Eucommia ulmoides Ameliorates Glucotoxicity by Suppressing Advanced Glycation End-Products in Diabetic Mice Kidney

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Ultraperformance Liquid Chromatography (UPLC)-Tandem Mass Spectrometry (MS/MS) Analyses
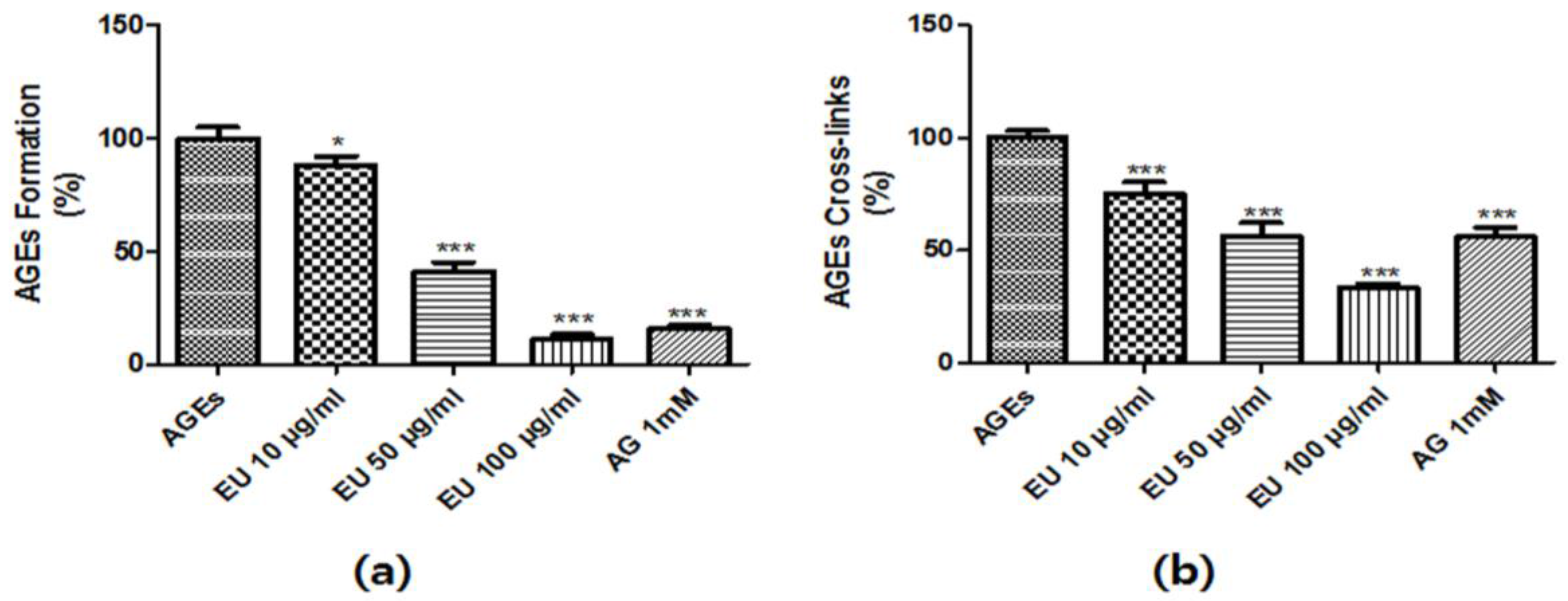
2.4. Inhibitory Effects of EU on AGE Formation
2.5. AGE Cross-Link Inhibitory Effect of EU
2.6. Animals
2.7. Blood Measurements
2.8. Measurement of Total AGEs
2.9. Western Blotting Analysis
2.10. Glo1 Activity Assay
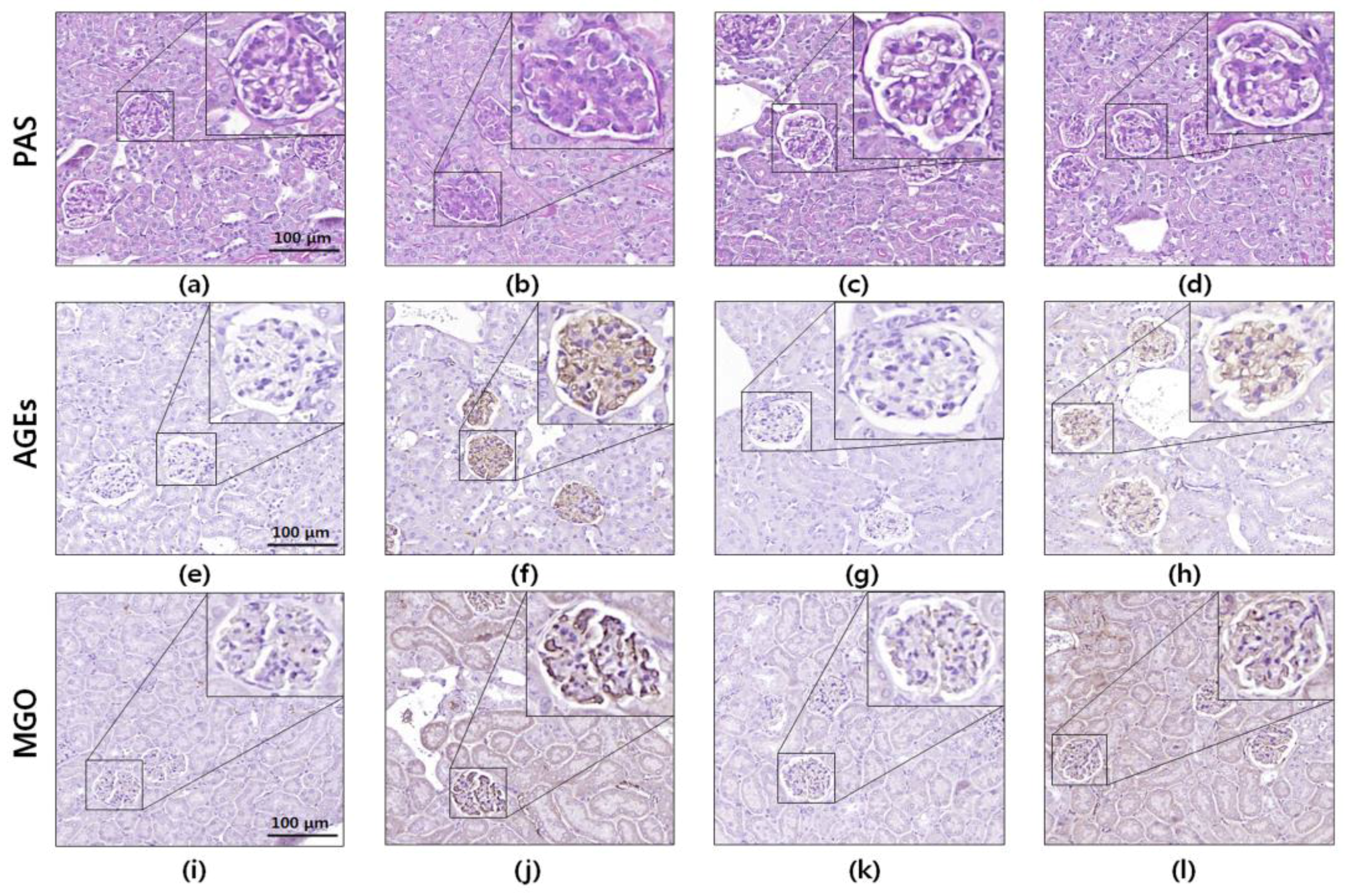
2.11. Histology and Immunohistochemistry
2.12. Statistical Analysis
3. Results
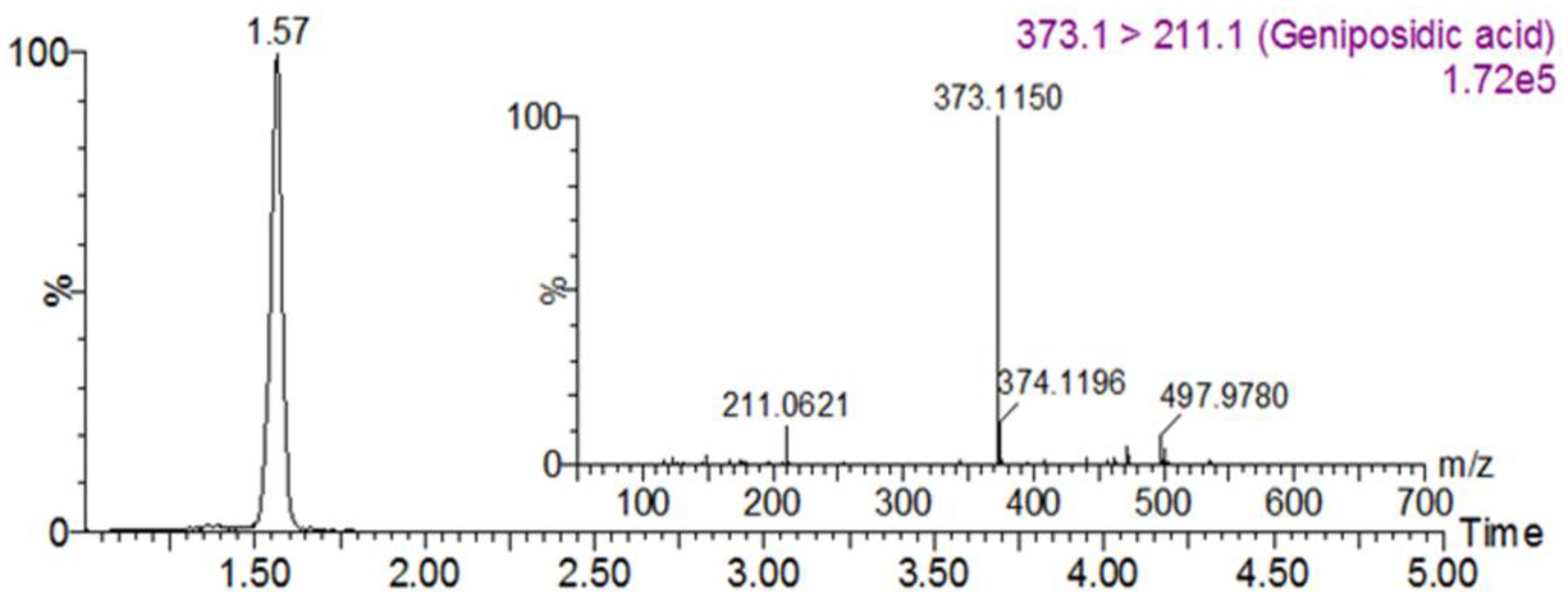
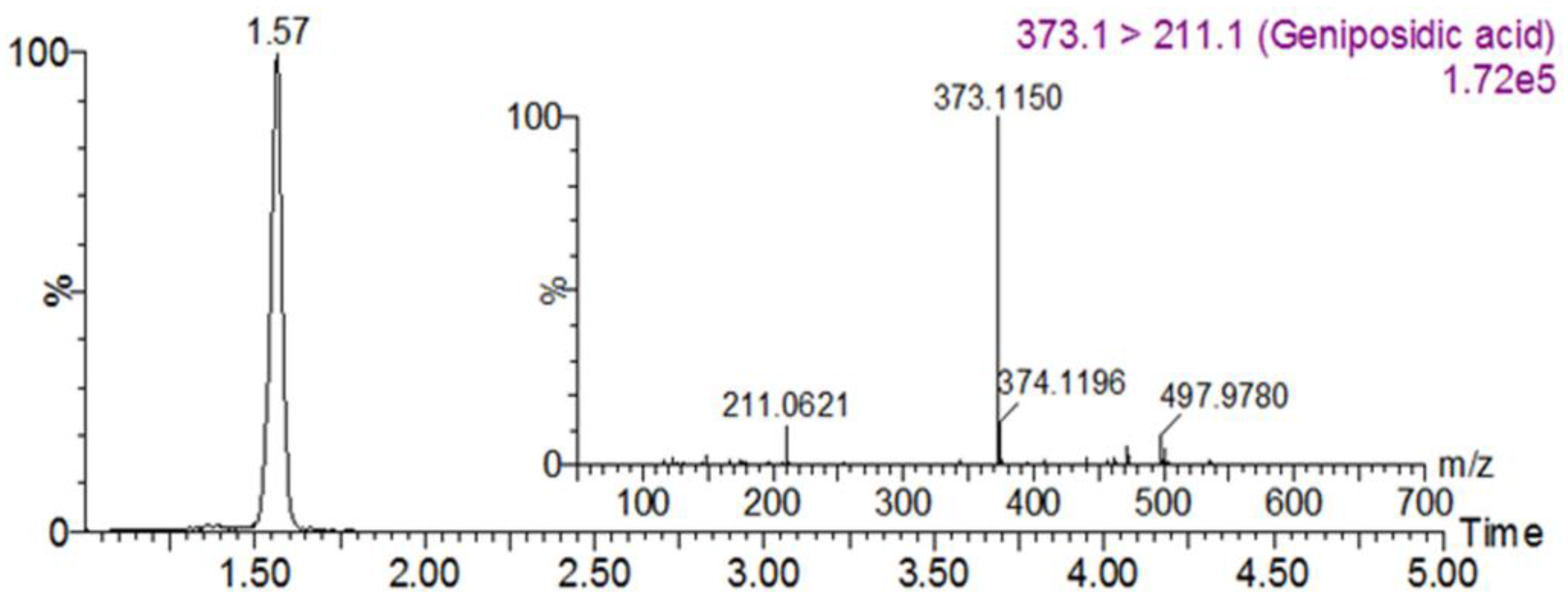
3.1. Geniposidic Acid Concentration in EU
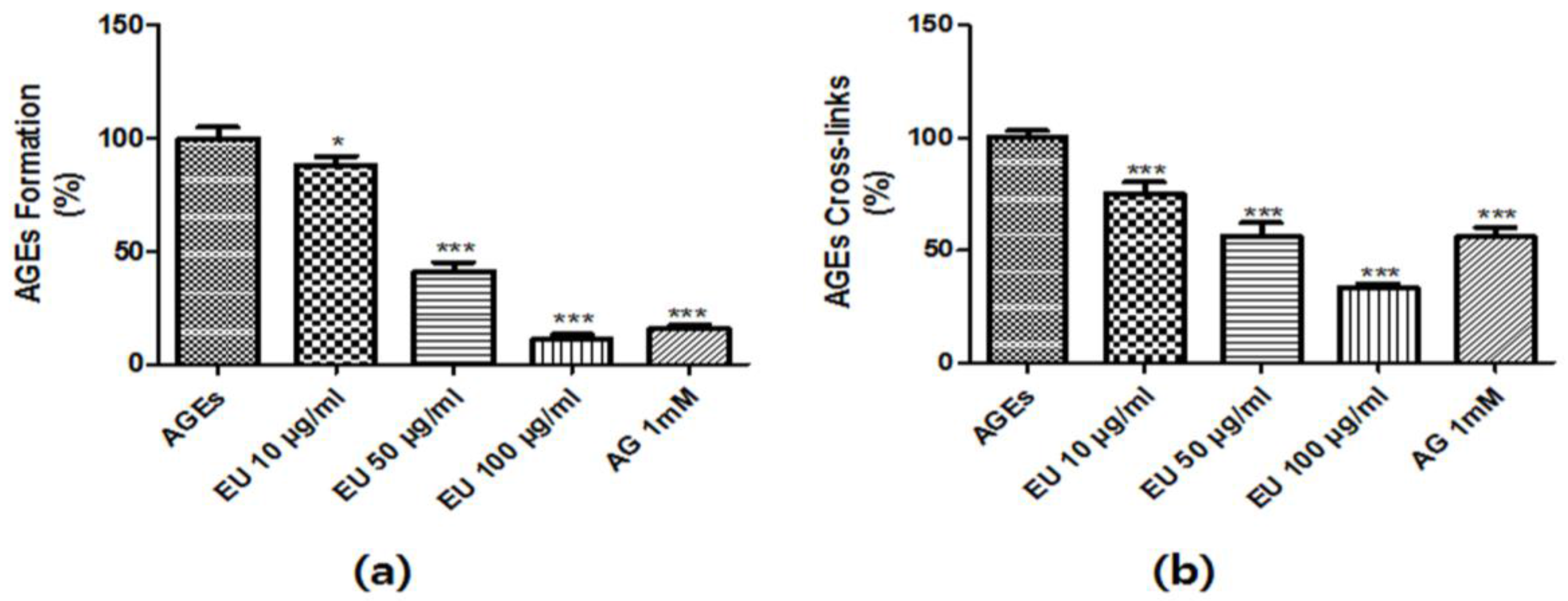
3.2. Inhibitory Effect of EU on AGE Formation and Cross-Linking
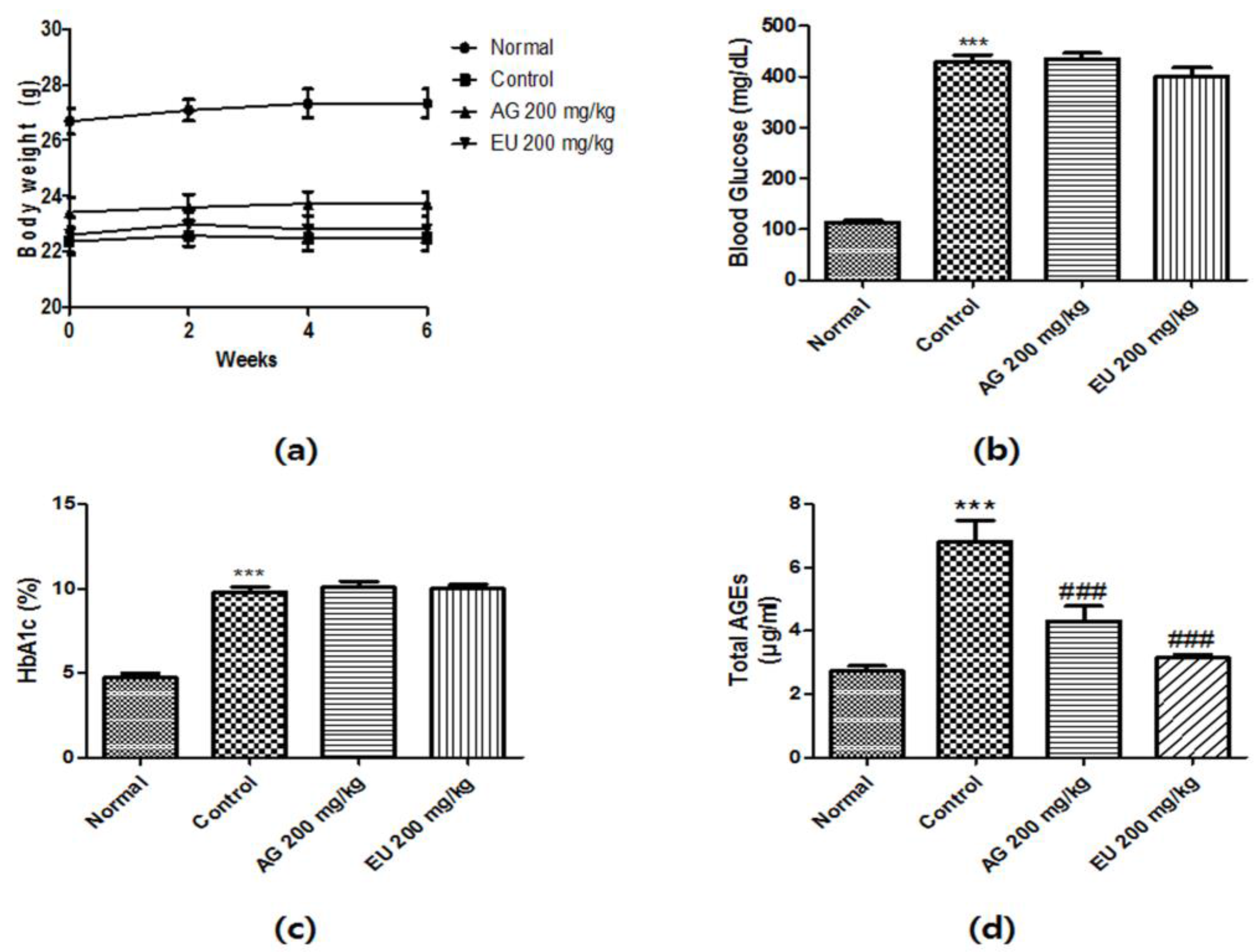
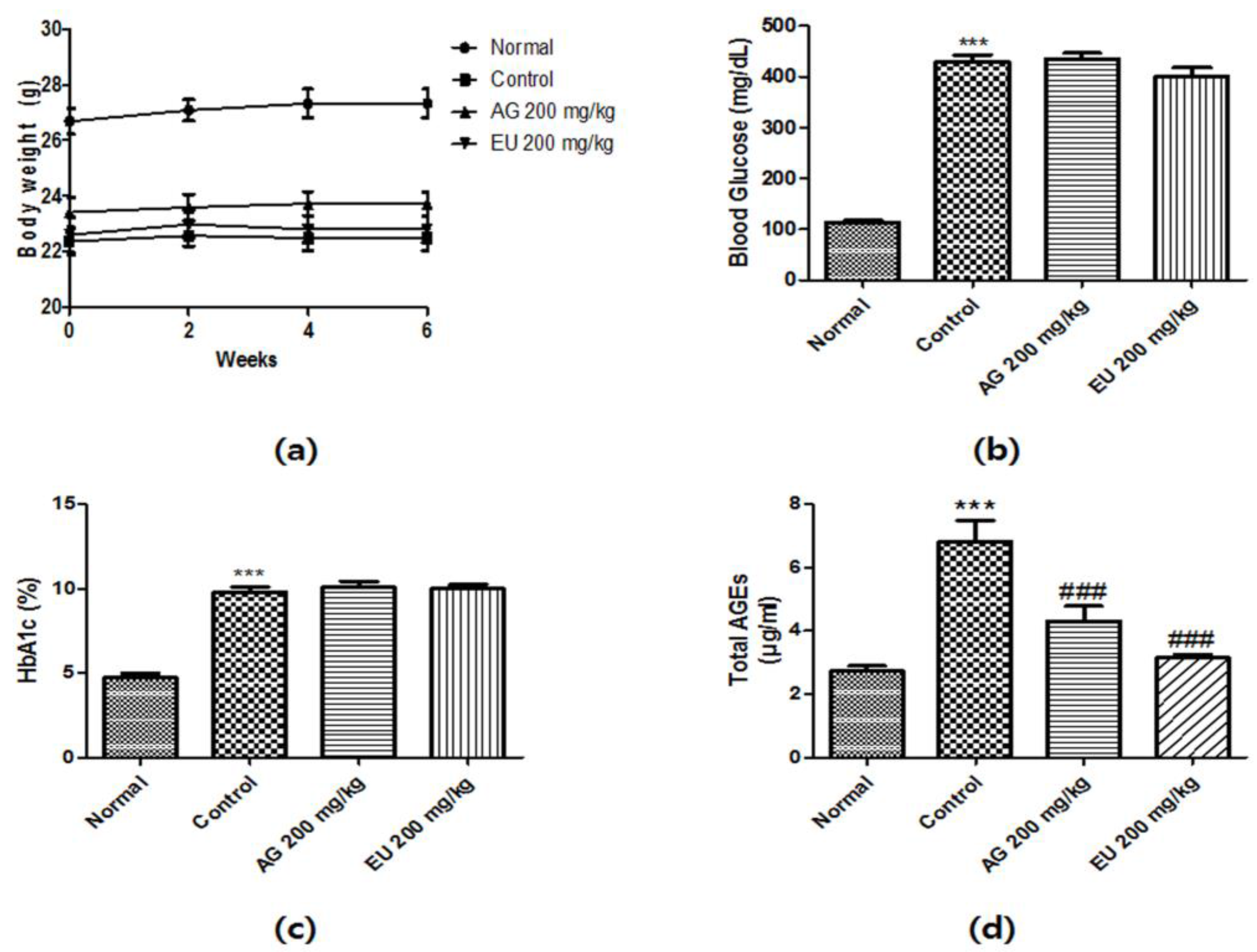
3.3. Effects of EU on Diabetic Parameters
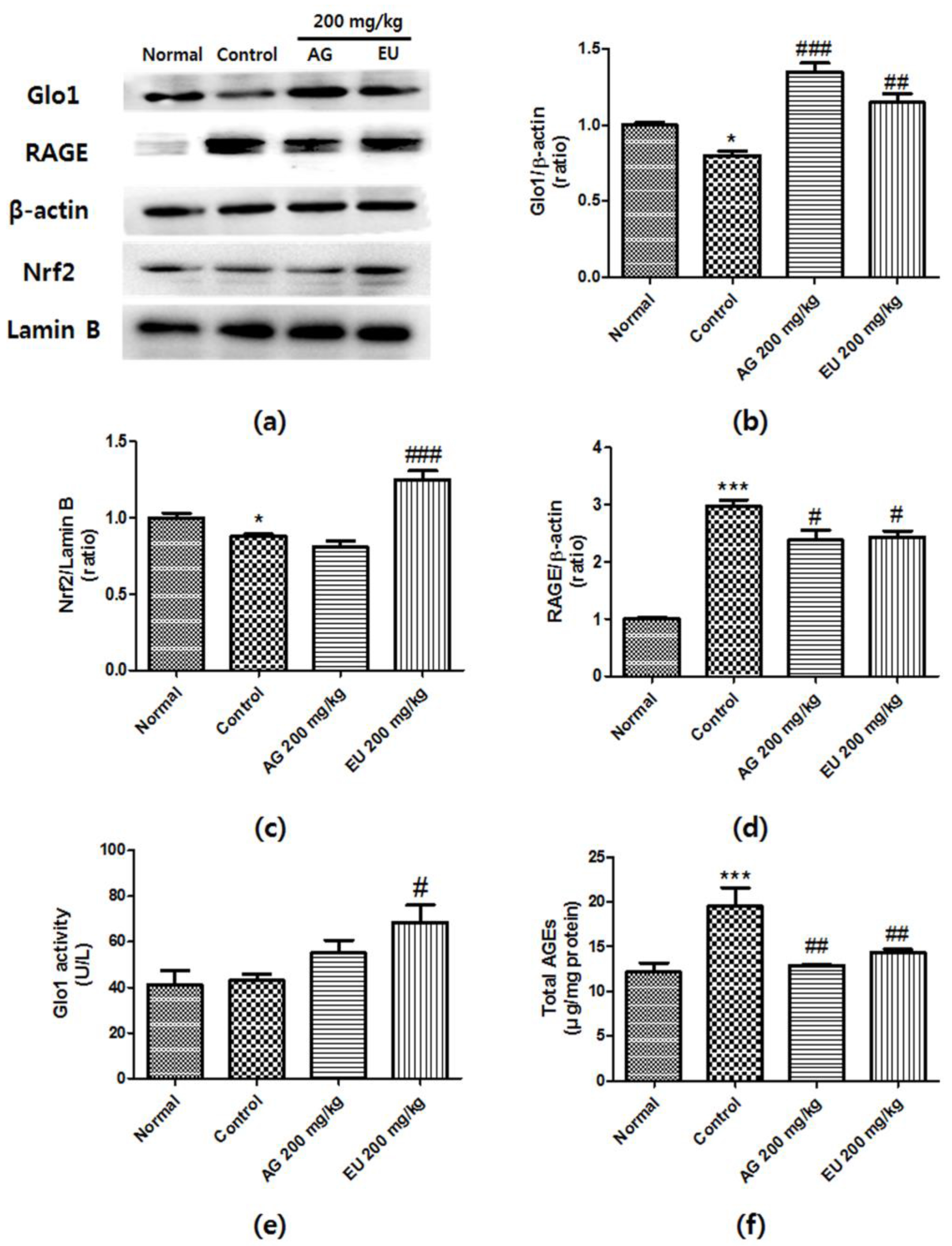
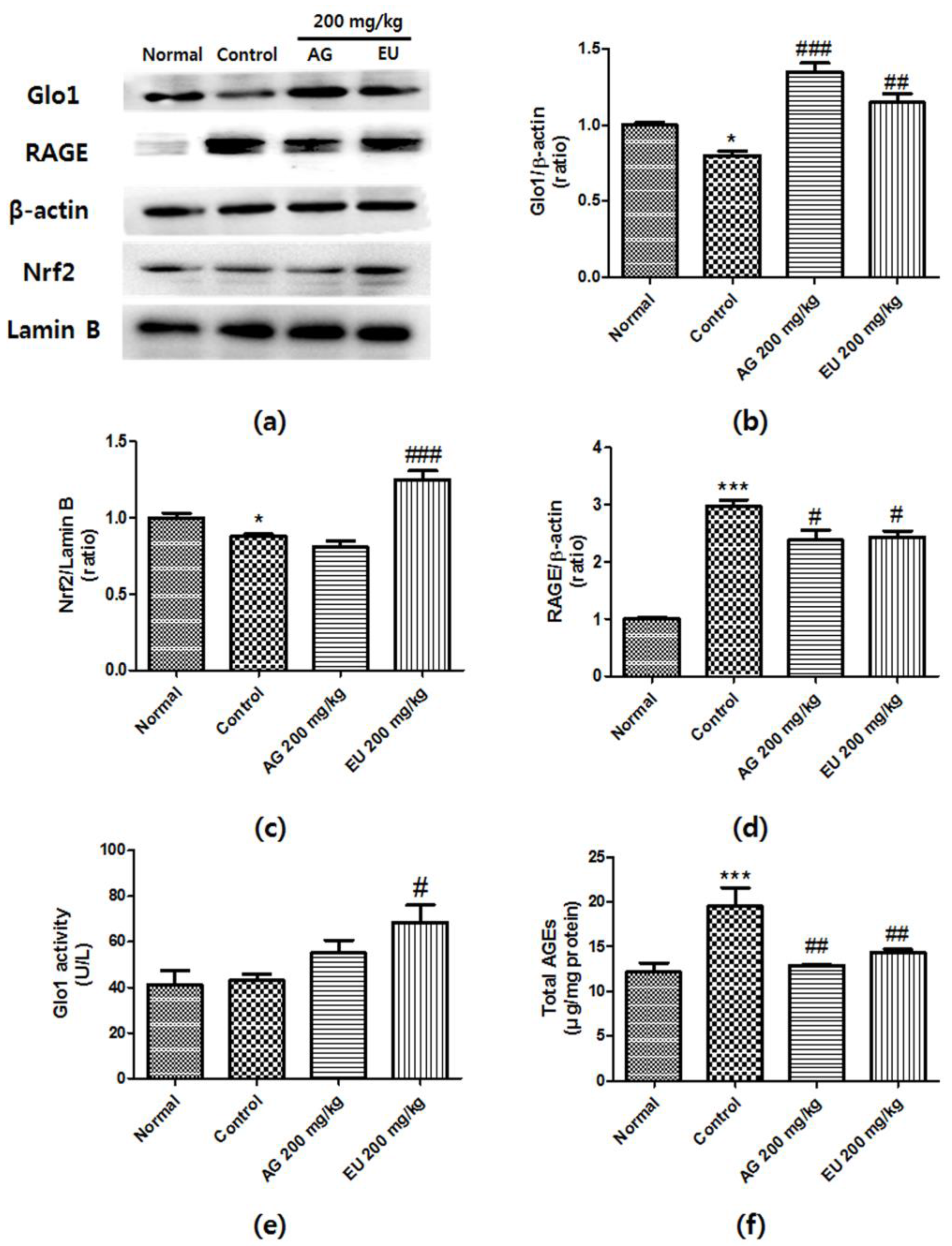
3.4. Effects of EU on Glo1 Expression and Activity
3.5. Effects of EU on Nrf2 and RAGE Expression
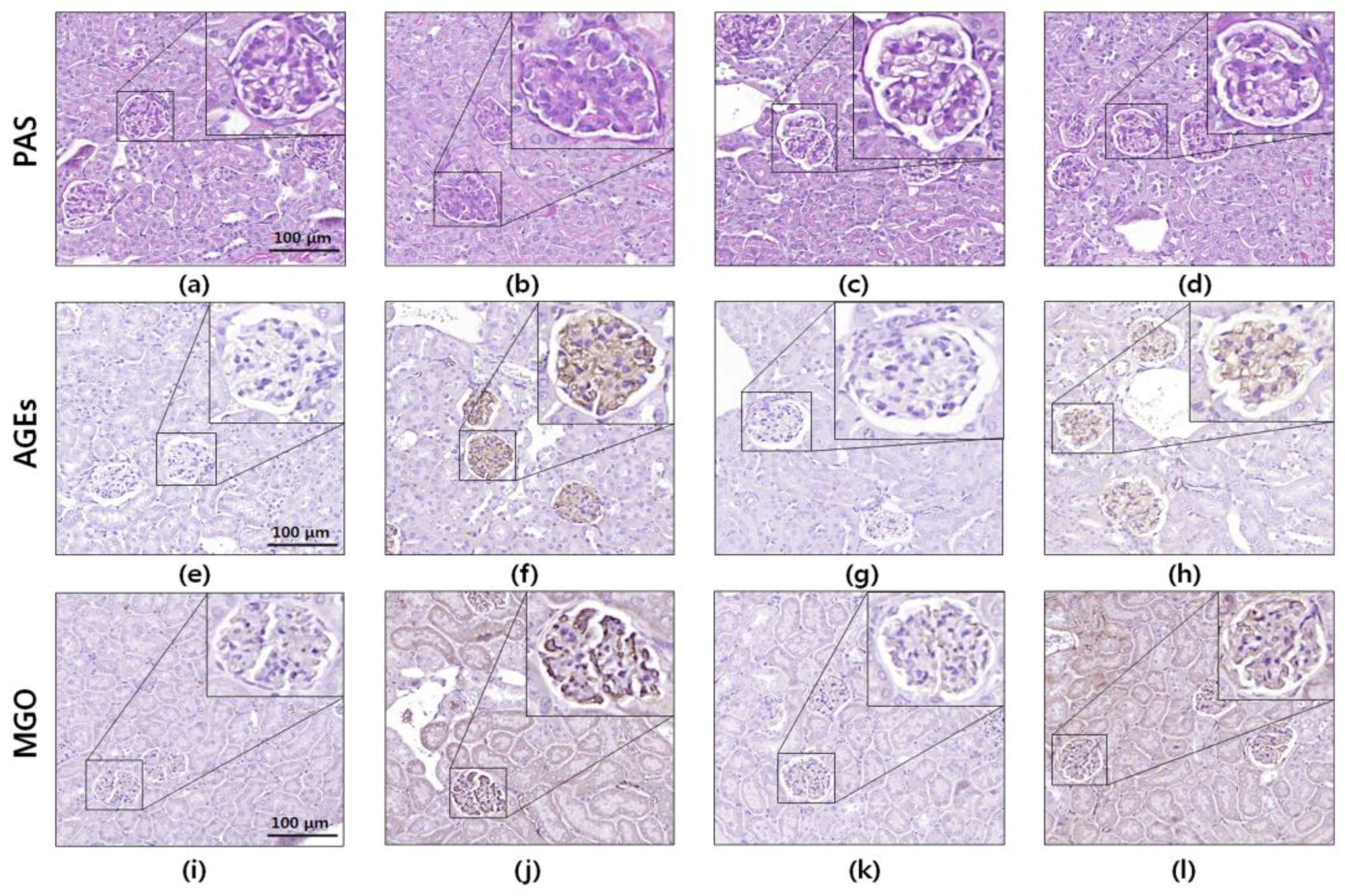
3.6. Histology and Immunohistochemistry
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced glycation end products and diabetic complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Stolzenberg-Solomon, R.; Zimmerman, T.P.; Duan, Z.; Chen, L.; Kahle, L.; Risch, A.; Subar, A.F.; Cross, A.J.; Hollenbeck, A. Dietary consumption of advanced glycation end products and pancreatic cancer in the prospective NIH-AARP diet and health study. Am. J. Clin. Nutr. 2015, 101, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.