
Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin


Abstract
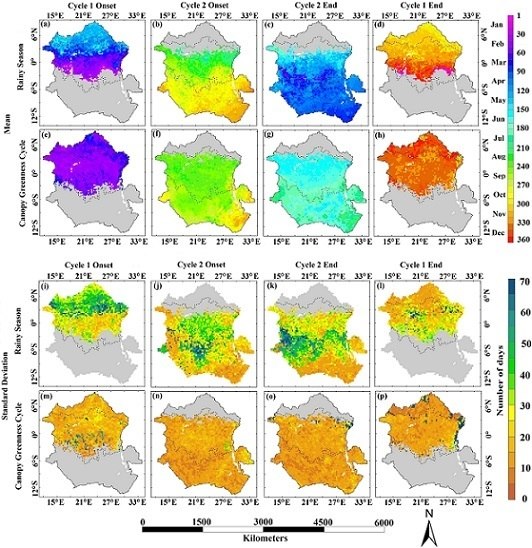
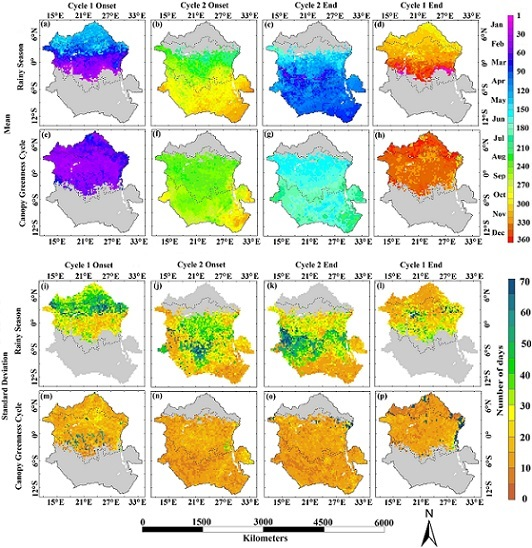
:
{kind=link}
{kind=link}
{kind=link}
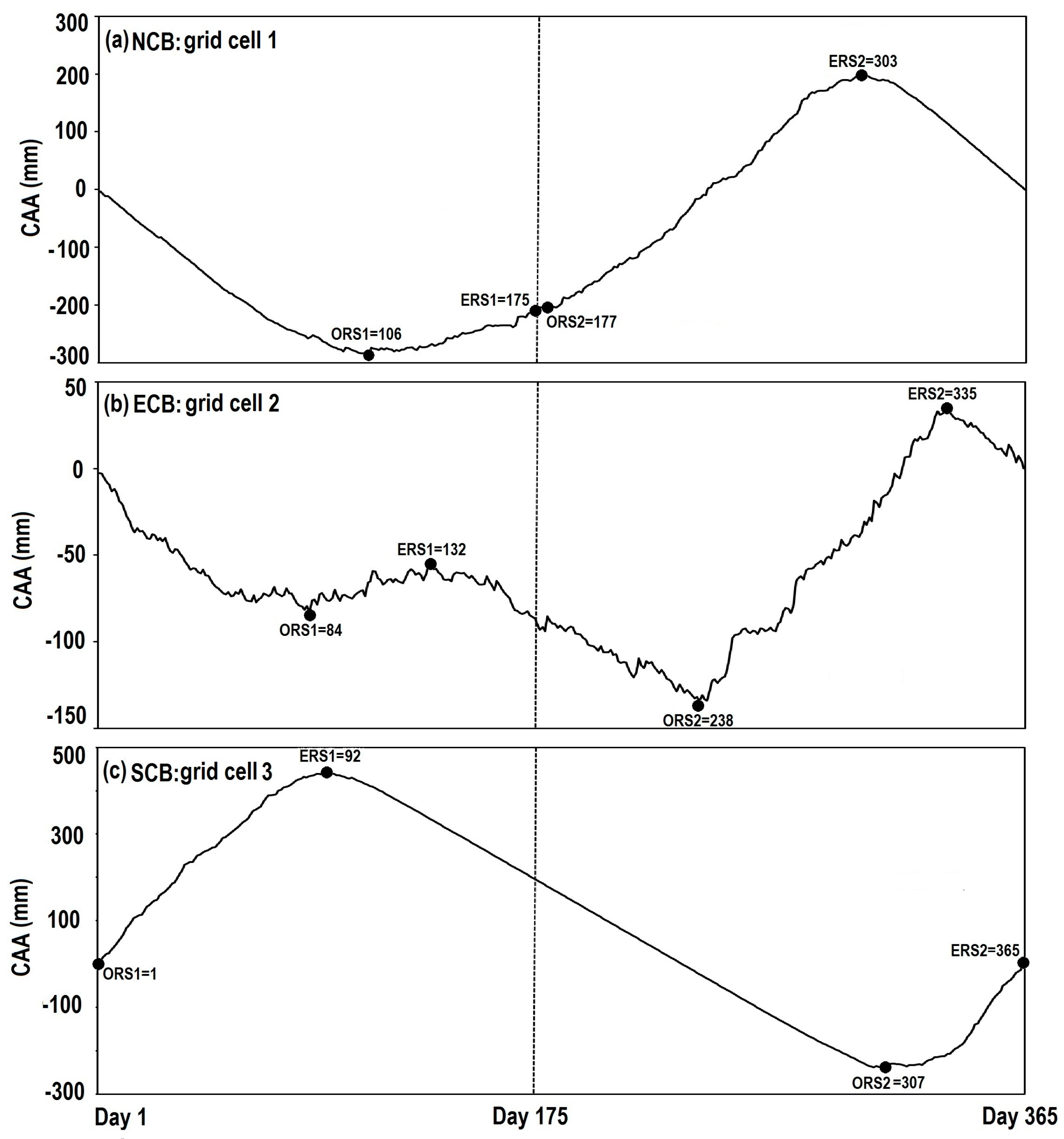{kind=link}
{kind=link}
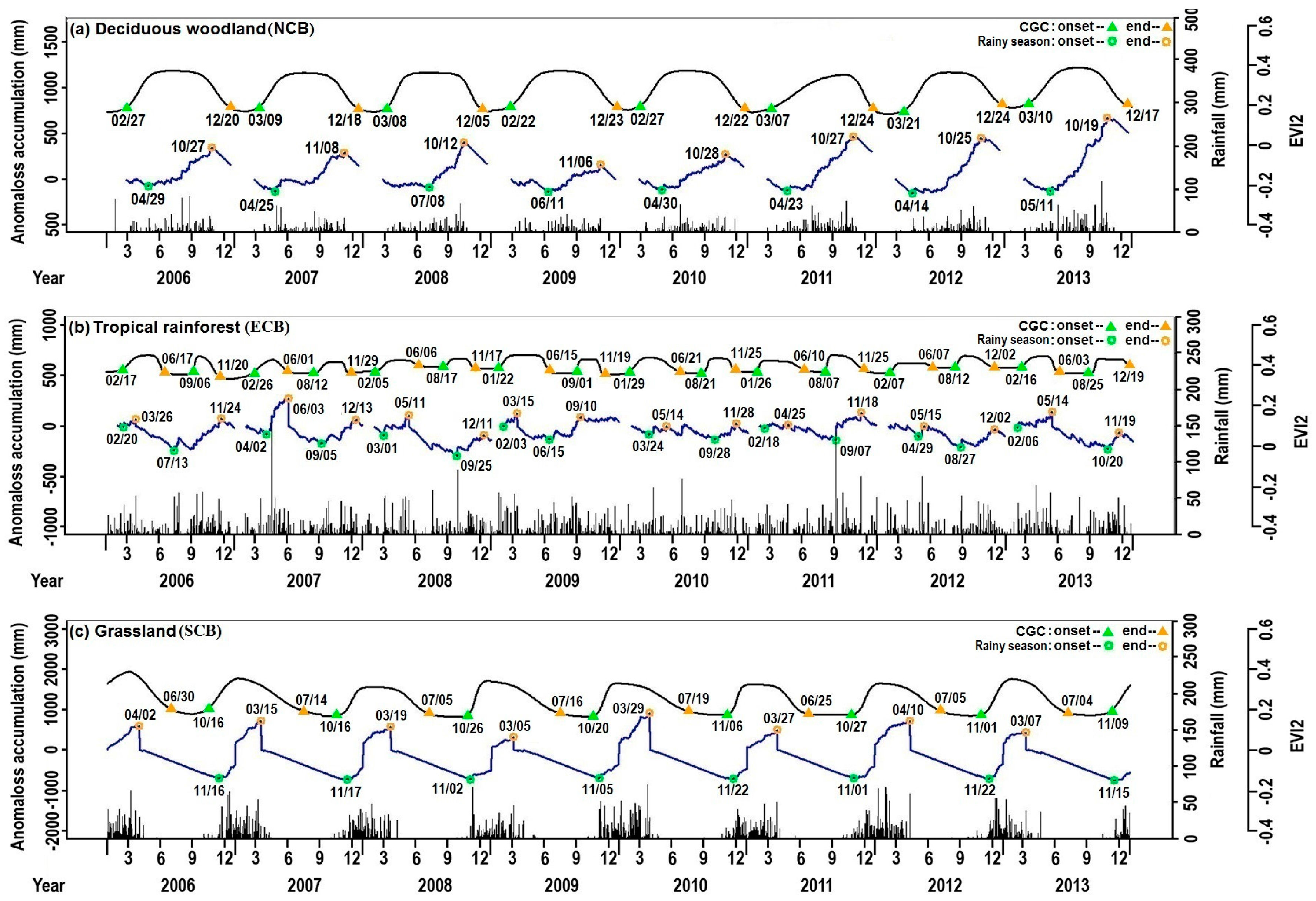{kind=link}
{kind=link}
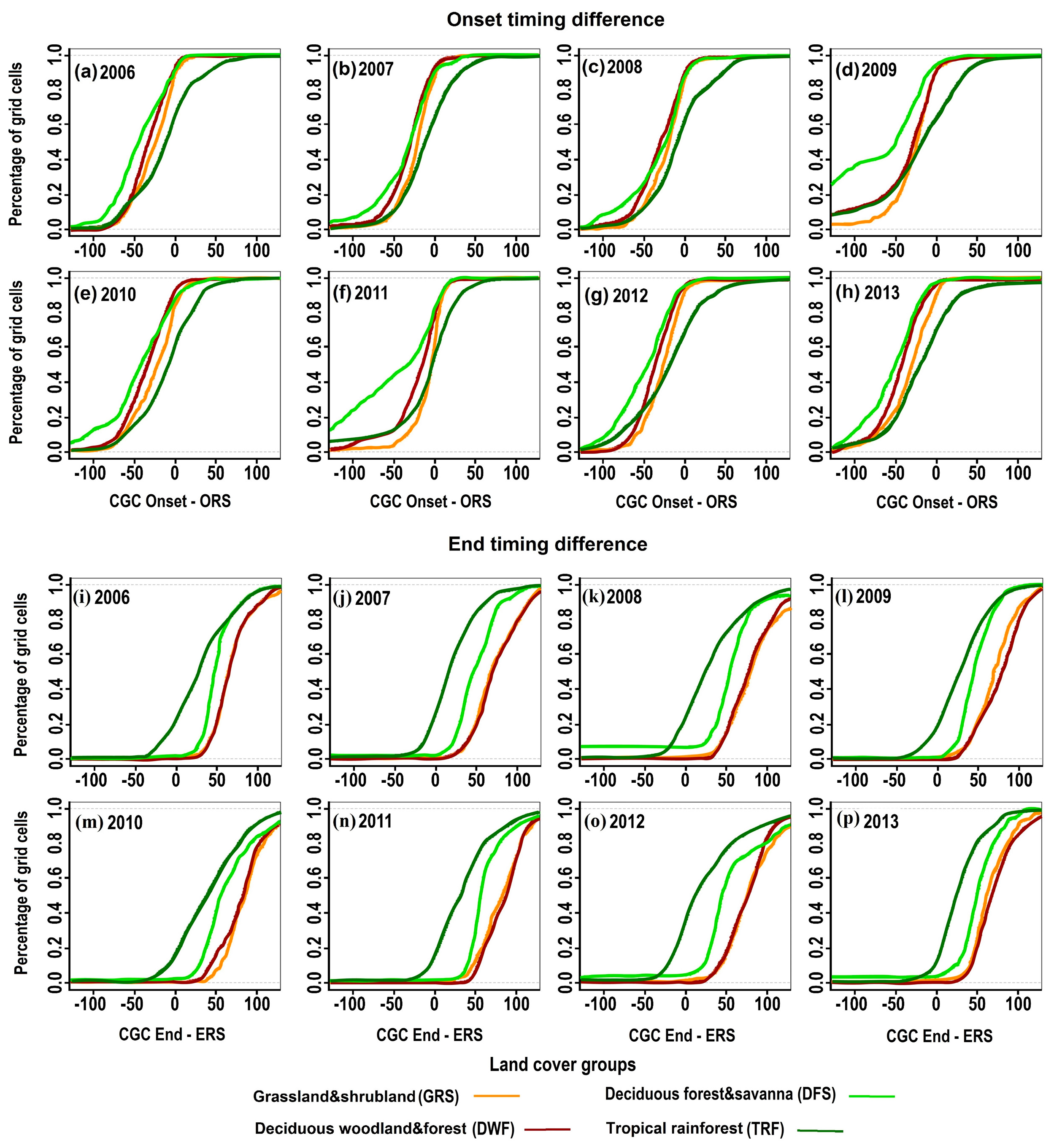{kind=link}
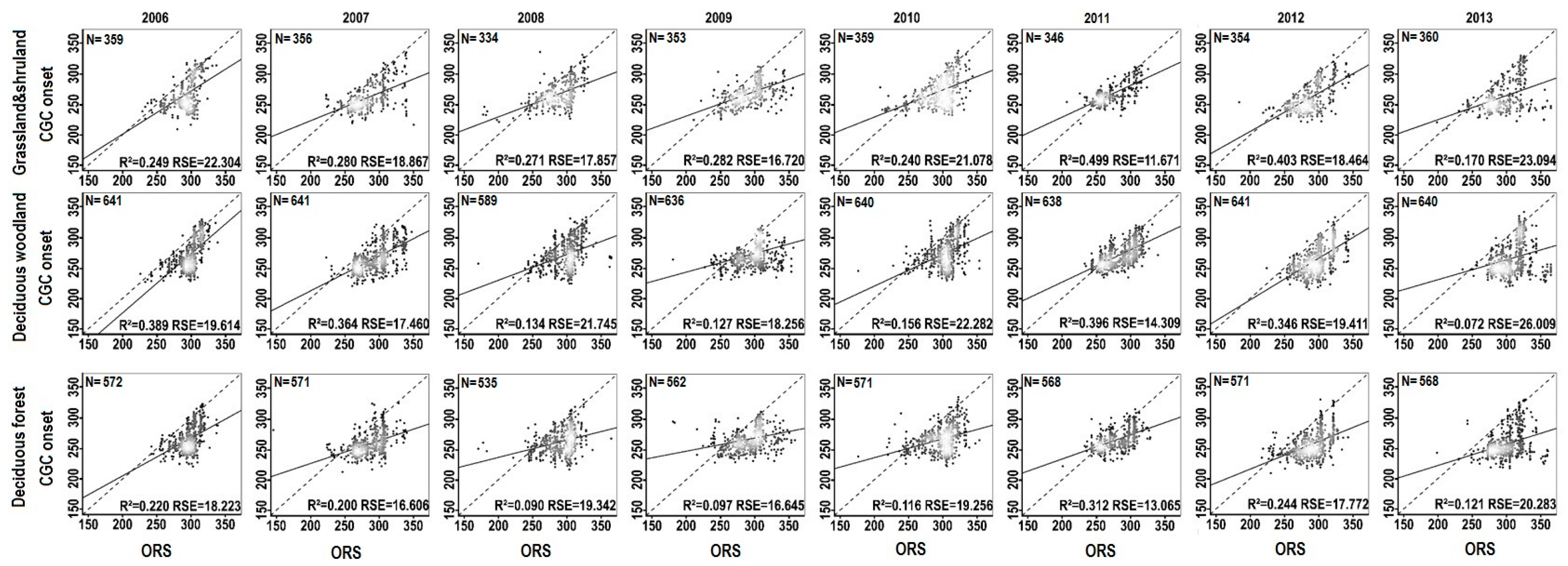{kind=link}
1. Introduction
2. Background
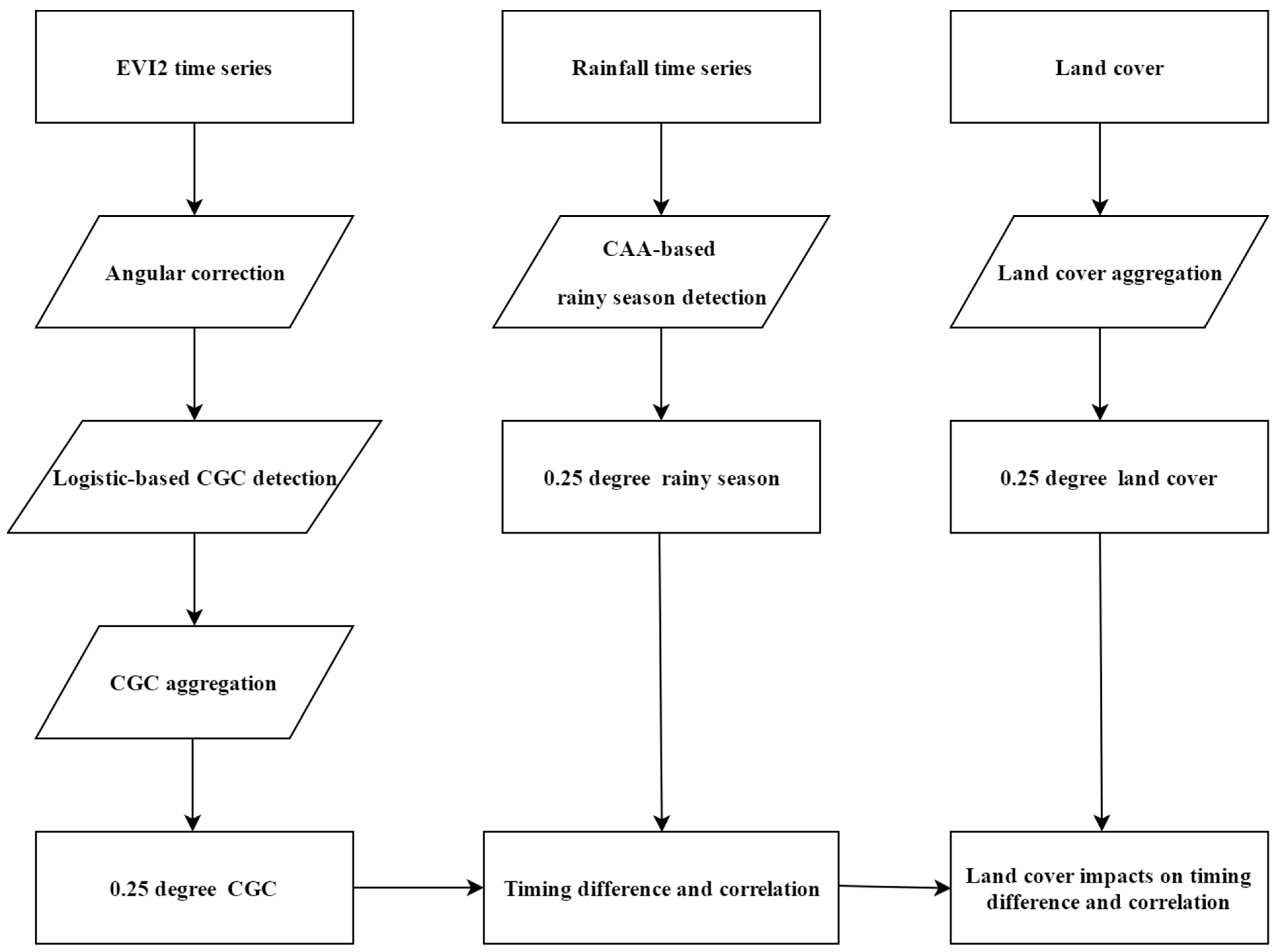
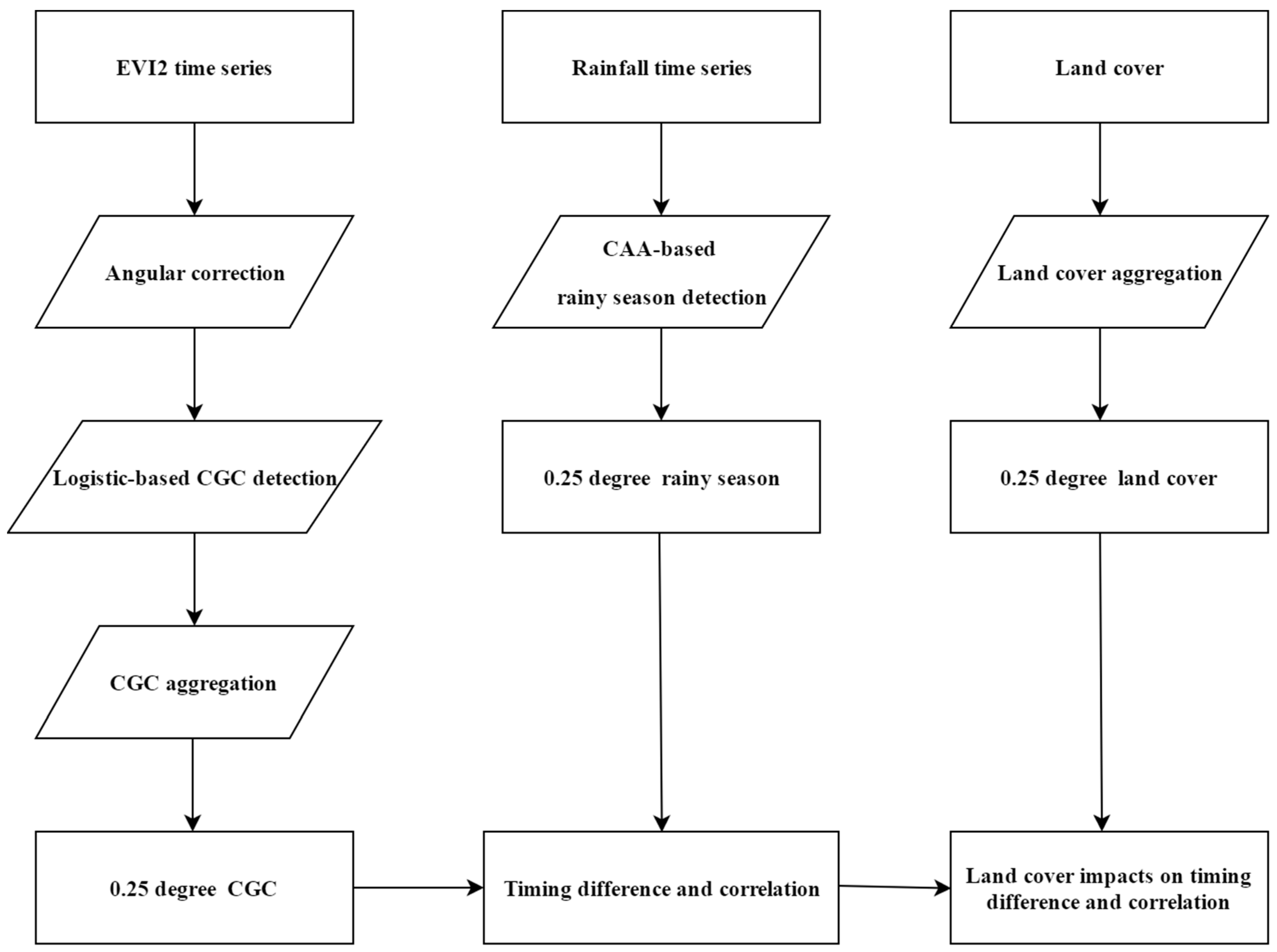
3. Materials and Methods
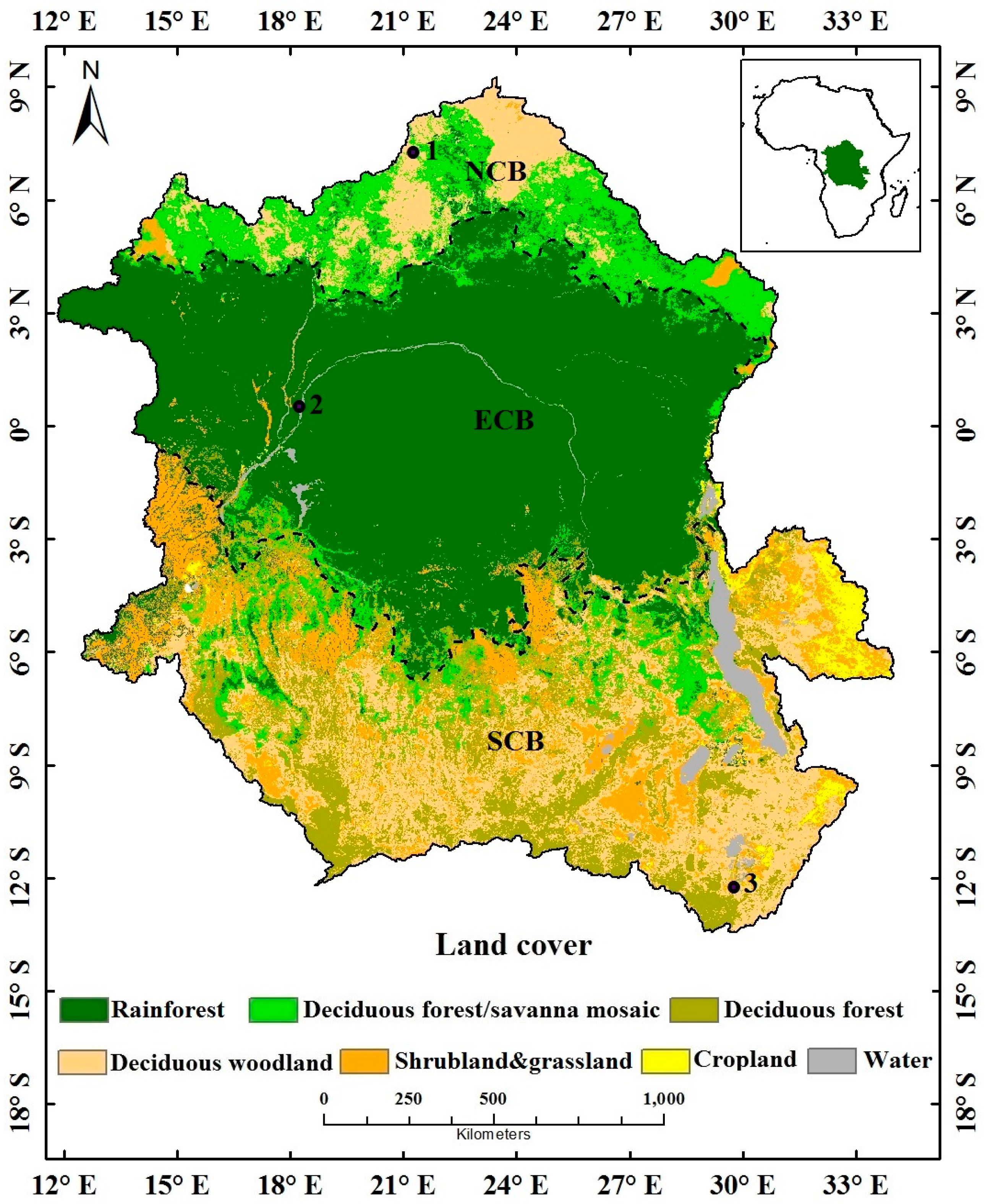
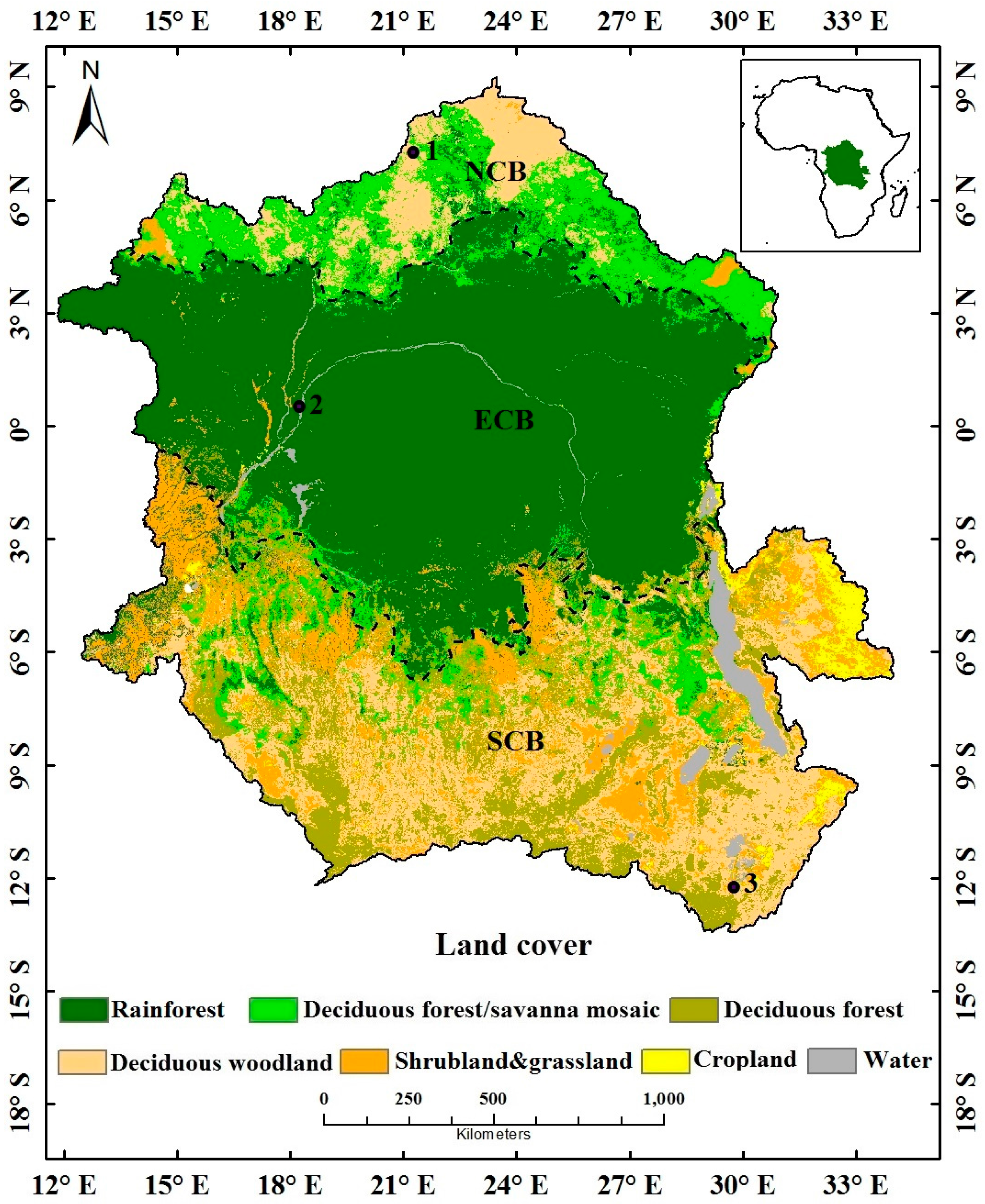
3.1. Land Cover Data
3.2. Detection of the Rainy Season from Tropical Rainfall Measurement Mission Product
3.3. Generation of Angularly Corrected Vegetation Index and Detection of Canopy Creenness Cycles
3.4. Investigation of Land Cover Impacts on LSP Responses to the Rainy Season
4. Results
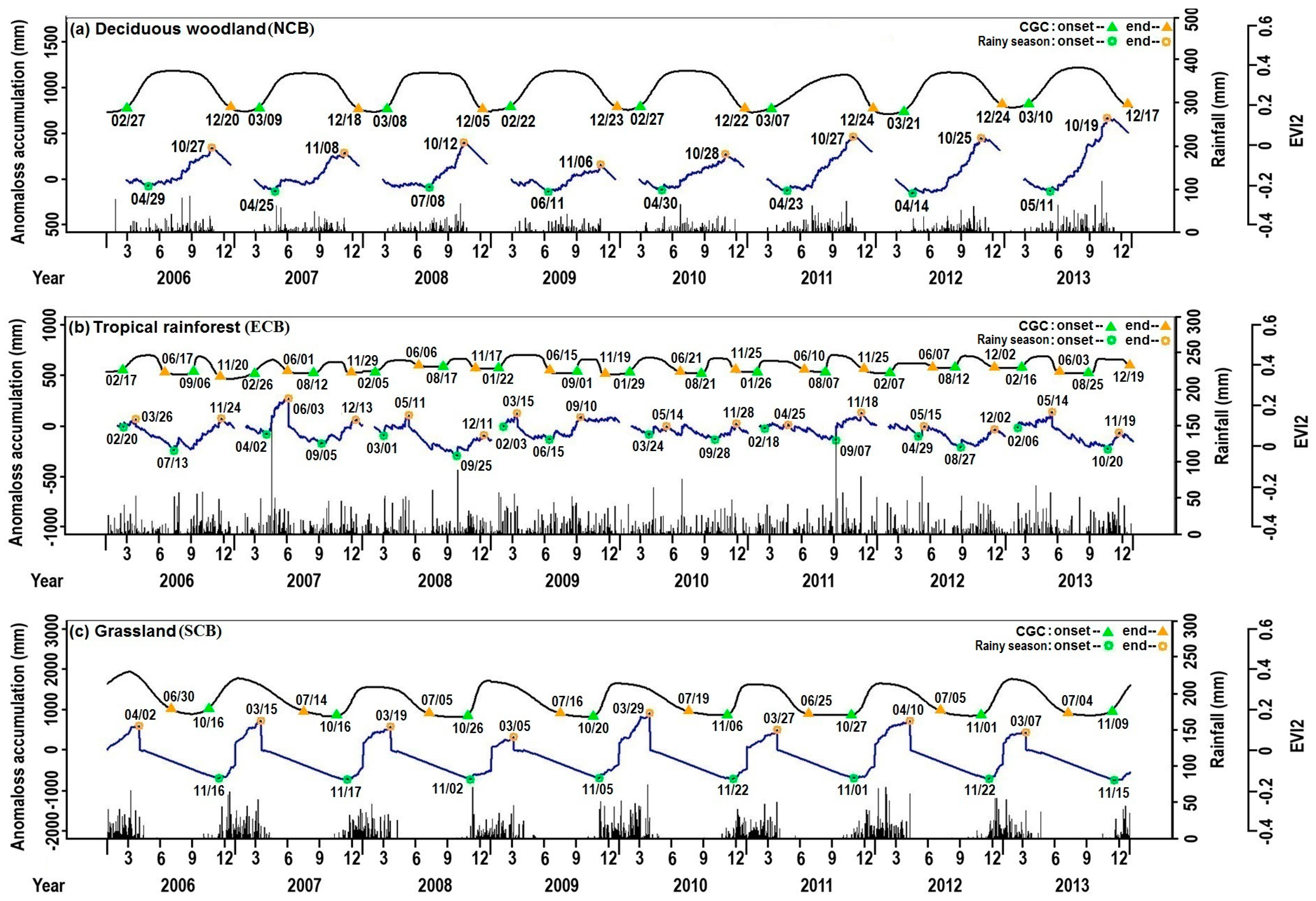
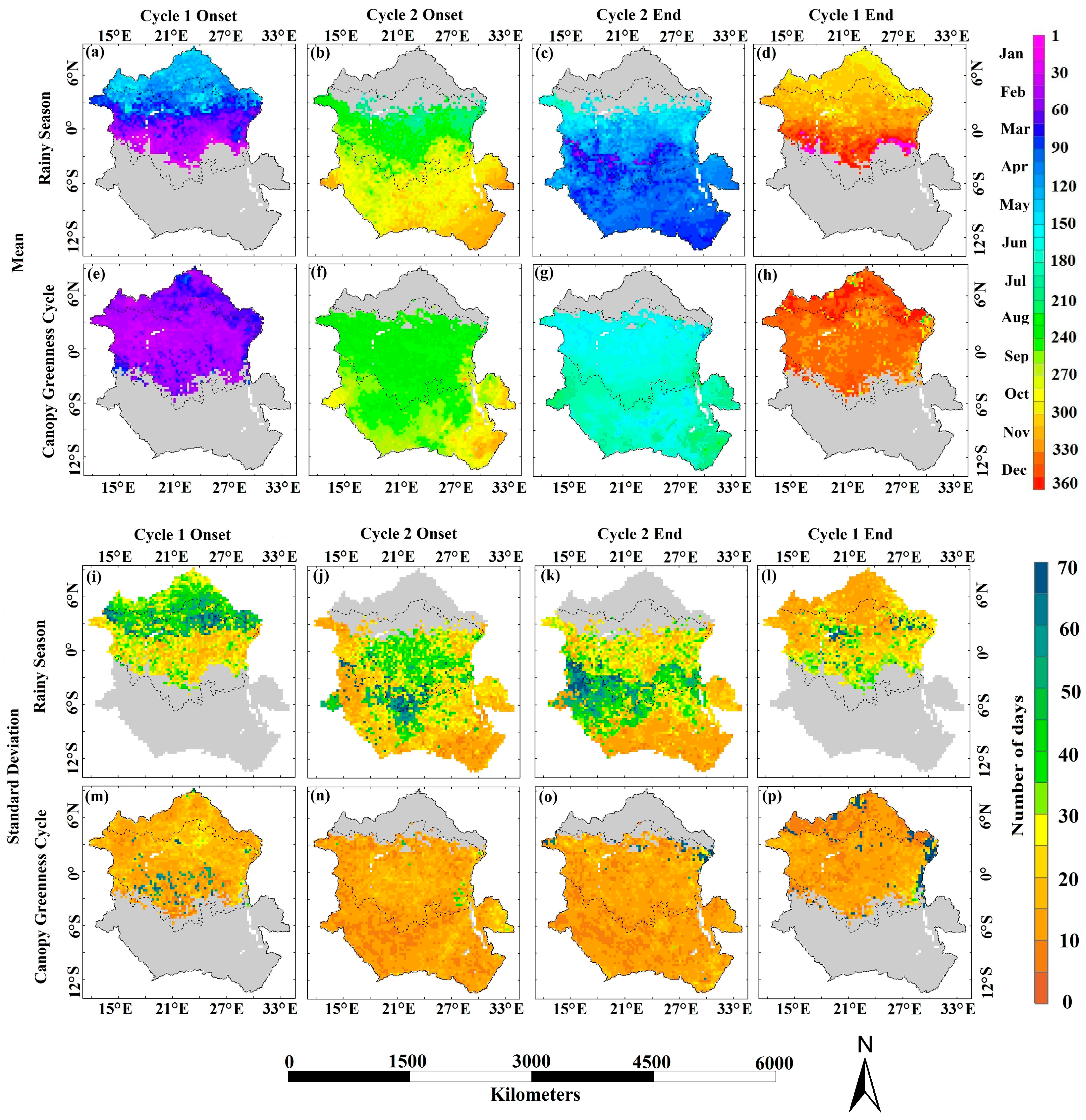
4.1. Inter-Annual Variations in the Timings of Both the Rainy Season and CGC
4.2. Spatial Patterns of the Rainy Season and CGC Timings
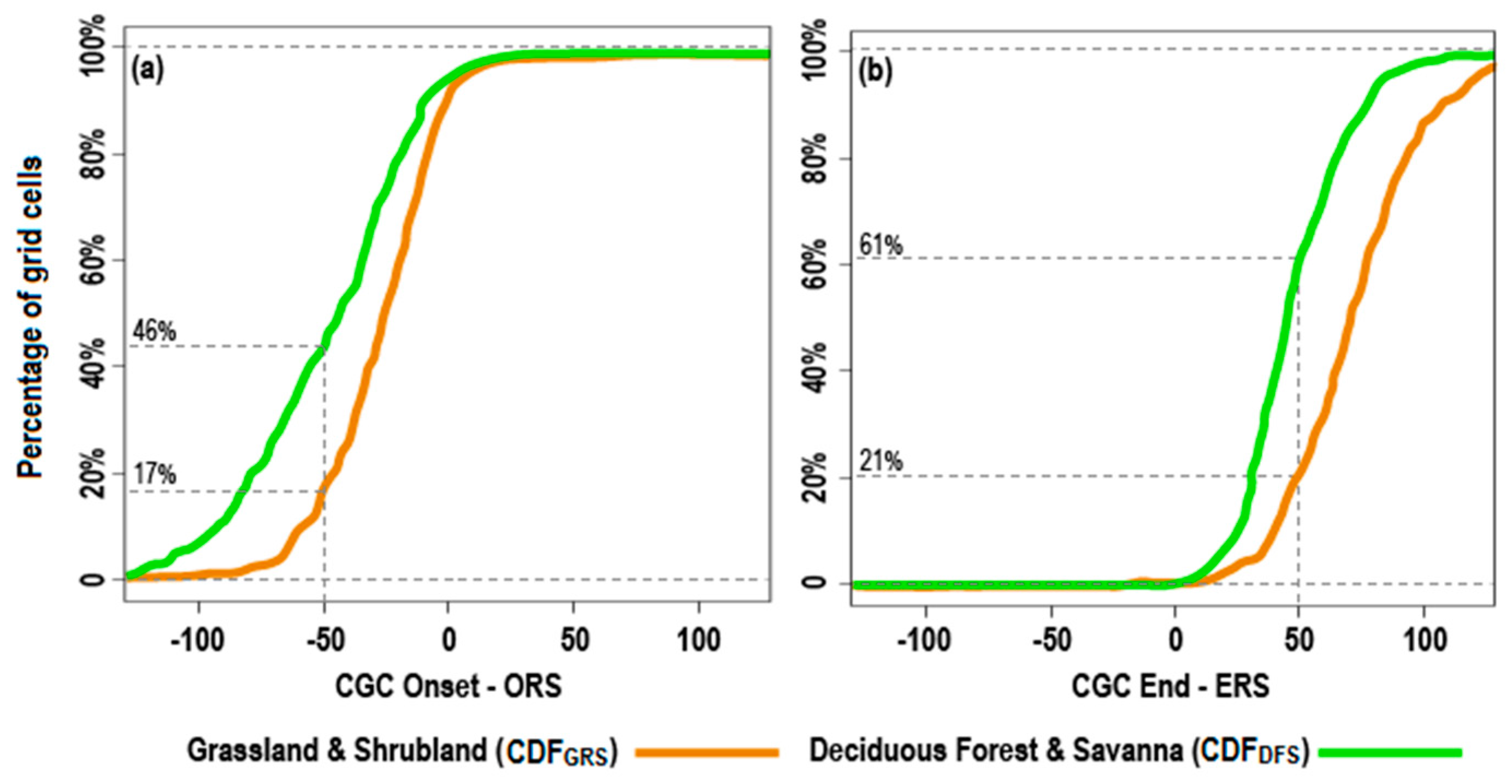
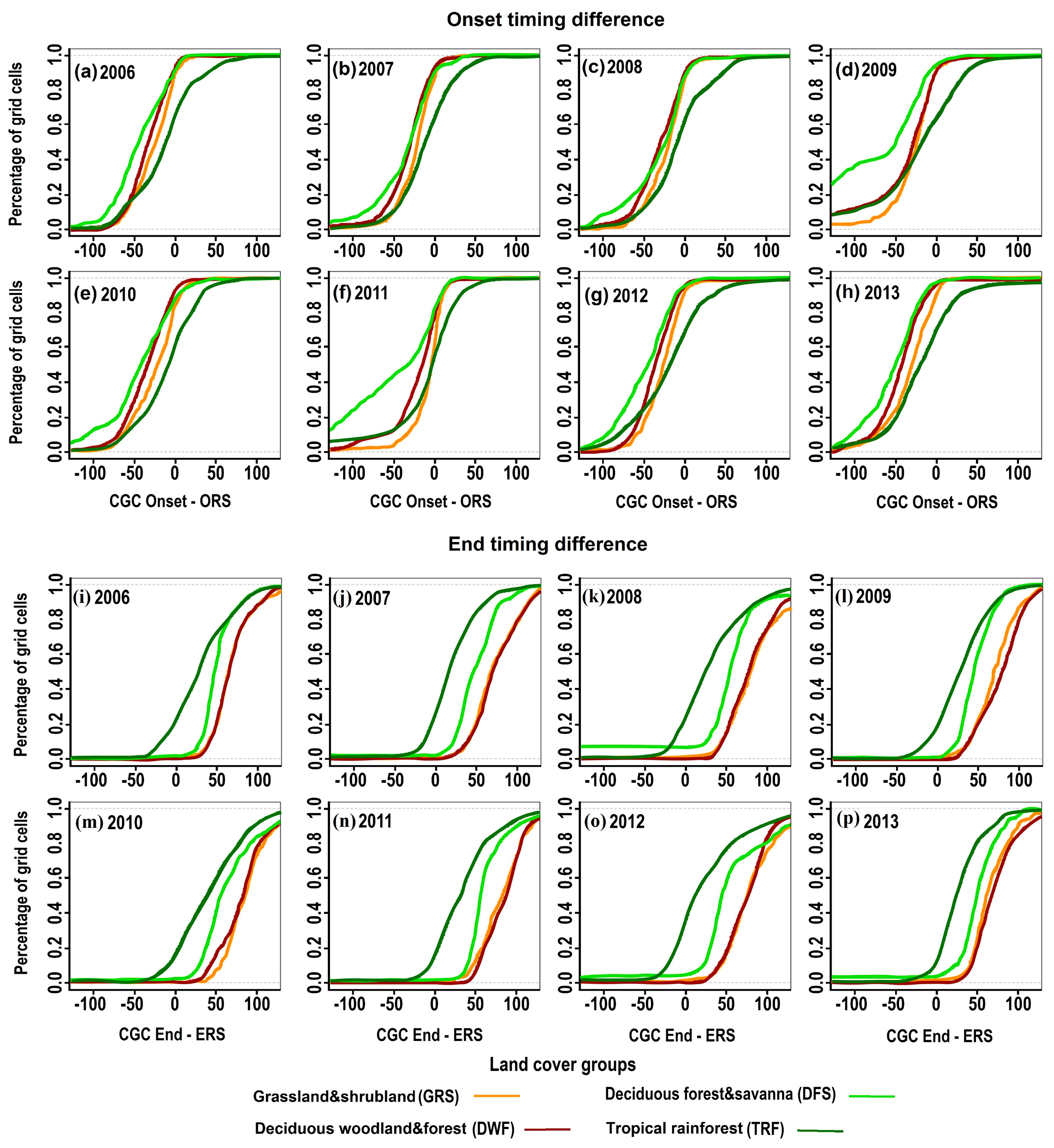
4.3. Impacts of Land Cover on the Responses of CGC Timing to the Rainy Season
5. Discussion
5.1. The Regulation of the Rainy Season by Intertropical Convergence Zone and the Detection of Rainy Season Using the CAA Method
5.2. The Impacts of Land Cover on the Responses of CGC to the Rainy Season
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CAA | Climatological Anomalous Accumulation |
| CDF | Cumulative Distribution Function |
| CGC | Canopy Greenness Cycle |
