Reducing Pup Litter Size Alters Early Postnatal Calcium Homeostasis and Programs Adverse Adult Cardiovascular and Bone Health in Male Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
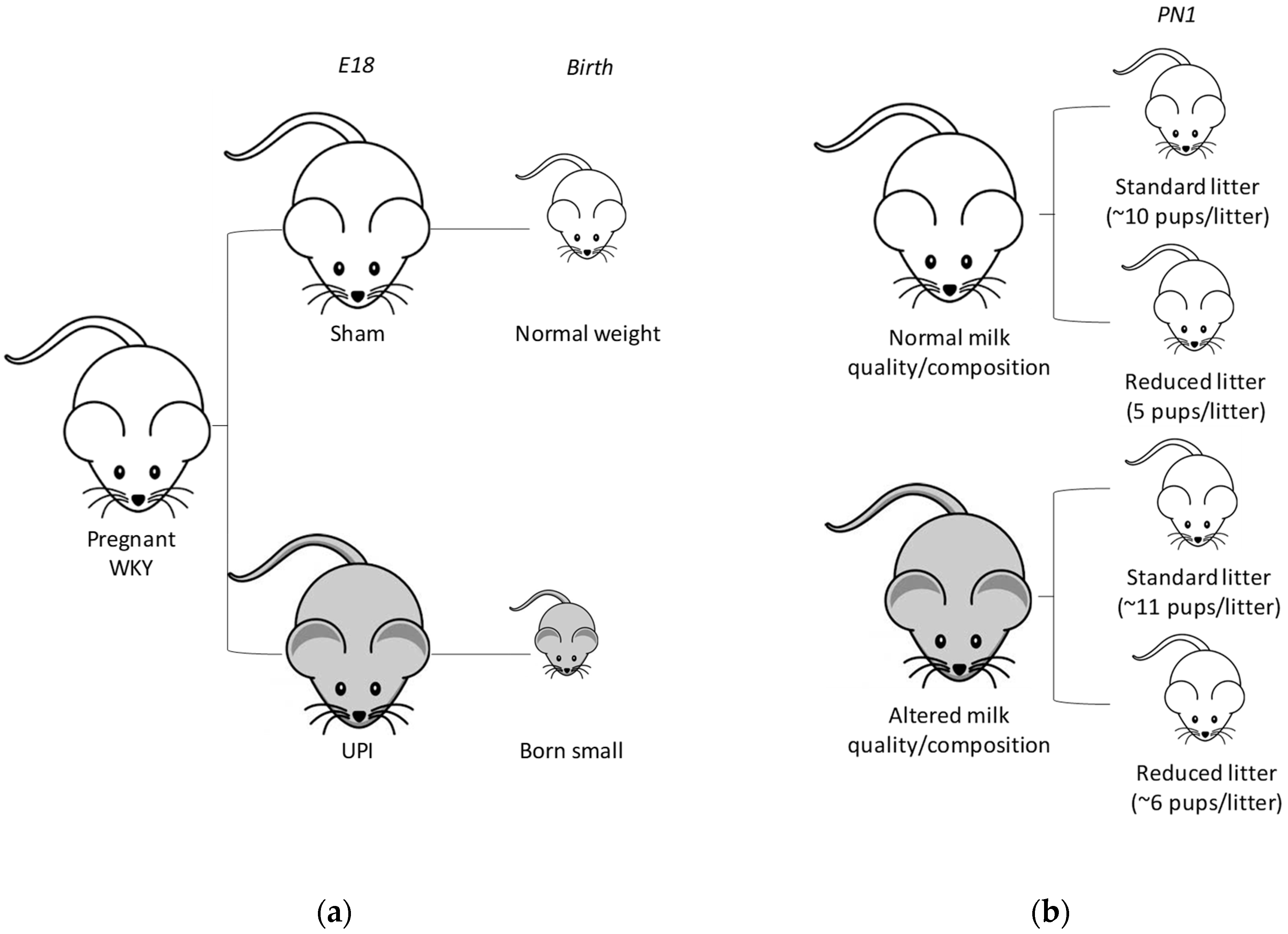
2.2. Study 1: Postnatal Study
2.2.1. Mammary ‘Real-Time’ PCR and Histology
2.2.2. PTHrP, Corticosterone, Calcium and Electrolyte Measurements
2.2.3. Fatty Acid Analysis
2.3. Study 2: Lactation and Adult Study
2.3.1. Bone Analyses
2.3.2. Plasma Analyses
2.4. Statistical Analysis
3. Results
3.1. Growth to Weaning
3.2. Mammary Structure and Maternal Plasma Analysis
3.3. Pup Plasma Analysis and Milk Consumption
3.4. Milk Fatty Acid Composition
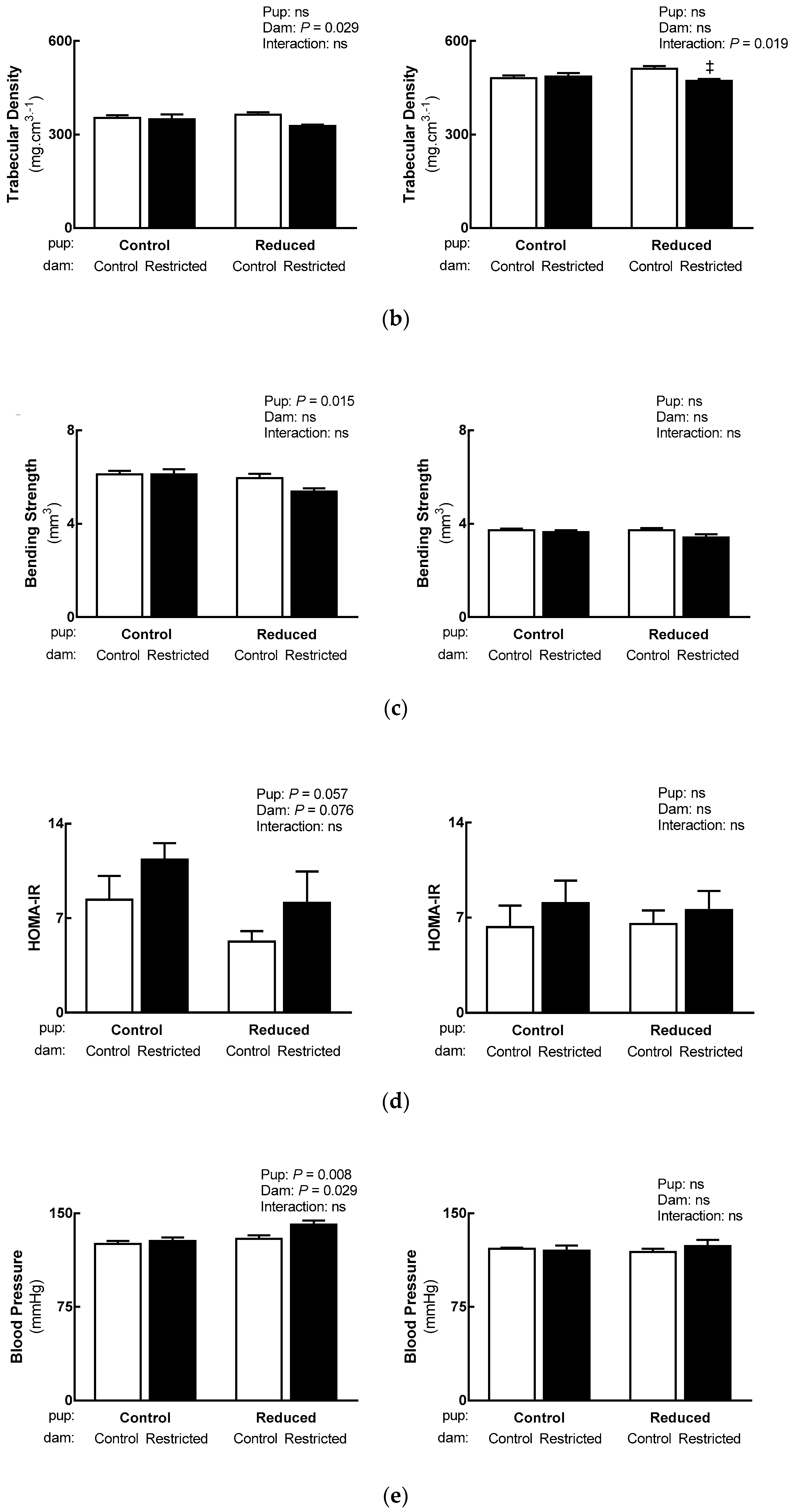
3.5. Adult Health
4. Discussion
4.1. Effects of Reducing Litter Size
4.2. Effects of Altered Milk Quality/Composition
4.3. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.; Winter, P.D.; Osmond, C.; Margetts, B.; Simmonds, S.J. Weight in infancy and death from ischaemic heart disease. Lancet 1989, 2, 577–580. [Google Scholar] [CrossRef]
- Barker, D.J.P.; Godfrey, K.M.; Osmond, C.; Bull, A. The relation of fetal length, ponderal index and head circumference to blood pressure and the risk of hypertension in adult life. Paediatr. Perinat. Epidemiol. 1992, 6, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Gluckman, P.D.; Harding, J.E. Fetal growth retardation: Underlying endocrine mechanisms and postnatal consequences. Acta Paediatr. Scand. 1997, 422, 69–72. [Google Scholar] [CrossRef]
- Wlodek, M.E.; Owens, J.A.; Siebel, A.L.; Moritz, K. Reduced nephron endowment and hypertension emerge following placental restriction in the rat. Pediatr. Res. 2005, 58, 1024. [Google Scholar]
- Painter, R.C.; Roseboom, T.J.; Bleker, O.P. Prenatal exposure to the dutch famine and disease in later life: An overview. Reprod. Toxicol. 2005, 20, 345–352. [Google Scholar] [CrossRef]
- Jaquiery, A.L.; Oliver, M.H.; Honeyfield-Ross, M.; Harding, J.E.; Bloomfield, F.H. Periconceptional undernutrition in sheep affects adult phenotype only in males. J. Nutr. Metab. 2012, 2012, 123610. [Google Scholar] [CrossRef]
- Sokol, R.J.; Delaney-Black, V.; Nordstrom, B. Fetal alcohol spectrum disorder. JAMA 2003, 290, 2996–2999. [Google Scholar] [CrossRef]
- Gray, S.P.; Denton, K.M.; Cullen-McEwen, L.; Bertram, J.F.; Moritz, K.M. Prenatal exposure to alcohol reduces nephron number and raises blood pressure in progeny. J. Am. Soc. Nephrol. 2010, 21, 1891–1902. [Google Scholar] [CrossRef]
- Virk, J.; Li, J.; Vestergaard, M.; Obel, C.; Kristensen, J.K.; Olsen, J. Prenatal exposure to bereavement and type-2 diabetes: A danish longitudinal population based study. PLoS ONE 2012, 7, e43508. [Google Scholar]
- Singh, R.R.; Cuffe, J.S.; Moritz, K.M. Short- and long-term effects of exposure to natural and synthetic glucocorticoids during development. Clin. Exp. Pharmacol. Physiol. 2012, 39, 979–989. [Google Scholar] [CrossRef]