-I. Role of advanced glycation end products (ages) and receptor for ages (rage) in vascular damage in diabetes. Exp. Gerontol. 2011, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Jung, D.H.; Sohn, E.; Lee, Y.M.; Kim, C.-S.; Kim, J.S. Extract of cassiae semen attenuates diabetic nephropathy via inhibition of advanced glycation end products accumulation in streptozotocin-induced diabetic rats. Phytomedicine 2014, 21, 734–739. [Google Scholar] [CrossRef] [PubMed]
- Kamalov, M.; Harris, P.; Cooper, G.; Brimble, M. Site-specific cross-linking of collagen peptides by lysyl advanced glycation endproducts. Chem. Commun. 2014, 50, 4944–4946. [Google Scholar] [CrossRef] [PubMed]
- Maessen, D.E.; Stehouwer, C.D.; Schalkwijk, C.G. The role of methylglyoxal and the glyoxalase system in diabetes and other age-related diseases. Clin. Sci. 2015, 128, 839–861. [Google Scholar] [CrossRef] [PubMed]
- Matafome, P.; Rodrigues, T.; Sena, C.; Seiça, R. Methylglyoxal in metabolic disorders: Facts, myths, and promises. Med. Res. Rev. 2017, 37, 368–403. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Wang, L.; Zhou, Q.; Yan, S.; Li, Z.; Sheng, J.; Zhang, W. (+)-Catechin ameliorates diabetic nephropathy by trapping methylglyoxal in type 2 diabetic mice. Mol. Nutr. Food Res. 2014, 58, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Thornalley, P.; Giardino, I.; Beisswenger, P.; Thorpe, S.R.; Onorato, J.; Brownlee, M. Overexpression of glyoxalase-I in bovine endothelial cells inhibits intracellular advanced glycation endproduct formation and prevents hyperglycemia-induced increases in macromolecular endocytosis. J. Clin. Investig. 1998, 101, 1142. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, O.; Niessen, P.M.; Miyata, T.; Østergaard, J.A.; Flyvbjerg, A.; Peutz-Kootstra, C.J.; Sieber, J.; Mundel, P.H.; Brownlee, M.; Janssen, B.J. Glyoxalase-1 overexpression reduces endothelial dysfunction and attenuates early renal impairment in a rat model of diabetes. Diabetologia 2014, 57, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Geoffrion, M.; Du, X.; Irshad, Z.; Vanderhyden, B.C.; Courville, K.; Sui, G.; D’Agati, V.D.; Ott-Braschi, S.; Rabbani, N.; Thornalley, P.J.; et al. Differential effects of glyoxalase 1 overexpression on diabetic atherosclerosis and renal dysfunction in streptozotocin-treated, apolipoprotein e-deficient mice. Physiol. Rep. 2014, 2, e12043. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-H.; Lee, B.-H.; Chang, Y.-Y.; Hsu, Y.-W.; Pan, T.-M. A novel natural Nrf2 activator with pparγ-agonist (monascin) attenuates the toxicity of methylglyoxal and hyperglycemia. Toxicol. Appl. Pharmacol. 2013, 272, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Mastrocola, R. Ages and neurodegeneration: The Nrf2/glyoxalase-1 interaction. Oncotarget 2017, 8, 5645. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-H.; Feng, L.; Zhu, M.-M.; Gu, J.-F.; Jiang, J.; Cheng, X.-D.; Ding, S.-M.; Wu, C.; Jia, X.-B. The anti-inflammation effect of moutan cortex on advanced glycation end products-induced rat mesangial cells dysfunction and high-glucose–fat diet and streptozotocin-induced diabetic nephropathy rats. J. Ethnopharmacol. 2014, 151, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Engelen, L.; Stehouwer, C.D.; Schalkwijk, C.G. Current therapeutic interventions in the glycation pathway: Evidence from clinical studies. Diabetes Obes. Metab. 