Modulation of Adhesion Process, E-Selectin and VEGF Production by Anthocyanins and Their Metabolites in an In Vitro Model of Atherosclerosis

 ,
,  ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Anthocyanin and Metabolite Standards
2.3. Cell Culture
2.4. Cytotoxicity Assay
2.5. Evaluation of Monocytes Adhesion on Activated Human Umbilical Vein Endothelial Cells
2.6. ELISA Quantification of Soluble VCAM-1, E-Selectin and VEGF
2.7. Data Analysis
3. Results
3.1. Effect of Peo-3-glc, Pet-3-glc, VA and MetGA on Cell Cytotoxicity
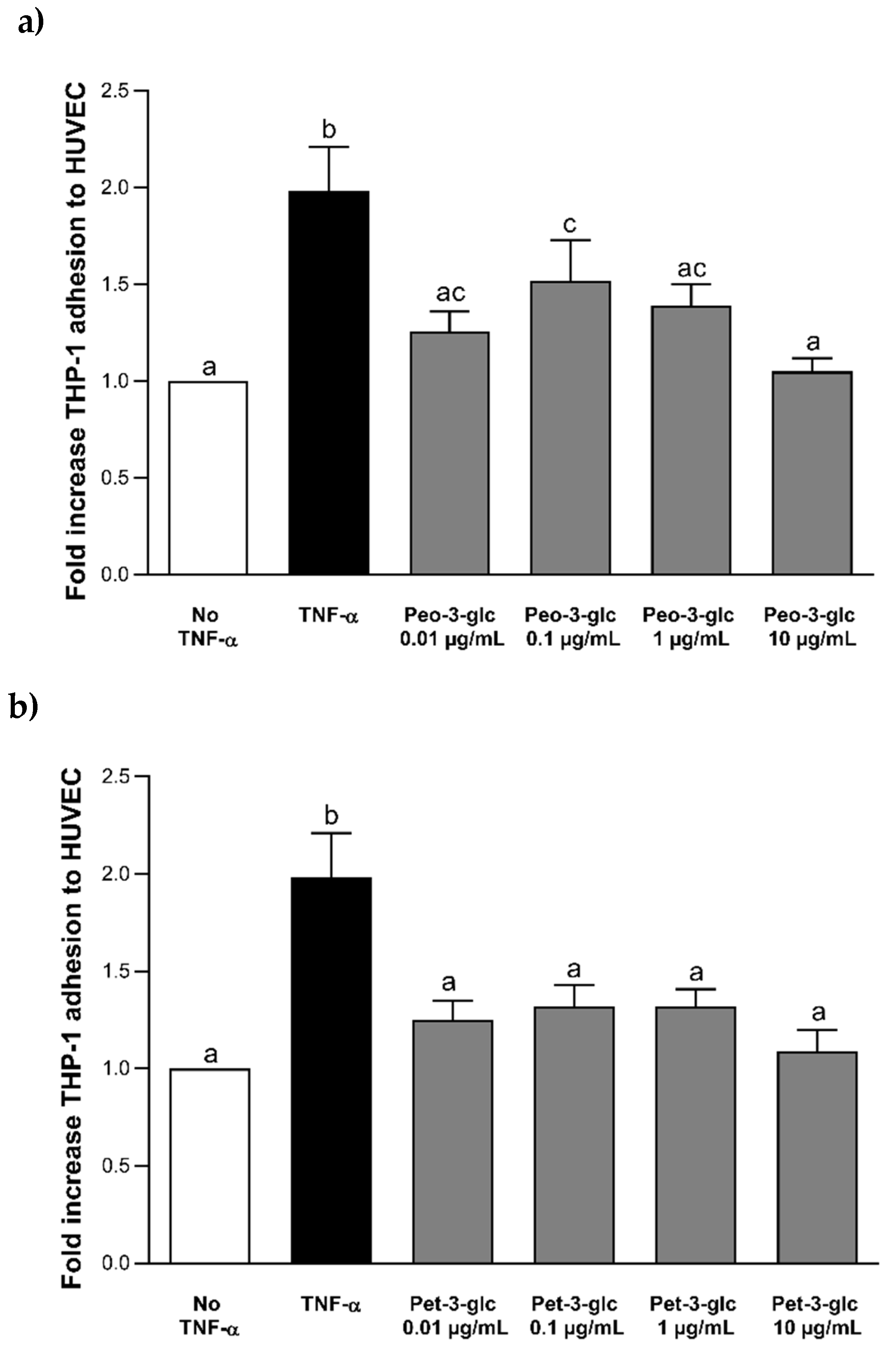
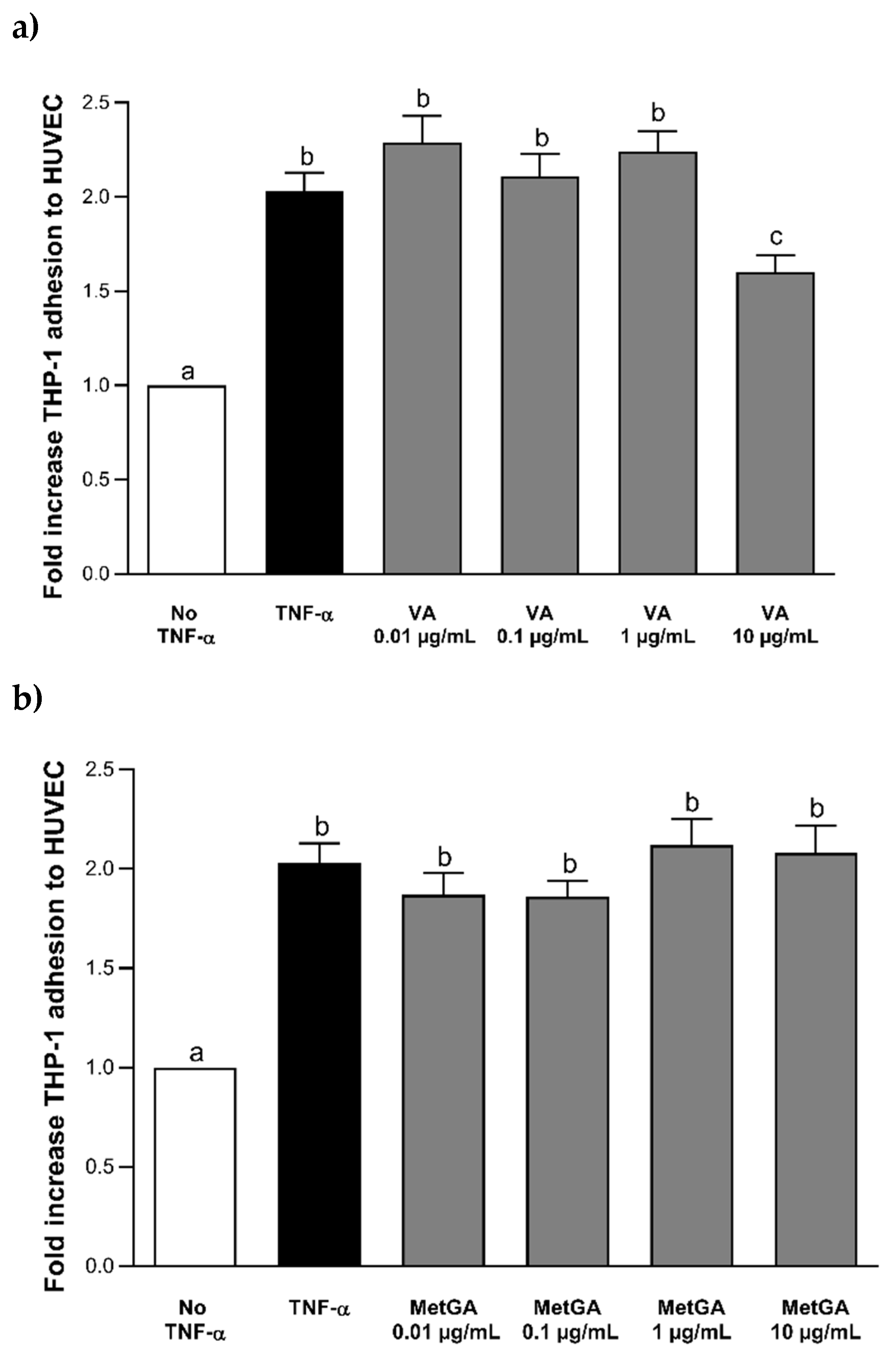
3.2. Effect of Peo-3-glc, Pet-3-glc, VA and MetGA on THP-1 Adhesion to HUVECs
3.3. Effect of Peo-3-glc, Pet-3-glc, VA and MetGA on the Levels of E-Selectin
3.4. Effect of Peo-3-glc, Pet-3-glc, VA and MetGA on the Levels of Soluble VCAM-1
3.5. Effect of Peo-3-glc, Pet-3-glc, VA and MetGA on the Levels of VEGF
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef]
- Čejková, S.; Králová-Lesná, I.; Poledne, R. Monocyte adhesion to the endothelium is an initial stage of atherosclerosis development. Cor et Vasa 2016, 58, e419–e425. [Google Scholar] [CrossRef]
- Liao, J.K. Linking endothelial dysfunction with endothelial cell activation. Clin. Investig. 2013, 123, 540–541. [Google Scholar] [CrossRef]
- Camaré, C.; Pucelle, M.; Nègre-Salvayre, A.; Salvayre, R. Angiogenesis in the atherosclerotic plaque. Redox Biol. 2017, 12, 18–34. [Google Scholar] [CrossRef]
- Michel, J.B.; Martin-Ventura, J.L.; Nicoletti, A.; Ho-Tin-Noé, B. Pathology of human plaque vulnerability: Mechanisms and consequences of intraplaque haemorrhages. Atherosclerosis 2014, 234, 311–319. [Google Scholar] [CrossRef]
- Crozier, A.; Yokota, T.; Jaganath, I.B.; Marks, S.; Saltmarsh, M.; Clifford, M.N. Secondary metabolites as dietary components in plant-based foods and beverages. In Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; Crozier, A., Clifford, M.N., Ashi-hara, H., Eds.; Blackwell Publishing: Oxford, UK, 2006; pp. 208–302. [Google Scholar]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Calhau, C.; de Freitas, V.; Mateus, M. Bioavailability of anthocyanins and derivatives. J. Funct. Foods 2014, 7, 54–66. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Krga, I.; Milenkovic, D. Anthocyanins: From Sources and Bioavailability to Cardiovascular-Health Benefits and Molecular Mechanisms of Action. J. Agric. Food Chem. 2019, 67, 1771–1783. [Google Scholar] [CrossRef]
- de Ferrars, R.M.; Czank, C.; Zhang, Q.; Botting, N.P.; Kroon, P.A.; Cassidy, A.; Kay, C.D. The pharmacokinetics of anthocyanins and their metabolites in humans. Br. J. Pharmacol. 2014, 171, 3268–3282. [Google Scholar] [CrossRef] [Green Version]
- Martini, D.; Marino, M.; Angelino, D.; Del Bo’, C.; Del Rio, D.; Riso, P.; Porrini, M. Role of berries in vascular function: A systematic review of human intervention studies. Nutr. Rev. 2019, 78, 189–206. [Google Scholar] [CrossRef]
- Del Bo’, C.; Deon, V.; Campolo, J.; Lanti, C.; Parolini, M.; Porrini, M.; Klimis-Zacas, D.; Riso, P. A serving of blueberry (V. corymbosum) acutely improves peripheral arterial dysfunction in young smokers and non-smokers: Two randomized, controlled, crossover pilot studies. Food Funct. 2017, 8, 4108. [Google Scholar] [CrossRef]
- Leyva-Soto, A.; Chavez-Santoscoy, R.A.; Lara-Jacobo, L.R.; Chavez-Santoscoy, A.V.; Gonzalez-Cobian, L.N. Daily Consumption of Chocolate Rich in Flavonoids Decreases Cellular Genotoxicity and Improves Biochemical Parameters of Lipid and Glucose Metabolism. Molecules 2018, 23, 2220. [Google Scholar] [CrossRef]
- Bondonno, N.P.; Bondonno, C.P.; Blekkenhorst, L.C.; Considine, M.J.; Maghzal, G.; Stocker, R.; Woodman, R.J.; Ward, N.C.; Hodgson, J.M.; Croft, K.D. Flavonoid-Rich Apple Improves Endothelial Function in Individuals at Risk for Cardiovascular Disease: A Randomized Controlled Clinical Trial. Mol. Nutr. Food Res. 2018, 62, 1700674. [Google Scholar] [CrossRef]
- Grassi, D.; Draijer, R.; Schalkwijk, C.; Desideri, G.; D’Angeli, A.; Francavilla, S.; Mulder, T.; Ferri, C. Black Tea Increases Circulating Endothelial Progenitor Cells and Improves Flow Mediated Dilatation Counteracting Deleterious Effects from a Fat Load in Hypertensive Patients: A Randomized Controlled Study. Nutrients 2016, 8, 727. [Google Scholar] [CrossRef]
