Protective Effects of LSGYGP from Fish Skin Gelatin Hydrolysates on UVB-Induced MEFs by Regulation of Oxidative Stress and Matrix Metalloproteinase Activity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Regents
2.2. Cell Culture
2.3. UVB Irradiation and Cell Viability Assay
2.4. Determination of Generated Intracellular ROS
2.5. Analysis of Intracellular SOD and MDA
2.6. ELISA Assays of Collagen I, MMP-1, and MMP-9
2.7. Molecular Docking Analysis
2.8. ELISA Analysis of JNK, ERK, and p38
2.9. Statistical Analysis
3. Results
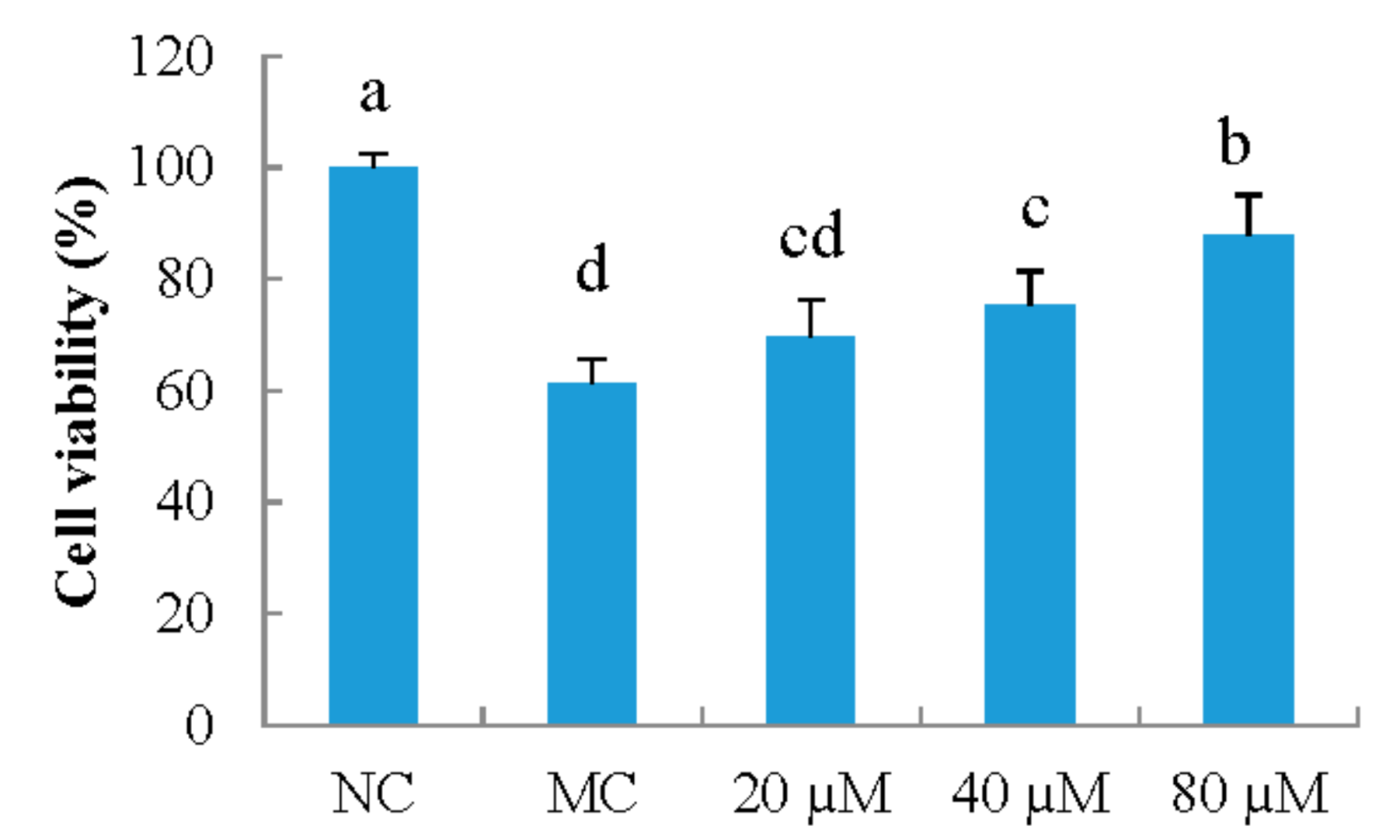
3.1. Cell Viability
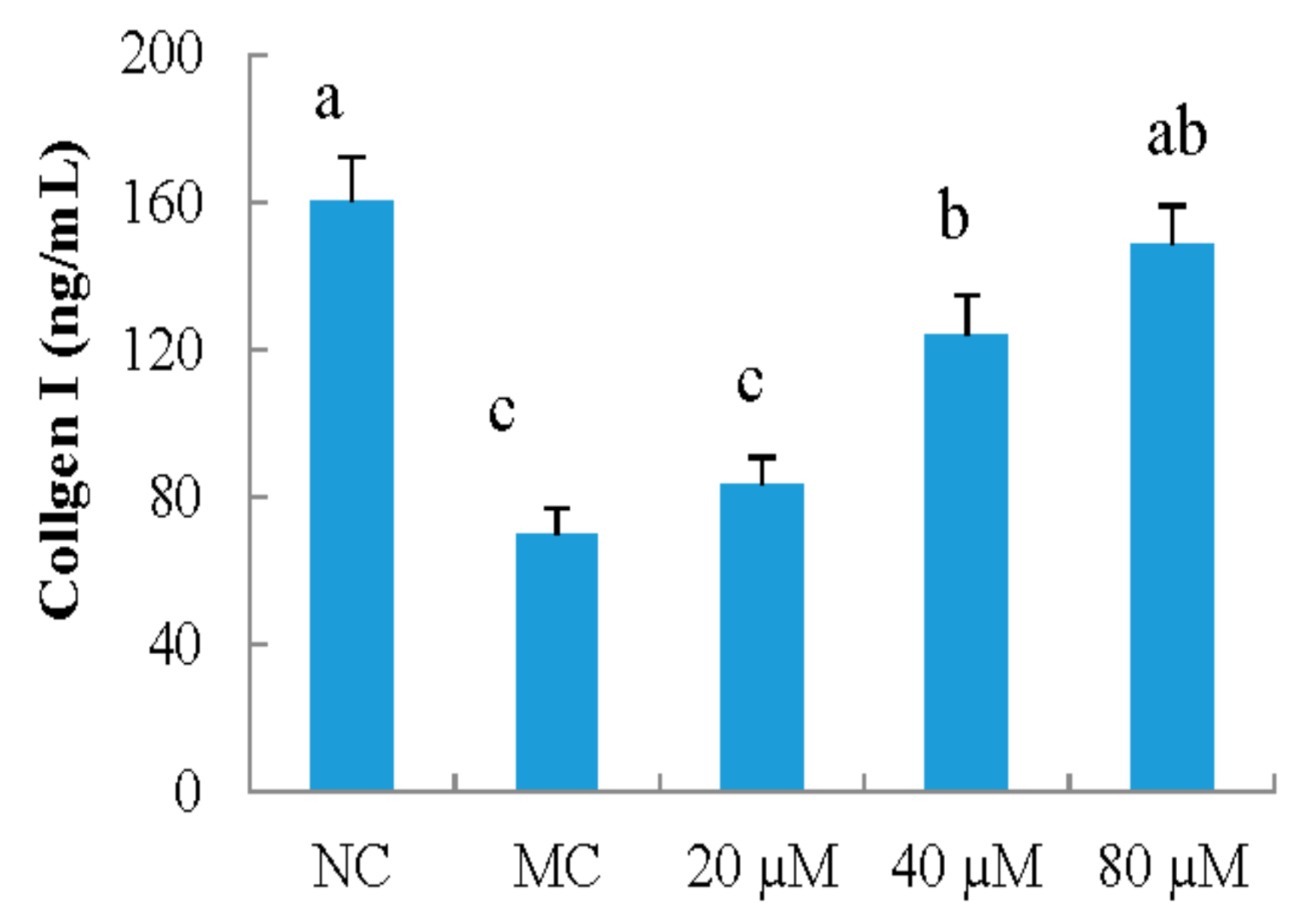
3.2. Collagen I
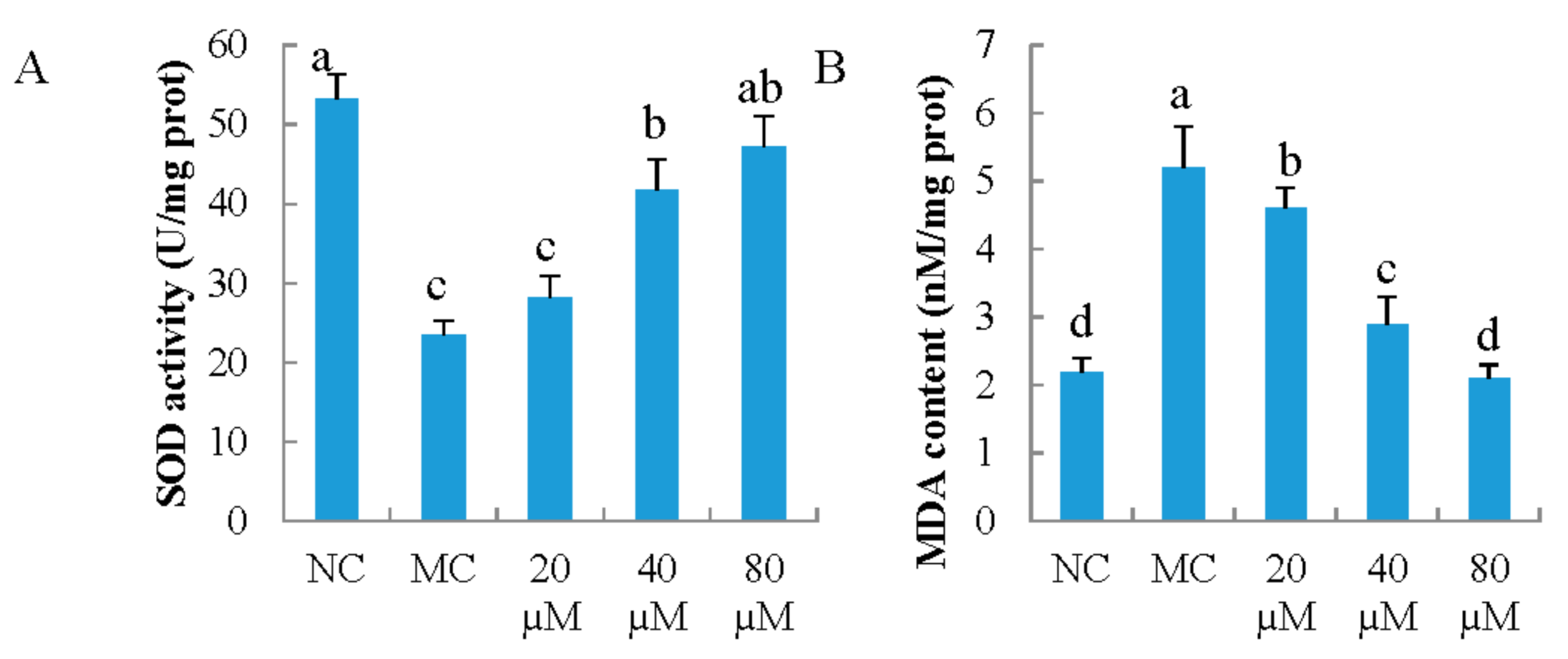
3.3. Antioxidant Indicators
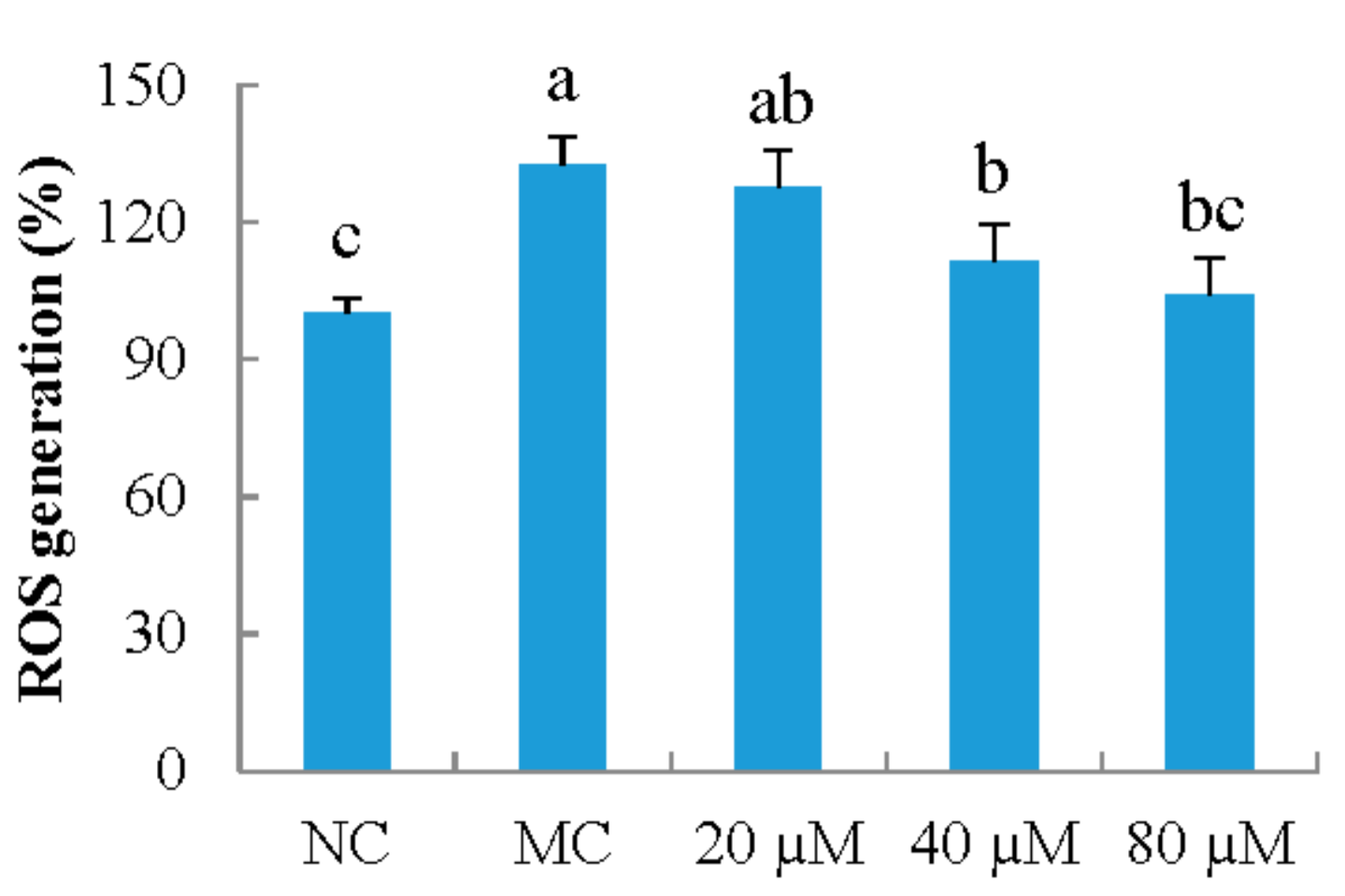
3.4. ROS Generation
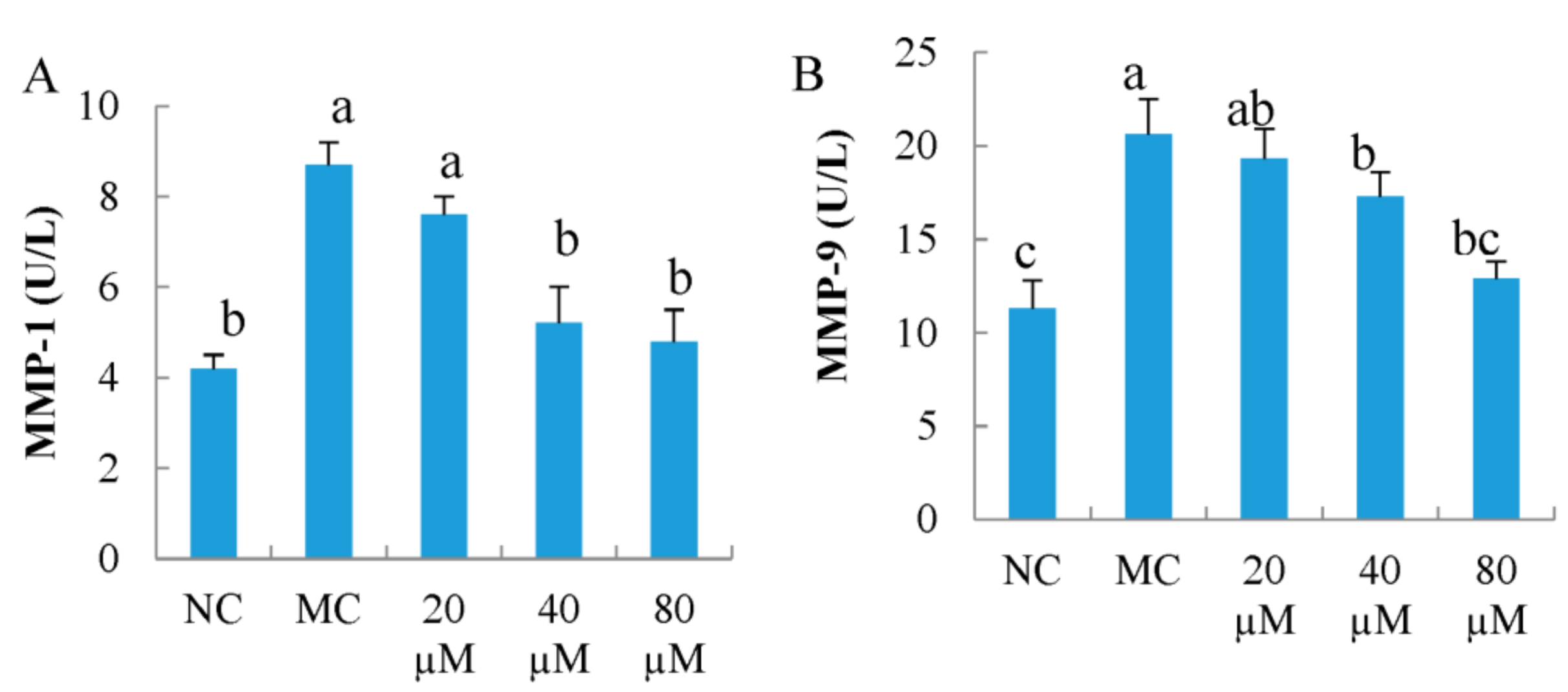
3.5. Intercellular MMP-1 and MMP-9 Activity
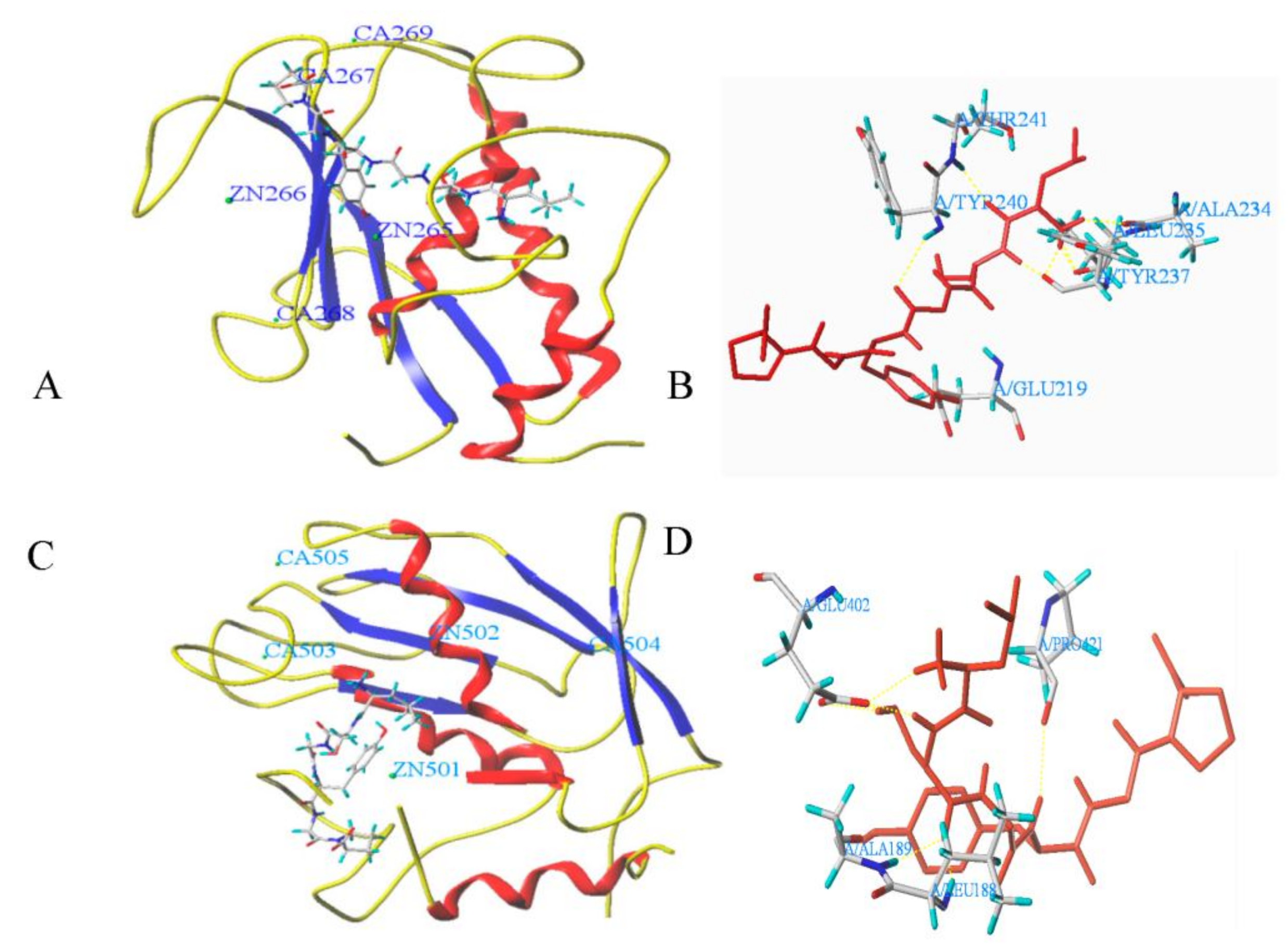
3.6. Molecular Docking
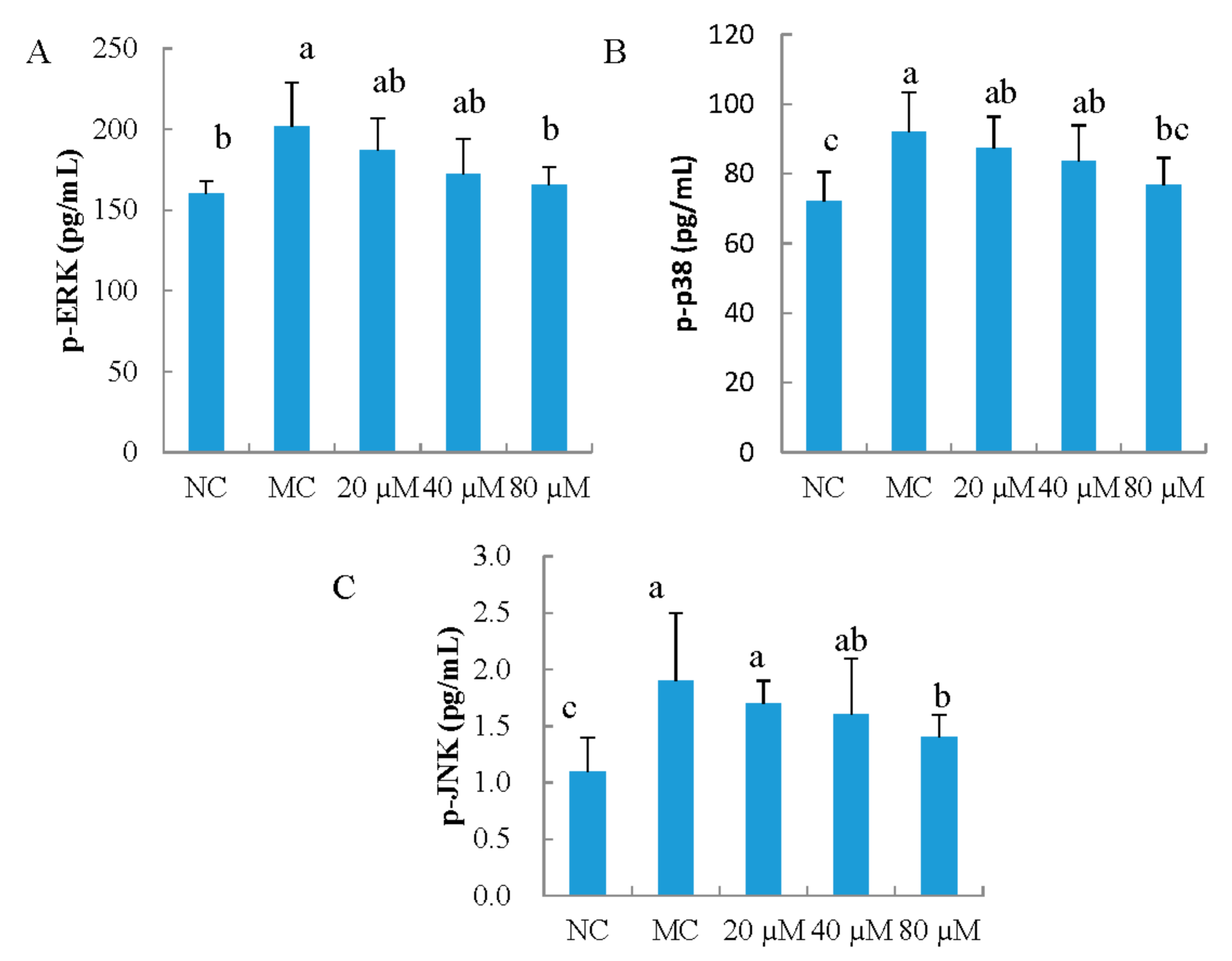
3.7. The Phosphorylation of ERK, JNK, and p38 in MEFs
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fan, J.; Zhuang, Y.