Differential Stimulation of Post-Exercise Myofibrillar Protein Synthesis in Humans Following Isonitrogenous, Isocaloric Pre-Exercise Feeding

 ,
,  , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Participants
2.3. Study Conduct
2.4. Resistance Exercise
2.5. Body Water Enrichment
2.6. Protein Bound Alanine Enrichment
2.7. Calculation of Fractional Synthetic Rate
2.8. Postprandial Aminoacidemia
2.9. Statistical Analysis
3. Results
3.1. Participants and Resistance Training Performance
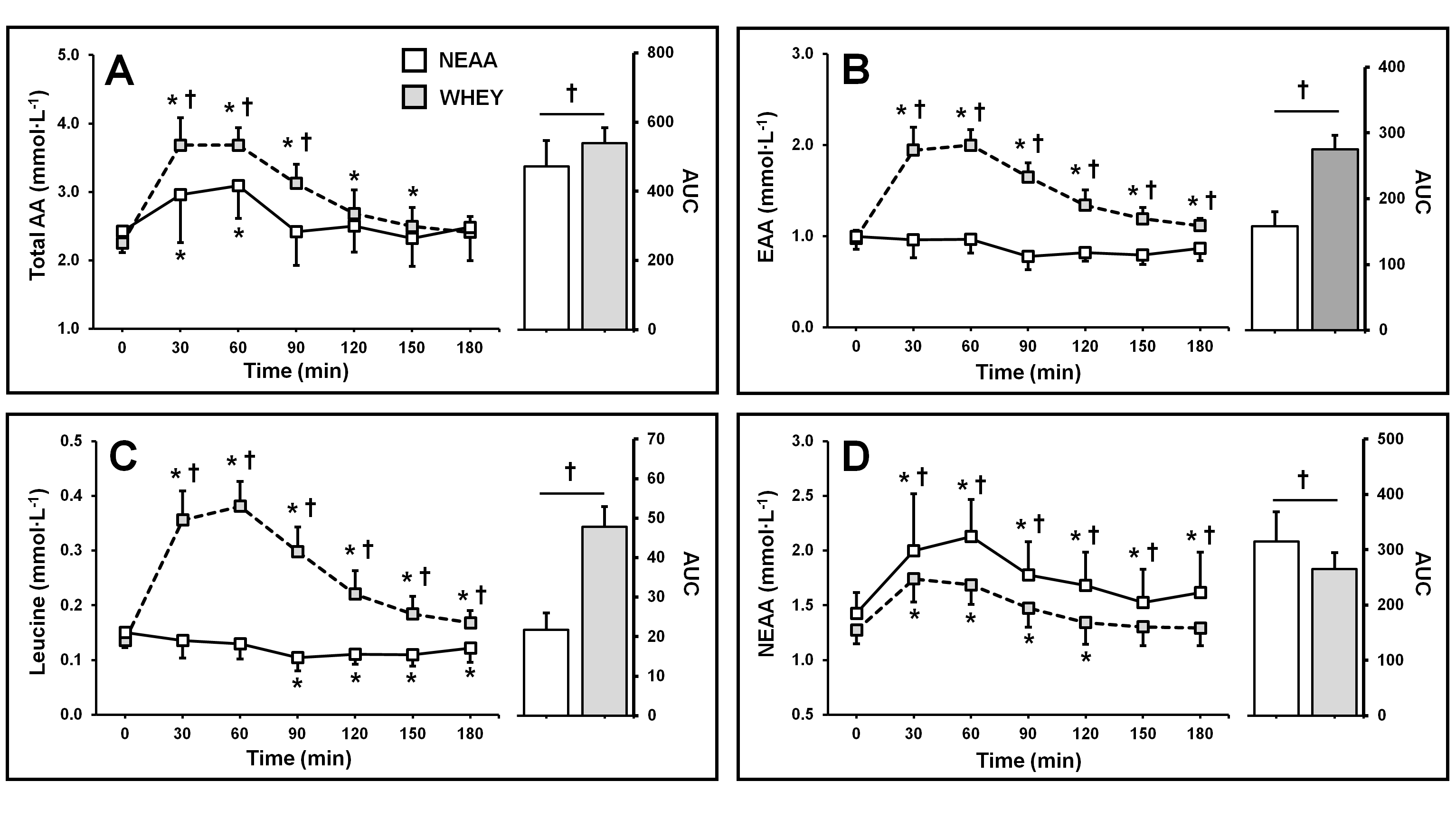
3.2. Aminoacidemia
3.2.1. Plasma Amino Acid Concentration
3.2.2. Plasma Essential Amino Acid Concentration
3.2.3. Plasma Leucine Concentration
3.2.4. Plasma Nonessential Amino Acid Concentration
3.3. Body Water Enrichment
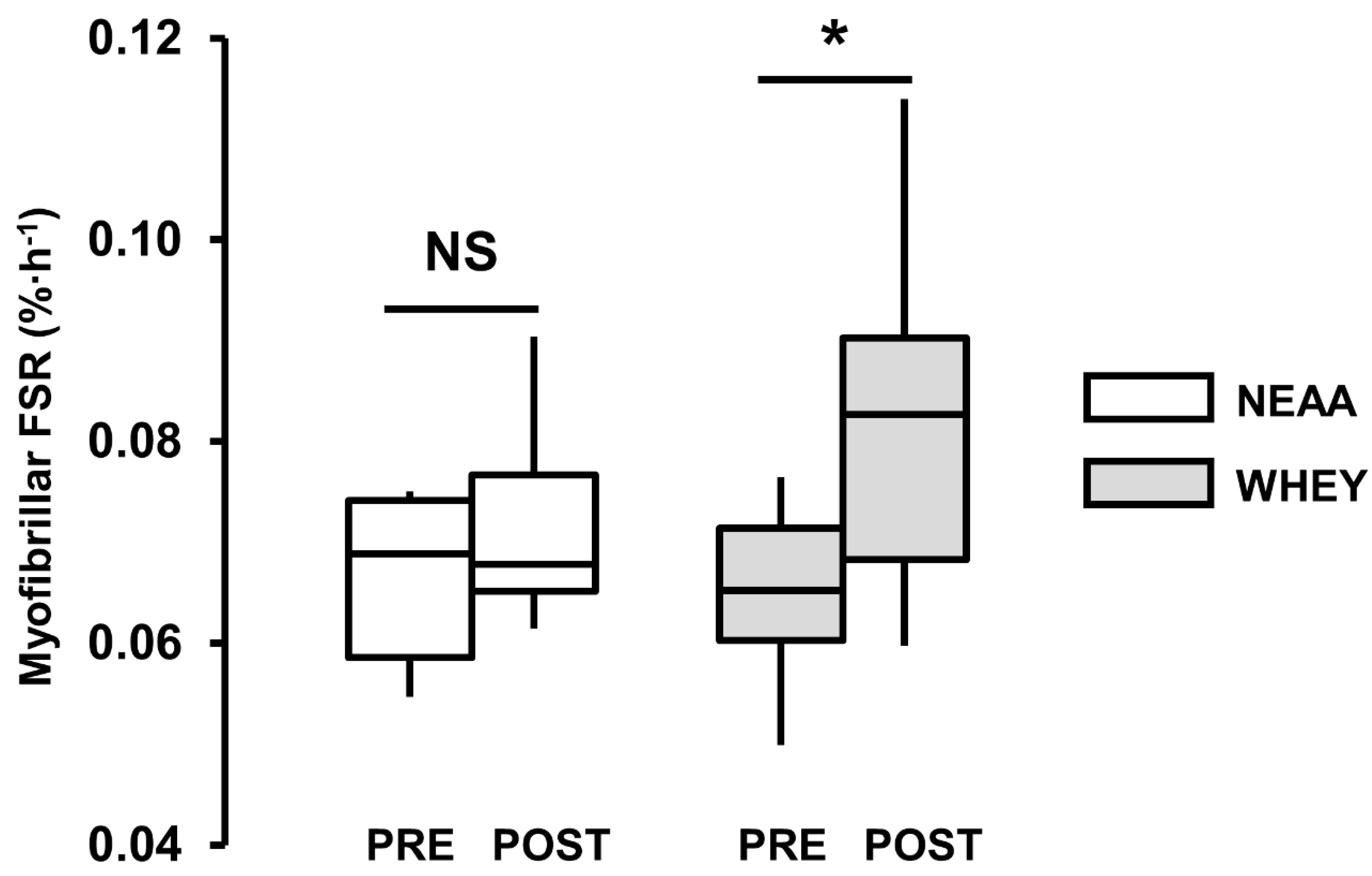
3.4. Myofibrillar Protein Synthesis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
| NEAA (n = 8) | WPC (n = 7) | WPH (n = 7) | |
| myoFSR PRE (%·h−1) | 0.066 (0.008) | 0.068 (0.006) | 0.063 (0.008) |
| myoFSR POST (%·h−1) | 0.072 (0.010) | 0.084 (0.017) | 0.079 (0.016) |
| % change | 10 (−5, 25) | 27 (2, 51) * | 30 (5, 55) * |
References
- Biolo, G.; Maggi, S.P.; Williams, B.D.; Tipton, K.D.; Wolfe, R.R. Increased rates of muscle protein turnover and amino acid transport after resistance exercise in humans. Am. J. Physiol. 1995, 268, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Bohe, J.; Low, A.; Wolfe, R.R.; Rennie, M.J. Human muscle protein synthesis is modulated by extracellular, not intramuscular amino acid availability: A dose-response study. J. Physiol. 2003, 552, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Jeong, S.J.; Park, M.C.; Kim, G.; Kwon, N.H.; Kim, H.K.; Ha, S.H.; Ryu, S.H.; Kim, S. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway. Cell 2012, 149, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Knockenhauer, K.E.; Wolfson, R.L.; Chantranupong, L.; Pacold, M.E.; Wang, T.; Schwartz, T.U.; Sabatini, D.M. Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway. Science 2016, 351, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2014, 99, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Borsheim, E.; Tipton, K.D.; Wolf, S.E.; Wolfe, R.R. Essential amino acids and muscle protein recovery from resistance exercise. Am. J. Physiol. Endocrinol. Metab. 2002, 283, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.; Gurkin, B.; Matin, S.; Wolfe, R. Nonessential amino acids are not necessary to stimulate net muscle protein synthesis in healthy volunteers. J. Nutr. Biochem. 1999, 10, 89–95. [Google Scholar] [CrossRef]