- Mennitti, L.V.; Oliveira, J.L.; Morais, C.A.; Estadella, D.; Oyama, L.M.; Oller do Nascimento, C.M.; Pisani, L.P. Type of fatty acids in maternal diets during pregnancy and/or lactation and metabolic consequences of the offspring. J. Nutr. Biochem. 2015, 26, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Ailhaud, G.; Massiera, F.; Weill, P.; Legrand, P.; Alessandri, J.M.; Guesnet, P. Temporal changes in dietary fats: Role of n-6 polyunsaturated fatty acids in excessive adipose tissue development and relationship to obesity. Prog. Lipid Res. 2006, 45, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Gallo, L.A.; Tran, M.; Moritz, K.M.; Wlodek, M.E. Developmental programming: Variations in early growth and adult disease. Clin. Exp. Pharmacol. Physiol. 2013, 40, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Pomar, C.A.; van, N.R.; Sanchez, J.; Pico, C.; Keijer, J.; Palou, A. Maternal consumption of a cafeteria diet during lactation in rats leads the offspring to a thin-outside-fat-inside phenotype. Int. J. Obes. (Lond.) 2017, 41, 1279–1287. [Google Scholar] [CrossRef] [PubMed]
- Lozano, G.; Elmaghrabi, A.; Salley, J.; Siddique, K.; Gattineni, J.; Baum, M. Effect of prenatal programming and postnatal rearing on glomerular filtration rate in adult rats. Am. J. Physiol. Ren. Physiol. 2015, 308, F411–F419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddique, K.; Guzman, G.L.; Gattineni, J.; Baum, M. Effect of postnatal maternal protein intake on prenatal programming of hypertension. Reprod. Sci. 2014, 21, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, H.; Moss, T.J.; Gatford, K.L.; Moritz, K.M.; Akison, L.; Fullston, T.; Hryciw, D.H.; Maloney, C.A.; Morris, M.J.; Wooldridge, A.L.; et al. A review of fundamental principles for animal models of dohad research: An australian perspective. J. Dev. Orig. Health Dis. 2016, 7, 449–472. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, F.J.; Duque-Guimaraes, D.; Carpenter, A.A.; Loche, E.; Ozanne, S.E. A post-weaning obesogenic diet exacerbates the detrimental effects of maternal obesity on offspring insulin signaling in adipose tissue. Sci. Rep. 2017, 7, 44949. [Google Scholar] [CrossRef]
- Galyon, K.D.; Farshidi, F.; Han, G.; Ross, M.G.; Desai, M.; Jellyman, J.K. Maternal bisphenol a exposure alters rat offspring hepatic and skeletal muscle insulin signaling protein abundance. Am. J. Obstet. Gynecol. 2017, 216, 290.e1–290.e9. [Google Scholar] [CrossRef]
- Maniam, J.; Antoniadis, C.P.; Wang, K.W.; Morris, M.J. Early life stress induced by limited nesting material produces metabolic resilience in response to a high-fat and high-sugar diet in male rats. Front. Endocrinol. (Lausanne) 2015, 6, 138. [Google Scholar] [CrossRef]
- Firth, E.C.; Gamble, G.D.; Cornish, J.; Vickers, M.H. Neonatal leptin treatment reverses the bone-suppressive effects of maternal undernutrition in adult rat offspring. Sci. Rep. 2017, 7, 7686. [Google Scholar] [CrossRef] [PubMed]