2013, 15, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Yen, G.-C.; Hsieh, C.-L. Reactive oxygen species scavenging activity of du-zhong (Eucommia ulmoides Oliv.) and its active compounds. J. Agric. Food Chem. 2000, 48, 3431–3436. [Google Scholar] [CrossRef] [PubMed]
- Hosoo, S.; Koyama, M.; Kato, M.; Hirata, T.; Yamaguchi, Y.; Yamasaki, H.; Wada, A.; Wada, K.; Nishibe, S.; Nakamura, K. The restorative effects of Eucommia ulmoides Oliver leaf extract on vascular function in spontaneously hypertensive rats. Molecules 2015, 20, 21971–21981. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-K.; Kim, M.-J.; Cho, S.-Y.; Park, S.A.; Park, K.-K.; Jung, U.J.; Park, H.-M.; Choi, M.-S. Hypoglycemic effect of du-zhong (Eucommia ulmoides oliv.) leaves in streptozotocin-induced diabetic rats. Diabetes Res. Clin. Pract. 2005, 67, 22–28. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, J.; Li, M.; Hao, D.; Yang, Y.; Zhang, C.; He, R.; Tao, R. Eucommia ulmoides oliv.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2014, 151, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Sugawa, H.; Ohno, R.; Shirakawa, J.; Nakajima, A.; Kanagawa, A.; Hirata, T.; Ikeda, T.; Moroishi, N.; Nagai, M.; Nagai, R. Eucommia ulmoides extracts prevent the formation of advanced glycation end products. Food Funct. 2016, 7, 2566–2573. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-Y.; Oh, J.-G.; Kim, J.S.; Lee, K.