Mustard Leaf Extract Suppresses Psychological Stress in Chronic Restraint Stress-Subjected Mice by Regulation of Stress Hormone, Neurotransmitters, and Apoptosis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of a Mustard Leaf (ML) Extract
2.3. Animals and Treatments
2.4. Chronic Restraint Stress (CRS) Procedure in Mice
2.5. Stress Hormone Estimation
2.6. Neurotransmitter Estimation
2.7. Real-Time PCR Analysis
2.8. Western Blot Analysis
2.9. Inflammatory Cytokine Estimation
2.10. Statistical Analyses
3. Results
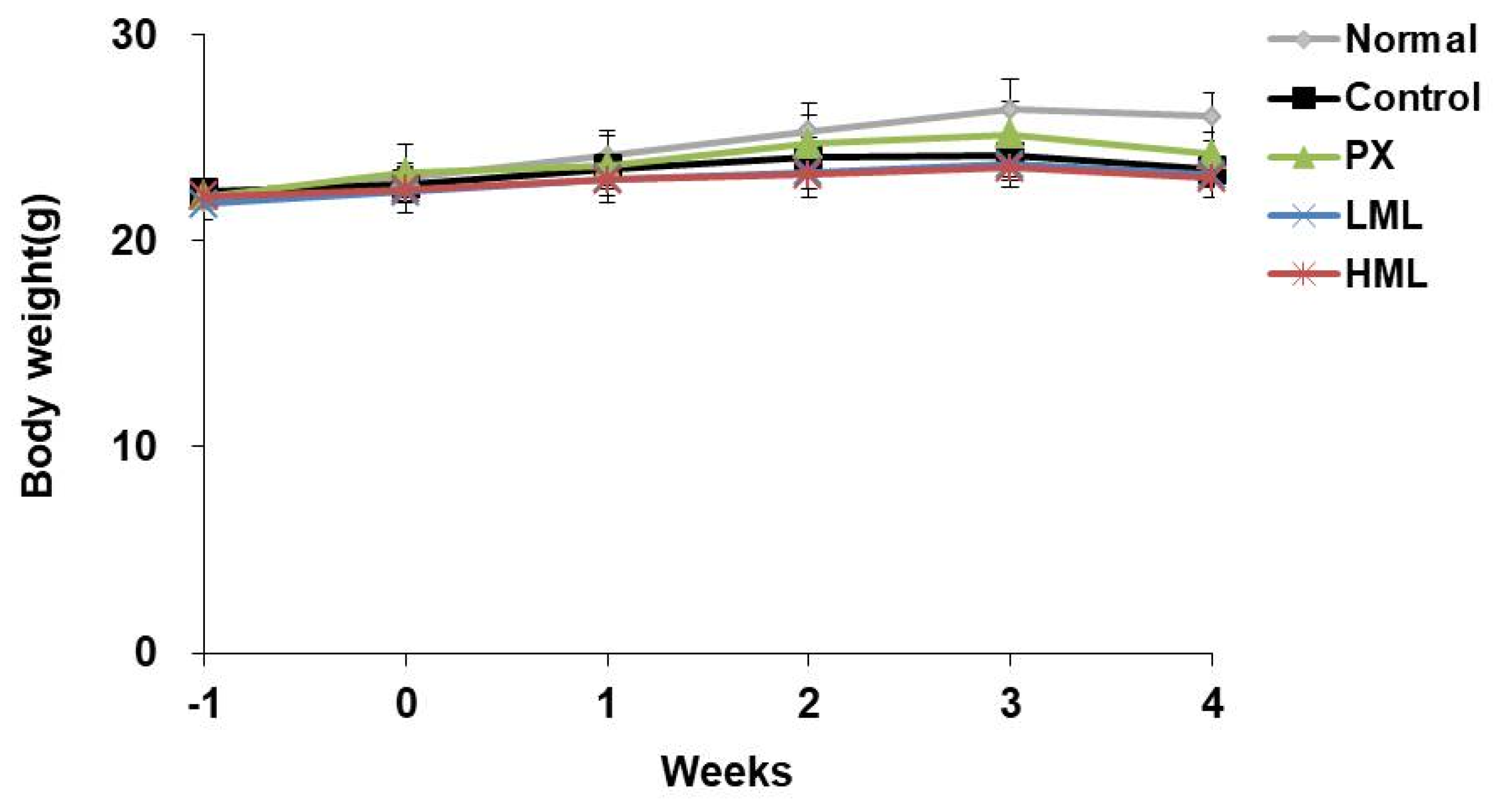
3.1. Effect of CRS and ML Extract on Body Weight
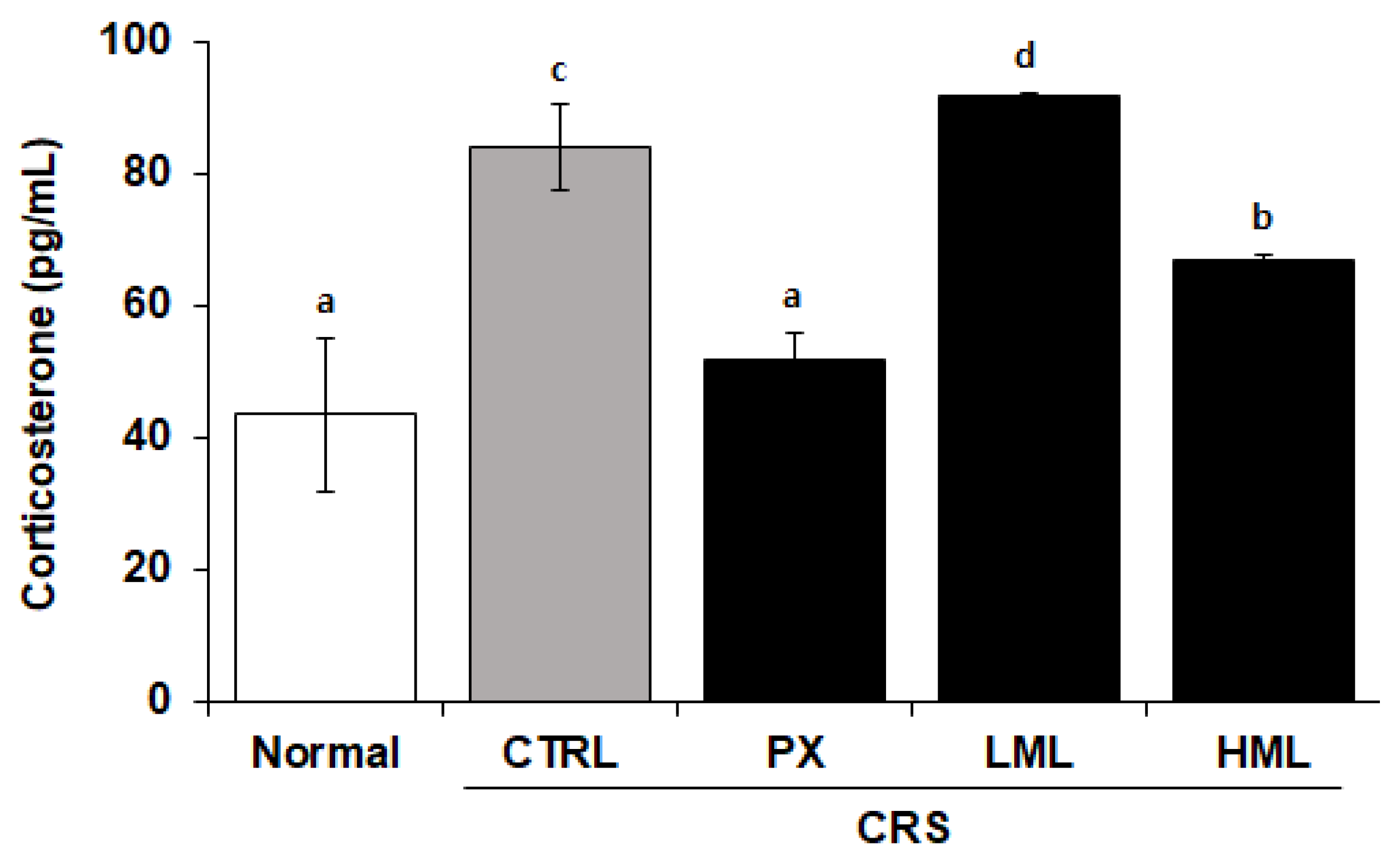
3.2. Effect of the ML Extract on Corticosterone Levels in CRS-Subjected Mice
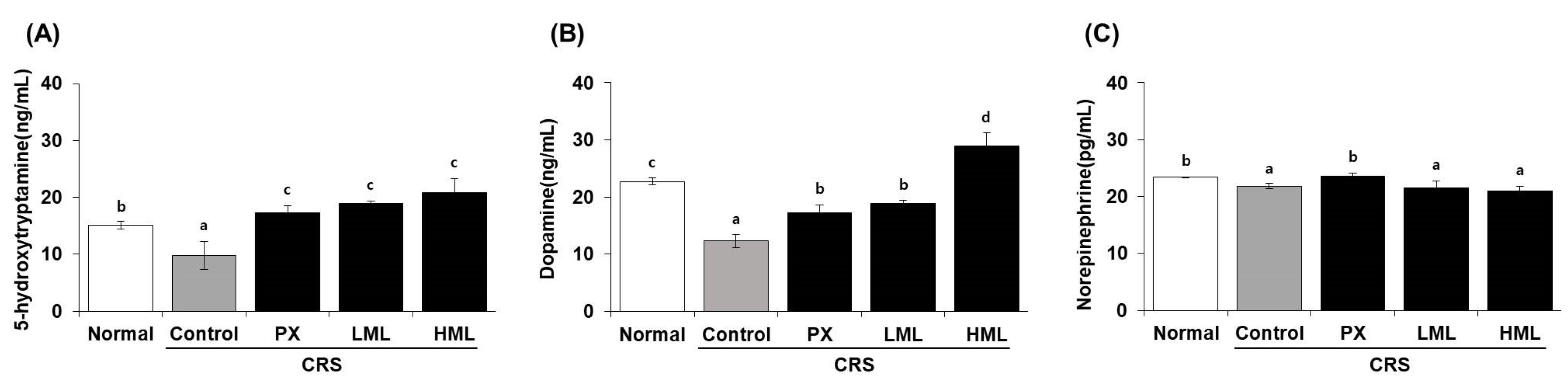
3.3. Effect of the ML Extract on Neurotransmitter Levels in CRS-Subjecetd Mice
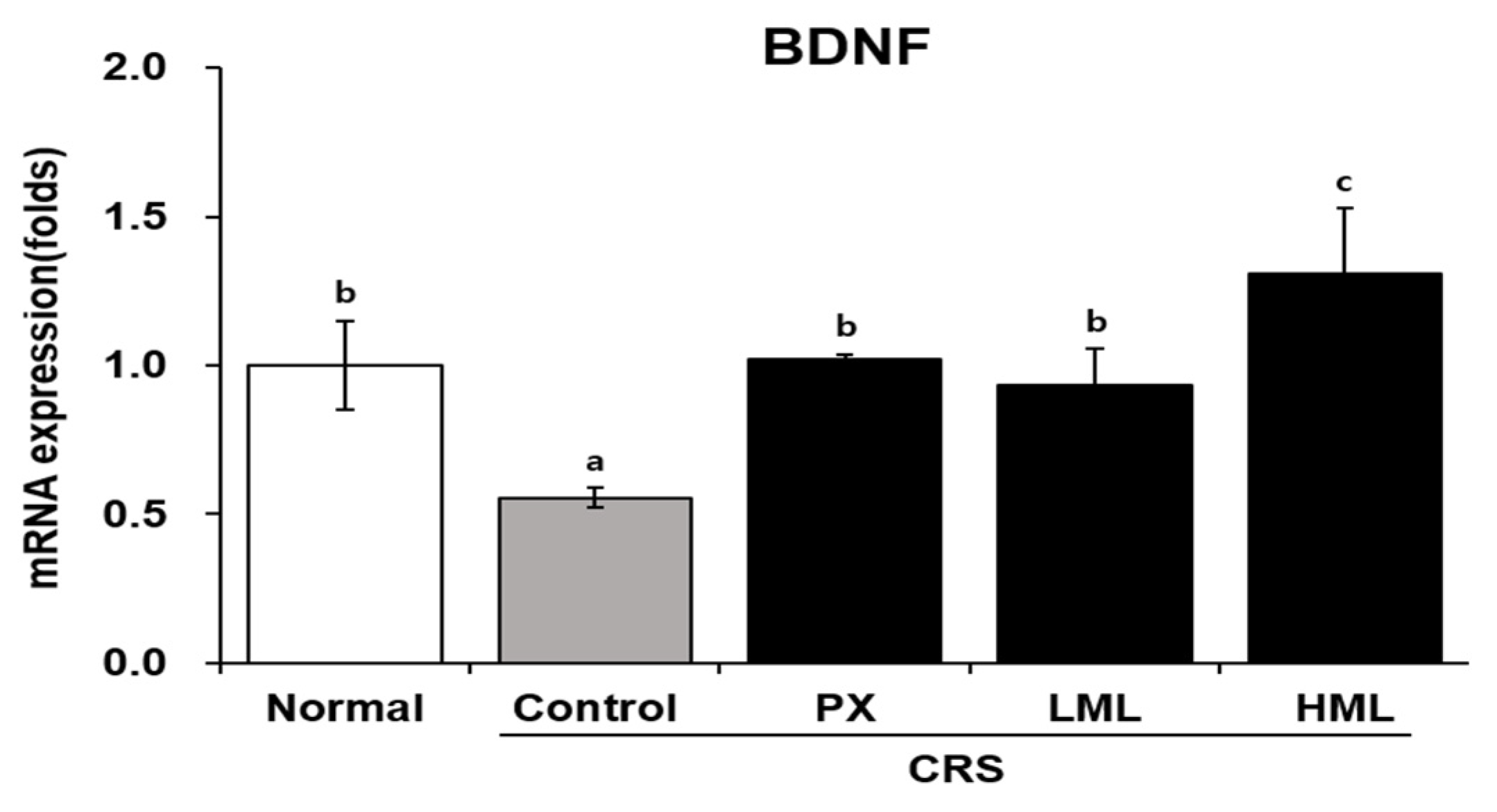
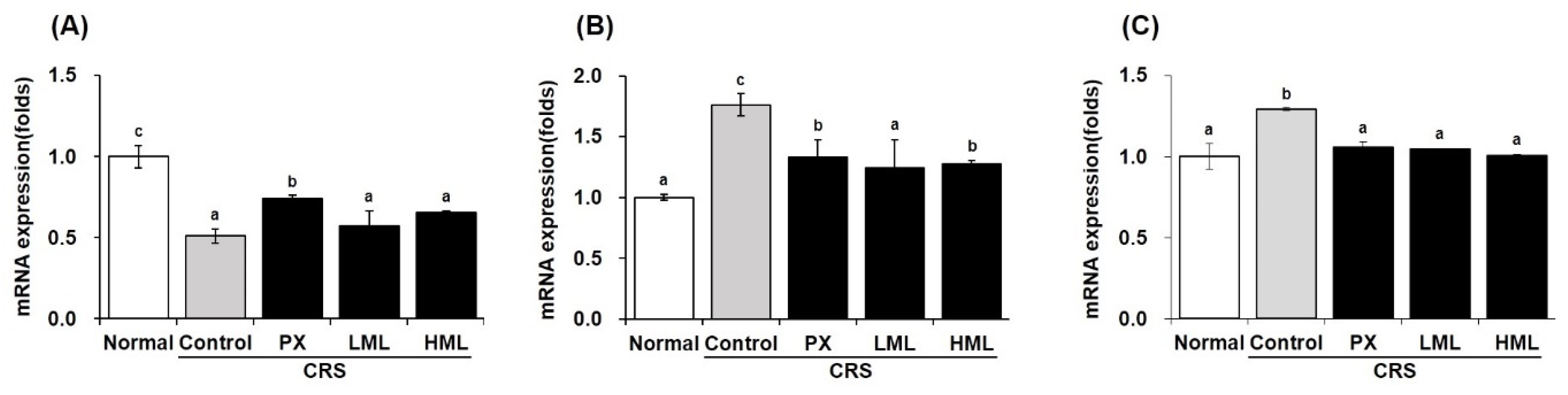
3.4. Effect of the ML Extract on BDNF mRNA Expression in CRS-Subjetced Mice
3.5. Effect of the ML Extract on Apoptosis-Related mRNA Expression in CRS-Subjected Mice
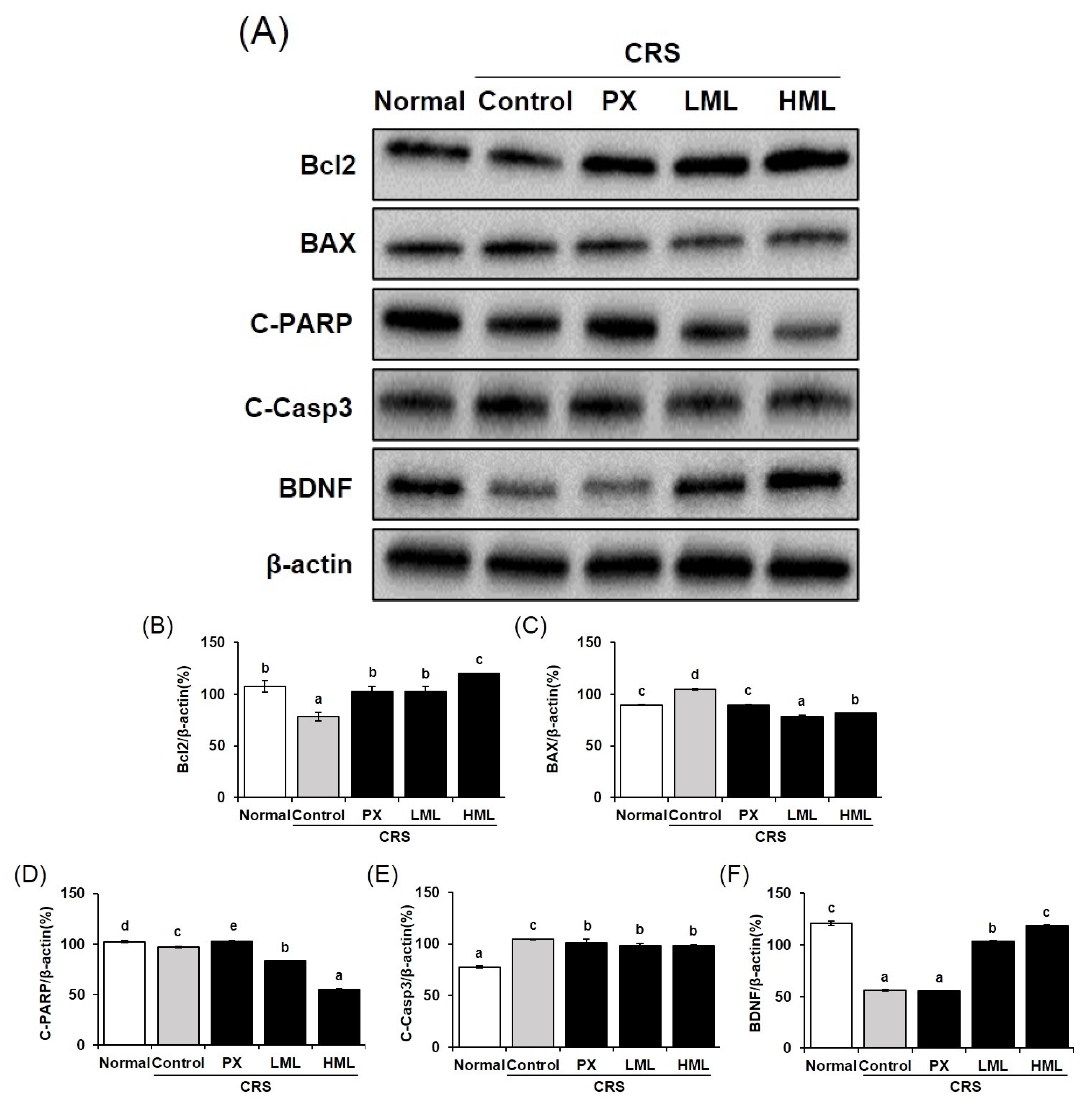
3.6. Effect of the ML Extract on Apoptosis-Related Protein Expression in CRS-Induced Mice
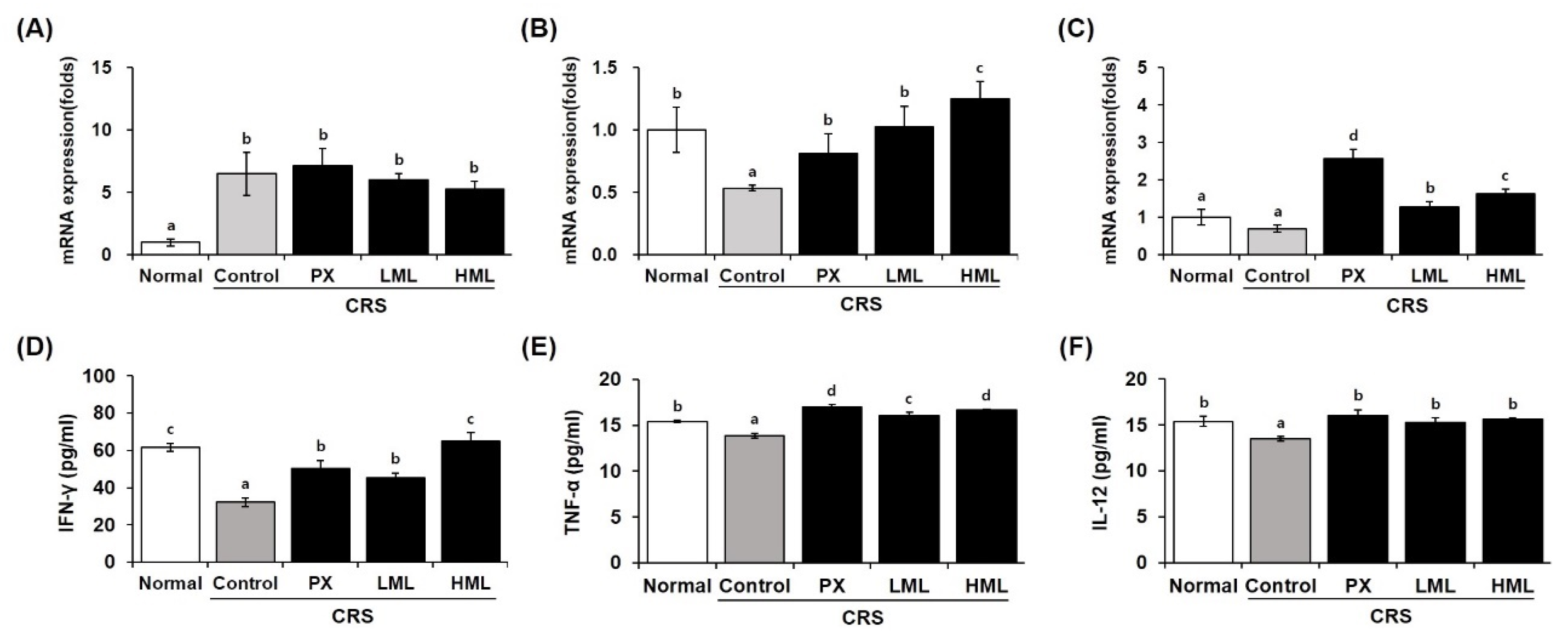
3.7. Effect of the ML Extract on Anti- and Pro-Inflammatory Cytokine mRNA Expression in CRS-Subjected Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chiba, S.; Numakawa, T.; Ninomiya, M.; Richards, M.C.; Wakabayashi, C.; Kunugi, H. Chronic restraint stress causes anxiety- and depression-like behaviors: Downregulates glucocorticoid receptor expression, and attenuates glutamate release induced by brain-derived neurotrophic factor in the prefrontal cortex. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.N.; Krigbaum, A.M.; Hanna, J.J.; Conrad, C.D. Sex differences and phase of light cycle modify chronic stress effects on anxiety and depressive-like behavior. Behav. Brain Res. 2011, 222, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Woolley, C.S.; Gould, E.; McEwen, B.S. Exposure to excess glucocorticoids alters dendritic morphology of adult hippocampal pyramidal neurons. Brain Res. 1990, 531, 225–231. [Google Scholar] [CrossRef]
- Watanabe, Y.; Gould, E.; McEwen, B.S. Stress induces atrophy of apical dendrites of hippocampal CA3 pyramidal neurons. Brain Res. 1992, 588, 341–345. [Google Scholar] [CrossRef]
- McAllister, A.K.; Katz, L.C.; Lo, D.C. Opposing roles for endogenous BDNF and NT-3 in regulating cortical dendritic growth. Neuron 1997, 18, 767–778. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, P.M.; Borghesani, P.R.; Levy, R.L.; Pomeroy, S.L.; Segal, R.A. Abnormal cerebellar development and foliation in BDNF-/- mice reveals a role for neurotrophins in CNS patterning. Neuron 1997, 19, 269–281. [Google Scholar] [CrossRef] [Green Version]
- Horch, H.W.; Krüttgen, A.; Portbury, S.D.; Katz, L.C. Destabilization of cortical dendrites and spines by BDNF. Neuron 1999, 23, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Thoenen, H.; Zafra, F.; Hengerer, B.; Lindholm, D. The synthesis of nerve growth factor and brain-derived neurotrophic factor in hippocampal and cortical neurons is regulated by specific transmitter systems. Ann. N. Y. Acad. Sci. 1991, 640, 86–90. [Google Scholar] [CrossRef]
- Lindholm, D.; Castrén, E.; Berzaghi, M.; Blöchl, A.; Thoenen, H. Activity-dependent and hormonal regulation of neurotrophin mRNA levels in brain-implications for neuronal plasticity. J. Neurobiol. 1994, 25, 1362–1372. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P. Stress hormones, proinflammatory and antiinflammatory cytokines, and autoimmunity. Ann. N. Y. Acad. Sci. 2002, 966, 290–303. [Google Scholar] [CrossRef]