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish umbrella on histological and immunity changes of mice photoaging. Nutrients 2013, 5, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Leirós, G.J.; Kusinsky, A.G.; Balañá, M.E.; Hagelin, K. Triolein reduces MMP-1 upregulation in dermal fibroblasts generated by ROS production in UVB-irradiated keratinocytes. J. Dermatol. Sci. 2017, 85, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.A.; Jung, M.; Lee, S.M.; Lee, T.H.; Kim, J. Protective effect of Disporum sessile D. Don extract against UVB-induced photoaging via suppressing MMP-1 expression and collagen degradation in human skin cells. J. Photochem. Photobiol. B 2014, 133, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.M.; Lin, T.J.; Chiu, C.Y.; Chang, C.W.; Hsu, K.C.; Fan, P.C.; Wena, K.C. Coffeaarabica extract and its constituents prevent photoaging by suppressing MMPs expression and MAP kinase pathway. Food Chem. Toxicol. 2011, 49, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.F.; Lee, H.; Jung, B.J.; Jang, S.; Chung, D.K.; Kim, H. Lipoteichoic acid isolated from Lactobacillus plantarum down-regulates UV-induced MMP-1 expression and up-regulates type I procollagen through the inhibition of reactive oxygen species generation. Mol. Immunol. 2015, 67, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Muller, F.L.; Lustgarten, M.S.; Jang, Y.; Richardson, A.; Van Remmen, H. Trends in oxidative aging theories. Free Radic. Biol. Med. 2007, 43, 477–503. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Cheung, I.W.Y.; Cheung, L.K.Y.; Tan, N.Y.; Li-Chan, E.C.Y. The role of molecular size in antioxidant activity of peptide fractions from Pacific hake (Merlucciu sproductus) hydrolysates. Food Chem. 2012, 134, 1297–1306. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Shi, Q.; Gninguue, A.; Wei, R.; Luo, H. Purification and identification of a novel peptide derived from by-products fermentation of spiny head croaker (Collichthys lucidus) with antifungal effects on phytopathogens. Process Biochem. 2017, 62, 184–192. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, C.; Jiang, A. Antioxidant peptides isolated from sea cucumber Stichopus Japonicus. Eur. Food Res. Technol. 2012, 234, 441–447. [Google Scholar] [CrossRef]
- Zhuang, Y.; Hou, H.; Zhao, X.; Zhang, Z.; Li, B. Effects of collagen and collagen hydrolysate from jellyfish (Rhopilema esculentum) on mice skin photoaging induced by UV irradiation. J. Food Sci. 2009, 74, H183–H188. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, H.; Fan, Y.; Wang, S.; Chen, Q.; Si, L.; Li, B. Protective effect of gelatin peptides from pacific cod skin against photoaging by inhibiting the expression of MMPs via MAPK signaling pathway. J. Photochem. Photobiol. B 2016, 165, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Liou, S.F.; Chen, S.J.; Shih, M.F. Protective effects of Chlorella-derived peptide on UVB-induced production of MMP-1 and degradation of procollagen genes in human skin fibroblasts. Regul. Toxicol. Pharm. 2011, 60, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Qian, Z.J.; Ryu, B.M.; Kim, K.N.; Kim, D.; Kim, Y.M.; Jeon, Y.J.; Park, W.S.; Choi, I.W.; Kim, G.H.; et al. Matrix metalloproteinases (MMPs) inhibitory effects of an octameric oligopeptide isolated from abalone Haliotis discus hannai. Food Chem. 2013, 141, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.M.; Qian, Z.J.; Kim, S.K. Purification of a peptide from seahorse, that inhibits TPA-induced MMP, iNOS and COX-2 expression through MAPK and NF-kappaB activation, and induces human osteoblastic and chondrocytic differentiation. Chem. Biol. Int. 2010, 184, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Hou, H.; Fan, Y.; Yang, T.; Li, B. Identification of MMP-1 inhibitory peptides from cod skin gelatin hydrolysates and the inhibition mechanism by MAPK signaling pathway. J. Func. Foods 2017, 33, 251–260. [Google Scholar] [CrossRef]
