Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients

 ,
,  ,
,  ,
,  and
and 
Abstract
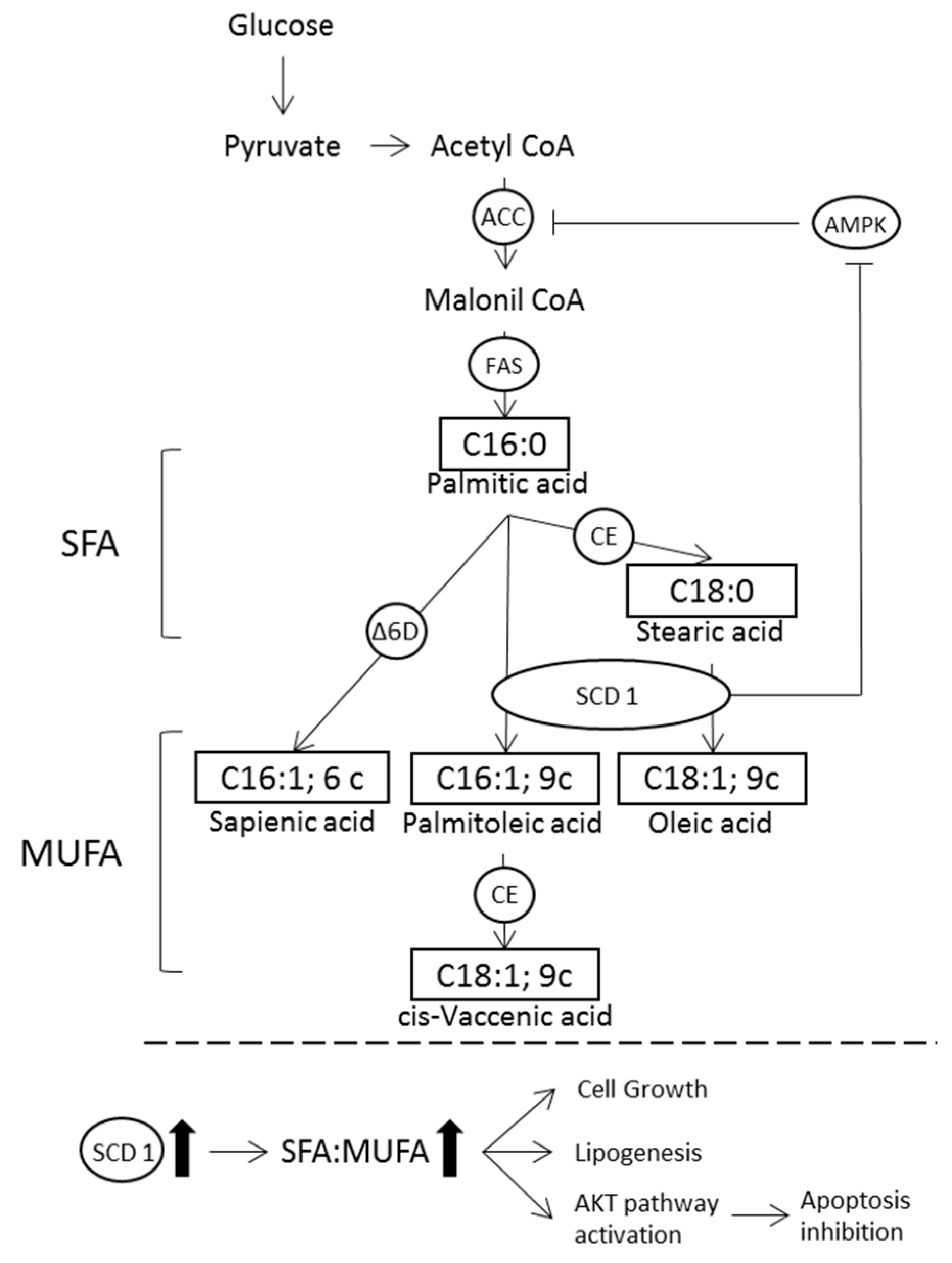
:1. Introduction
2. Materials and Methods
2.1. Subjects and Study Design
2.2. Nutritional Status and Dietary Intake
2.3. Red Blood Cell Membrane Fatty Acid Profile Analysis
2.4. Membrane Fatty Acid Cluster
2.5. Statistical Analysis
3. Results
3.1. Population Characteristics
3.2. Dietary Intake
3.3. Red blood cell Membrane Fatty Acid Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stewart, B.W.; Wild, C. International Agency for Research on Cancer, World Health Organization. World Cancer Rep. 2014, 14, 630. [Google Scholar]
- Mayne, S.T.; Playdon, M.C.; Rock, C.L. Diet, nutrition, and cancer: Past, present and future. Nat. Rev. Clin. Oncol. 2016, 13, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.; Wakefield, C.E.; Laing, D.G. Smell and taste disorders resulting from cancer and chemotherapy. Curr. Pharm. Des. 2016, 22, 2253–2263. [Google Scholar] [CrossRef] [PubMed]
- Bozzetti, F. Nutritional support of the oncology patient. Crit. Rev. Oncol. Hematol. 2013, 87, 172–200. [Google Scholar] [CrossRef] [PubMed]
- Tueros, I.; Uriarte, M. Innovative food products for cancer patients: Future directions. J. Sci. Food Agric. 2018, 98, 1647–1652. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Baenke, F.; Peck, B.; Miess, H.; Schulze, A. Hooked on fat: The role of lipid synthesis in cancer metabolism and tumour development. Dis. Model. Mech. 2013, 6, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Rohrig, F.; Schulze, A. The multifaceted roles of fatty acid synthesis in cancer. Nat. Rev. Cancer. 2016, 16, 732–749. [Google Scholar] [CrossRef] [PubMed]
- Hopperton, K.E.; Duncan, R.E.; Bazinet, R.P.; Archer, M.C. Fatty acid synthase plays a role in cancer metabolism beyond providing fatty acids for phospholipid synthesis or sustaining elevations in glycolytic activity. Exp. Cell. Res. 2014, 320, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Beloribi-Djefaflia, S.; Vasseur, S.; Guillaumond, F. Lipid metabolic reprogramming in cancer cells. Oncogenesis 2016, 5, e189. [Google Scholar] [CrossRef] [PubMed]
- MacLean, C.H.; Newberry, S.J.; Mojica, W.A.; Khanna, P.; Issa, A.M.; Suttorp, M.J.; Lim, Y.W.; Traina, S.B.; Hilton, L.; Garland, R.; et al. Effects of omega-3 fatty acids on cancer risk: A systematic review. JAMA 2006, 295, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Moloudizargari, M.; Mortaz, E.; Asghari, M.H.; Adcock, I.M.; Redegeld, F.A.; Garssen, J. Effects of the polyunsaturated fatty acids, epa and dha, on hematological malignancies: A systematic review. Oncotarget 2018, 9, 11858–11875. [Google Scholar] [CrossRef] [PubMed]