- Dasinger, J.H.; Intapad, S.; Backstrom, M.A.; Carter, A.J.; Alexander, B.T. Intrauterine growth restriction programs an accelerated age-related increase in cardiovascular risk in male offspring. Am. J. Physiol. Ren. Physiol. 2016, 311, F312–F319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, E.; Sosa-Larios, T.; Calzada, L.; Ibanez, C.A.; Mendoza-Rodriguez, C.A.; Morales, A.; Morimoto, S. Decreased basal insulin secretion from pancreatic islets of pups in a rat model of maternal obesity. J. Endocrinol. 2016, 231, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Herrera, E.A.; Evans, R.D.; Giussani, D.A. Antioxidant treatment improves neonatal survival and prevents impaired cardiac function at adulthood following neonatal glucocorticoid therapy. J. Physiol. 2013, 591, 5083–5093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, A.; Reyes, L.M.; Morton, J.S.; Fung, D.; Schneider, J.; Davidge, S.T. Effect of resveratrol on metabolic and cardiovascular function in male and female adult offspring exposed to prenatal hypoxia and a high-fat diet. J. Physiol. 2016, 594, 1465–1482. [Google Scholar] [CrossRef] [PubMed]
- Chahoud, I.; Paumgartten, F.J. Influence of litter size on the postnatal growth of rat pups: Is there a rationale for litter-size standardization in toxicity studies? Environ. Res. 2009, 109, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Wlodek, M.E.; Westcott, K.; Siebel, A.L.; Owens, J.A.; Moritz, K.M. Growth restriction before or after birth reduces nephron number and increases blood pressure in male rats. Kidney Int. 2008, 74, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tare, M.; Parkington, H.C.; Bubb, K.J.; Wlodek, M.E. Uteroplacental insufficiency and lactational environment separately influence arterial stiffness and vascular function in adult male rats. Hypertension 2012, 60, 378–386. [Google Scholar] [CrossRef]
- Tran, M.; Young, M.E.; Jefferies, A.J.; Hryciw, D.H.; Ward, M.M.; Fletcher, E.L.; Wlodek, M.E.; Wadley, G.D. Uteroplacental insufficiency leads to hypertension, but not glucose intolerance or impaired skeletal muscle mitochondrial biogenesis, in 12-month-old rats. Physiol. Rep. 2015, 3, e12556. [Google Scholar] [CrossRef]
- O’Dowd, R.; Kent, J.C.; Moseley, J.M.; Wlodek, M.E. Effects of uteroplacental insufficiency and reducing litter size on maternal mammary function and postnatal offspring growth. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R539–R548. [Google Scholar] [CrossRef]
- Romano, T.; Wark, J.D.; Owens, J.A.; Wlodek, M.E. Prenatal growth restriction and postnatal growth restriction followed by accelerated growth independently program reduced bone growth and strength. Bone 2009, 45, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Habbout, A.; Guenancia, C.; Lorin, J.; Rigal, E.; Fassot, C.; Rochette, L.; Vergely, C. Postnatal overfeeding causes early shifts in gene expression in the heart and long-term alterations in cardiometabolic and oxidative parameters. PLoS ONE 2013, 8, e56981. [Google Scholar] [CrossRef] [PubMed]