-W. Effects of chebulic acid on advanced glycation endproducts-induced collagen cross-links. Biol. Pharm. Bull. 2014, 37, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Beisswenger, P.J.; Howell, S.K.; Russell, G.B.; Miller, M.E.; Rich, S.S.; Mauer, M. Early progression of diabetic nephropathy correlates with methylglyoxal-derived advanced glycation end products. Diabetes Care 2013, 36, 3234–3239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Feng, L.; Gu, J.; Ma, L.; Qin, D.; Wu, C.; Jia, X. The attenuation of moutan cortex on oxidative stress for renal injury in AGEs-induced mesangial cell dysfunction and streptozotocin-induced diabetic nephropathy rats. Oxid. Med. Cell. Longev. 2014, 2014, 463815. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Wang, T.; Managi, H.; Yoshikawa, M. Structural requirements of flavonoids for inhibition of protein glycation and radical scavenging activities. Bioorg. Med. Chem. 2003, 11, 5317–5323. [Google Scholar] [CrossRef] [PubMed]
- Dearlove, R.P.; Greenspan, P.; Hartle, D.K.; Swanson, R.B.; Hargrove, J.L. Inhibition of protein glycation by extracts of culinary herbs and spices. J. Med. Food 2008, 11, 275–281. [Google Scholar] [CrossRef] [PubMed]
- West, B.J.; Deng, S.; Uwaya, A.; Isami, F.; Abe, Y.; Yamagishi, S.-I.; Jensen, C.J. Iridoids are natural glycation inhibitors. Glycoconj. J. 2016, 33, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Xue, H.-Y.; Jin, L.-J.; Li, S.-Y.; Xu, Y.-P. Antioxidant and pancreas-protective effect of aucubin on rats with streptozotocin-induced diabetes. Eur. J. Pharmacol. 2008, 582, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, Z.-Y.; Liang, X.; Xie, C.; Chen, Y.-F. Anti-atherosclerotic effect of geniposidic acid in a rabbit model and related cellular mechanisms. Pharm. Biol. 2015, 53, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nakazawa, Y.; Onizuka, S.; Satoh, S.; Chiba, A.; Sekihashi, K.; Miura, A.; Yasugahira, N.; Sasaki, Y.F. Antimutagenicity of tochu tea (an aqueous extract of Eucommia ulmoides leaves): 1. The clastogen-suppressing effects of tochu tea in CHO cells and mice. Mutat. Res. 1997, 388, 7–20. [Google Scholar] [CrossRef]