- Khankari, N.K.; Bradshaw, P.T.; Steck, S.E.; He, K.; Olshan, A.F.; Shen, J.; Ahn, J.; Chen, Y.; Ahsan, H.; Terry, M.B.; et al. Polyunsaturated fatty acid interactions and breast cancer incidence: A population-based case-control study on long island, new york. Ann. Epidemiol. 2015, 25, 929–935. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dubois, R.N. Eicosanoids and cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Lands, W.E. Metabolism of glycerolipides; a comparison of lecithin and triglyceride synthesis. J. Biol. Chem. 1958, 231, 883–888. [Google Scholar] [PubMed]
- Nicolson, G.L.; Ash, M.E. Membrane lipid replacement for chronic illnesses, aging and cancer using oral glycerolphospholipid formulations with fructooligosaccharides to restore phospholipid function in cellular membranes, organelles, cells and tissues. Biochim. Biophys. Acta Biomembr. 2017, 9, 18. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.A. Fat and fatty acid intake and metabolic effects in the human body. Ann. Nutr. Metab. 2009, 55, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Chatgilialoglu, C. Membrane Lipidomics for Personalized Health; John Wiley & Sons: Chichester, UK, 2015; pp. 135–155. [Google Scholar]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Ferreri, C.; Chatgilialoglu, C. Role of fatty acid-based functional lipidomics in the development of molecular diagnostic tools. Expert Rev. Mol. Diagn. 2012, 12, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Mikirova, N.; Riordan, H.D.; Jackson, J.A.; Wong, K.; Miranda-Massari, J.R.; Gonzalez, M.J. Erythrocyte membrane fatty acid composition in cancer patients. PR Health Sci. J. 2004, 23, 107–113. [Google Scholar]
- Okuno, M.; Hamazaki, K.; Ogura, T.; Kitade, H.; Matsuura, T.; Yoshida, R.; Hijikawa, T.; Kwon, M.; Arita, S.; Itomura, M.; et al. Abnormalities in fatty acids in plasma, erythrocytes and adipose tissue in japanese patients with colorectal cancer. In Vivo 2013, 27, 203–210. [Google Scholar] [PubMed]
- Pala, V.; Krogh, V.; Muti, P.; Chajes, V.; Riboli, E.; Micheli, A.; Saadatian, M.; Sieri, S.; Berrino, F. Erythrocyte membrane fatty acids and subsequent breast cancer: A prospective italian study. J. Natl. Cancer Inst. 2001, 93, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Coviello, G.; Tutino, V.; Notarnicola, M.; Caruso, M.G. Erythrocyte membrane fatty acids profile in colorectal cancer patients: A preliminary study. Anticancer Res. 2014, 34, 4775–4779. [Google Scholar] [PubMed]