- Habbout, A.; Delemasure, S.; Goirand, F.; Guilland, J.C.; Chabod, F.; Sediki, M.; Rochette, L.; Vergely, C. Postnatal overfeeding in rats leads to moderate overweight and to cardiometabolic and oxidative alterations in adulthood. Biochimie 2012, 94, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Briffa, J.F.; O’Dowd, R.; Moritz, K.M.; Romano, T.; Jedwab, L.R.; McAinch, A.J.; Hryciw, D.H.; Wlodek, M.E. Uteroplacental insufficiency reduces rat plasma leptin concentrations and alters placental leptin transporters: Ameliorated with enhanced milk intake and nutrition. J. Physiol. 2017, 595, 3389–3407. [Google Scholar] [CrossRef] [PubMed]
- Wlodek, M.E.; Westcott, K.T.; O’Dowd, R.; Serruto, A.; Wassef, L.; Moritz, K.M.; Moseley, J.M. Uteroplacental restriction in the rat impairs fetal growth in association with alterations in placental growth factors including pthrp. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1620–R1627. [Google Scholar] [CrossRef] [PubMed]
- Wlodek, M.E.; Mibus, A.; Tan, A.; Siebel, A.L.; Owens, J.A.; Moritz, K.M. Normal lactational environment restores nephron endowment and prevents hypertension after placental restriction in the rat. J. Am. Soc. Nephrol. 2007, 18, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Moritz, K.M.; Mazzuca, M.Q.; Siebel, A.L.; Mibus, A.; Arena, D.; Tare, M.; Owens, J.A.; Wlodek, M.E. Uteroplacental insufficiency causes a nephron deficit, modest renal insufficiency but no hypertension with ageing in female rats. J. Physiol. 2009, 587, 2635–2646. [Google Scholar] [CrossRef] [Green Version]
- Siebel, A.L.; Mibus, A.; De Blasio, M.J.; Westcott, K.T.; Morris, M.J.; Prior, L.; Owens, J.A.; Wlodek, M.E. Improved lactational nutrition and postnatal growth ameliorates impairment of glucose tolerance by uteroplacental insufficiency in male rat offspring. Endocrinology 2008, 149, 3067–3076. [Google Scholar] [CrossRef]
- Wlodek, M.E.; Westcott, K.T.; Ho, P.W.M.; Serruto, A.; Di Nicolantonio, R.; Farrugia, W.; Moseley, J.M. Reduced fetal, placental, and amniotic fluid pthrp in the growth-restricted spontaneously hypertensive rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R31–R38. [Google Scholar] [CrossRef]
- Wlodek, M.E.; Westcott, K.T.; Serruto, A.; O’Dowd, R.; Wassef, L.; Ho, P.W.M.; Moseley, J.M. Impaired mammary function and parathyroid hormone-related protein during lactation in growth-restricted spontaneously hypertensive rats. J. Endocrinol. 2003, 177, 233–245. [Google Scholar] [CrossRef]
- Wlodek, M.E.; Ho, P.W.M.; Rice, G.E.; Moseley, J.M.; Martin, T.J.; Brennecke, S.P. Parathyroid hormone-related protein (pthrp) concentrations in human amniotic fluid during gestation and at the time of labour. Reprod. Fertil. Dev. 1995, 7, 1509–1513. [Google Scholar] [CrossRef] [PubMed]
- Wlodek, M.E.; Ceranic, V.; O’Dowd, R.; Westcott, K.T.; Siebel, A.L. Maternal progesterone treatment rescues the mammary impairment following uteroplacental insufficiency and improves postnatal pup growth in the rat. Reprod. Sci. 2009, 16, 380–390. [Google Scholar] [CrossRef]
- LePage, G.; Roy, C.C. Direct transesterification of all lipid classes in a one-step reaction. J. Lipid Res. 1986, 27, 114–120. [Google Scholar] [PubMed]
- Weisinger, H.S.; Vingrys, A.; Bui, B.; Sinclair, A.J. Effects of dietary n-3 fatty acid deficiency and repletion in the guinea pig retina. Investig. Ophthalmol. Vis. Sci. 1999, 40, 327–338. [Google Scholar]
