Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen


Abstract
1. Introduction
2. Gluten as the Trigger for Autoimmunity
3. Immune Activating Potential of Gluten Peptides
4. Analysis of Gluten Peptides
5. Factors Affecting Gluten Content Estimations
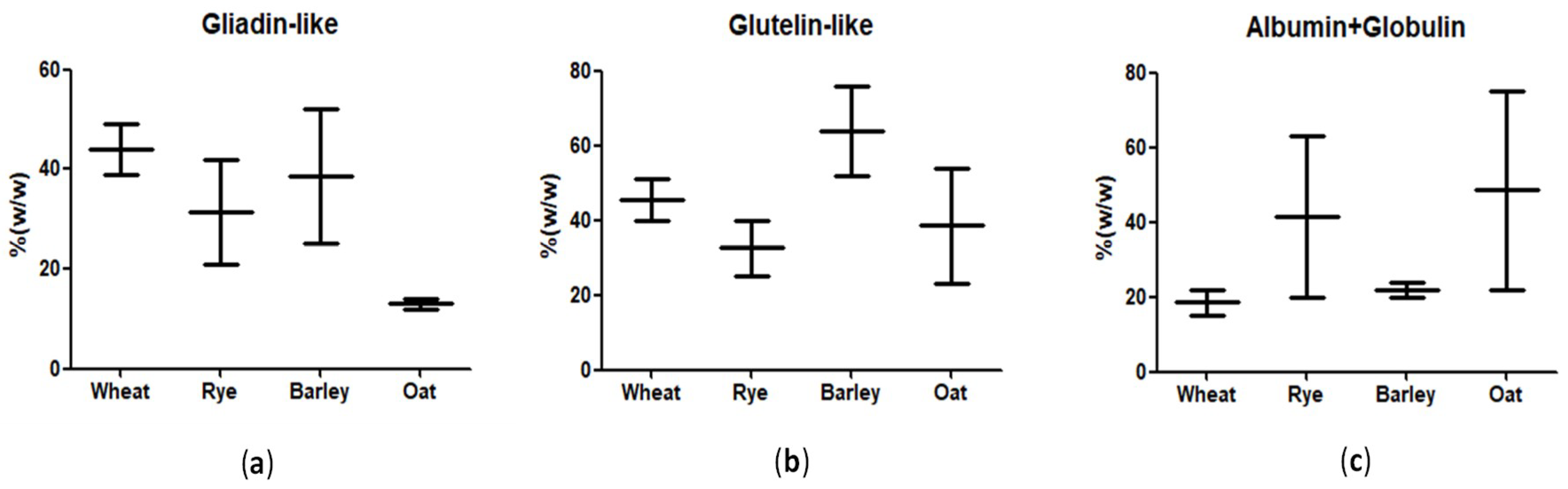
5.1. Gliadin as Standard
5.2. Is Gluten Equivalent in Wheat, Barley, and Rye?
5.3. Can Oats Contain Gluten?
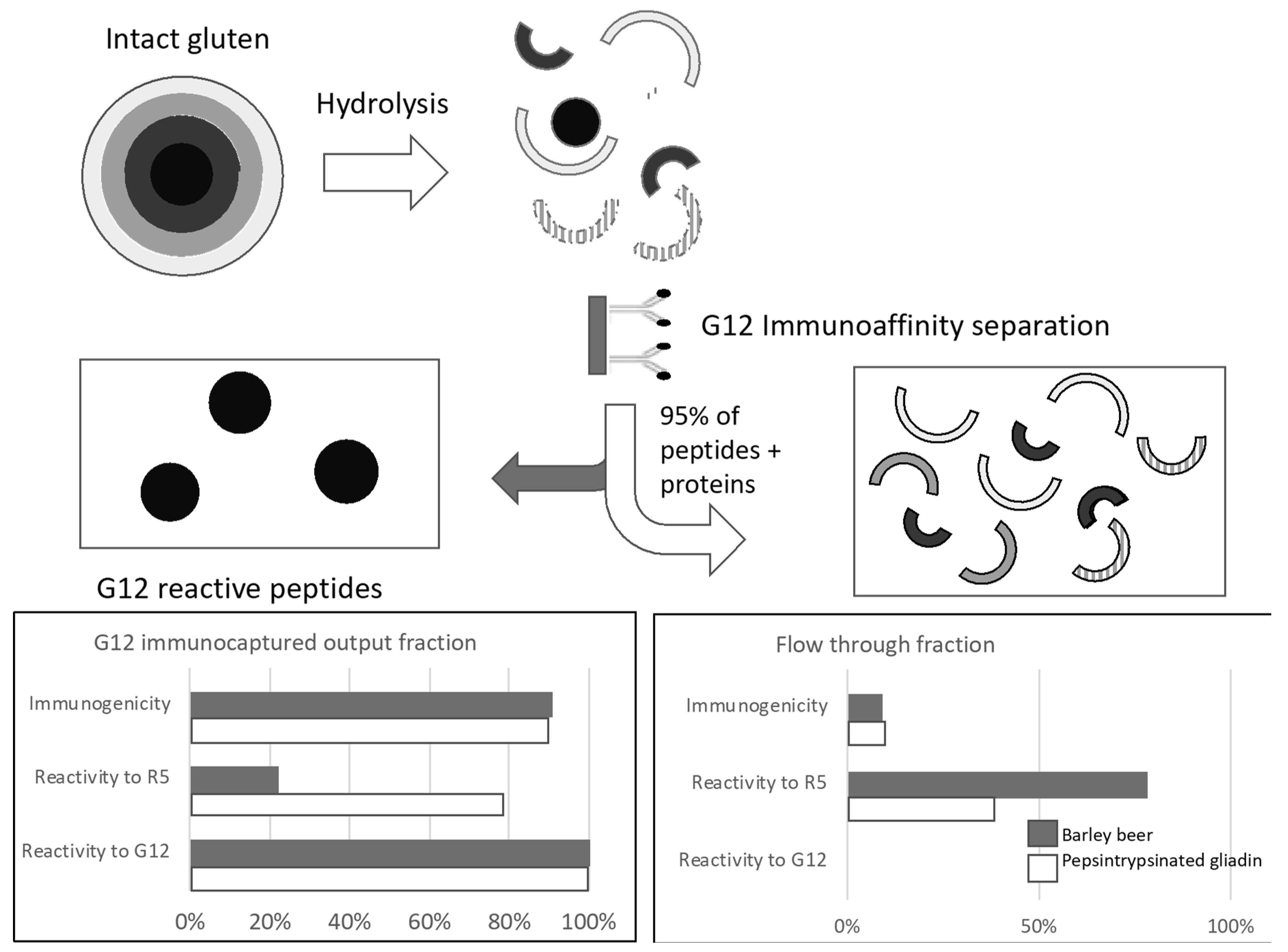
5.4. How Much Immunogenic Gluten is There in Hydrolyzed Gluten?
5.5. How Much Gluten Does Genetically-Modified Low Immunogenic Cereal Contain?
5.6. Gluten Immunogenic Peptides Modified by Bacterial Enzymes
5.7. Gluten Immunogenic Peptide Can be Excreted in Stools and Urine
6. Conclusions and Future Directions
7. Patents
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mena, M.C.; Sousa, C. Analytical tools for gluten detection: Policies and regulation. OmniaSci. Monogra. 2015, 527–564. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Martínez-Esteso, M.J.; Nørgaard, J.; Brohée, M.; Haraszi, R.; Maquet, A.; O’Connor, G. Defining the wheat gluten peptide fingerprint via a discovery and targeted proteomics approach. J. Proteomics 2016, 147, 156–168. [Google Scholar] [CrossRef] [PubMed]
- Mamone, G.; Picariello, G.; Addeo, F.; Ferranti, P. Proteomic analysis in allergy and intolerance to wheat products. Expert Rev. Proteomics 2011, 8, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Javed, A.A. Dissolvable Sample Collection Matrices and Methods of Using the Same. U.S. Patent 15/265607, 15 February 2018. [Google Scholar]
- Shewry, P.R.; Halford, N.G.; Tatham, A.S.; Popineau, Y.; Lafiandra, D.; Belton, P.S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res. 2003, 45, 219–302. [Google Scholar] [PubMed]
- Osborne, T.B. The Proteins of the Wheat Kernel; Carnegie Institution of Washington: Washington, DC, USA, 1907. [Google Scholar]
- Eliasson, A.C.; Larsson, K. Cereals in Breadmaking: A Molecular Colloidal Approach; Marcel Dekker, Inc.: New York, NY, USA, 1993. [Google Scholar]
- Alais, C.; Linden, G.; Morton, I.; Whitehead, A. Food Biochemistry; Springer-Science+Business Media, B.V: Dordrecht, The Netherlands, 1991. [Google Scholar]
- Giuberti, G.; Gallo, A.; Masoero, F. A comparison of methods to quantify prolamin contents in cereals. Ital. J. Anim. Sci. 2011, 10, 7–13. [Google Scholar] [CrossRef]
- Žilić, S.; Barać, M.; Pešić, M.; Dodig, D.; Ignjatović-Micić, D. Genetic variability of albumin-globulin content, and lipoxygenase, peroxidase activities among bread and durum wheat genotypes. Genetika 2011, 43, 503–516. [Google Scholar] [CrossRef]
- Real, A.; Comino, I.; de Lorenzo, L.; Merchán, F.; Gil-Humanes, J.; Giménez, M.J.; López-Casado, M.Á.; Isabel Torres, M.; Cebolla, Á.; Sousa, C.; et al. Molecular and immunological characterization of gluten proteins isolated from oat cultivars that differ in toxicity for celiac disease. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Kanerva, P.; Brinck, O.; Sontag-Strohm, T.; Salovaara, H.; Loponen, J. Deamidation of gluten proteins and peptides decreases the antibody affinity in gluten analysis assays. J. Cereal Sci. 2011, 53, 335–339. [Google Scholar] [CrossRef]
- Chirdo, F.G.; Arranz, E. Cereal proteins: Immunostimulatory and toxic peptides. Adv. Underst. Gluten Relat. Pathol. Evol. Gluten-Free Foods 2015, 141–162. [Google Scholar] [CrossRef]
- Balakireva, A.V.; Zamyatnin, A.A. Properties of gluten intolerance: Gluten structure, evolution, pathogenicity and detoxification capabilities. Nutrients 2016, 8, 644. [Google Scholar] [CrossRef] [PubMed]