- Turck, D.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Knutsen, H.K.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Guidance on the scientific requirements for health claims related to muscle function and physical performance. EFSA J. 2018, 16, e05434. [Google Scholar] [CrossRef]
- Phillips, S.M. The impact of protein quality on the promotion of resistance exercise-induced changes in muscle mass. Nutr. Metab. 2016, 13, 64. [Google Scholar] [CrossRef]
- Moore, D.R.; Tang, J.E.; Burd, N.A.; Rerecich, T.; Tarnopolsky, M.A.; Phillips, S.M. Differential stimulation of myofibrillar and sarcoplasmic protein synthesis with protein ingestion at rest and after resistance exercise. J. Physiol. 2009, 587, 897–904. [Google Scholar] [CrossRef]
- Reitelseder, S.; Agergaard, J.; Doessing, S.; Helmark, I.C.; Lund, P.; Kristensen, N.B.; Frystyk, J.; Flyvbjerg, A.; Schjerling, P.; van Hall, G. Whey and casein labeled with L-[1-13C] leucine and muscle protein synthesis: Effect of resistance exercise and protein ingestion. Am. J. Physiol. Endocrinol. Metab. 2010, 300, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Elliott, T.A.; Cree, M.G.; Wolf, S.E.; Sanford, A.P.; Wolfe, R.R. Ingestion of casein and whey proteins result in muscle anabolism after resistance exercise. Med. Sci. Sports. Exerc. 2004, 36, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Koopman, R.; Wagenmakers, A.J.; Manders, R.J.; Zorenc, A.H.; Senden, J.M.; Gorselink, M.; Keizer, H.A.; van Loon, L.J. Combined ingestion of protein and free leucine with carbohydrate increases postexercise muscle protein synthesis in vivo in male subjects. Am. J. Physiol. Endocrinol. Metab. 2005, 288, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.E.; Manolakos, J.J.; Kujbida, G.W.; Lysecki, P.J.; Moore, D.R.; Phillips, S.M. Minimal whey protein with carbohydrate stimulates muscle protein synthesis following resistance exercise in trained young men. Appl. Physiol. Nutr. Metab. 