar] [CrossRef] [PubMed]
- Breman, H.; Kessler, J.-J. Woody Plants in Agro-Ecosystems of Semi-Arid Regions: With an Emphasis on the Sahelian Countries, 1st ed.; Springer: Berlin, Germany, 1995. [Google Scholar]
- Brandt, M.; Mbow, C.; Diouf, A.A.; Verger, A.; Samimi, C.; Fensholt, R. Ground- and satellite-based evidence of the biophysical mechanisms behind the greening Sahel. Glob. Chang. Biol. 2015, 21, 1610–1620. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, S.M.; Tappan, G.G. Vegetation impoverishment despite greening: A case study from central Senegal. J. Arid Environ. 2013, 90, 55–66. [Google Scholar] [CrossRef]
- Gonzalez, P.; Tucker, C.J.; Sy, H. Tree density and species decline in the African Sahel attributable to climate. J. Arid Environ. 2012, 78, 55–64. [Google Scholar] [CrossRef]
- Hiernaux, P.; Diarra, L.; Trichon, V.; Mougin, E.; Soumaguel, N.; Baup, F. Woody plant population dynamics in response to climate changes from 1984 to 2006 in Sahel (Gourma, Mali). J. Hydrol. 2009, 375, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Brandt, M.; Grau, T.; Mbow, C.; Samimi, C. Modeling soil and woody vegetation in the Senegalese Sahel in the context of environmental change. Land 2014, 3, 770–792. [Google Scholar] [CrossRef]
- Vincke, C.; Diédhiou, I.; Grouzis, M. Long term dynamics and structure of woody vegetation in the Ferlo (Senegal). J. Arid Environ. 2010, 74, 268–276. [Google Scholar] [CrossRef]
- Tappan, G.G.; Sall, M.; Wood, E.C.; Cushing, M. Ecoregions and land cover trends in Senegal. J. Arid Environ. 2004, 59, 427–462. [Google Scholar] [CrossRef]
- Budde, M.E.; Tappan, G.; Rowland, J.; Lewis, J.; Tieszen, L.L. Assessing land cover performance in Senegal, West Africa using 1-km integrated NDVI and local variance analysis. J. Arid Environ. 2004, 59, 481–498. [Google Scholar] [CrossRef]
- Tian, F.; Brandt, M.; Liu, Y.Y.; Verger, A.; Tagesson, T.; Diouf, A.A.; Rasmussen, K.; Mbow, C.; Wang, Y.; Fensholt, R. Remote sensing of vegetation dynamics in drylands: Evaluating vegetation optical depth (VOD) using AVHRR NDVI and in situ green biomass data over West African Sahel. Remote Sens. Environ. 2016, 177, 265–276. [Google Scholar] [CrossRef]
- Tucker, C.J.; Vanpraet, C.; Boerwinkel, E.; Gaston, A. Satellite remote sensing of total dry matter production in the Senegalese Sahel. Remote Sens. Environ. 1983, 13, 461–474. [Google Scholar] [CrossRef]
- Brandt, M.; Hiernaux, P.; Rasmussen, K.; Mbow, C.; Kergoat, L.; Tagesson, T.; Ibrahim, Y.Z.; Wélé, A.; Tucker, C.J.; Fensholt, R. Assessing woody vegetation trends in Sahelian drylands using MODIS based seasonal metrics. Remote Sens. Environ. 2016, 183, 215–225. [Google Scholar] [CrossRef]
- Helman, D.; Lensky, I.; Tessler, N.; Osem, Y. A phenology-based method for monitoring woody and herbaceous vegetation in Mediterranean forests from NDVI time series. Remote Sens. 2015, 7, 12314–12335. [Google Scholar] [CrossRef]
- Dye, D.G.; Middleton, B.R.; Vogel, J.M.; Wu, Z.; Velasco, M. Exploiting differential vegetation Phenology for satellite-based mapping of semiarid grass vegetation in the Southwestern United States and Northern Mexico. Remote Sens. 2016, 8, 889. [Google Scholar] [CrossRef]
- De Keersmaecker, W.; Lhermitte, S.; Honnay, O.; Farifteh, J.; Somers, B.; Coppin, P. How to measure ecosystem stability? An evaluation of the reliability of stability metrics based on remote sensing time series across the major global ecosystems. Glob. Chang. Biol. 2014, 20, 2149–2161. [Google Scholar] [CrossRef] [PubMed]