- Denton, M.J.; Spencer, N.; Arnstein, H.R. Biochemical and enzymic changes during erythrocyte differentiation. The significance of the final cell division. Biochem. J. 1975, 146, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghezzo, A.; Visconti, P.; Abruzzo, P.M.; Bolotta, A.; Ferreri, C.; Gobbi, G.; Malisardi, G.; Manfredini, S.; Marini, M.; Nanetti, L.; et al. Oxidative stress and erythrocyte membrane alterations in children with autism: Correlation with clinical features. PloS ONE 2013, 8, e66418. [Google Scholar] [CrossRef] [PubMed]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R.; et al. Hexadecenoic fatty acid isomers in human blood lipids and their relevance for the interpretation of lipidomic profiles. PloS ONE 2016, 11, e0152378. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, G.; Ferreri, C.; Sansone, A.; Chatgilialoglu, C.; Marzetti, C.; Spyratou, E.; Georgakilas, A.G.; Marini, M.; Abruzzo, P.M.; Bolotta, A.; et al. High predictive values of rbc membrane-based diagnostics by biophotonics in an integrated approach for autism spectrum disorders. Sci. Rep. 2017, 7, 9854. [Google Scholar] [CrossRef] [PubMed]
- Amezaga, J.; Alfaro, B.; Rios, Y.; Larraioz, A.; Ugartemendia, G.; Urruticoechea, A.; Tueros, I. Assessing taste and smell alterations in cancer patients undergoing chemotherapy according to treatment. Support. Care cancer 2018, 31, 4077–4086. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ballart, J.D.; Pinol, J.L.; Zazpe, I.; Corella, D.; Carrasco, P.; Toledo, E.; Perez-Bauer, M.; Martinez-Gonzalez, M.A.; Salas-Salvado, J.; Martin-Moreno, J.M. Relative validity of a semi-quantitative food-frequency questionnaire in an elderly mediterranean population of spain. Br. J. Nutr. 2010, 103, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.M.; Lopez-Sobaler, A.M.; Andres, P.; Requejo, A.M.; Aparicio, A.; Molinero, L.M. Dial Software for Assessing Diets and Food Calculations, (for Windows, Version 3.5.0.3); Departament of Nutrition (UCM) & Alce Ingeniería: Madrid, Spain, 2017. [Google Scholar]
- van der Vegt, S.G.; Ruben, A.M.; Werre, J.M.; Palsma, D.M.; Verhoef, C.W.; de Gier, J.; Staal, G.E. Counterflow centrifugation of red cell populations: A cell age related separation technique. Br. J. Haematol. 1985, 61, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Breil, C.; Abert Vian, M.; Zemb, T.; Kunz, W.; Chemat, F. “Bligh and dyer” and folch methods for solid-liquid-liquid extraction of lipids from microorganisms. Comprehension of solvatation mechanisms and towards substitution with alternative solvents. Int. J. Mol. Sci. 2017, 18, 708. [Google Scholar] [CrossRef] [PubMed]