- Van de Wal, Y.; Kooy, Y.M.C.; van Veelen, P.A.; Pena, S.A.; Mearin, L.M.; Molberg, O.; Lundin, K.E.A.; Sollid, L.M.; Mutis, T.; Benckhuijsen, W.E.; et al. Small intestinal T cells of celiac disease patients recognize a natural pepsin fragment of gliadin. Proc. Natl. Acad. Sci. 1998, 95, 10050–10054. [Google Scholar] [CrossRef] [PubMed]
- McAllister, C.S.; Kagnoff, M.F. The immunopathogenesis of celiac disease reveals possible therapies beyond the gluten-free diet. Semin. Immunopathol. 2012, 34, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Jabri, B.; Sollid, L.M. T cells in celiac disease. J. Immunol. 2017, 198, 3005–3014. [Google Scholar] [CrossRef] [PubMed]
- Mearin, M.L. The prevention of coeliac disease. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Neuhausen, S.L.; Steele, L.; Ryan, S.; Mousavi, M.; Pinto, M.; Osann, K.E.; Flodman, P.; Zone, J.J. Co-occurrence of Celiac Disease and Other Autoimmune Diseases in Celiacs and Their First-Degree Relatives. J. Autoimmun. 2008, 31, 160–165. [Google Scholar] [CrossRef]
- Lauret, E.; Rodrigo, L. Celiac disease and autoimmune-associated conditions. BioMed Res. Int. 2013, 2013, 127589. [Google Scholar] [CrossRef]
- Bruins, M.J. The clinical response to gluten challenge: A review of the literature. Nutrients 2013, 5, 4614–4641. [Google Scholar] [CrossRef]
- Venhoff, N.; Emmerich, F.; Neagu, M.; Salzer, U.; Koehn, C.; Driever, S.; Kreisel, W.; Rizzi, M.; Effelsberg, N.M.; Kollert, F.; et al. The role of HLA DQ2 and DQ8 in dissecting celiac-like disease in common variable immunodeficiency. J. Clin. Immunol. 2013, 33, 909–916. [Google Scholar] [CrossRef]
- Koning, F. Celiac disease: Quantity matters. Semin. Immunopathol. 2012, 34, 541–549. [Google Scholar] [CrossRef]
- Vader, W.; Kooy, Y.; Van Veelen, P.; De Ru, A.; Harris, D.; Benckhuijsen, W.; Pea, S.; Mearin, L.; Drijfhout, J.W.; Koning, F. The gluten response in children with celiac disease is directed toward multiple gliadin and glutenin peptides. Gastroenterology 2002, 122, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Molberg, Ø.; Mcadam, S.N.; Körner, R.; Quarsten, H.; Kristiansen, C.; Madsen, L.; Fugger, L.; Scott, H.; Norën, O.; Roepstorff, P.; et al. Tissue transglutaminase selectively modifies gliadin peptides that are recognized by gut-derived T cells in celiac disease. Nat. Med. 1998, 4, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Bodd, M.; Ráki, M.; Tollefsen, S.; Fallang, L.E.; Bergseng, E.; Lundin, K.E.A.; Sollid, L.M. HLA-DQ2-restricted gluten-reactive T cells produce IL-21 but not IL-17 or IL-22. Mucosal Immunol. 2010, 3, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Tye-Din, J.A.; Stewart, J.A.; Dromey, J.A.; Beissbarth, T.; Van Heel, D.A.; Tatham, A.; Henderson, K.; Mannering, S.I.; Gianfrani, C.; Jewell, D.P.; et al. Comprehensive, quantitative mapping of T cell epitopes in gluten in celiac disease. Sci. Transl. Med. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Willemijn Vader, L.; Stepniak, D.T.; Bunnik, E.M.; Kooy, Y.M.C.; De Haan, W.; Drijfhout, J.W.; Van Veelen, P.A.; Koning, F. Characterization of cereal toxicity for celiac disease patients based on protein homology in grains. Gastroenterology 2003, 125, 1105–1113. [Google Scholar] [CrossRef]
- van Herpen, T.W.J.M.; Goryunova, S.V.; van der Schoot, J.; Mitreva, M.; Salentijn, E.; Vorst, O.; Schenk, M.F.; van Veelen, P.A.; Koning, F.; van Soest, L.J.M.; et al. Alpha-gliadin genes from the A, B, and D genomes of wheat contain different sets of celiac disease epitopes. BMC Genomics 2006, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ciccocioppo, R.; Di Sabatino, A.; Corazza, G.R. The immune recognition of gluten in coeliac disease. Clin. Exp. Immunol. 2005, 140, 408–416. [Google Scholar] [CrossRef]
