Intestinal Anti-Inflammatory Effect of a Peptide Derived from Gastrointestinal Digestion of Buffalo (Bubalus bubalis) Mozzarella Cheese

 ,
, 
 ,
,  ,
,  ,
,  ,
,  , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Peptide Synthesis
2.2. Drugs and Reagents
2.3. Cell Culture
2.4. Animals
2.5. Cytotoxicity Studies
2.6. Alkaline Phosphatase (ALP) Activity
2.7. Western Blotting
2.8. Immunostaining and Confocal Microscopy
2.9. Permeability Assay on Caco-2 Cells
2.10. Induction of Experimental Colitis
2.11. Haematoxylin-Eosin Staining
2.12. Immuno-Fluorescence Microscopy
2.13. Intestinal Permeability Measurement
2.14. Induction of Intestinal Hypermotility and Upper Gastrointestinal Transit in Mice
2.15. Statistical Analysis
3. Results
3.1. In Vitro Study
3.1.1. MBCP Induces Cytotoxic Effects in Caco-2 Cells at Very High Concentrations
3.1.2. MBCP Stimulates Differentiation in Growing Caco-2 Cells
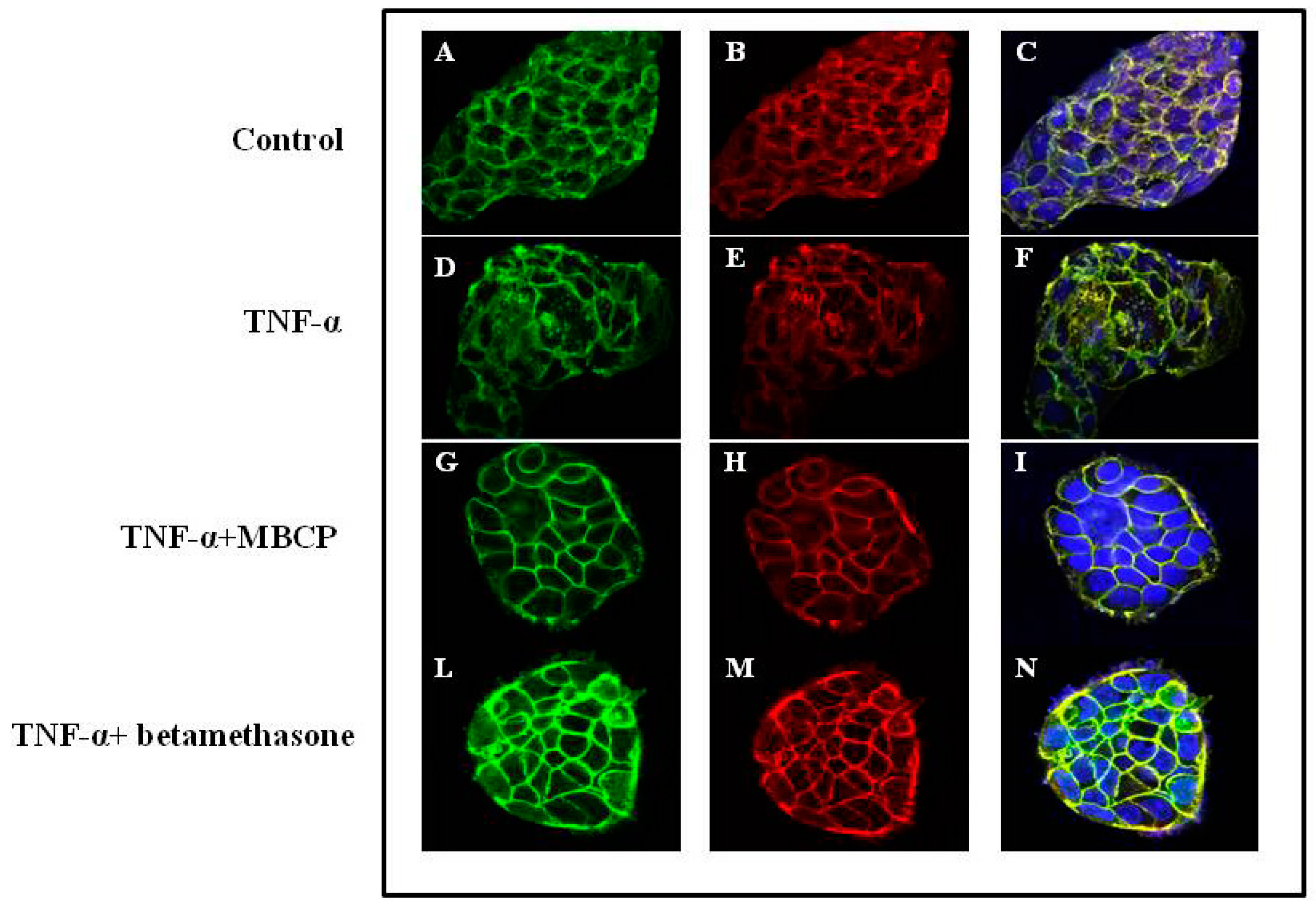
3.1.3. MBCP Modulates Adherent Junctions Formation in Control and TNF-α-Stimulated Caco-2 Cells
3.1.4. MBCP Counteracts the TNF-α Inflammatory Effect by Modulating the NF-κB Pathway
3.1.5. MBCP Reduces Atropine and Tubocurarine-Induced Adherens Junctions Disorganization
3.1.6. MBCP Inhibits the TNF-α-Increased Caco-2 Permeability
3.2. In Vivo Study
3.2.1. MBCP Reduces the Inflammation in the DNBS Model of Colitis
3.2.2. MBCP Reduces the Intestinal Permeability in Vivo
3.2.3. MBCP Counteracts the Accelerated Upper Gastrointestinal Transit Induced by Croton Oil
4. Discussion
4.1. MBPC Modulates the Differentiation and Permeability in Caco-2 Cells
4.2. MBPC Ameliorates Murine Colitis
4.3. MBPC Normalizes Inflammation-Induced Murine Intestinal Hypermotility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- Sairenji, T.; Collins, K.L.; Evans, D.V. An Update on Inflammatory Bowel Disease. Prim Care 2017, 44, 673–692. [Google Scholar] [CrossRef]
- Lucafò, M.; Franca, R.; Selvestrel, D.; Curci, D.; Pugnetti, L.; Decorti, G.; Stocco, G. Pharmacogenetics of treatments for inflammatory bowel disease. Expert Opin. Drug Metab. Toxicol. 2018, 14, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediat. Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef]
- Fasano, A.; Shea-Donohue, T. Mechanisms of disease: The role of intestinal barrier function in the pathogenesis of gastrointestinal autoimmune diseases. Nat. Clin. Pract. Gastroenterol. Hepatol. 2005, 2, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Coopman, P.; Djiane, A. Adherens Junction and E-Cadherin complex regulation by epithelial polarity. Cell. Mol. Life Sci. 2016, 73, 3535–3553. [Google Scholar] [CrossRef]
- Brüser, L.; Bogdan, S. Adherens Junctions on the Move-Membrane Trafficking of E-Cadherin. Cold Spring Harb. Perspect. Biol. 2017, 9, a029140. [Google Scholar] [CrossRef] [PubMed]
- Edelblum, K.L.; Turner, J.R. The tight junction in inflammatory disease: Communication breakdown. Curr. Opin. Pharmacol. 2009, 9, 715–720. [Google Scholar] [CrossRef]