| DFS | Deciduous Forest and Savanna Mosaic |
| DWF | Deciduous Woodland and Deciduous Forest |
| ECB | Equatorial Congo Basin |
| ERS | End of Rainy Season |
| EVI2 | Two-band Enhanced Vegetation Index |
| GRS | Grassland and Shrubland |
| LSP | Land surface phenology |
| NCB | Northern Congo Basin |
| ORS | Onset of Rainy Season |
| SCB | Southern Congo Basin |
| SEVIRI | Spinning Enhanced Visible and Infrared Imager |
| TRF | tropical rainforest |
References
- Phillips, O.L.; Malhi, Y.; Higuchi, N.; Laurance, W.F.; Núñez, P.V.; Vásquez, R.M.; Laurance, S.G.; Ferreira, L.V.; Stern, M.; Brown, S.; et al. Changes in the carbon balance of tropical forests: Evidence from long-term plots. Science 1998, 282, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y.; Adu-Bredu, S.; Asare, R.A.; Lewis, S.L.; Mayaux, P. African rainforests: Past, present and future. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120312. [Google Scholar] [CrossRef] [PubMed]
- Megevand, C.; Mosnier, A.; Hourticq, J.; Sanders, K.; Doetinchem, N.; Streck, C. Deforestation Trends in the Congo Basin: Reconciling Economic Growth and Forest Protection; World Bank: Washington, DC, USA, 2013. [Google Scholar]
- De Wasseige, C.; De Marcken, P.; Bayol, N.; Hiol Hiol, F.; Mayaux, P.; Desclée, B.; Nasi, R.; Billand, A.; Defourny, P.; Eba’a Atyi, R. The Forests of the Congo Basin—State of the Forest 2010; Publications Office of the European Union: Luxembourg, 2012. [Google Scholar]