- Pousset, F.; Cremona, S.; Dantzer, R.; Kelley, K.; Parnet, P. Interleukin-4 and interleukin-10 regulate IL1-β induced mouse primary astrocyte activation: A comparative stud. GLIA 1999, 26, 12–21. [Google Scholar] [CrossRef]
- Hwang, K.-A.; Hwang, Y.-J.; Hwang, I.-G.; Song, J.; Kim, Y.J. Low temperature-aged garlic extract suppresses psychological stress by modulation of stress hormones and oxidative stress response in brain. J. Chin. Med. Assoc. 2019, 82, 191–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cory, S.; Adams, J.M. The Bcl2 family: Regulators of the cellular life-or-death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Kosten, T.A.; Galloway, M.P.; Duman, R.S.; Russell, D.S.; D’sa, C. Repeated unpredictable stress and antidepressants differentially regulate expression of the bcl-2 family of apoptotic genes in rat cortical, hippocampal, and limbic brain structures. Neuropsychopharmacology 2008, 33, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Esther, P.N.; Núria, G.; Elena, G.; Jordi, A. Brain-derived neurotrophic factor prevents changes in Bcl-2 family members and caspase-3 activation induced by excitotoxicity in the striatum. J. Neurochem. 2005, 92, 678–691. [Google Scholar]
- Sapolsky, R.M. The possibility of neurotoxicity in the hippocampus in major depression: A primer on neuron death. Biol. Psychiatry. 2000, 48, 755–765. [Google Scholar] [CrossRef]
- Farahani, M.S.; Bahramsoltani, R.; Farzaei, M.H.; Abdollahi, M.; Rahimi, R. Plant-derived natural medicines for the management of depression: An overview of mechanisms of action. Rev. Neurosci. 2015, 26, 305–321. [Google Scholar] [CrossRef]