- Utech, M.; Brüwer, M.; Nusrat, A. Tight junctions and cell-cell interactions. Methods Mol. Biol. 2006, 341, 185–195. [Google Scholar]
- De Simone, C.; Picariello, G.; Mamone, G.; Stiuso, P.; Dicitore, A.; Vanacore, D.; Chianese, L.; Addeo, F.; Ferranti, P. Characterisation and cytomodulatory properties of peptides from Mozzarella di Bufala Campana cheese whey. J. Pept. Sci. 2009, 15, 251–258. [Google Scholar] [CrossRef] [PubMed]
- De Simone, C.; Ferranti, P.; Picariello, G.; Scognamiglio, I.; Dicitore, A.; Addeo, F.; Chianese, L.; Stiuso, P. Peptides from water buffalo cheese whey induced senescence cell death via ceramide secretion in human colon adenocarcinoma cell line. Mol. Nutr. Food Res. 2011, 55, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, D.P.; Mohapatra, S.; Misra, S.; Sahu, P.S. Milk derived bioactive peptides and their impact on human health—A review. Saudi J. Biol. Sci. 2016, 23, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Chanu, K.V.; Thakuria, D.; Kumar, S. Antimicrobial peptides of buffalo and their role in host defenses. Vet. World 2018, 11, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basilicata, M.G.; Pepe, G.; Adesso, S.; Ostacolo, C.; Sala, M.; Sommella, E.; Scala, M.C.; Messore, A.; Autore, G.; Marzocco, S.; et al. Antioxidant Properties of Buffalo-Milk Dairy Products: A β-Lg Peptide Released after Gastrointestinal Digestion of Buffalo Ricotta Cheese Reduces Oxidative Stress in Intestinal Epithelial Cells. Int. J. Mol. Sci. 2018, 19, 1955. [Google Scholar] [CrossRef] [PubMed]
- Tenore, G.C.; Ritieni, A.; Campiglia, P.; Stiuso, P.; Di Maro, S.; Sommella, E.; Pepe, G.; D’Urso, E.; Novellino, E. Antioxidant peptides from “Mozzarella di Bufala Campana DOP” after simulated gastrointestinal digestion: In vitro intestinal protection, bioavailability, and anti-haemolytic capacity. J. Funct. Foods 2015, 15, 365–375. [Google Scholar]
- Pagano, E.; Capasso, R.; Piscitelli, F.; Romano, B.; Parisi, O.A.; Finizio, S.; Lauritano, A.; Marzo, V.D.; Izzo, A.A.; Borrelli, F. An Orally Active Cannabis Extract with High Content in Cannabidiol attenuates Chemically-induced Intestinal Inflammation and Hypermotility in the Mouse. Front. Pharmacol. 2016, 7, 341. [Google Scholar] [CrossRef]
- Venditti, M.; Fasano, C.; Santillo, A.; Aniello, F.; Minucci, S. First evidence of DAAM1 localization in mouse seminal vesicles and its possible involvement during regulated exocytosis. C. R. Biol. 2018, 341, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Capasso, R.; Orlando, P.; Pagano, E.; Aveta, T.; Buono, L.; Borrelli, F.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide normalizes intestinal motility in a model of post-inflammatory accelerated transit: Involvement of CB1 receptors and TRPV1 channels. Br. J. Pharmacol. 2014, 171, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Pol, O.; Puig, M.M. Reversal of tolerance to the antitransit effects of morphine during acute intestinal inflammation in mice. Br. J. Pharmacol. 1997, 122, 1216–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, H.; Erickson, R.H.; Gum, J.R.; Yoshioka, M.; Gum, E.; Kim, Y.S. Biosynthesis of alkaline phosphatase during differentiation of the human colon cancer cell line Caco-2. Gastroenterology 1990, 98, 1199–1207. [Google Scholar] [CrossRef]