- Guglielmi, F.; Luceri, C.; Giovannelli, L.; Dolara, P.; Lodovici, M. Effect of 4-coumaric and 3,4-dihydroxybenzoic acid on oxidative DNA damage in rat colonic mucosa. Br. J. Nutr. 2003, 89, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Martín, M.A.; Serrano, A.B.; Ramos, S.; Pulido, M.I.; Bravo, L.; Goya, L. Cocoa flavonoids up-regulate antioxidant enzyme activity via the ERK1/2 pathway to protectagainst oxidative stress-induced apoptosis in HepG2 cells. J. Nutr. Biochem. 2010, 21, 196–205. [Google Scholar] [CrossRef]
- Iskender, H.; Yenice, G.; Dokumacioglu, E.; Kaynar, O.; Hayirli, A.; Kaya, A. The Effects of Dietary Flavonoid Supplementation on the Antioxidant Status of Laying Hens. Br. J. Poult. Sci. 2016, 18, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xu, L.; Sang, R.; Yu, Y.; Ge, B.; Zhang, X. Immunomodulatory and anti-inflammatory effects of total flavonoids of Astragalus by regulating NF-ΚB and MAPK signalling pathways in RAW 264.7 macrophages. Die Pharm. Int. J. Pharm. Sci. 2018, 73, 589–593. [Google Scholar] [CrossRef]
- Le Phuong Nguyen, T.; Fenyvesi, F.; Remenyik, J.; Homoki, J.R.; Gogolák, P.; Bácskay, I.; Fehér, P.; Ujhelyi, Z.; Vasvári, G.; Vecsernyés, M.; et al. Protective Effect of Pure Sour Cherry Anthocyanin Extract on Cytokine-Induced Inflammatory Caco-2 Monolayers. Nutrients 2018, 10, 861. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, D.; Cimino, F.; Fratantonio, D.; Molonia, M.S.; Bashllari, R.; Busà, R.; Saija, A.; Speciale, A. Cyanidin-3-O-Glucoside Modulates the In Vitro Inflammatory Crosstalk between Intestinal Epithelial and Endothelial Cells. Mediat. Inflamm. 2017, 2017, 3454023. [Google Scholar] [CrossRef] [PubMed]