2007, 32, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- West, D.; Abou Sawan, S.; Mazzulla, M.; Williamson, E.; Moore, D. Whey protein supplementation enhances whole body protein metabolism and performance recovery after resistance exercise: A double-blind crossover study. Nutrients 2017, 9, 735. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.E.; Moore, D.R.; Kujbida, G.W.; Tarnopolsky, M.A.; Phillips, S.M. Ingestion of whey hydrolysate, casein, or soy protein isolate: Effects on mixed muscle protein synthesis at rest and following resistance exercise in young men. J. Appl. Physiol. 2009, 107, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.B.; Tarnopolsky, M.A.; Macdonald, M.J.; Macdonald, J.R.; Armstrong, D.; Phillips, S.M. Consumption of fluid skim milk promotes greater muscle protein synthesis after resistance exercise than does consumption of an isonitrogenous and isoenergetic soy-protein beverage. Am. J. Clin. Nutr. 2007, 85, 1031–1040. [Google Scholar] [CrossRef]
- Carson, B.; Davies, R.; Bass, J.; Norton, C.; Patel, B.; Amigo-Benavent, M.; Murphy, S.; Kiely, P.; Jakeman, P. PL-033 A translational model of muscle protein synthetic bioactivity in vitro, ex vivo and in vivo. Exerc. Biochem. Rev. 2018, 1. [Google Scholar] [CrossRef]
- Davies, R.W.; Carson, B.P.; Bass, J.J.; Holohan, S.; Jakeman, P.M. Acute reduction of lower-body contractile function following a microbiopsy of m. vastus lateralis. Scand. J. Med. Sci. Sports 2018, 28, 2638–2642. [Google Scholar] [CrossRef]
- Davies, R.W.; Carson, B.P.; Jakeman, P.M. Sex Differences in the Temporal Recovery of Neuromuscular Function Following Resistance Training in Resistance Trained Men and Women 18 to 35 Years. Front. Physiol. 2018, 9, 1480. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, D.J.; Franchi, M.V.; Brook, M.S.; Narici, M.V.; Williams, J.P.; Mitchell, W.K.; Szewczyk, N.J.; Greenhaff, P.L.; Atherton, P.J.; Smith, K. A validation of the application of (D2O) stable isotope tracer techniques for monitoring day-to-day changes in muscle protein subfraction synthesis in humans. Am. J. Physiol. Endocrinol. Metab. 