- Martins, J.; Brijesh, S. Anti-depressant activity of Erythrina variegate bark extract and regulation of monoamine oxidase activities in mice. J. Ethnopharmacol. 2020, 248, 112280. [Google Scholar] [CrossRef]
- Setorki, M. Medicinal herbs with anti-depressant effects. J. Herbmed Pharmacol. 2020, 9, 309–317. [Google Scholar] [CrossRef]
- Lee, S.Y.; Weon, H.Y.; Kim, J.J.; Han, J.H. Biocontrol of leaf mustard powdery mildew caused by Erysiphe cruciferarm using Bacillus velezensis YP2. Korean J. Pestic. Sci. 2016, 20, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Mahmudah, R.; Hasanuddin, S.; Saleh, A.; Yuliastri, W.O.; Isrul, M. Antidepressant activity and identification of chemical compounds extract mustard leaves (Brassica juncea L.). Res. J. Pharm. Technol. 2019, 12, 3223–3227. [Google Scholar] [CrossRef]
- Verma, D.; Lakhanpal, N.; Singh, K. Genome-wide identification and characterization of abiotic-stress responsive SOD (superoxide dismutase) gene family in Brassica juncea and B. rapa. BMC Genom. 2019, 20, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Xie, K.; Yang, X.; Gu, J.; Ge, L.; Wang, X.; Wang, Z. Resveratrol reverses the effects of chronic unpredictable mild stress on behavior, serum corticosterone levels and BDNF expression in rats. Behav. Brain Res. 2014, 264, 9–16. [Google Scholar] [CrossRef]
- Afsar, T.; Razak, S.; Khan, R.A.; Almajwal, A. Anti-depressant and anxiolytic potential of Acacia hydaspica R. Parker aerial parts extract: Modulation of brain antioxidant enzyme status. BMC Complement. Altern. Med. 2017, 17, 228. [Google Scholar] [CrossRef]