- Anevska, K.; Gallo, L.A.; Tran, M.; Jefferies, A.J.; Wark, J.D.; Wlodek, M.E.; Romano, T. Pregnant growth restricted female rats have bone gains during late gestation which contributes to second generation adolescent and adult offspring having normal bone health. Bone 2015, 74, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Romano, T.; Wark, J.D.; Wlodek, M.E. Developmental programming of bone deficits in growth-restricted offspring. Reprod. Fertil. Dev. 2014, 27, 823–833. [Google Scholar] [CrossRef]
- Anevska, K.; Cheong, J.N.; Wark, J.D.; Wlodek, M.E.; Romano, T. Maternal stress does not exacerbate long-term bone deficits in female rats born growth restricted, with differential effects on offspring bone health. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 314, R161–R170. [Google Scholar] [CrossRef]
- Wadley, G.D.; Siebel, A.L.; Cooney, G.J.; McConell, G.K.; Wlodek, M.E.; Owens, J.A. Uteroplacental insufficiency and reducing litter size alters skeletal muscle mitochondrial biogenesis in a sex specific manner in the adult rat. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E861–E869. [Google Scholar] [CrossRef]
- Rijnkels, M.; Kabotyanski, E.; Montazer-Torbati, M.B.; Hue, B.C.; Vassetzky, Y.; Rosen, J.M.; Devinoy, E. The epigenetic landscape of mammary gland development and functional differentiation. J. Mammary Gland Biol. Neoplasia 2010, 15, 85–100. [Google Scholar] [CrossRef]
- Kukreja, S.C.; D’Anza, J.J.; Melton, M.E.; Wimbiscus, S.A.; Grill, V.; Martin, T.J. Lack of effects of neutralisation of parathyroid hormone-related protein on calcium homeostasis in neonatal mice. J. Bone Miner. Res. 1991, 6, 1197–1201. [Google Scholar] [CrossRef]
- Kovacs, C.S. Calcium metabolism during pregnancy and lactation. J. Mammary Gland Biol. Neoplasia 2005, 10, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, C.S. Bone development and mineral homeostasis in the fetus and neonate: Roles of the calciotropic and phosphotropic hormones. Physiol. Rev. 2014, 94, 1143–1218. [Google Scholar] [CrossRef] [PubMed]
- Hirahatake, K.M.; Slavin, J.L.; Maki, K.C.; Adams, S.H. Associations between dairy foods, diabetes, and metabolic health: Potential mechanisms and future directions. Metabolism 2014, 63, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Permyakov, E.A.; Berliner, L.J. Alpha-lactalbumin: Structure and function. FEBS Lett. 2000, 473, 269–274. [Google Scholar] [CrossRef]
- Gregersen, S.; Bystrup, S.; Overgaard, A.; Jeppesen, P.B.; Sonderstgaard Thorup, A.C.; Jensen, E.; Hermansen, K. Effects of whey proteins on glucose metabolism in normal wistar rats and zucker diabetic fatty (zdf) rats. Rev. Diabet. Stud. 2013, 10, 252–269. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, J.; Forsen, T.; Tuomilehto, J.; Osmond, C.; Barker, D. Fetal and childhood growth and hypertension in adult life. Hypertension 2000, 36, 790–794. [Google Scholar] [CrossRef]
- Lucas, A.; Fewtrell, M.S.; Davies, P.S.; Bishop, N.J.; Clough, H.; Cole, T.J. Breastfeeding and catch-up growth in infants born small for gestational age. Acta Paediatr. 1997, 86, 564–569. [Google Scholar] [CrossRef]