- Bruins Slot, I.D.; Van Der Fels-Klerx, H.J.; Bremer, M.G.E.G.; Hamer, R.J. Immunochemical detection methods for gluten in food products: Where do we go from here? Crit. Rev. Food Sci. Nutr. 2015, 56, 2455–2466. [Google Scholar] [CrossRef]
- Silano, M.; De Vincenzi, M. In Vitro Screening of food peptides toxic for coeliac and other gluten-sensitive patients: A review. Toxicology 1999, 132, 99–110. [Google Scholar] [CrossRef]
- Camarca, A.; Anderson, R.P.; Mamone, G.; Fierro, O.; Facchiano, A.; Costantini, S.; Zanzi, D.; Sidney, J.; Auricchio, S.; Sette, A.; et al. Intestinal T cell responses to gluten peptides are largely heterogeneous: Implications for a peptide-based therapy in celiac disease. J. Immunol. 2009, 182, 4158–4166. [Google Scholar] [CrossRef]
- Shan, L.; Qiao, S.W.; Arentz-Hansen, H.; Molberg, Ø.; Gray, G.M.; Sollid, L.M.; Khosla, C. Identification and analysis of multivalent proteolytically resistant peptides from gluten: implications for celiac sprue. J. Proteome Res. 2005, 4, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Qiao, S.-W.; Bergseng, E.; Molberg, O.; Jung, G.; Fleckenstein, B.; Sollid, L.M. Refining the rules of gliadin T cell epitope binding to the disease-associated DQ2 molecule in celiac disease: Importance of proline spacing and glutamine deamidation. J. Immunol. 2005, 175, 254–261. [Google Scholar] [CrossRef] [PubMed]
- DØrum, S.; Arntzen, M.O.; Qiao, S.W.; Holm, A.; Koehler, C.J.; Thiede, B.; Sollid, L.M.; Fleckenstein, B. The preferred substrates for transglutaminase 2 in a complex wheat gluten digest are peptide fragments harboring celiac disease T-cell epitopes. PLoS ONE 2010, 5. [Google Scholar] [CrossRef]
- Sollid, L.M.; Qiao, S.W.; Anderson, R.P.; Gianfrani, C.; Koning, F. Nomenclature and listing of celiac disease relevant gluten T-cell epitopes restricted by HLA-DQ molecules. Immunogenetics 2012, 64, 455–460. [Google Scholar] [CrossRef]
- Arentz-Hansen, H.; Korner, R.; Molberg, O.; Quarsten, H.; Vader, W.; Kooy, Y.M.C.; Lundin, K.E.A.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The intestinal T cell response to a-gliadin in adult celiac disesae is focused on a single deamidated glutamine targeted by tissue transglutaminase. J. Exp. Med. 2000, 191, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.P.; Degano, P.; Godkin, A.J.; Jewell, D.P.; Hill, A.V.S. In vivo antigen challenge in celiac disease identifies a single transglutaminase-modified peptide as the dominant a-gliadin T-cell epitope. Nat. Med. 2000, 6, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Nanayakkara, M.; Lania, G.; Maglio, M.; Discepolo, V.; Sarno, M.; Gaito, A.; Troncone, R.; Auricchio, S.; Auricchio, R.; Barone, M.V. An undigested gliadin peptide activates innate immunity and proliferative signaling in enterocytes: The role in celiac disease. Am. J. Clin. Nutr. 2013, 98, 1123–1135. [Google Scholar] [CrossRef] [PubMed]
- Mamone, G.; Ferranti, P.; Rossi, M.; Roepstorff, P.; Fierro, O.; Malorni, A.; Addeo, F. Identification of a peptide from α-gliadin resistant to digestive enzymes: Implications for celiac disease. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 855, 236–241. [Google Scholar] [CrossRef]
- Comino, I.; Real, A.; Gil-Humanes, J.; Pistón, F.; de Lorenzo, L.; Moreno, M.; de, L.; López-Casado, M.Á.; Lorite, P.; Cebolla, Á.; et al. Significant differences in coeliac immunotoxicity of barley varieties. Mol. Nutr. Food Res. 2012, 56, 1697–1707. [Google Scholar] [CrossRef]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in celiac sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef]