- Asefi-Najafabady, S.; Saatchi, S. Response of african humid tropical forests to recent rainfall anomalies. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tian, Y.; Myneni, R.B.; Ciais, P.; Saatchi, S.; Liu, Y.Y.; Piao, S.; Chen, H.; Vermote, E.F.; Song, C.; et al. Widespread decline of congo rainforest greenness in the past decade. Nature 2014, 509, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Fauset, S.; Baker, T.R.; Lewis, S.L.; Feldpausch, T.R.; Affum-Baffoe, K.; Foli, E.G.; Hamer, K.C.; Swaine, M.D. Drought-induced shifts in the floristic and functional composition of tropical forests in ghana. Ecol. Lett. 2012, 15, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.M.; Williams, M.; Grace, J.; Woollen, E.; Lehmann, C.E.R. Pre-rain green-up is ubiquitous across southern tropical africa: Implications for temporal niche separation and model representation. New Phytol. 2017, 213, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Buitenwerf, R.; Rose, L.; Higgins, S.I. Three decades of multi-dimensional change in global leaf phenology. Nat. Clim. Chang. 2015, 5, 364–368. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. War, drought, and phenology: Changes in the land surface phenology of afghanistan since 1982. J. Land Use Sci. 2008, 3, 95–111. [Google Scholar] [CrossRef]
- Guan, K.; Wood, E.F.; Medvigy, D.; Kimball, J.; Pan, M.; Caylor, K.K.; Sheffield, J.; Xu, X.; Jones, M.O. Terrestrial hydrological controls on land surface phenology of african savannas and woodlands. J. Geophys. Res. Biogeosci. 2014, 119, 1652–1669. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in africa by coupling modis and trmm instruments. J. Geophys. Res. Atmos. 2005, 110, D12103. [Google Scholar] [CrossRef]
- Gond, V.; Fayolle, A.; Pennec, A.; Cornu, G.; Mayaux, P.; Camberlin, P.; Doumenge, C.; Fauvet, N.; Gourlet-Fleury, S. Vegetation structure and greenness in central africa from modis multi-temporal data. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120309. [Google Scholar] [CrossRef] [PubMed]