2014, 306, 571–579; [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.J.; Cegielski, J.; Phillips, B.E.; Boereboom, C.; Lund, J.N.; Atherton, P.J.; Smith, K. Internal comparison between deuterium oxide (D2O) and L-[ring-13C6] phenylalanine for acute measurement of muscle protein synthesis in humans. Physiol. Rep. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Holwerda, A.M.; Paulussen, K.J.M.; Overkamp, M.; Smeets, J.S.J.; Gijsen, A.P.; Goessens, J.P.B.; Verdijk, L.B.; van Loon, L.J.C. Daily resistance-type exercise stimulates muscle protein synthesis in vivo in young men. J. Appl. Physiol. 2018, 124, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Power-Grant, O.; McCormack, W.G.; Ramia De Cap, M.; Amigo-Benavent, M.; Fitzgerald, R.J.; Jakeman, P. Evaluation of the antioxidant capacity of a milk protein matrix in vitro and in vivo in women aged 50–70 years. Int. J. Food Sci. Nutr. 2016, 67, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 863. [Google Scholar] [CrossRef] [PubMed]
- Reidy, P.T.; Rasmussen, B.B. Role of Ingested Amino Acids and Protein in the Promotion of Resistance Exercise-Induced Muscle Protein Anabolism. J. Nutr. 2016, 146, 155–183. [Google Scholar] [CrossRef]
- Smith, G.I.; Patterson, B.W.; Mittendorfer, B. Human muscle protein turnover--why is it so variable? J. Appl. Physiol. 2011, 110, 480–491. [Google Scholar] [CrossRef]
- Atherton, P.J.; Miller, B.F.; Burd, N.A.; Macnaughton, L.S.; Murton, A.J.; Camera, D.M.; Pancheva, A.V.; Wang, X.; Vagula, M.C.; Phillips, B.E.; et al. Commentaries on Viewpoint: What is the relationship between acute measure of muscle protein synthesis and changes in muscle mass? J. Appl. Physiol. 2015, 118, 498–503. [Google Scholar] [CrossRef]
- Mitchell, C.J.; Churchward-Venne, T.A.; Cameron-Smith, D.; Phillips, S.M. What is the relationship between the acute muscle protein synthesis response and changes in muscle mass? J. Appl. Physiol. 2015, 118, 495–497. [Google Scholar] [CrossRef] [Green Version]
- Brook, M.S.; Wilkinson, D.J.; Mitchell, W.K.; Lund, J.N.; Szewczyk, N.J.; Greenhaff, P.L.; Smith, K.; Atherton, P.J. Skeletal muscle hypertrophy adaptations predominate in the early stages of resistance exercise training, matching deuterium oxide-derived measures of muscle protein synthesis and mechanistic target of rapamycin complex 1 signaling. FASEB J. 2015, 29, 4485–4496. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| WHEY | NEAA | |||