- Maiuri, L.; Ciacci, C.; Ricciardelli, I.; Vacca, L.; Raia, V.; Auricchio, S.; Picard, J.; Osman, M.; Quaratino, S.; Londei, M. Association between innate response to gliadin and activation of pathogenic T cells in coeliac disease. Lancet 2003, 362, 30–37. [Google Scholar] [CrossRef]
- Sturgess, R.; Day, P.; Ellis, H.J.; Lundin, K.E.A.; Gjertsen, H.A.; Kontakou, M.; Ciclitira, C.P. Wheat peptide challenge in coeliac disease. Lancet 1994, 343, 758–761. [Google Scholar] [CrossRef]
- Marsh, M.N.; Morgan, S.; Ensari, A.; Wardle, T.; Lobley, R.; Mills, C.; Auricchio, S. In-vivo activity of peptides 31–43, 44–55, 56–68 of a-gliadin in Gluten Sensitive Enteropathy (GSE). Gastroenterology 1995, 108, A871. [Google Scholar] [CrossRef]
- Schalk, K.; Lexhaller, B.; Koehler, P.; Scherf, K.A. Isolation and characterization of gluten protein types from wheat, rye, barley and oats for use as reference materials. PLoS ONE 2017, 12, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gianfrani, C.; Auricchio, S.; Troncone, R. Adaptive and innate immune responses in celiac disease. Immunol. Lett. 2005, 99, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Lebreton, C.; Schumann, M.; Matysiak-Budnik, T.; Dugave, C.; Bouhnik, Y.; Malamut, G.; Cellier, C.; Allez, M.; Crenn, P.; et al. Paracellular versus transcellular intestinal permeability to gliadin peptides in active celiac disease. Am. J. Pathol. 2012, 180, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Matysiak-Budnik, T.; Candalh, C.; Dugave, C.; Namane, A.; Cellier, C.; Cerf-Bensussan, N.; Heyman, M. Alterations of the intestinal transport and processing of gliadin peptides in celiac disease. Gastroenterology 2003, 125, 696–707. [Google Scholar] [CrossRef]
- Zimmermann, C.; Rudloff, S.; Lochnit, G.; Arampatzi, S.; Maison, W.; Zimmer, K.P. Epithelial transport of immunogenic and toxic gliadin peptides in vitro. PLoS ONE 2014, 9, 1–17. [Google Scholar] [CrossRef]
- Hollon, J.; Puppa, E.; Greenwald, B.; Goldberg, E.; Guerrerio, A.; Fasano, A. Effect of gliadin on permeability of intestinal biopsy explants from celiac disease patients and patients with non-celiac gluten sensitivity. Nutrients 2015, 7, 1565–1576. [Google Scholar] [CrossRef]
- Bruun, S.W.; Josefsen, K.; Tanassi, J.T.; Marek, A.; Pedersen, M.H.F.; Sidenius, U.; Haupt-Jorgensen, M.; Antvorskov, J.C.; Larsen, J.; Heegaard, N.H.; et al. Large gliadin peptides detected in the pancreas of NOD and healthy mice following oral administration. J. Diabetes Res. 2016, 2016, 1–11. [Google Scholar] [CrossRef]
- Moreno, M.D.L.; Cebolla, Á.; Munõz-Suano, A.; Carrillo-Carrion, C.; Comino, I.; Pizarro, Á.; León, F.; Rodríguez-Herrera, A.; Sousa, C. Detection of gluten immunogenic peptides in the urine of patients with coeliac disease reveals transgressions in the gluten-Free diet and incomplete mucosal healing. Gut 2017, 66, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Syage, J.A.; Kelly, C.P.; Dickason, M.A.; Ramirez, A.C.; Leon, F.; Dominguez, R.; Sealey-Voyksner, J.A. Determination of gluten consumption in celiac disease patients on a gluten-free diet. Am. J. Clin. Nutr. 2018, 107, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Tanner, G.J.; Blundell, M.J.; Colgrave, M.L.; Howitt, C.A. Quantification of hordeins by ELISA: The correct standard makes a magnitude of difference. PLoS ONE 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Brottveit, M.; Ráki, M.; Bergseng, E.; Fallang, L.-E.; Simonsen, B.; Løvik, A.; Larsen, S.; Løberg, E.M.; Jahnsen, F.L.; Sollid, L.M.; et al. Assessing possible celiac disease by an HLA-DQ2-Gliadin tetramer test. Am. J. Gastroenterol. 2011, 106, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Sarna, V.K.; Skodje, G.I.; Reims, H.M.; Risnes, L.F.; Dahal-Koirala, S.; Sollid, L.M.; Lundin, K.E.A. HLA-DQ:Gluten Tetramer Test in Blood Gives Better Detection of Coeliac Patients than Biopsy after 14-Day Gluten Challenge. Gut 2018, 67, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Comino, I.; Real, A.; De Lorenzo, L.; Cornell, H.; López-Casado, M.Á.; Barro, F.; Lorite, P.; Torres, M.I.; Cebolla, Á.; Sousa, C. Diversity in oat potential immunogenicity: Basis for the selection of oat varieties with no toxicity in coeliac disease. Gut 2011, 60, 915–922. [Google Scholar] [CrossRef] [PubMed]