- Pironi, L.; Guidetti, M.; Verrastro, O.; Iacona, C.; Agostini, F.; Pazzeschi, C.; Sasdelli, A.S.; Melchiorre, M.; Ferreri, C. Functional lipidomics in patients on home parenteral nutrition: Effect of lipid emulsions. World J. Gastroenterol. 2017, 23, 4604–4614. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Von Schacky, C. The omega-3 index: A new risk factor for death from coronary heart disease? Prev. Med. 2004, 39, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Abbott, S.K.; Else, P.L.; Atkins, T.A.; Hulbert, A.J. Fatty acid composition of membrane bilayers: Importance of diet polyunsaturated fat balance. Biochim. Biophys. Acta 2012, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Jolliffe, I.T. Principal Component Analysis, 2nd ed.; Springer: New York, NY, 2002; pp. 63–165. [Google Scholar]
- Merino Salvador, M.; Gomez de Cedron, M.; Moreno Rubio, J.; Falagan Martinez, S.; Sanchez Martinez, R.; Casado, E.; Ramirez de Molina, A.; Sereno, M. Lipid metabolism and lung cancer. Crit. Rev. Oncol. Hematol. 2017, 112, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Robey, R.B.; Weisz, J.; Kuemmerle, N.B.; Salzberg, A.C.; Berg, A.; Brown, D.G.; Kubik, L.; Palorini, R.; Al-Mulla, F.; Al-Temaimi, R.; et al. Metabolic reprogramming and dysregulated metabolism: Cause, consequence and/or enabler of environmental carcinogenesis? Carcinogenesis 2015, 36. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R. Membrane phospholipids, lipoxidative damage and molecular integrity: A causal role in aging and longevity. Biochim. Biophys. Acta 2008, 10, 5. [Google Scholar] [CrossRef] [PubMed]
- Maulucci, G.; Cohen, O.; Daniel, B.; Sansone, A.; Petropoulou, P.I.; Filou, S.; Spyridonidis, A.; Pani, G.; De Spirito, M.; Chatgilialoglu, C.; et al. Fatty acid-related modulations of membrane fluidity in cells: Detection and implications. Free Radic. Res. 2016, 50, S40–S50. [Google Scholar] [CrossRef] [PubMed]
- Peck, B.; Schug, Z.T.; Zhang, Q.; Dankworth, B.; Jones, D.T.; Smethurst, E.; Patel, R.; Mason, S.; Jiang, M.; Saunders, R.; et al. Inhibition of fatty acid desaturation is detrimental to cancer cell survival in metabolically compromised environments. Cancer Metab. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Igal, R.A. Stearoyl coa desaturase-1: New insights into a central regulator of cancer metabolism. Biochim. Biophys. Acta 2016, 12, 14. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Loser, K.; Thurmer, M. Stearoyl-coa desaturase-1 and adaptive stress signaling. Biochim. Biophys. Acta 2016, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Fan, X.X.; He, J.; Pan, H.; Li, R.Z.; Huang, L.; Jiang, Z.; Yao, X.J.; Liu, L.; Leung, E.L.; et al. Scd1 is associated with tumor promotion, late stage and poor survival in lung adenocarcinoma. Oncotarget 2016, 7, 39970–39979. [Google Scholar] [CrossRef] [PubMed]
- Cottet, V.; Collin, M.; Gross, A.S.; Boutron-Ruault, M.C.; Morois, S.; Clavel-Chapelon, F.; Chajes, V. Erythrocyte membrane phospholipid fatty acid concentrations and risk of colorectal adenomas: A case-control nested in the french e3n-epic cohort study. Cancer Epidemiol. Biomarkers Prev. 2013, 22, 1417–1427. [Google Scholar] [CrossRef] [PubMed]
- Mancini, R.; Noto, A.; Pisanu, M.E.; De Vitis, C.; Maugeri-Sacca, M.; Ciliberto, G. Metabolic features of cancer stem cells: The emerging role of lipid metabolism. Oncogene 2018, 37, 2367–2378. [Google Scholar] [CrossRef] [PubMed]
- Warensjo, E.; Ohrvall, M.; Vessby, B. Fatty acid composition and estimated desaturase activities are associated with obesity and lifestyle variables in men and women. Nutr. Metab. Cardiovasc. Dis. 2006, 16, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Tahergorabi, Z.; Khazaei, M.; Moodi, M.; Chamani, E. From obesity to cancer: A review on proposed mechanisms. Cell Biochem. Funct. 2016, 34, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Kyrgiou, M.; Kalliala, I.; Markozannes, G.; Gunter, M.J.; Paraskevaidis, E.; Gabra, H.; Martin-Hirsch, P.; Tsilidis, K.K. Adiposity and cancer at major anatomical sites: Umbrella review of the literature. BMJ 2017. [Google Scholar] [CrossRef] [PubMed]
- Merino, D.M.; Ma, D.W.; Mutch, D.M. Genetic variation in lipid desaturases and its impact on the development of human disease. Lipids Health Dis. 2010, 9, 9–63. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-Y.; Chapkin, R.S. Importance of dietary γ-linolenic acid in human health and nutrition. J. Nutr. 1998, 128, 1411–1414. [Google Scholar] [CrossRef] [PubMed]
- Vernieri, C.; Nichetti, F.; Raimondi, A.; Pusceddu, S.; Platania, M.; Berrino, F.; de Braud, F. Diet and supplements in cancer prevention and treatment: Clinical evidences and future perspectives. Crit. Rev. Oncol. Hematol. 2018, 123, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Ma, J.; Campos, H.; Hankinson, S.E.; Hu, F.B. Comparison between plasma and erythrocyte fatty acid content as biomarkers of fatty acid intake in us women. Am. J. Clin. Nutr. 2007, 86, 74–81. [Google Scholar] [CrossRef] [PubMed]