- Yan, T.; He, B.; Wan, S.; Xu, M.; Yang, H.; Xiao, F.; Bi, K.; Jia, Y. Antidepressant-like effects and cognitive enhancement of Schisandra chinensis in chronic unpredictable mild stress mice and its related mechanism. Sci. Rep. 2017, 7, 6903. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.J.; Pack, S.H.; Shin, M.H. The effect of the meridian massage on the change of the low back pain RPE (rating of perceived exertion) and the changes of serum cortisol as well as stress index of middle aged women. J. Korean Soc. Cosmet. 2013, 19, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Deng, A.; Zhou, T.; Ding, F. Pretreatment with 2-(4-Methoxyphenyl)ethyl-2-acetamido-2-deoxy-β-D-pyranoside Attenuates Cerebral Ischemia/Reperfusion-Induced Injury In Vitro and In Vivo. PLoS ONE 2014, 9, e100126. [Google Scholar] [CrossRef]
- Zhou, C.H.; Zhang, Y.H.; Xue, F.; Xue, S.S.; Chen, Y.C.; Gu, T.; Peng, Z.W.; Wang, H.N. Isoflurane exposure regulates the cell viability and BDNF expression of astrocytes via upregulation of TREK-1. Mol. Med. Rep. 2017, 16, 7305–7314. [Google Scholar] [CrossRef] [Green Version]
- Haskó, G.; Szabó, C. IL-12 as a therapeutic target for pharmacological modulation in immune-mediated and inflammatory diseases: Regulation of T helper 1/T helper 2 responses. Br. J. Pharmacol. 1999, 127, 1295–1304. [Google Scholar]
- Harro, J.; Tõnissaar, M.; Eller, M.; Kask, A.; Oreland, L. Chronic variable stress and partial 5-HT denervation by parachloroamphetamine treatment in the rat: Effects on behavior and monoamine neurochemistry. Brain Res. 2001, 899, 227–239. [Google Scholar] [CrossRef]
- Fasipe, O.J. Neuropharmacological classification of antidepressant agents based on their mechanisms of action. Arch. Med. Health Sci. 2018, 6, 81–94. [Google Scholar] [CrossRef]
- Cunha, A.B.M.; Frey, B.N.; Andreazza, A.C.; Goi, J.D.; Rosa, A.D.; Goncalves, C.A.; Santin, A.; Kapczinski, F. Serum brain-derived neurotrophic factor is decreased in bipolar disorder during depressive and manic episodes. Neurosci. Lett. 2006, 398, 215–219. [Google Scholar] [CrossRef]