- Comino, I.; Real, A.; Vivas, S.; Síglez, M.Á.; Caminero, A.; Nistal, E.; Casqueiro, J.; Rodríguez-Herrera, A.; Cebolla, Á.; Sousa, C. Monitoring of gluten-free diet compliance in celiac patients by assessment of gliadin 33-mer equivalent epitopes in feces. Am. J. Clin. Nutr. 2012, 95, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Comino, I.; Real, A.; de Lourdes Moreno, M.; Montes, R.; Cebolla, Á.; Sousa, C. Immunological determination of gliadin 33-mer equivalent peptides in beers as a specific and practical analytical method to assess safety for celiac patients. J. Sci. Food Agric. 2013, 93, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.D.L.; Muñoz-Suano, A.; López-Casado, M.Á.; Torres, M.I.; Sousa, C.; Cebolla, Á. Selective capture of most celiac immunogenic peptides from hydrolyzed gluten proteins. Food Chem. 2016, 205, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Bruins Slot, I.D.; Bremer, M.G.E.G.; Van Der Fels-Klerx, I.; Hamer, R.J. Evaluating the performance of gluten ELISA test kits: The numbers do not tell the tale. Cereal Chem. 2015, 92, 513–521. [Google Scholar] [CrossRef]
- Rzychon, M.; Brohée, M.; Cordeiro, F.; Haraszi, R.; Ulberth, F.; O’Connor, G. The feasibility of harmonizing gluten ELISA measurements. Food Chem. 2017, 234, 144–154. [Google Scholar] [CrossRef] [PubMed]
- Skerritt, J.H.; Hill, A.S. Monoclonal antibody sandwich enzyme immunoassays for determination of gluten in foods. J. Agric. Food Chem. 1990, 38, 1771–1778. [Google Scholar] [CrossRef]
- Skerritt, J.H.; Hill, A.S. Enzyme immunoassay for determination of gluten in foods: Collaborative study. J. Assoc. Off. Anal. Chem. 1991, 74, 257–264. [Google Scholar] [PubMed]
- Ellis, H.J.; Rosen-Bronson, S.; O’Reilly, N.; Ciclitira, P.J. Measurement of gluten using a monoclonal antibody to a coeliac toxic peptide of a gliadin. Gut 1998, 43, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Mitea, C.; Kooy-Winkelaar, Y.; Van Veelen, P.; De Ru, A.; Drijfhout, J.W.; Koning, F.; Dekking, L. Fine specificity of monoclonal antibodies against celiac disease-inducing peptides in the gluteome. Am. J. Clin. Nutr. 2008, 88, 1057–1066. [Google Scholar] [CrossRef] [PubMed]
- Valdés, I.; García, E.; Llorente, M.; Méndez, E. Innovative approach to low-level gluten determination in foods using a novel sandwich enzyme-linked immunosorbent assay protocol. Eur. J. Gastroenterol. Hepatol. 2003, 15, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Méndez, E.; Vela, C.; Immer, U.; Janssen, F.W. Report of a collaborative trial to investigate the performance of the r5 enzyme linked immunoassay to determine gliadin in gluten-free food. Eur. J. Gastroenterol. Hepatol. 2005, 17, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Torgler, C.; Vilchez, F.; Sousa, C. Analytical tools to detect gluten immuno-toxic fractions in food based on monoclonal antibodies raised against the gliadin 33-mer peptide. Proc. 24th Meet. Work. Gr. prolamin Anal. Toxic. 2011. Available online: http://foodsafety.biomedal.com/wp-content/uploads/2013/10/Working-group-on-prolamin-Analytical-Tools-p-47-51.pdf (accessed on 26 November 2018).