- Diouf, A.A.; Brandt, M.; Verger, A.; Jarroudi, M.E.; Djaby, B.; Fensholt, R.; Ndione, J.A.; Tychon, B. Fodder biomass monitoring in Sahelian rangelands using phenological metrics from FAPAR time series. Remote Sens. 2015, 7, 9122–9148. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hall, F.G. The interpretation of spectral vegetation indexes. IEEE Trans. Geosci. Remote Sens. 1995, 33, 481–486. [Google Scholar] [CrossRef]
- Dinku, T.; Ceccato, P.; Grover-Kopec, E.; Lemma, M.; Connor, S.J.; Ropelewski, C.F. Validation of satellite rainfall products over East Africa’s complex topography. Int. J. Remote Sens. 2007, 28, 1503–1526. [Google Scholar] [CrossRef]
- Toté, C.; Patricio, D.; Boogaard, H.; van der Wijngaart, R.; Tarnavsky, E.; Funk, C. Evaluation of satellite rainfall estimates for drought and flood monitoring in Mozambique. Remote Sens. 2015, 7, 1758–1776. [Google Scholar] [CrossRef]
- Novella, N.S.; Thiaw, W.M. African rainfall climatology version 2 for famine early warning systems. J. Appl. Meteorol. Climatol. 2013, 52, 588–606. [Google Scholar] [CrossRef]
- Brandt, M.; Hiernaux, P.; Tagesson, T.; Verger, A.; Rasmussen, K.; Diouf, A.A.; Mbow, C.; Mougin, E.; Fensholt, R. Woody plant cover estimation in drylands from Earth Observation based seasonal metrics. Remote Sens. Environ. 2016, 172, 28–38. [Google Scholar] [CrossRef]
- Bégué, A.; Vintrou, E.; Ruelland, D.; Claden, M.; Dessay, N. Can a 25-year trend in Soudano-Sahelian vegetation dynamics be interpreted in terms of land use change? A remote sensing approach. Glob. Environ. Chang. 2011, 21, 413–420. [Google Scholar] [CrossRef]
- Stevens, N.; Lehmann, C.E.R.; Murphy, B.P.; Durigan, G. Savanna woody encroachment is widespread across three continents. Glob. Chang. Biol. 2017, 23, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Mitchard, E.T.A.; Flintrop, C.M. Woody encroachment and forest degradation in sub-Saharan Africa’s woodlands and savannas 1982–2006. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, T.G.; Puttick, J.R.; Hoffman, M.T. Bush encroachment in southern Africa: Changes and causes. Afr. J. Range Forage Sci. 2014, 31, 67–88. [Google Scholar] [CrossRef]
- Buitenwerf, R.; Bond, W.J.; Stevens, N.; Trollope, W.S.W. Increased tree densities in South African savannas: >50 years of data suggests CO2 as a driver. Glob. Chang. Biol. 2012, 18, 675–684. [Google Scholar] [CrossRef]
- Stevens, N.; Erasmus, B.F.N.; Archibald, S.; Bond, W.J. Woody encroachment over 70 years in South African savannahs: Overgrazing, global change or extinction aftershock? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150437. [Google Scholar] [CrossRef] [PubMed]
- Wigley, B.J.; Bond, W.J.; Hoffman, M.T. Thicket expansion in a South African savanna under divergent land use: Local vs. global drivers? Glob. Chang. Biol. 2010, 16, 964–976. [Google Scholar] [CrossRef]
- Tian, F.; Brandt, M.; Liu, Y.Y.; Rasmussen, K.; Fensholt, R. Mapping gains and losses in woody vegetation across global tropical drylands. Glob. Chang. Biol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Gaze, S.R.; Brouwer, J.; Simmonds, L.P.; Bromley, J. Dry season water use patterns under Guiera senegalensis L. shrubs in a tropical savanna. J. Arid Environ. 1998, 40, 53–67. [Google Scholar] [CrossRef]
- Hill, M.J.; Hanan, N.P. Ecosystem Function in Savannas: Measurement and Modeling at Landscape to Global Scales, 1st ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- The End of Desertification? Behnke, R.; Mortimore, M. (Eds.) Springer Earth System Sciences; Springer: Berlin, Germany, 2016.