- Zuo, T.; Xu, W.; Li, H.; Song, H.; Zhu, M. Geniposide and geniposidic acid, modified forms of genipin, attenuate genipin-induced mitochondrial apoptosis without altering the anti-inflammatory ability in KGN cell line. Med. Chem. Res. 2017, 26, 499–508. [Google Scholar] [CrossRef]
- Ramkissoon, J.S.; Mahomoodally, M.F.; Subratty, A.H.; Ahmed, N. Inhibition of glucose-and fructose-mediated protein glycation by infusions and ethanolic extracts of ten culinary herbs and spices. Asian Pac. J. Trop. Biomed. 2016, 6, 492–500. [Google Scholar] [CrossRef]
- Khan, N.; Gothalwal, R. An anti-glycating property roots ethanol and methanol extracts of Withania somnifera on in vitro glycation of elastin protein. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 899–908. [Google Scholar]
- Rabbani, N.; Thornalley, P.J. The critical role of methylglyoxal and glyoxalase 1 in diabetic nephropathy. Diabetes 2014, 63, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-W.; Cheng, Y.-Q.; Liu, X.-L.; Hao, Y.-C.; Li, Y.; Zhu, X.; Zhang, F.; Yin, X.-X. Mangiferin upregulates glyoxalase 1 through activation of nrf2/are signaling in central neurons cultured with high glucose. Mol. Neurobiol. 2017, 54, 4060–4070. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, K.; Jung, T.; Höhn, A.; Weber, D.; Grune, T. Advanced glycation end products and oxidative stress in type 2 diabetes mellitus. Biomolecules 2015, 5, 194–222. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, E.; Friess, U. Oxidative stress, age, and atherosclerosis. Kidney Int. 2007, 72, S17–S26. [Google Scholar] [CrossRef] [PubMed]