- Dunham, J.S.; Deakin, J.F.W.; Miyajima, F.; Payton, A.; Toro, C.T. Expression of hippocampal brain-derived neurotrophic factor and its receptors in Stanley consortium brains. J. Psychiatr. Res. 2009, 43, 1175–1184. [Google Scholar] [CrossRef] [Green Version]
- Nibuya, M.; Takahashi, M.; Russell, D.S.; Duman, R.S. Repeated stress increases catalytic TrkB mRNA in rat hippocampus. Neurosci. Lett. 1999, 267, 81–84. [Google Scholar] [CrossRef]
- Chen, B.; Dowlatshahi, D.; MacQueen, G.M.; Wang, J.-F.; Young, L.T. Increased hippocampal BDNF immunoreactivity in subjects treated with antidepressant medication. Biol. Psychiatry 2001, 50, 260–265. [Google Scholar] [CrossRef]
- Karege, F.; Vaudan, G.; Schwald, M.; Perroud, N.; La Harpe, R. Neurotrophin levels in postmortem brains of suicide victims and the effects of antemortem diagnosis and psychotropic drugs. Mol. Brain Res. 2005, 136, 29–37. [Google Scholar] [CrossRef]
- Shirayama, Y.; Chen, A.C.-H.; Nakagawa, S.; Russell, D.S.; Duman, R.S. Brain-derived neurotrophic factor produces antidepressant effects in behavioral models of depression. J. Neurosci. 2002, 22, 3251–3261. [Google Scholar] [CrossRef] [Green Version]
- You, Z.; Luo, C.; Zhang, W.; Chen, Y.; He, J.; Zhao, Q.; Zuo, R.; Wu, Y. Pro- and anti-inflammatory cytokines expression in rat’s brain and spleen exposed to chronic mild stress: Involvement in depression. Behav. Brain Res. 2011, 225, 135–141. [Google Scholar] [CrossRef]
- Molina-Holgado, E.; Arévalo-Martín, A.; Castrillo, A.; Boscá, L.; Vela, J.M.; Guaza, C. Interleukin-4 and interleukin-10 modulate nuclear factor κB activity and nitric oxide synthase-2 expression in Theiler’s virus-infected brain astrocytes. J. Neurochem. 2002, 81, 1242–1252. [Google Scholar] [CrossRef] [Green Version]