- Zhang, J.; Portela, S.B.; Horrell, J.B.; Leung, A.; Weitmann, D.R.; Artiuch, J.B.; Wilson, S.M.; Cipriani, M.; Slakey, L.K.; Burt, A.M.; et al. An integrated, accurate, rapid, and economical handheld consumer gluten detector. Food Chem. 2019, 275, 446–456. [Google Scholar] [CrossRef]
- Morón, B.; Bethune, M.T.; Comino, I.; Manyani, H.; Ferragud, M.; López, M.C.; Cebolla, Á.; Khosla, C.; Sousa, C. Toward the assessment of food toxicity for celiac patients: Characterization of monoclonal antibodies to a main immunogenic gluten peptide. PLoS ONE 2008, 3, 1–13. [Google Scholar] [CrossRef]
- Skerritt, J.H.; Hill, A.S.; Andrews, J.L. Antigenicity of wheat prolamins: Detailed epitope analysis using a panel of monoclonal antibodies. J. Cereal Sci. 2000, 32, 259–279. [Google Scholar] [CrossRef]
- Van den Broeck, H.C.; de Jong, H.C.; Salentijn, E.M.J.; Dekking, L.; Bosch, D.; Hamer, R.J.; Gilissen, L.J.W.J.; van der Meer, I.M.; Smulders, M.J.M. Presence of Celiac Disease Epitopes in Modern and Old Hexaploid Wheat Varieties: Wheat Breeding May Have Contributed to Increased Prevalence of Celiac Disease. Theor. Appl. Genet. 2010, 121, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Morón, B.; Cebolla, Á,; Manyani, H.; Álvarez-Maqueda, M.; Megías, M.; Thomas, M.; del, C.; López, M.C.; Sousa, C. Sensitive detection of cereal fractions that are toxic to celiac disease patients by using monoclonal antibodies to a main immunogenic wheat peptide. Am. J. Clin. Nutr. 2008, 87, 405–414. [Google Scholar] [CrossRef]
- Real, A.; Comino, I.; Moreno, M.D.L.; López-Casado, M.Á.; Lorite, P.; Isabel Torres, M.; Cebolla, Á.; Sousa, C. Identification and in vitro reactivity of celiac immunoactive peptides in an apparent gluten-free beer. PLoS ONE 2014, 9, 12–14. [Google Scholar] [CrossRef] [PubMed]
- Comino, I.; Fernández-Bañares, F.; Esteve, M.; Ortigosa, L.; Castillejo, G.; Fambuena, B.; Ribes-Koninckx, C.; Sierra, C.; Rodríguez-Herrera, A.; Salazar, J.C.; et al. Fecal gluten peptides reveal limitations of serological tests and food questionnaires for monitoring gluten-free diet in celiac disease patients. Am. J. Gastroenterol. 2016, 111, 1456–1465. [Google Scholar] [CrossRef] [PubMed]
- Soler, M.; Estevez, M.C.; Moreno, M.; de, L.; Cebolla, A.; Lechuga, L.M. Label-free SPR detection of gluten peptides in urine for non-invasive celiac disease follow-up. Biosens. Bioelectron. 2016, 79, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Ehren, J.; Móron, B.; Martin, E.; Bethune, M.T.; Gray, G.M.; Khosla, C. A food-grade enzyme preparation with modest gluten detoxification properties. PLoS ONE 2009, 4, 1–10. [Google Scholar] [CrossRef]
- Taylor, S.L.; Nordlee, J.A.; Jayasena, S.; Baumert, J.L. Evaluation of a handheld gluten detection device. J. Food Prot. 2018, 81, 1723–1728. [Google Scholar] [CrossRef]
- Diaz-Amigo, C.; Popping, B. Accuracy of ELISA detection methods for gluten and reference materials: A realistic assessment. J. Agric. Food Chem. 2013, 61, 5681–5688. [Google Scholar] [CrossRef]
- Gessendorfer, B.; Koehler, P.; Wieser, H. Preparation and characterization of enzymatically hydrolyzed prolamins from wheat, rye, and barley as references for the immunochemical quantitation of partially hydrolyzed gluten. Anal. Bioanal. Chem. 2009, 395, 1721–1728. [Google Scholar] [CrossRef]
- Spaenij-Dekking, L.; Kooy-Winkelaar, Y.; Van Veelen, P.; Drijfhout, J.W.; Jonker, H.; Van Soest, L.; Smulders, M.J.M.; Bosch, D.; Gilissen, L.J.W.J.; Koning, F. Natural variation in toxicity of wheat: potential for selection of nontoxic varieties for celiac disease patients. Gastroenterology 2005, 129, 797–806. [Google Scholar] [CrossRef]
- Gregorini, A.; Colomba, M.; Julia Ellis, H.; Ciclitira, P.J. Immunogenicity characterization of two ancient wheat α-gliadin peptides related to coeliac disease. Nutrients 2009, 1, 276–290. [Google Scholar] [CrossRef] [PubMed]
- Hernando, A.; Mujico, J.R.; Mena, M.C.; Lombardía, M.; Méndez, E. Measurement of wheat gluten and barley hordeins in contaminated oats from europe, the united states and canada by sandwich R5 ELISA. Eur. J. Gastroenterol. Hepatol. 2008, 20, 545–554. [Google Scholar] [CrossRef]