- Verhegghen, A.; Mayaux, P.; De Wasseige, C.; Defourny, P. Mapping congo basin vegetation types from 300 m and 1 km multi-sensor time series for carbon stocks and forest areas estimation. Biogeosciences 2012, 9, 5061–5079. [Google Scholar] [CrossRef]
- Guan, K.; Medvigy, D.; Wood, E.F.; Caylor, K.K.; Shi, L.; Su-Jong, J. Deriving vegetation phenological time and trajectory information over africa using seviri daily lai. IEEE Trans. Geosci. Remote Sens. 2014, 52, 1113–1130. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, X.; Yu, Y.; Guo, W. A comparison of tropical rainforest phenology retrieved from geostationary (seviri) and polar-orbiting (modis) sensors across the congo basin. IEEE Trans. Geosci. Remote Sens. 2016, 54, 4867–4881. [Google Scholar] [CrossRef]
- Sobrino, J.S.; Julien, Y.; Soria, G. Phenology estimation from meteosat second generation data. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2013, 6, 1653–1659. [Google Scholar] [CrossRef]
- Herrmann, S.M.; Mohr, K.I. A continental-acale classification of rainfall seasonality regimes in africa based on gridded precipitation and land surface temperature products. J. Appl. Meteorol. Climatol. 2011, 50, 2504–2513. [Google Scholar] [CrossRef]
- Liebmann, B.; Bladé, I.; Kiladis, G.N.; Carvalho, L.M.V.; Senay, G.B.; Allured, D.; Leroux, S.; Funk, C. Seasonality of african precipitation from 1996 to 2009. J. Clim. 2012, 25, 4304–4322. [Google Scholar] [CrossRef]