- Antonsson, B.; Martinou, J.C. The Bcl-2 protein family. Exp. Cell Res. 2000, 256, 50–57. [Google Scholar] [CrossRef]
- Jürgensmeier, J.M.; Xie, Z.; Deveraux, Q.; Ellerby, L.; Bredesen, D.; Reed, J.C. Bax directly induces release of cytochrome c from isolated mitochondria. Proc. Natl. Acad. Sci. USA 1998, 95, 4997–5002. [Google Scholar]
- Nagata, S. Apopotosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.J.; Lee, D.J.; Kim, M.O.; Jung, B.; Kim, S.O.; Sohn, N.W.; Lee, E.H. Reduction but not cleavage of poly(ADP-ribose) polymerase during stress-mediated cell death in the rat hippocampus. Neuroreport 2003, 14, 935–939. [Google Scholar] [CrossRef]
- Kim, K.-W.; Ha, K.-Y.; Lee, J.-S.; Rhyu, K.-W.; An, H.S.; Woo, Y.K. The apoptotic effects of oxidative stress and antiapoptotic effects of caspase inhibitors on rat notochordal cells. Spine 2007, 32, 2443–2448. [Google Scholar] [CrossRef] [Green Version]
- Wahjuni, S.; Gunawan, I.W.G.; Malindo, I.Y.D. The effect of mustard greens (Brassica rapa l.) ethanol extract on blood glucose and malondialdehyde levels of hyperglycemic Wistar rats. Bali Med. J. 2019, 8, 35–40. [Google Scholar] [CrossRef]
- Jaganathan, S.K.; Supriyanto, E. Antiproliferative and Molecular Mechanism of Eugenol-Induced Apoptosis in Cancer Cells. Molecules 2012, 17, 6290–6304. [Google Scholar] [CrossRef]
- Said, M.M.; Abd Rabo, M.M. Neuroprotective effects of eugenol against aluminiuminduced toxicity in the rat brain. Arh. Hig. Rada. Toksikol. 2017, 68, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Garabadu, D.; Shah, A.; Ahmad, A.; Joshi, V.B.; Saxena, B.; Palit, G.; Krishnamurthy, S. Eugenol as an anti-stress agent: Modulation of hypothalamic–pituitary–adrenal axis and brain monoaminergic systems in a rat model of stress. Stress 2010, 14, 145–155. [Google Scholar] [CrossRef]