- Parthasarathy, A.; Gopi, V.; Devi, K.M.S.; Balaji, N.; Vellaichamy, E. Aminoguanidine inhibits ventricular fibrosis and remodeling process in isoproterenol-induced hypertrophied rat hearts by suppressing ROS and MMPs. Life Sci. 2014, 118, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhang, Y.; Zhuang, Y. Antiphotoaging effect and purification of an antioxidant peptide from tilapia (Oreochromis niloticus) gelatin peptides. J. Func. Foods 2013, 5, 154–162. [Google Scholar] [CrossRef]
- Ren, S.W.; Li, J.; Wang, W.; Guan, H.S. Protective effects of j-ca3000 + CP against ultraviolet-induced damage in HaCaT and MEF cells. J. Photochem. Photobiol. B. 2010, 101, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Ma, Y.; Sun, D.; Fan, J.; Cai, S. In vitro DNA damage protection and anti-inflammatory effects of Tartary buckwheats (FagopyrumtataricumL. Gaertn) fermented by filamentous fungi. Int. J. Food Sci. Technol. 2017, 52, 2006–2017. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Protective effects of rambutan (Nephelium lappaceum) peel phenolics on H2O2-induced oxidative damages in HepG2 cells and d-galactose-induced aging mice. Food Chem. Toxicol. 