- von Roemeling, C.A.; Caulfield, T.R.; Marlow, L.; Bok, I.; Wen, J.; Miller, J.L.; Hughes, R.; Hazlehurst, L.; Pinkerton, A.B.; Radisky, D.C.; et al. Accelerated bottom-up drug design platform enables the discovery of novel stearoyl-coa desaturase 1 inhibitors for cancer therapy. Oncotarget 2017, 9, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Benesch, M.G.K.; MacIntyre, I.T.K.; McMullen, T.P.W.; Brindley, D.N. Coming of age for autotaxin and lysophosphatidate signaling: Clinical applications for preventing, detecting and targeting tumor-promoting inflammation. Cancers 2018, 10, 73. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Controls | Patients | p-value | |
|---|---|---|---|
| n (%) | n (%) | ||
| Gender | 0.578 | ||
| Men | 9 (24) | 16 (30) | |
| Women | 28 (76) | 38 (70) | |
| Age (years) (median, Q1–Q3) | 42 (32–57) | 59 (51–64) | <0.001 |
| BMI (kg/m2) (mean ± SD) | 22.7 ± 4.0 | 24.6 ± 4.7 | 0.124 |
| Cancer diagnosis | |||
| Breast | - | 20 (37) | - |
| Colon | - | 13 (24) | - |
| Lung | - | 9 (17) | - |
| Prostate | - | 4 (7) | - |
| Ovarian | - | 2 (4) | - |
| Lymphoma | - | 1 (2) | - |
| Others | - | 5 (9) | - |
| Chemotherapy regimen | |||
| Paclitaxel | - | 10 (19) | - |
| Oxaliplatin-based regimens | - | 12 (22) | - |
| Docetaxel | - | 4 (8) | - |
| Carboplatin | - | 6 (11) | - |
| Anthracyclines (doxorubicin, TAC, AC, CHOP) | - | 5 (9) | - |
| Cisplatin (with pemetrexed or gemcitabine) | - | 6 (11) | - |
| 5-Fluorouracil | - | 2 (4) | - |
| Vinorelbine | - | 4 (7) | |
| Others | - | 5 (9) | - |
| Total Nutrient/Food Intake | |||
|---|---|---|---|
| Control | Cancer | p | |
| Daily energy and nutrient intake | |||
| Calories (Kcal/day) | 1716 (1475–2052) | 1568 (1306–1989) | 0.304 |
| Carbohydrates (% of energy/day) * | 37 (35–42) | 35 (33–39) | 0.025 |
| Simple sugars (g/day) | 90 (74–106) | 42 (26–57) | <0.001 |
| Protein (% of energy/day) | 18 (16–19) | 19 (17–21) | 0.142 |
| Fiber (g/day) * | 25 (21–27) | 17 (12–22) | <0.001 |
| Alcohol (g/day) * | 6 (2–10) | 1 (0–5) | <0.001 |
| Fat (% of energy/day) | 39 (33–42) | 42 (39–45) | 0.006 |
| Total SFA (% of energy/day) | 11 (9–12) | 11 (9–13) | 0.773 |
| C14:0 (g/day) | 1.6 (1.1–2.0) | 1.4 (0.7–2.2) | 0.541 |
| C16:0 (g/day) * | 12 (9–14) | 9 (7–12) | 0.004 |
| C18:0 (g/day) * | 4.4 (3.7–6.2) | 3.4 (2.2–4.6) | <0.001 |
| Total MUFA (% of energy/day) | 5 (5–8) | 7 (6–8) | 0.011 |