|---|---|---|---|---|
| mg·kg−1 | % | mg·kg1 | % | |
| Alanine | 17 | 5 | 33 | 10 |
| Arginine | 8 | 2 | 0 | 0 |
| Aspartic acid | 35 | 11 | 41 | 12 |
| Cysteine | 8 | 2 | 0 | 0 |
| Glutamic acid | 56 | 17 | 120 | 36 |
| Glycine | 6 | 2 | 13 | 4 |
| Histidine | 6 | 2 | 0 | 0 |
| Isoleucine | 20 | 6 | 0 | 0 |
| Leucine | 34 | 10 | 0 | 0 |
| Lysine | 30 | 9 | 0 | 0 |
| Methionine | 7 | 2 | 0 | 0 |
| Phenylalanine | 10 | 3 | 0 | 0 |
| Proline | 19 | 6 | 53 | 16 |
| Serine | 17 | 5 | 45 | 14 |
| Threonine | 23 | 7 | 0 | 0 |
| Tryptophan | 7 | 2 | 0 | 0 |
| Tyrosine | 9 | 3 | 25 | 8 |
| Valine | 19 | 6 | 0 | 0 |
| EAA | 0.16 | 48 | 0 | 0 |
| NEAA | 0.17 | 52 | 0.33 | 100 |
| Total | 0.33 | 0.33 |
| NEAA (n = 8) | WHEY (n = 14) | p-Value | |
|---|---|---|---|
| Age (y) | 23 (5) | 23 (4) | 0.666 |
| Stature (m) | 1.77 (0.04) | 1.82 (0.07) | 0.120 |
| Body mass (kg) | 77 (17) | 81 (10) | 0.455 |
| 1RM (kg·kg−1) | 1.5 (0.3) | 1.5 (0.2) | 0.985 |
| RET experience (y) | 2.6 (1.5) | 2.4 (1.1) | 0.602 |
| RET performance (sets) | 8.7 (1.5) | 7.6 (1.8) | 0.287 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, R.W.; Bass, J.J.; Carson, B.P.; Norton, C.; Kozior, M.; Amigo-Benavent, M.; Wilkinson, D.J.; Brook, M.S.; Atherton, P.J.; Smith, K.; et al. Differential Stimulation of Post-Exercise Myofibrillar Protein Synthesis in Humans Following Isonitrogenous, Isocaloric Pre-Exercise Feeding. Nutrients 2019, 11, 1657. https://doi.org/10.3390/nu11071657
Davies RW, Bass JJ, Carson BP, Norton C, Kozior M, Amigo-Benavent M, Wilkinson DJ, Brook MS, Atherton PJ, Smith K, et al. Differential Stimulation of Post-Exercise Myofibrillar Protein Synthesis in Humans Following Isonitrogenous, Isocaloric Pre-Exercise Feeding. Nutrients. 2019; 11(7):1657. https://doi.org/10.3390/nu11071657
Chicago/Turabian StyleDavies, Robert W., Joseph J. Bass, Brian P. Carson, Catherine Norton, Marta Kozior, Miryam Amigo-Benavent, Daniel J. Wilkinson, Matthew S. Brook, Philip J. Atherton, Kenneth Smith, and et al. 2019. "Differential Stimulation of Post-Exercise Myofibrillar Protein Synthesis in Humans Following Isonitrogenous, Isocaloric Pre-Exercise Feeding" Nutrients 11, no. 7: 1657. https://doi.org/10.3390/nu11071657