2017, 108, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, Q.; Fan, J.; Li, X.; Zhuang, Y. Purification and Characterization of Peptides Inhibiting MMP-1 Activity with C Terminate of Gly-Leu from Simulated Gastrointestinal Digestion Hydrolysates of Tilapia (Oreochromis niloticus) Skin Gelatin. J. Agric. Food Chem. 2018, 66, 593–601. [Google Scholar]
- Roomi, M.W.; Ivanov, V.; Niedzwiecki, A.; Rath, M. Synergistic antitumor effect of ascorbic acid, lysine, proline, and epigallocatechin gallate on human fibrosarcoma cells HT1080. Ann. Cancer Res. Ther. 2004, 12, 146–156. [Google Scholar] [CrossRef]
- Hou, H.; Li, B.; Zhao, X.; Zhuang, Y.; Ren, G.; Yan, M.; Zhang, X.; Chen, L.; Fan, Y. The effect of pacific cod (Gadusmacrocephalus) skin gelatin polypeptides on UV radiation induced skin photoaging in ICR mice. Food Chem. 2009, 115, 945–950. [Google Scholar] [CrossRef]
- Ma, Q.; Guo, Y.; Sun, L.; Zhuang, Y. Anti-diabetic effects of phenolic extract from rambutan peels (Nephelium lappaceum) in high-fat diet and streptozotocin-induced diabetic mice. Nutrients 2017, 9, 801. [Google Scholar] [CrossRef]