- Giménez, M.J.; Real, A.; García-Molina, M.D.; Sousa, C.; Barro, F. Characterization of celiac disease related oat proteins: bases for the development of high quality oat varieties suitable for celiac patients. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benoit, L.; Masiri, J.; Del Blanco, I.A.; Meshgi, M.; Gendel, S.M.; Samadpour, M. Assessment of avenins from different oat varieties using R5-based sandwich ELISA. J. Agric. Food Chem. 2017, 65, 1467–1472. [Google Scholar] [CrossRef]
- Comino, I.; Bernardo, D.; Bancel, E.; Moreno, M.; de, L.; Sánchez, B.; Barro, F.; Šuligoj, T.; Ciclitira, P.J.; Cebolla, Á.; Knight, S.C.; et al. Identification and molecular characterization of oat peptides implicated on coeliac immune response. Food Nutr. Res. 2016, 60, 30324. [Google Scholar] [CrossRef] [PubMed]
- Arentz-Hansen, H.; Fleckenstein, B.; Molberg, Ø.; Scott, H.; Koning, F.; Jung, G.; Roepstorff, P.; Lundin, K.E.A.; Sollid, L.M. The molecular basis for oat intolerance in patients with celiac disease. PLoS Med. 2004, 1, 084–095. [Google Scholar] [CrossRef] [PubMed]
- Silano, M.; Penas Pozo, E.; Uberti, F.; Manferdelli, S.; Del Pinto, T.; Felli, C.; Budelli, A.; Vincentini, O.; Restani, P. Diversity of oat varieties in eliciting the early inflammatory events in celiac disease. Eur. J. Nutr. 2014, 53, 1177–1186. [Google Scholar] [CrossRef] [PubMed]
- Sey, M.S.L.; Parfitt, J.; Gregor, J. prospective study of clinical and histological safety of pure and uncontaminated canadian oats in the management of celiac disease. J. Parenter. Enter. Nutr. 2011, 35, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Picariello, G.; Mamone, G.; Cutignano, A.; Fontana, A.; Zurlo, L.; Addeo, F.; Ferranti, P. Proteomics, peptidomics, and immunogenic potential of wheat beer (weissbier). J. Agric. Food Chem. 2015, 63, 3579–3586. [Google Scholar] [CrossRef] [PubMed]
- Dostálek, P.; Hochel, I.; Méndez, E.; Hernando, A.; Gabrovská, D. Immunochemical determination of gluten in malts and beers. Food Addit. Contam. 2006, 23, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Tian, N.; Siezen, R.; Schuppan, D.; Helmerhorst, E.J. Identification of food-grade subtilisins as gluten-degrading enzymes to treat celiac disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G571–G580. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Pistón, F.; Giménez, M.J.; Martín, A.; Barro, F. The introgression of rnai silencing of γ-gliadins into commercial lines of bread wheat changes the mixing and technological properties of the dough. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Pistón, F.; Barro, F.; Rosell, C.M. The shutdown of celiac disease-related gliadin epitopes in bread wheat by rnai provides flours with increased stability and better tolerance to over-mixing. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Piston, F.; Tollefsen, S.; Sollid, L.M.; Barro, F. Effective Shutdown in the Expression of Celiac Disease-Related Wheat Gliadin T-Cell Epitopes by RNA Interference. Proc. Natl. Acad. Sci. 2010, 107, 17023–17028. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-Gluten, Nontransgenic Wheat Engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef]
- García-Molina, M.D.; García-Olmo, J.; Barro, F. Effective Identification of Low-Gliadin Wheat Lines by Near Infrared Spectroscopy (NIRS): Implications for the Development and Analysis of Foodstuffs Suitable for Celiac Patients. PLoS ONE 2016, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schalk, K.; Koehler, P.; Scherf, K.A. Quantitation of specific barley, rye, and oat marker peptides by targeted liquid chromatography-mass spectrometry to determine gluten concentrations. J. Agric. Food Chem. 2018, 66, 3581–3592. [Google Scholar] [CrossRef] [PubMed]
- Santos, K.F.D.N.; Silveira, R.D.D.; Martin-Didonet, C.C.G.; Brondani, C. Storage protein profile and amino acid content in wild rice oryza glumaepatula. Pesqui. Agropecu. Bras. 2013, 48, 66–72. [Google Scholar] [CrossRef]