- Washington, R.; James, R.; Pearce, H.; Pokam, W.M.; Moufouma-Okia, W. Congo basin rainfall climatology: Can we believe the climate models? Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120296. [Google Scholar] [CrossRef] [PubMed]
- Mayaux, P.; Bartholomé, E.; Fritz, S.; Belward, A. A new land-cover map of africa for the year 2000. J. Biogeogr. 2004, 31, 861–877. [Google Scholar] [CrossRef]
- Couralet, C.; Van den Bulcke, J.; Ngoma, L.M.; Van Acker, J.; Beeckman, H. Phenology in functional groups of central african rainforest trees. J. Trop. For. Sci. 2013, 25, 361–374. [Google Scholar]
- Viennois, G.; Barbier, N.; Fabre, I.; Couteron, P. Multiresolution quantification of deciduousness in west-central african forests. Biogeosciences 2013, 10, 6957–6967. [Google Scholar] [CrossRef]
- De Weirdt, M.; Verbeeck, H.; Maignan, F.; Peylin, P.; Poulter, B.; Bonal, D.; Ciais, P.; Steppe, K. Seasonal leaf dynamics for tropical evergreen forests in a process-based global ecosystem model. Geosci. Model Dev. 2012, 5, 1091–1108. [Google Scholar] [CrossRef]
- Myneni, R.B.; Yang, W.; Nemani, R.R.; Huete, A.R.; Dickinson, R.E.; Knyazikhin, Y.; Didan, K.; Fu, R.; Negrón Juárez, R.I.; Saatchi, S.S.; et al. Large seasonal swings in leaf area of amazon rainforests. Proc. Natl Acad. Sci. USA 2007, 104, 4820–4823. [Google Scholar] [CrossRef] [PubMed]
- Friedl, M.A.; Sulla-Menashe, D.; Tan, B.; Schneider, A.; Ramankutty, N.; Sibley, A.; Huang, X. Modis collection 5 global land cover: Algorithm refinements and characterization of new datasets. Remote Sens. Environ. 2010, 114, 168–182. [Google Scholar] [CrossRef]