- Busch, M.; Franke, S.; Rüster, C.; Wolf, G. Advanced glycation end-products and the kidney. Eur. J. Clin. Investig. 2010, 40, 742–755. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.C.; Frasnelli, S.C.; Bastos Ade, S.; Orrico, S.R.; Rossa, C., Jr. Modulation of cell proliferation, survival and gene expression by rage and TLR signaling in cells of the innate and adaptive immune response: Role of p38 MAPK and NF-KB. J. Appl. Oral Sci. 2014, 22, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Schlöndorff, D. Choosing the right mouse model for diabetic nephropathy. Kidney Int. 2010, 77, 749–750. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Xu, X.; Wang, K.; Qi, X.; Wu, Y. Paeoniflorin attenuates incipient diabetic nephropathy in streptozotocin-induced mice by the suppression of the Toll-like receptor-2 signaling pathway. Drug Des. Dev. Ther. 2017, 11, 3221. [Google Scholar] [CrossRef] [PubMed]
- Miyata, T.; Ueda, Y.; Horie, K.; Nangaku, M.; Tanaka, S.; van Ypersele de Strihou, C.; Kurokawa, K. Renal catabolism of advanced glycation end products: The fate of pentosidine. Kidney Int. 1998, 53, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Vlassara, H.; Striker, L.J.; Teichberg, S.; Fuh, H.; Li, Y.M.; Steffes, M. Advanced glycation end products induce glomerular sclerosis and albuminuria in normal rats. Proc. Natl. Acad. Sci. USA 1994, 91, 11704–11708. [Google Scholar] [CrossRef] [PubMed]
- Stinghen, A.E.; Massy, Z.A.; Vlassara, H.; Striker, G.E.; Boullier, A. Uremic toxicity of advanced glycation end products in CKD. J. Am. Soc. Nephrol. 2016, 27, 354–370. [Google Scholar] [CrossRef] [PubMed]