- Eriksson, J.G.; Forsén, T.; Tuomilehto, J.; Osmond, C.; Barker, D.J.P. Early growth and coronary heart disease in later life: Longitudinal study. BMJ 2001, 322, 948–953. [Google Scholar] [CrossRef]
- Galante, L.; Milan, A.M.; Reynolds, C.M.; Cameron-Smith, D.; Vickers, M.H.; Pundir, S. Sex-specific human milk composition: The role of infant sex in determining early life nutrition. Nutrients 2018, 10, 1194. [Google Scholar] [CrossRef]
- Thakkar, S.K.; Giuffrida, F.; Cristina, C.H.; De Castro, C.A.; Mukherjee, R.; Tran, L.A.; Steenhout, P.; Lee le, Y.; Destaillats, F. Dynamics of human milk nutrient composition of women from singapore with a special focus on lipids. Am. J. Hum. Biol. 2013, 25, 770–779. [Google Scholar] [CrossRef]
- Curley, J.P.; Champagne, F.A. Influence of maternal care on the developing brain: Mechanisms, temporal dynamics and sensitive periods. Front. Neuroendocrinol. 2016, 40, 52–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enes-Marques, S.; Giusti-Paiva, A. Litter size reduction accentuates maternal care and alters behavioral and physiological phenotypes in rat adult offspring. J. Physiol. Sci. 2018, 68, 789–798. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pup-on-Dam | Standard-on-Normal | Standard-on-Altered | Reduced-on-Normal | Reduced-on-Altered | Two-Way ANOVA | |||
|---|---|---|---|---|---|---|---|---|
| Pup | Dam | Interaction | ||||||
| Litter Size | ||||||||
| PN1 | 10.4 ± 0.4 | 11.5 ± 0.9 | 5.0 ± 0.0 | 6.0 ± 0.7 | p = 0.0001 | ns 1 | ns | |
| PN6 | 8.9 ± 0.6 | 8.6 ± 0.8 | 4.7 ± 0.3 | 5.25 ± 0.8 | p = 0.0001 | ns | ns | |
| Cannibalism | 1.5 ± 0.5 | 2.9 ± 1.0 | 1.0 ± 0.7 | 0.8 ± 0.3 | ns | ns | ns | |
| Body Weight (g) | ||||||||
| PN1 | Male | 4.17 ± 0.12 | 4.13 ± 0.10 | 4.23 ± 0.15 | 3.99 ± 0.13 | ns | ns | ns |
| Female | 3.87 ± 0.12 | 3.98 ± 0.06 | 3.98 ± 0.06 | 3.94 ± 0.11 | ns | ns | ns | |
| PN3 | Male | 5.62 ± 0.11 | 5.42 ± 0.15 | 5.41 ± 0.14 | 5.12 ± 0.15 | ns | ns | ns |
| Female | 5.38 ± 0.12 | 5.16 ± 0.20 | 5.23 ± 0.14 | 4.91 ± 0.14 | ns | ns | ns | |
| PN6 | Male | 8.54 ± 0.21 | 8.31 ± 0.23 | 8.66 ± 0.38 | 7.65 ± 0.30 | ns | p = 0.039 | ns |
| Female | 8.17 ± 0.21 | 7.91 ± 0.37 | 8.29 ± 5.14 | 7.26 ± 0.28 | ns | p = 0.049 | ns | |
| PN10 | Male | 14.6 ± 0.3 | 14.1 ± 0.4 | 14.6 ± 0.5 | 13.1 ± 0.5 | ns | p = 0.037 | ns |
| Female | 14.2 ± 0.3 | 13.6 ± 0.6 | 14.1 ± 0.5 | 12.7 ± 0.5 | ns | p = 0.040 | ns | |
| PN14 | Male | 22.1 ± 0.5 | 21.2 ± 0.6 | 21.5 ± 0.7 | 19.6 ± 1.0 | ns | p = 0.043 | ns |
| Female | 21.7 ± 0.4 | 20.6 ± 0.8 | 20.6 ± 0.8 | 19.0 ± 0.9 | ns | ns | ns | |
| PN17 | Male | 27.4 ± 0.6 | 26.3 ± 0.6 | 26.9 ± 0.8 | 24.5 ± 1.2 | ns | p = 0.030 | ns |
| Female | 26.9 ± 0.5 | 25.6 ± 0.7 | 26.0 ± 0.8 | 24.1 ± 1.1 | ns | p = 0.047 | ns | |
| PN21 | Male | 34.3 ± 0.6 | 33.6 ± 1.0 | 34.7 ± 0.7 | 31.4 ± 1.1 | ns | p = 0.020 | ns |
| Female | 33.8 ± 0.6 | 32.8 ± 1.0 | 33.5 ± 0.9 | 31.4 ± 1.0 | ns | ns | ns | |
| PN24 | Male | 43.7 ± 1.1 | 42.3 ± 0.9 | 44.3 ± 1.0 | 39.2 ± 1.4 | ns | p = 0.004 | ns |