- Defourny, P.; Vancutsem, C.; Bicheron, P.; Brockmann, C.; Niño, F.; Schouten, L.; Leroy, M.M. Globcover: A 300 m global land cover product for 2005 using envisat meris time series. In Proceedings of the ISPRS Comission VII Mid-Term Symposium: Remote Sensing: From Pixels to Processes, Enschede, The Netherlands, 6 December 2006; pp. 1–4. [Google Scholar]
- Mayaux, P.; Eva, H.; Gallego, J.; Strahler, A.H.; Herold, M.; Agrawal, S.; Naumov, S.; Miranda, E.E.D.; Bella, C.M.D.; Ordoyne, C.; et al. Validation of the global land cover 2000 map. IEEE Trans. Geosc. Remote Sens. 2006, 44, 1728–1739. [Google Scholar] [CrossRef]
- Bai, L. Comparison and Validation of Five Land Cover Products over the African Continent; Lund University: Lund, Sweden, 2010. [Google Scholar]
- Milewski, A.; Elkadiri, R.; Durham, M. Assessment and comparison of tmpa satellite precipitation products in varying climatic and topographic regimes in morocco. Remote Sens. 2015, 7, 5697. [Google Scholar] [CrossRef]
- Brown, M.E.; de Beurs, K.M. Evaluation of multi-sensor semi-arid crop season parameters based on ndvi and rainfall. Remote Sens. Environ. 2008, 112, 2261–2271. [Google Scholar] [CrossRef]
- Fensholt, R.; Sandholt, I.; Stisen, S.; Tucker, C. Analysing ndvi for the african continent using the geostationary meteosat second generation seviri sensor. Remote Sens. Environ. 2006, 101, 212–229. [Google Scholar] [CrossRef]
- Jiang, Z.; Huete, A.R.; Didan, K.; Miura, T. Development of a two-band enhanced vegetation index without a blue band. Remote Sens. Environ. 2008, 112, 3833–3845. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the modis vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Tian, Y.; Romanov, P.; Yunyue, Y.; Hui, X.; Tarpley, D. Analysis of vegetation index ndvi anisotropy to improve the accuracy of the goes-r green vegetation fraction product. In Proceedings of the Geoscience and Remote Sensing Symposium (IGARSS), Honolulu, HI, USA, 25–30 July 2010; IEEE: New York, NY, USA, 2010; pp. 2091–2094. [Google Scholar]