- Hiernaux, P. Implications of the ‘new rangeland paradigm’ for natural resource management. In Proceedings of the 12th Danish Sahel Workshop, Copenhagen, Denmark, 3–5 January 2000; pp. 113–142.
- Kgosikoma, O.E.; Mogotsi, K. Understanding the causes of bush encroachment in Africa: The key to effective management of savanna grasslands. Trop. Grassl. Forrajes Trop. 2013, 1, 215–219. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Height | Phenology | Characteristics | Resprouting | Sites | Palatability | Deep Roots |
|---|---|---|---|---|---|---|---|
| Acacia raddiana | + | Semi-deciduous | Drought resistant | 0 | S | + | 0 |
| Commiphora africana | 0 | Deciduous | Fire and termite resistant | - | S | + | - |
| Balanites aegyptiaca | 0 | Evergreen | Drought resistant | + | S, F | + | 0 |
| Boscia senegalensis | - | Evergreen | Pioneer | 0 | F | + | - |
| Guiera senegalensis | - | Semi-deciduous | Pioneer | +++ | F, S | 0 | 0 |
| Grewia biclor | 0 | Deciduous | Drought resistant | + | S, F | + | - |
| Combretum glutinosum | 0 | Evergreen | Drought and fire resistant | ++ | S, F | 0 | 0 |
| Combretum micranthum | - | Deciduous | Drought and fire resistant | 0 | F | 0 | 0 |
| Pterocarpus lucens | 0 | Deciduous | Fire resistant | 0 | F | + | 0 |
| Soils | No Lag | 1 Year Lag | 2 Year Lag | 3 Year Lag | 2 Year Sum | 3 Year Sum | 4 Year Sum | 5 Year Sum | 6 Year Sum | 7 Year Sum |
|---|---|---|---|---|---|---|---|---|---|---|
| All | 0.51 | 0.58 | 0.61 | 0.54 | 0.58 | 0.63 | 0.69 | 0.71 | 0.73 | 0.72 |
| Sandy | 0.61 | 0.62 | 0.7 | 0.6 | 0.67 | 0.7 | 0.79 | 0.79 | 0.82 | 0.84 |
| Shallow | −0.43 | −0.11 | −0.08 | 0.03 | −0.33 | −0.29 | −0.35 | −0.28 | −0.18 | −0.18 |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brandt, M.; Tappan, G.; Diouf, A.A.; Beye, G.; Mbow, C.; Fensholt, R. Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel. Remote Sens. 2017, 9, 39. https://doi.org/10.3390/rs9010039
Brandt M, Tappan G, Diouf AA, Beye G, Mbow C, Fensholt R. Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel. Remote Sensing. 2017; 9(1):39. https://doi.org/10.3390/rs9010039
Chicago/Turabian StyleBrandt, Martin, Gray Tappan, Abdoul Aziz Diouf, Gora Beye, Cheikh Mbow, and Rasmus Fensholt. 2017. "Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel" Remote Sensing 9, no. 1: 39. https://doi.org/10.3390/rs9010039
APA StyleBrandt, M., Tappan, G., Diouf, A. A., Beye, G., Mbow, C., & Fensholt, R. (2017). Woody Vegetation Die off and Regeneration in Response to Rainfall Variability in the West African Sahel. Remote Sensing, 9(1), 39. https://doi.org/10.3390/rs9010039