- Park, S.A.; Choi, M.-S.; Jung, U.J.; Kim, M.-J.; Kim, D.J.; Park, H.M.; Park, Y.B.; Lee, M.K. Eucommia ulmoides Oliver leaf extract increases endogenous antioxidant activity in type 2 diabetic mice. J. Med. Food 2006, 9, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.-S.; Liu, I.-M.; Niu, C.-S.; Ku, P.-M.; Hsu, C.-T.; Cheng, J.-T. Eucommia bark (du-zhong) improves diabetic nephropathy without altering blood glucose in type 1-like diabetic rats. Drug Des. Dev. Ther. 2016, 10, 971. [Google Scholar] [CrossRef]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Do, M.H.; Hur, J.; Choi, J.; Kim, M.; Kim, M.J.; Kim, Y.; Ha, S.K. Eucommia ulmoides Ameliorates Glucotoxicity by Suppressing Advanced Glycation End-Products in Diabetic Mice Kidney. Nutrients 2018, 10, 265. https://doi.org/10.3390/nu10030265
Do MH, Hur J, Choi J, Kim M, Kim MJ, Kim Y, Ha SK. Eucommia ulmoides Ameliorates Glucotoxicity by Suppressing Advanced Glycation End-Products in Diabetic Mice Kidney. Nutrients. 2018; 10(3):265. https://doi.org/10.3390/nu10030265
Chicago/Turabian StyleDo, Moon Ho, Jinyoung Hur, Jiwon Choi, Mina Kim, Min Jung Kim, Yoonsook Kim, and Sang Keun Ha. 2018. "Eucommia ulmoides Ameliorates Glucotoxicity by Suppressing Advanced Glycation End-Products in Diabetic Mice Kidney" Nutrients 10, no. 3: 265. https://doi.org/10.3390/nu10030265
APA StyleDo, M. H., Hur, J., Choi, J., Kim, M., Kim, M. J., Kim, Y., & Ha, S. K. (2018). Eucommia ulmoides Ameliorates Glucotoxicity by Suppressing Advanced Glycation End-Products in Diabetic Mice Kidney. Nutrients, 10(3), 265. https://doi.org/10.3390/nu10030265