| Female | 42.1 ± 0.8 | 40.4 ± 1.1 | 42.4 ± 1.1 | 38.5 ± 1.2 | ns | p = 0.010 | ns | |
| PN28 | Male | 58.7 ± 1.2 | 57.4 ± 1.1 | 59.6 ± 1.2 | 54.0 ± 1.8 | ns | p = 0.011 | ns |
| Female | 54.9 ± 1.0 | 54.1 ± 1.4 | 55.0 ± 1.4 | 51.3 ± 1.3 | ns | ns | ns | |
| PN35 | Male | 87.7 ± 1.7 | 86.6 ± 1.7 | 87.1 ± 1.3 | 82.8 ± 2.8 | ns | ns | ns |
| Female | 79.1 ± 1.2 | 77.8 ± 1.8 | 77.0 ± 1.4 | 75.7 ± 1.8 | ns | ns | ns | |
| Pup-on-Dam | Standard-on-Normal | Standard-on-Altered | Reduced-on-Normal | Reduced-on-Altered | Two-Way ANOVA | ||
|---|---|---|---|---|---|---|---|
| Pup | Dam | Interaction | |||||
| Maternal (n = 4–9 per group) | |||||||
| Plasma PTHrP (pmol/L) | 8.2 ± 0.4 | 9.1 ± 1.2 | 10.5 ± 0.8 | 9.6 ± 0.7 | ns 1 | ns | ns |
| Plasma total calcium (mmol/L) | 2.6 ± 0.08 | 2.6 ± 0.04 | 2.6 ± 0.05 | 2.4 ± 0.04 | ns | ns | ns |
| Plasma corticosterone (ng/mL) | 593 ± 59 | 726 ± 106 | 551 ± 86 | 553 ± 80 | ns | ns | ns |
| Mammary (n = 3–6 per group) | |||||||
| Alveolar number | 14.5 ± 1.6 | 15.9 ± 0.8 | 11.4 ± 1.2 | 16.5 ± 1.7 | ns | p = 0.038 | ns |
| Alveolar area | 75.2 ± 3.0 | 75.1 ± 1.9 | 79.8 ± 6.5 | 78.5 ± 3.9 | ns | ns | ns |
| PTHrP mRNA | 1.0 ± 0.1 | 0.8 ± 0.3 | 1.1 ± 0.2 | 1.4 ± 0.3 | ns | ns | ns |
| Milk (n = 4–8 per group) | |||||||
| Na/K | 7.6 ± 1.2 | 6.5 ± 1.6 | 9.8 ± 2.5 | 6.4 ± 0.9 | ns | ns | ns |
| Ionic calcium (mmol/L) | 8.9 ± 0.6 | 8.7 ± 1.0 | 7.4 ± 0.9 | 9.1 ± 0.4 | ns | ns | ns |
| Total calcium (mmol/L) | 64.7 ± 4.0 | 65.8 ± 3.7 | 60.4 ± 5.6 | 57.9 ± 2.8 | ns | ns | ns |
| Total protein (mg/L) | 28.2 ± 4.4 | 22.0 ± 3.7 | 25.4 ± 4.5 | 28.1 ± 3.6 | ns | ns | ns |
| Lactose (mM) | 30.2 ± 7.9 | 38.4 ± 3.8 | 29.5 ± 6.0 | 41.7 ± 10.4 | ns | ns | ns |
| Pup-on-Dam | Standard-on-Normal | Standard-on-Altered | Reduced-on-Normal | Reduced-on-Altered | Two-Way ANOVA | ||
|---|---|---|---|---|---|---|---|
| Pup | Dam | Interaction | |||||
| Fatty Acids | |||||||
| Omega-6 PUFA | |||||||
| Linoleic (18:2n-6) | 11.79 ± 0.25 | 14.12 ± 0.43 ‡1 | 14.23 ± 0.26 *2 | 13.85 ± 0.50 | p = 0.010 | p = 0.019 | p = 0.002 |
| Arachidonic (20:4n-6) | 1.03 ± 0.06 | 1.33 ± 0.12 | 1.52 ± 0.08 * | 1.35 ± 0.09 | p = 0.014 | ns 3 | p = 0.023 |
| Total n-6 PUFA | 14.4 ± 0.4 | 17.4 ± 0.7 ‡ | 17.9 ± 0.4 * | 17.1 ± 0.7 | p = 0.009 | ns | p = 0.003 |
| Omega-3 PUFA | |||||||
| α-linolenic (18:3n-3) | 1.40 ± 0.06 | 1.48 ± 0.09 | 1.49 ± 0.05 | 1.46 ± 0.11 | ns | ns | ns |
| Eicosapentaenoic (20:5n-3) | 0.21 ± 0.01 | 0.17 ± 0.03 | 0.21 ± 0.02 | 0.18 ± 0.04 | ns | ns | ns |
| Docosahexaenoic (22:6n-3) | 0.41 ± 0.03 | 0.45 ± 0.06 | 0.55 ± 0.04 | 0.58 ± 0.09 | p = 0.047 | ns | ns |
| Total n-3 PUFA | 2.4 ± 0.09 | 2.4 ± 0.18 | 2.7 ± 0.15 | 2.7 ± 0.21 | ns | ns | ns |
| n-6:n-3 | 6.07 ± 0.15 | 7.11 ± 0.27 | 6.58 ± 0.28 | 6.63 ± 0.54 | ns | ns | ns |
| Total LC-MUFA | 22.4 ± 0.9 | 22.4 ± 1.4 | 25.8 ± 0.9 | 23.9 ± 0.8 | p = 0.032 | ns | ns |
| Saturated Fatty Acids | |||||||
| Medium (6–12) | 19.8 ± 1.0 | 21.1 ± 0.7 | 21.1 ± 0.7 | 20.8 ± 0.9 | ns | ns | ns |
| Long (14–20) | 40.4 ± 1.1 | 35.2 ± 2.1 | 30.9 ± 1.4 * | 34.2 ± 1.9 | p = 0.006 | ns | p = 0.022 |