- Oak, M.H.; Auger, C.; Belcastro, E.; Park, S.H.; Lee, H.H.; Schini-Kerth, V.B. Potential mechanisms underlying cardiovascular protection by polyphenols: Role of the endothelium. Free Radic. Biol. Med. 2018, 122, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Luján-Barroso, L.; Slimani, N.; Romieu, I.; Touillaud, M.; Kaaks, R.; Teucher, B.; Mattiello, A.; Grioni, S.; et al. Estimation of the intake of anthocyanidins and their food sources in the European Prospective Investigation into Cancer and Nutrition (EPIC) study. Br. J. Nutr. 2011, 106, 1090–1099. [Google Scholar] [CrossRef] [Green Version]
- Jaipersad, A.S.; Lip, G.Y.; Silverman, S.; Shantsila, E. The role of monocytes in angiogenesis and atherosclerosis. J. Am. Coll. Cardiol. 2014, 63, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Del Bo’, C.; Marino, M.; Riso, P.; Møller, P.; Porrini, M. Anthocyanins and metabolites resolve TNF-α-mediated production of E-selectin and adhesion of monocytes to endothelial cells. Chem. Biol. Interact. 2019, 300, 49–55. [Google Scholar] [CrossRef]
- Del Bo’, C.; Roursgaard, M.; Porrini, M.; Loft, S.; Møller, P.; Riso, P. Different effects of anthocyanins and phenolic acids from wild blueberry (Vaccinium angustifolium) on monocytes adhesion to endothelial cells in a TNF-α stimulated proinflammatory environment. Mol. Nutr. Food Res. 2016, 60, 2355–2366. [Google Scholar] [CrossRef]
- Krga, I.; Monfoulet, L.M.; Konic-Ristic, A.; Mercier, S.; Glibetic, M.; Morand, C.; Milenkovic, D. Anthocyanins and their gut metabolites reduce the adhesion of monocyte to TNFa-activated endothelial cells at physiologically relevant concentrations. Arch. Biochem. Biophys. 2016, 599, 51–59. [Google Scholar] [CrossRef]
- McEver, R.P. Selectins: Initiators of leucocyte adhesion and signalling at the vascular wall. Cardiovasc. Res. 2015, 107, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Klaus, L.; Yuqing, H. VCAM-1 is critical in atherosclerosis. J. Clin. Investig. 2001, 107, 1209–1210. [Google Scholar] [CrossRef]
- Jong-Gil, P.; Su, Y.R.; In-Hyuk, J.; You-Han, L.; Kyung, J.K.; Mi-Ran, L.; Mi-Ni, L.; Seong, K.S.; Jeong, H.L.; Hang, L.; et al. Evaluation of VCAM-1 antibodies as therapeutic agent for atherosclerosis in apolipoprotein E-deficient mice. Atherosclerosis 2013, 226, 356–363. [Google Scholar] [CrossRef]