| C16:1 (g/day) * | 1 (0.8–1.3) | 1.1 (0.8–1.5) | 0.547 |
| C18:1 (g/day) * | 34 (29–36) | 33 (26–39) | 0.771 |
| Total PUFA (% of energy/day) * | 5.4 (4.7–7.6) | 6.6 (5.8–7.8) | 0.008 |
| Total Omega 3 (% of energy/day) * | 1.0 (0.9–1.2) | 1.2 (1.0–1.4) | 0.004 |
| C18:3 (g/day) * | 1.6 (1.3–2.5) | 1.1 (0.8–1.5) | <0.001 |
| C20:5 (g/day) * | 0.09 (0.05–0.13) | 0.30 (0.29–0.31) | <0.001 |
| C22:5 (g/day) * | 0.05 (0.03–0.07) | 0.09 (0.08–0.10) | <0.001 |
| C22:6 (g/day) * | 0.24 (0.17–0.31) | 0.58 (0.55–0.61) | <0.001 |
| Total Omega 6 (% of energy/day) * | 4.3 (3.6–6.2) | 5.2 (4.3–5.9) | 0.062 |
| C18:2 (g/day) * | 9 (7–14) | 9 (7–12) | 0.633 |
| C20:4 (g/day) * | 0.11 (0.08–0.13) | 0.13 (0.11–0.15) | 0.024 |
| Food groups (g/day) | |||
| Oily fish * | 250 (117–250) | 500 (500–500) | <0.001 |
| Lean fish * | 250 (250–500) | 500 (500–500) | 0.001 |
| Shellfish * | 93 (0–93) | 50 (0–138) | 0.934 |
| Olive Oil * | 175 (70–175) | 245 (105–245) | <0.001 |
| Nuts * | 73 (35–275) | 75 (0–131) | 0.081 |
| Fruit * | 2800 (1493–3200) | 2800 (800–4200) | 0.478 |
| Vegetables * | 1600 (1200–2000) | 1400 (925–1950) | 0.161 |
| Dairy products * | 50 (0–425) | 700 (313–1384) | <0.001 |
| Eggs * | 138 (138–207) | 173 (138–276) | 0.265 |
| Red meat * | 150 (70–220) | 113 (0–300) | 0.241 |
| RBC Membrane FA (% rel) | Unadjusted Control | Unadjusted Cancer | p | Adjusted Control | Adjusted Cancer | p | Difference |
|---|---|---|---|---|---|---|---|
| (n = 37) | (n = 54) | (n = 37) | (n = 54) | (%) | |||
| Saturated fatty acids | |||||||
| Palmitic acid (16:0) | 26.0 ± 0.2 | 24.6 ± 0.2 | <0.001 | 26.1 ± 0.3 | 24.6 ± 0.2 | 0.003 | −5.7 |
| Stearic acid (18:0) | 21.0 ± 0.2 | 17.4 ± 0.1 | <0.001 | 20.8 ± 0.3 | 17.5 ± 0.2 | <0.001 | −15.9 |
| Monounsaturated fatty acids | |||||||
| Palmitoleic acid (16:1 n-7) * | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.506 | 0.4 ± 0.1 | 0.4 ± 0.1 | 0.305 | - |
| Oleic acid (18:1 n-9) * | 15.5 ± 0.2 | 17.6 ± 0.2 | <0.001 | 15.5 ± 0.3 | 17.5 ± 0.2 | <0.001 | 12.9 |
| 18:1 n-11 * | 1.1 ± 0.0 | 1.7 ± 0.0 | <0.001 | 1.1 ± 0.1 | 1.7 ± 0.1 | <0.001 | 54.5 |
| Polyunsaturated acid | |||||||
| Linoleic acid (18:2n-6) | 11.5 ± 0.2 | 11.9 ± 0.2 | 0.140 | 10.8 ± 0.3 | 12.5 ± 0.2 | <0.001 | 15.7 |
| Dihomo-γ-linoleic acid (20:3) * | 1.8 ± 0.1 | 2.2 ± 0.1 | <0.001 | 1.8 ± 0.1 | 2.2 ± 0.1 | 0.006 | 22.2 |
| Arachidonic acid (20:4) * | 16.1 ± 0.2 | 16.9 ± 0.2 | 0.033 | 16.8 ± 0.4 | 16.4 ± 0.3 | 0.472 | - |
| EPA (20:5) * | 0.8 ± 0.0 | 1.0 ± 0.1 | 0.026 | 0.8 ± 0.1 | 0.9 ± 0.1 | 0.323 | - |
| DHA (22:6) | 5.8 ± 0.1 | 6.0 ± 0.2 | 0.517 | 5.9 ± 0.2 | 5.9 ± 0.2 | 0.894 | - |