- Fanjul-Fernández, M.; Folgueras, A.R.; Cabrera, S.; López-Otín, C. Matrix metalloproteinases: Evolution, gene regulation and functional analysis in mouse models. Biochim. Biophys. Acta 2010, 1803, 3–19. [Google Scholar] [CrossRef] [PubMed]
- You, G.E.; Jung, B.J.; Kim, H.R.; Kim, H.G.; Kim, T.R.; Chung, D.K. Lactobacillus sakei lipoteichoic acid inhibits MMP-1 induced by UVA in normal dermal fibroblasts of human. J. Microbiol. Biotechnol. 2013, 23, 1357–1364. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Lu, W.; Wang, L.; Shan, L.; Li, H.; Huang, J.; Sun, Q.; Zhang, W. Synthesis of derivatives of methyl rosmarinate and their inhibitory activities against matrix metalloproteinase-1 (MMP-1). Eur. J. Med. Chem. 2013, 62, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by green tea catechins and prediction of their interaction by molecular docking analysis. Biomed. Pharm. 2016, 84, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; Deng, J.; Huang, S.; Li, P.; Liang, Y.; Huang, G. 3, 4-dihydroxybenzalacetone attenuates lipopolysaccharide-induced inflammation in acute lung injury via down-regulation of MMP-2 and MMP-9 activities through suppressing ROS-mediated MAPK and PI3K/AKT signaling pathways. Int. Immunopharm. 2017, 50, 77–86. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LSGYGP | T-Score | C-Score | Hydrogen Bond Number | Distance (Å) |