- Warner, E.F.; Zhang, Q.; Raheem, K.; O’Hagan, D.; O’Connell, M.A.; Kay, C.D. Common Phenolic Metabolites of Flavonoids, but Not Their Unmetabolized Precursors, Reduce the Secretion of Vascular Cellular Adhesion Molecules by Human Endothelial Cells. J. Nutr. 2016, 146, 465–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntz, S.; Kunz, C.; Domann, E.; Würdemann, N.; Unger, F.; Römpp, A.; Rudloff, S. Inhibition of Low-Grade Inflammation by Anthocyanins after Microbial Fermentation in Vitro. Nutrients 2016, 8, 411. [Google Scholar] [CrossRef] [PubMed]
- Amin, H.P.; Czank, C.; Raheem, S.; Zhang, Q.; Botting, N.P.; Cassidy, A.; Kay, C.D. Anthocyanins and their physiologically relevant metabolites alter the expression of IL-6 and VCAM-1 in CD40L and oxidized LDL challenged vascular endothelial cells. Mol. Nutr. Food Res. 2015, 59, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Calabriso, N.; Massaro, M.; Scoditti, E.; Pasqualone, A.; Laddomada, B.; Carluccio, M.A. Phenolic extracts from whole wheat biofortified bread dampen overwhelming inflammatory response in human endothelial cells and monocytes: Major role of VCAM-1 and CXCL-10. Eur. J. Nutr. 2019, 1–13. [Google Scholar] [CrossRef]
- Tang, J.S.; Bozonet, S.M.; McKenzie, J.L.; Anderson, R.F.; Melton, L.D.; Vissers, M.C.M. Physiological Concentrations of Blueberry-Derived Phenolic Acids Reduce Monocyte Adhesion to Human Endothelial Cells. Mol. Nutr. Food Res. 2019, 63, e1900478. [Google Scholar] [CrossRef]
- Katsi, V.; Ioannis, Z.; Stavroula, M.; Thomas, M.; Petros, N.; Dimitris, T.; Ioannis, K. Anti-VEGF Anticancer Drugs: Mind the Hypertension. Recent Adv. Cardiovasc. Drug Dis. 2014, 9, 63–72. [Google Scholar] [CrossRef]
- Sueishi, K.; Yonemitsu, Y.; Nakagawa, K.; Kaneda, Y.; Kumamoto, M.; Nakashima, Y. Atherosclerosis and angiogenesis. Its pathophysiological significance in humans as well as in an animal model induced by the gene transfer of vascular endothelial growth factor. Ann. N. Y. Acad. Sci. 1997, 811, 311–324. [Google Scholar] [CrossRef]