- Tanner, G.J.; Blundell, M.J.; Colgrave, M.L.; Howitt, C.A. Creation of the first ultra-low gluten barley (hordeum vulgare l.) for coeliac and gluten-intolerant populations. Plant Biotechnol. J. 2016, 14, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Malalgoda, M.; BomOhm, J.; Meinhardt, S.; Chao, S.; Simsek, S. Cluster analysis of historical and modern hard red spring wheat cultivars based on parentage and HPLC analysis of gluten-forming proteins. Cereal Chem. 2017, 94, 560–567. [Google Scholar] [CrossRef]
- Francavilla, R.; De Angelis, M.; Rizzello, C.G.; Cavallo, N.; Dal Bello, F.; Gobbetti, M. Selected probiotic lactobacilli have the capacity to hydrolyze gluten peptides during simulated gastrointestinal digestion. Appl. Environ. Microbiol. 2017, 83, 1–12. [Google Scholar] [CrossRef]
- Caminero, A.; Galipeau, H.J.; McCarville, J.L.; Johnston, C.W.; Bernier, S.P.; Russell, A.K.; Jury, J.; Herran, A.R.; Casqueiro, J.; Tye-Din, J.A.; et al. Duodenal bacteria from patients with celiac disease and healthy subjects distinctly affect gluten breakdown and immunogenicity. Gastroenterology 2016, 151, 670–683. [Google Scholar] [CrossRef] [PubMed]
- Janssen, G.; Christis, C.; Kooy-Winkelaar, Y.; Edens, L.; Smith, D.; Van Veelen, P.; Koning, F. Ineffective degradation of immunogenic gluten epitopes by currently available digestive enzyme supplements. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gerasimidis, K.; Zafeiropoulou, K.; Mackinder, M.; Ijaz, U.Z.; Duncan, H.; Buchanan, E.; Cardigan, T.; Edwards, C.A.; McGrogan, P.; Russell, R.K. Comparison of clinical methods with the faecal gluten immunogenic peptide to assess gluten intake in coeliac disease. J. Pediatr. Gastroenterol. Nutr. 2018, 67, 356–360. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Peptide Sequence | Origin, Description and Practical Evidences | References |
|---|---|---|
| LQLQPFP(QPQLPYP)3QPQPF | α-gliadin 33-mer. Correlation between food immunoreactivity with T-cells and reactivity of anti-33-mer antibodies among different oat and barley cultivars, and beer fractions. Suppression of 90% immunogenicity with G12 anti-33-mer antibody gluten peptides | [12,43,60,61,62,63] |
| EPEQPIPEQPQPYPQQ | α-gliadin | [57] |
| EQPFPQPEQPFPWQP | ω-gliadin/C-hordein | [57] |
| ELQPFPQPELPYPQPQ | β-hordein | [28] |
| QLQPFPQPELPY | DQ2.5-glia-α1a. Use in HLA-DQ Gluten tetramer tests | [59] |
| PQPELPYPQPE | β-hordein. DQ2.5-glia-α2. Use in HLA-DQ Gluten tetramer tests | [28,59] |
| QQPFPQPEQPFP | DQ2.5-glia-ω1. Use in HLA-DQ Gluten tetramer tests | [59] |
| EQPFPQPEQPFPWQP | ω-gliadin/C-hordein. DQ2.5-glia-α2. Use in HLA-DQ Gluten tetramer tests | [28,59] |
| EPEQPIPEQPQPYPQ | α-gliadin. DQ2.5-hor-3. Use in HLA-DQ Gluten tetramer tests | [28,59] |
| Monoclonal Antibodies | Prefered Epitope | References |
|---|---|---|
| G12 | QPQ-(L/Q)-P-(Y/F) | [74] |
| A1 | Q-(Q/L)-P-(Y/F)-PQP | [74] |
| R5 | QQPFP | [70] |
| 401.21 | PQ-(PQ/QP)-PFP-(QE/EES) | [75] |
| α20 | F-RPQQPYP-Q | [76] |
| 14G11 | α-gliadin 33-mer, ND | [73] |
| 13F6 | α-gliadin 33-mer, ND | [73] |
| Cereal | Prolamin (%) | Glutelin (%) | Immunoreactivity α | Detection Method | References |
|---|---|---|---|---|---|
| Wheat | 5.94 a | 2.98 a | ++ | G12 and A1 moAbs | [77,79,80] |
| Transgenic wheat | 1.96 b | 4.16 b | ND. 85% G12 reactivity reduction | R5 and G12 moAbs | [100,101] |
| Barley | 3.13 a | 1.1 a | ++ β | G12 moAb | [43,74,78] |
| Rye | 2.53 a | 0.55 a | ND | G12 and A1 moAbs | [74,77] |
| Oats | 1.29 a | 1.01 a | +/− β | G12 moAb | [60,74,77,78 |
| Rice | 0.54 c | 6.66 c | − | G12 moAb | [60,74,77,78] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cebolla, Á.; Moreno, M.d.L.; Coto, L.; Sousa, C. Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen. Nutrients 2018, 10, 1927. https://doi.org/10.3390/nu10121927
Cebolla Á, Moreno MdL, Coto L, Sousa C. Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen. Nutrients. 2018; 10(12):1927. https://doi.org/10.3390/nu10121927
Chicago/Turabian StyleCebolla, Ángel, María de Lourdes Moreno, Laura Coto, and Carolina Sousa. 2018. "Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen" Nutrients 10, no. 12: 1927. https://doi.org/10.3390/nu10121927
APA StyleCebolla, Á., Moreno, M. d. L., Coto, L., & Sousa, C. (2018). Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen. Nutrients, 10(12), 1927. https://doi.org/10.3390/nu10121927