|---|---|---|---|---|
| MMP-1 | 7.30 | 5 | 8 | Glu219: 2.06; Tyr240: 2.62; Thr241: 2.02; Tyr237: 2.10/2.05; Leu235: 1.90/1.81/2.58 Zn265: 1.98 |
| MMP-9 | 8.53 | 4 | 7 | Pro421: 2.33; Leu188: 1.65; Ala189: 2.27; GLU402: 2.44/2.27/1.93/2.31; Zn1450: 2.02 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Liu, Q.; Yuan, L.; Zhuang, Y. Protective Effects of LSGYGP from Fish Skin Gelatin Hydrolysates on UVB-Induced MEFs by Regulation of Oxidative Stress and Matrix Metalloproteinase Activity. Nutrients 2018, 10, 420. https://doi.org/10.3390/nu10040420
Ma Q, Liu Q, Yuan L, Zhuang Y. Protective Effects of LSGYGP from Fish Skin Gelatin Hydrolysates on UVB-Induced MEFs by Regulation of Oxidative Stress and Matrix Metalloproteinase Activity. Nutrients. 2018; 10(4):420. https://doi.org/10.3390/nu10040420
Chicago/Turabian StyleMa, Qingyu, Qiuming Liu, Ling Yuan, and Yongliang Zhuang. 2018. "Protective Effects of LSGYGP from Fish Skin Gelatin Hydrolysates on UVB-Induced MEFs by Regulation of Oxidative Stress and Matrix Metalloproteinase Activity" Nutrients 10, no. 4: 420. https://doi.org/10.3390/nu10040420