- Qiu, J.; Lei, D.; Hu, J.; Yin, T.; Zhang, K.; Yu, D.; Wang, G. Effect of intraplaque angiogenesis to atherosclerotic rupture-prone plaque induced by high shear stress in rabbit model. Regen. Biomater. 2017, 4, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Lamy, S.; Blanchette, M.; Michaud-Levesque, J.; Lafleur, R.; Durocher, Y.; Moghrabi, A.; Barrette, S.; Gingras, D.; Béliveau, R. Delphinidin, a dietary anthocyanidin, inhibits vascular endothelial growth factor receptor-2 phosphorylation. Carcinogenesis 2006, 27, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Keravis, T.; Favot, L.; Abusnina, A.A.; Anton, A.; Justiniano, H.; Soleti, R.; Alabed Alibrahim, E.; Simard, G.; Andriantsitohaina, R.; Lugnier, C. Delphinidin Inhibits Tumor Growth by Acting on VEGF Signalling in Endothelial Cells. PLoS ONE 2015, 10, e0145291. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Jeong, Y.J.; Cho, H.J.; Hoe, H.S.; Park, K.K.; Park, Y.Y.; Choi, Y.H.; Kim, C.H.; Chang, H.W.; Park, Y.J.; et al. Delphinidin inhibits angiogenesis through the suppression of HIF-1α and VEGF expression in A549 lung cancer cells. Oncol. Rep. 2017, 37, 777–784. [Google Scholar] [CrossRef]
- Tanaka, J.; Nakamura, S.; Tsuruma, K.; Shimazawa, M.; Shimoda, H.; Hara, H. Purple rice (Oryza sativa L.) extract and its constituents inhibit VEGF-induced angiogenesis. Phytother. Res. 2012, 26, 214–222. [Google Scholar] [CrossRef]
- Negrão, R.; Costa, R.; Duarte, D.; Gomes, T.T.; Azevedo, I.; Soares, R. Different effects of catechin on angiogenesis and inflammation depending on VEGF levels. J. Nutr. Biochem. 2013, 24, 435–444. [Google Scholar] [CrossRef]
- Calabriso, N.; Massaro, M.; Scoditti, E.; D’Amore, S.; Gnoni, A.; Pellegrino, M.; Storelli, C.; De Caterina, R.; Palasciano, G.; Carluccio, M.A. Extra virgin olive oil rich in polyphenols modulates VEGF-induced angiogenic responses by preventing NADPH oxidase activity and expression. J. Nutr. Biochem. 2016, 28, 19–29. [Google Scholar] [CrossRef]
- Tsakiroglou, P.; Weber, J.; Ashworth, S.; Del Bo’, C.; Klimis-Zacas, D. Phenolic and anthocyanin fractions from wild blueberries (V. angustifolium) differentially modulate endothelial cell migration partially through RHOA and RAC1. J. Cell. Biochem. 2019, 120, 11056–11067. [Google Scholar] [CrossRef]