| Trans fatty acids | |||||||
| Trans 18:1 * | 0.2 ± 0.0 | 0.1 ± 0.0 | <0.001 | 0.2 ± 0.0 | 0.1 ± 0.0 | <0.001 | −50 |
| Trans 20:4 * | 0.2 ± 0.0 | 0.1 ± 0.0 | <0.001 | 0.2 ± 0.0 | 0.1 ± 0.0 | <0.001 | −50 |
| Total fatty acids | |||||||
| Total SFA | 47.0 ± 0.4 | 42.0 ± 0.1 | <0.001 | 46.9 ± 0.4 | 42.0 ± 0.3 | ND | −10.4 |
| Total MUFA | 17.0 ± 0.2 | 19.7 ± 0.2 | <0.001 | 17.1 ± 0.3 | 19.7 ± 0.2 | <0.001 | 15.2 |
| Total PUFA | 36.0 ± 0.3 | 38.0 ± 0.2 | <0.001 | 36.0 ± 0.4 | 38.0 ± 0.3 | <0.001 | 5.3 |
| Total Omega 6 | 29.4 ± 0.3 | 31.0 ± 0.2 | <0.001 | 29.3 ± 0.4 | 31.0 ± 0.3 | 0.002 | 5.8 |
| Total Trans * | 0.4 ± 0.2 | 0.2 ± 0.0 | <0.001 | 0.4 ± 0.0 | 0.2 ± 0.0 | ND | |
| Fatty acid indexes | |||||||
| Saturation Index | 2.8 ± 0.0 | 2.1 ± 0.0 | <0.001 | 2.8 ± 0.1 | 2.2 ± 0.0 | ND | −27.3 |
| Unsaturation Index | 150.9 ± 1.4 | 156.9 ± 1.3 | 0.001 | 150.9 ± 1.9 | 156.8 ± 1.5 | 0.049 | 3.83 |
| Peroxidation Index | 135.8 ± 1.6 | 141.8 ±1.4 | 0.006 | 136.3 ± 3.3 | 141.5 ± 1.8 | 0.143 | - |
| Inflammatory risk Index * | 4.3 ± 0.3 | 4.7 ± 0.2 | 0.591 | 4.6 ± 0.3 | 4.7 ± 0.2 | 0.801 | - |
| PUFA Balance | 18.3 ± 0.5 | 18.3 ± 0.6 | 0.428 | 18.6 ± 0.7 | 18.1 ± 0.5 | 0.927 | - |
| n-3 cardiovascular risk index | 6.6 ± 0.2 | 7.0 ± 0.2 | 0.085 | 6.7 ± 0.3 | 6.9 ± 0.2 | 0.699 | - |
| Enzymatic indexes | |||||||
| Δ9D 18:0/18:1 * | 1.4 ± 0.0 | 1.0 ± 0.0 | <0.001 | 1.4 ± 0.1 | 1.0 ± 0.0 | <0.001 | −28.6 |
| Δ6D+ELO 18:2/20:3 | 6.7 ± 0.2 | 5.7 ± 0.2 | 0.001 | 6.4 ± 0.3 | 5.9 ± 0.2 | 0.219 | - |
| Δ5D 20:4/20:3 | 9.3 ± 0.3 | 8.2 ± 0.3 | 0.009 | 10.0 ± 0.5 | 7.7 ± 0.4 | 0.002 | −23.0 |
| Δ9D 16:0/16:1 * | 64.9 ± 2.7 | 59.0 ± 2.6 | 0.081 | 67.6 ± 4.0 | 57.1 ± 3.0 | 0.082 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amézaga, J.; Arranz, S.; Urruticoechea, A.; Ugartemendia, G.; Larraioz, A.; Louka, M.; Uriarte, M.; Ferreri, C.; Tueros, I. Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients. Nutrients 2018, 10, 1853. https://doi.org/10.3390/nu10121853
Amézaga J, Arranz S, Urruticoechea A, Ugartemendia G, Larraioz A, Louka M, Uriarte M, Ferreri C, Tueros I. Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients. Nutrients. 2018; 10(12):1853. https://doi.org/10.3390/nu10121853
Chicago/Turabian StyleAmézaga, Javier, Sara Arranz, Ander Urruticoechea, Gurutze Ugartemendia, Aitziber Larraioz, Maria Louka, Matxalen Uriarte, Carla Ferreri, and Itziar Tueros. 2018. "Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients" Nutrients 10, no. 12: 1853. https://doi.org/10.3390/nu10121853