- Solberg, L.C.; Ahmadiyeh, N.; Baum, A.E.; Vitaterna, M.H.; Takahashi, J.S.; Turek, F.W.; Redei, E.E. Depressive-like behavior and stress reactivity are independent traits in a Wistar Kyoto × Fisher 344 cross. Mol. Psychiatry 2003, 8, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Zhao, Y.; Wang, Y.; Liu, L.; Zhang, X.; Li, B.; Cuia, R. The Effects of Psychological Stress on Depression. Curr. Neuropharmacol. 2015, 4, 494–504. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment (mg/kg) | Condition |
|---|---|---|
| Normal | PBS | No CRS |
| Control | PBS | CRS |
| PX | Paroxetine (10) | CRS |
| LML | Low-concentration mustard leaf (100) | CRS |
| HML | High-concentration mustard leaf (500) | CRS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, K.-A.; Hwang, H.-J.; Hwang, Y.J.; Kim, Y.J. Mustard Leaf Extract Suppresses Psychological Stress in Chronic Restraint Stress-Subjected Mice by Regulation of Stress Hormone, Neurotransmitters, and Apoptosis. Nutrients 2020, 12, 3640. https://doi.org/10.3390/nu12123640
Hwang K-A, Hwang H-J, Hwang YJ, Kim YJ. Mustard Leaf Extract Suppresses Psychological Stress in Chronic Restraint Stress-Subjected Mice by Regulation of Stress Hormone, Neurotransmitters, and Apoptosis. Nutrients. 2020; 12(12):3640. https://doi.org/10.3390/nu12123640
Chicago/Turabian StyleHwang, Kyung-A., Hye-Jeong Hwang, Yu Jin Hwang, and Young Jun Kim. 2020. "Mustard Leaf Extract Suppresses Psychological Stress in Chronic Restraint Stress-Subjected Mice by Regulation of Stress Hormone, Neurotransmitters, and Apoptosis" Nutrients 12, no. 12: 3640. https://doi.org/10.3390/nu12123640