- Cerezo, A.B.; Winterbone, M.S.; Moyle, C.W.; Needs, P.W.; Kroon, P.A. Molecular structure-function relationship of dietary polyphenols for inhibiting VEGF-induced VEGFR-2 activity. Mol. Nutr. Food Res. 2015, 59, 2119–2131. [Google Scholar] [CrossRef] [Green Version]
- Perez-Moral, N.; Needs, P.W.; Moyle, C.W.A.; Kroon, P.A. Hydrophobic Interactions Drive Binding between Vascular Endothelial Growth Factor-A (VEGFA) and Polyphenolic Inhibitors. Molecules 2019, 24, 2785. [Google Scholar] [CrossRef] [Green Version]
- Tsakiroglou, P.; VandenAkker, N.E.; Del Bo’, C.; Riso, P.; Klimis-Zacas, D. Role of Berry Anthocyanins and Phenolic Acids on Cell Migration and Angiogenesis: An Updated Overview. Nutrients 2019, 11, 1075. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Trypan Blue Assay | Anthocyanins | Gut Metabolites | |||
|---|---|---|---|---|---|
| Concentrations | Peo-3-glc | Pet-3-glc | Concentrations | VA | MetGA |
| 0.02 µM | 99.7 ± 0.33 | 110 ± 0 | 0.05 µM | 100 ± 0 | 99.7 ± 0.33 |
| 0.2 µM | 100 ± 0 | 97.0 ± 1.0 | 0.5 µM | 99.7 ± 0.33 | 99.3 ± 0.67 |
| 2 µM | 99.3 ± 0.67 | 97.7 ± 0.33 | 5 µM | 99.7 ± 0.66 | 98.7 ± 1.33 |
| 20 µM | 99.3 ± 0.33 | 100 ± 0 | 50 µM | 99.3 ± 0.67 | 97.3 ± 1.77 |
| MTT assay | Anthocyanins | Gut metabolites | |||
| Concentration | Peo-3-glc | Pet-3-glc | Concentration | VA | MetGA |
| 20 µM | 98.5 ± 0.12 | 94.4 ± 0.45 | 50 µM | 99.7 ± 0.32 | 96.7 ± 0.43 |
| Anthocyanins | Gut Metabolites | ||||
|---|---|---|---|---|---|
| Concentrations | Peo-3-glc | Pet-3-glc | Concentrations | VA | MetGA |
| TNF-α 0 ng mL−1 | 160 ± 7.9 a | 164 ± 5.8 a | TNF-α 0 ng mL−1 | 160 ± 7.9 a | 164 ± 5.8 a |
| TNF-α 100 ng mL−1 | 316 ± 8.1 b | 317 ± 6.3 b | TNF-α 100 ng mL−1 | 316 ± 8.1 b | 317 ± 6.3 b |
| 0.02 µM | 143 ± 4.3 a | 115 ± 7.5 c | 0.05 µM | 312 ± 14.1 b | 299 ± 7.5 b |
| 0.2 µM | 108 ± 5.3 c | 123 ± 11.8 a,c | 0.5 µM | 312 ± 11.2 b | 297 ± 7.5 b |
| 2 µM | 109 ± 7.2 c | 104 ± 6.3 c | 5 µM | 305 ± 7.4 b | 297 ± 8.0 b |
| 20 µM | 76 ± 8.4 c | 88 ± 12.1 c | 50 µM | 95 ± 13.2 c | 295 ± 7.3 b |
| Anthocyanins | Gut Metabolites | ||||