- Pelissier-Rota, M.; Lainé, M.; Ducarouge, B.; Bonaz, B.; Jacquier-Sarlin, M. Role of cholinergic receptors in colorectal cancer: Potential therapeutic implications of vagus nerve stimulation? J. Cancer Ther. 2013, 4, 1116–1131. [Google Scholar] [CrossRef]
- Lesko, S.; Wessler, I.; Gäbel, G.; Petto, C.; Pfannkuche, H. Cholinergic modulation of epithelial integrity in the proximal colon of pigs. Cells Tissues Organs 2013, 197, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.R.; Uwada, J.; Yazawa, T.; Islam, M.T.; Krug, S.M.; Fromm, M.; Karaki, S.; Suzuki, Y.; Kuwahara, A.; Yoshiki, H.; et al. Activation of muscarinic cholinoceptor ameliorates tumor necrosis factor-α-induced barrier dysfunction in intestinal epithelial cells. FEBS Lett. 2015, 589, 3640–3647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Cao, L.; Liu, B.; McCaig, C.D.; Pu, J. The transition from proliferation to differentiation in colorectal cancer is regulated by the calcium activated chloride channel A1. PLoS ONE 2013, 8, e60861. [Google Scholar] [CrossRef]
- Tanoue, T.; Nishitani, Y.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. In vitro model to estimate gut inflammation using co-cultured Caco-2 and RAW264.7 cells. Biochem. Biophys. Res. Commun. 2008, 374, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Tatiya-Aphiradee, N.; Chatuphonprasert, W.; Jarukamjorn, K. Immune response and inflammatory pathway of ulcerative colitis. J. Basic Clin. Physiol. Pharmacol. 2018, 30, 1–10. [Google Scholar] [CrossRef]
- Buhrke, T.; Lengler, I.; Lampen, A. Analysis of proteomic changes induced upon cellular differentiation of the human intestinal cell line Caco-2. Dev. Growth Differ. 2011, 53, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Bilski, J.; Mazur-Bialy, A.; Wojcik, D.; Zahradnik-Bilska, J.; Brzozowski, B.; Magierowski, M.; Mach, T.; Magierowska, K.; Brzozowski, T. The Role of Intestinal Alkaline Phosphatase in Inflammatory Disorders of Gastrointestinal Tract. Mediat. Inflamm. 2017, 2017, 9074601. [Google Scholar] [CrossRef]
- Yi, J.Y.; Jung, Y.J.; Choi, S.S.; Chung, E. TNF-alpha Downregulates E-cadherin and Sensitizes Response to γ-irradiation in Caco-2 Cells. Cancer Res. Treat. 2009, 41, 164–170. [Google Scholar] [CrossRef]
- Christian, F.; Smith, E.L.; Carmody, R.J. The Regulation of NF-κB Subunits by Phosphorylation. Cells 2016, 5, 12. [Google Scholar] [CrossRef]
- Rioux, N.; Castonguay, A. The induction of cyclooxygenase-1 by a tobacco carcinogen in U937 human macrophages is correlated to the activation of NF-kappaB. Carcinogenesis 2000, 21, 1745–1751. [Google Scholar] [CrossRef] [PubMed]