| Pup-on-Dam | Standard-on-Normal | Standard-on-Altered | Reduced-on-Normal | Reduced-on-Altered | Two-Way ANOVA | |||
|---|---|---|---|---|---|---|---|---|
| Pup | Dam | Interaction | ||||||
| Body Weight (g) | Male | 386.1 ± 7.0 | 368.9 ± 2.3 | 368.4 ± 7.0 | 351.8 ± 9.5 | p = 0.020 | p = 0.023 | ns |
| Female | 239.3 ± 4.1 | 233.1 ± 6.5 | 238.3 ± 3.7 | 226.9 ± 4.7 | ns 1 | ns | ns | |
| Bone Parameters | ||||||||
| Femur Length (mm) | Male | 37.01 ± 0.12 | 36.79 ± 0.23 | 36.61 ± 0.21 | 36.01 ± 0.16 | ns | ns | ns |
| Female | 33.09 ± 0.14 | 32.75 ± 0.19 | 32.69 ± 0.14 | 32.42 ± 0.15 | ns | ns | ns | |
| Cortical content (mg·mm−1) | Male | 10.67 ± 0.15 | 10.63 ± 0.28 | 10.74 ± 0.18 | 9.93 ± 0.13 | ns | p = 0.005 | ns |
| Female | 7.87 ± 0.09 | 7.79 ± 0.11 | 7.90 ± 0.12 | 7.49 ± 0.11 | ns | ns | ns | |
| Cortical density (mg (mm3)−1) | Male | 1409.4 ± 1.8 | 1413.3 ± 3.7 | 1412.6 ± 3.8 | 1407.9 ± 1.9 | ns | ns | ns |
| Female | 1407.9 ± 2.5 | 1412.4 ± 1.2 | 1404.8 ± 1.8 | 1399.1 ± 2.0 | ns | ns | ns | |
| Metabolic Parameters | ||||||||
| Fasting plasma glucose (mmol·L−1) | Male | 5.77 ± 0.21 | 5.81 ± 0.19 | 5.52 ± 0.19 | 5.64 ± 0.27 | ns | ns | ns |
| Female | 5.15 ± 0.18 | 5.69 ± 0.18 | 5.50 ± 0.20 | 5.64 ± 0.22 | ns | ns | ns | |
| Fasting plasma insulin (ng·ML−1) | Male | 1.28 ± 0.24 | 1.82 ± 0.17 | 0.91 ± 0.12 | 1.32 ± 0.33 | p = 0.070 | p = 0.048 | ns |
| Female | 1.07 ± 0.24 | 1.23 ± 0.23 | 1.09 ± 0.14 | 1.23 ± 0.21 | ns | ns | ns | |
| Glucose AUC | Male | 757.3 ± 87.0 | 795.6 ± 79.8 | 742.2 ± 67.1 | 989.6 ± 62.1 | ns | P = 0.080 | ns |
| Female | 812.9 ± 74.1 | 701.9 ± 70.1 | 689.3 ± 52.8 | 635.2 ± 45.2 | ns | ns | ns | |
| First phase insulin AUC | Male | 14.25 ± 1.54 | 10.01 ± 2.08 | 11.31 ± 1.92 | 8.53 ± 1.17 | ns | p = 0.069 | ns |
| Female | 13.91 ± 2.02 | 11.29 ± 2.05 | 13.81 ± 1.74 | 14.01 ± 1.84 | ns | ns | ns | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briffa, J.F.; O’Dowd, R.; Romano, T.; Muhlhausler, B.S.; Moritz, K.M.; Wlodek, M.E. Reducing Pup Litter Size Alters Early Postnatal Calcium Homeostasis and Programs Adverse Adult Cardiovascular and Bone Health in Male Rats. Nutrients 2019, 11, 118. https://doi.org/10.3390/nu11010118
Briffa JF, O’Dowd R, Romano T, Muhlhausler BS, Moritz KM, Wlodek ME. Reducing Pup Litter Size Alters Early Postnatal Calcium Homeostasis and Programs Adverse Adult Cardiovascular and Bone Health in Male Rats. Nutrients. 2019; 11(1):118. https://doi.org/10.3390/nu11010118
Chicago/Turabian StyleBriffa, Jessica F., Rachael O’Dowd, Tania Romano, Beverly S. Muhlhausler, Karen M. Moritz, and Mary E. Wlodek. 2019. "Reducing Pup Litter Size Alters Early Postnatal Calcium Homeostasis and Programs Adverse Adult Cardiovascular and Bone Health in Male Rats" Nutrients 11, no. 1: 118. https://doi.org/10.3390/nu11010118
APA StyleBriffa, J. F., O’Dowd, R., Romano, T., Muhlhausler, B. S., Moritz, K. M., & Wlodek, M. E. (2019). Reducing Pup Litter Size Alters Early Postnatal Calcium Homeostasis and Programs Adverse Adult Cardiovascular and Bone Health in Male Rats. Nutrients, 11(1), 118. https://doi.org/10.3390/nu11010118