- Zhang, X. Reconstruction of a complete global time series of daily vegetation index trajectory from long-term avhrr data. Remote Sens. Environ. 2015, 156, 457–472. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using modis. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Arnold, T.B.; Emerson, J.W. Nonparametric goodness-of-fit tests for discrete null distributions. R J. 2011, 3, 34–39. [Google Scholar]
- Haensler, A.; Saeed, F.; Jacob, D. Assessment of projected climate change signals over central africa based on a multitude of global and regional climate projections. In Climate Change Scenarios for the Congo Basin; Climate Service Centre Report No. 11; Haensler, A., Jacob, D., Kabat, P., Ludwig, F., Eds.; Climate Service Center: Hamburg, Germany, 2013. [Google Scholar]
- Tadross, M.A.; Hewitson, B.C.; Usman, M.T. The interannual variability of the onset of the maize growing season over south africa and zimbabwe. J. Clim. 2005, 18, 3356–3372. [Google Scholar] [CrossRef]
- AGRHYMET. Méthodologie de Suivi Des Zones À Risque; Centre Regional AGRHYMET: Niamey, Niger, 1996. [Google Scholar]
- Guan, K.; Pan, M.; Li, H.; Wolf, A.; Wu, J.; Medvigy, D.; Caylor, K.K.; Sheffield, J.; Wood, E.F.; Malhi, Y.; et al. Photosynthetic seasonality of global tropical forests constrained by hydroclimate. Nat. Geosci. 2015, 8, 284–289. [Google Scholar] [CrossRef]
- Guan, K.; Wolf, A.; Medvigy, D.; Caylor, K.K.; Pan, M.; Wood, E.F. Seasonal coupling of canopy structure and function in african tropical forests and its environmental controls. Ecosphere 2013, 4, 1–21. [Google Scholar] [CrossRef]