- McGilligan, V.E.; Wallace, J.M.; Heavey, P.M.; Ridley, D.L.; Rowland, I.R. The effect of nicotine in vitro on the integrity of tight junctions in Caco-2 cell monolayers. Food. Chem. Toxicol. 2007, 45, 1593–1598. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Fasolino, I.; Romano, B.; Capasso, R.; Maiello, F.; Coppola, D.; Orlando, P.; Battista, G.; Pagano, E.; Di Marzo, V.; et al. Beneficial effect of the non-psychotropic plant cannabinoid cannabigerol on experimental inflammatory bowel disease. Biochem. Pharmacol. 2013, 85, 1306–1316. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, F.; Romano, B.; Petrosino, S.; Pagano, E.; Capasso, R.; Coppola, D.; Battista, G.; Orlando, P.; Di Marzo, V.; Izzo, A.A. Palmitoylethanolamide, a naturally occurring lipid, is an orally effective intestinal anti-inflammatory agent. Br. J. Pharmacol. 2015, 172, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Leong, R.W.; Wasinger, V.C.; Ip, M.; Yang, M.; Phan, T.G. Impaired Intestinal Permeability Contributes to Ongoing Bowel Symptoms in Patients with Inflammatory Bowel Disease and Mucosal Healing. Gastroenterology 2017, 153, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Brierley, S.M.; Linden, D.R. Neuroplasticity and dysfunction after gastrointestinal inflammation. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 611–627. [Google Scholar] [CrossRef]
- Corsetti, M.; Tack, J. Naloxegol, a new drug for the treatment of opioid-induced constipation. Expert Opin. Pharmacother. 2015, 16, 399–406. [Google Scholar] [CrossRef]
- Rao, S.S.; Rattanakovit, K.; Patcharatrakul, T. Diagnosis and management of chronic constipation in adults. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 295–305. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tenore, G.C.; Pagano, E.; Lama, S.; Vanacore, D.; Di Maro, S.; Maisto, M.; Capasso, R.; Merlino, F.; Borrelli, F.; Stiuso, P.; et al. Intestinal Anti-Inflammatory Effect of a Peptide Derived from Gastrointestinal Digestion of Buffalo (Bubalus bubalis) Mozzarella Cheese. Nutrients 2019, 11, 610. https://doi.org/10.3390/nu11030610
Tenore GC, Pagano E, Lama S, Vanacore D, Di Maro S, Maisto M, Capasso R, Merlino F, Borrelli F, Stiuso P, et al. Intestinal Anti-Inflammatory Effect of a Peptide Derived from Gastrointestinal Digestion of Buffalo (Bubalus bubalis) Mozzarella Cheese. Nutrients. 2019; 11(3):610. https://doi.org/10.3390/nu11030610
Chicago/Turabian StyleTenore, Gian Carlo, Ester Pagano, Stefania Lama, Daniela Vanacore, Salvatore Di Maro, Maria Maisto, Raffaele Capasso, Francesco Merlino, Francesca Borrelli, Paola Stiuso, and et al. 2019. "Intestinal Anti-Inflammatory Effect of a Peptide Derived from Gastrointestinal Digestion of Buffalo (Bubalus bubalis) Mozzarella Cheese" Nutrients 11, no. 3: 610. https://doi.org/10.3390/nu11030610
APA StyleTenore, G. C., Pagano, E., Lama, S., Vanacore, D., Di Maro, S., Maisto, M., Capasso, R., Merlino, F., Borrelli, F., Stiuso, P., & Novellino, E. (2019). Intestinal Anti-Inflammatory Effect of a Peptide Derived from Gastrointestinal Digestion of Buffalo (Bubalus bubalis) Mozzarella Cheese. Nutrients, 11(3), 610. https://doi.org/10.3390/nu11030610