|---|---|---|---|---|---|
| Concentrations | Peo-3-glc | Pet-3-glc | Concentrations | VA | MetGA |
| TNF-α 0 ng mL−1 | 59 ± 9.0 a | 64 ± 10 a | TNF-α 0 ng mL−1 | 59 ± 9.0 a | 64 ± 10 a |
| TNF-α 100 ng mL−1 | 316 ± 16 b | 307 ± 11 b | TNF-α 100 ng mL−1 | 316 ± 16 b | 307 ± 11 b |
| 0.02 µM | 107 ± 15 c | 311 ± 13 b | 0.05 µM | 308 ± 11 b | 299 ± 15 b |
| 0.2 µM | 104 ± 16 c | 297 ± 15 b | 0.5 µM | 299 ± 22 b | 297 ± 15 b |
| 2 µM | 186 ± 12 c | 300 ± 14 b | 5 µM | 295 ± 12 b | 297 ± 16 b |
| 20 µM | 149 ± 24 c | 83 ± 10 c | 50 µM | 315 ± 16 c | 295 ± 14 b |
| Anthocyanins | Gut Metabolites | ||||
|---|---|---|---|---|---|
| Concentrations | Peo-3-glc | Pet-3-glc | Concentrations | VA | MetGA |
| TNF-α 0 ng mL−1 | 120 ± 6.9 a | 121 ± 6.1 a | TNF-α 0 ng mL−1 | 120 ± 6.9 a | 121 ± 6.1 a |
| TNF-α 100 ng mL−1 | 170 ± 8.5 b | 172 ± 7.9 b | TNF-α 100 ng mL−1 | 170 ± 8.5 b | 172 ± 7.9 b |
| 0.02 µM | 120 ± 6.9 a | 129 ± 10 a | 0.05 µM | 149 ± 3.0 c | 153 ± 2.5 c |
| 0.2 µM | 123 ± 1.7 a | 123 ± 7.4 a | 0.5 µM | 141 ± 8.3 c | 142 ± 3.0 c |
| 2 µM | 123 ± 6.0 a | 123 ± 2.9 a | 5 µM | 147 ± 4.9 c | 141 ± 4.9 c |
| 20 µM | 119 ± 2.6 a | 117 ± 9.9 a | 50 µM | 135 ± 5.7 c | 137 ± 6.0 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marino, M.; Bo’, C.D.; Tucci, M.; Klimis-Zacas, D.; Riso, P.; Porrini, M. Modulation of Adhesion Process, E-Selectin and VEGF Production by Anthocyanins and Their Metabolites in an In Vitro Model of Atherosclerosis. Nutrients 2020, 12, 655. https://doi.org/10.3390/nu12030655
Marino M, Bo’ CD, Tucci M, Klimis-Zacas D, Riso P, Porrini M. Modulation of Adhesion Process, E-Selectin and VEGF Production by Anthocyanins and Their Metabolites in an In Vitro Model of Atherosclerosis. Nutrients. 2020; 12(3):655. https://doi.org/10.3390/nu12030655
Chicago/Turabian StyleMarino, Mirko, Cristian Del Bo’, Massimiliano Tucci, Dorothy Klimis-Zacas, Patrizia Riso, and Marisa Porrini. 2020. "Modulation of Adhesion Process, E-Selectin and VEGF Production by Anthocyanins and Their Metabolites in an In Vitro Model of Atherosclerosis" Nutrients 12, no. 3: 655. https://doi.org/10.3390/nu12030655