- Ryan, C.M.; Williams, M.; Hill, T.C.; Grace, J.; Woodhouse, I.H. Assessing the phenology of southern tropical africa: A comparison of hemispherical photography, scatterometry, and optical/nir remote sensing. IEEE Trans. Geosci. Remote Sens. 2014, 52, 519–528. [Google Scholar] [CrossRef]
- Higgins, S.I.; Delgado-Cartay, M.D.; February, E.C.; Combrink, H.J. Is there a temporal niche separation in the leaf phenology of savanna trees and grasses? J. Biogeogr. 2011, 38, 2165–2175. [Google Scholar] [CrossRef]
- Archibald, S.; Scholes, R.J. Leaf green-up in a semi-arid african savanna: Separating tree and grass responses to environmental cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar]
- Borchert, R. Soil and stem water storage determine phenology and distribution of tropical dry forest trees. Ecology 1994, 75, 1437–1449. [Google Scholar] [CrossRef]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, D.; Zhang, X.; Yu, Y.; Guo, W. Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin. Remote Sens. 2017, 9, 461. https://doi.org/10.3390/rs9050461
Yan D, Zhang X, Yu Y, Guo W. Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin. Remote Sensing. 2017; 9(5):461. https://doi.org/10.3390/rs9050461
Chicago/Turabian StyleYan, Dong, Xiaoyang Zhang, Yunyue Yu, and Wei Guo. 2017. "Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin" Remote Sensing 9, no. 5: 461. https://doi.org/10.3390/rs9050461
APA StyleYan, D., Zhang, X., Yu, Y., & Guo, W. (2017). Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin. Remote Sensing, 9(5), 461. https://doi.org/10.3390/rs9050461