
Buckwheat Flour and Its Starch Prevent Age-Related Cognitive Decline by Increasing Hippocampal BDNF Production in Senescence-Accelerated Mouse Prone 8 Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Animal Treatment
2.3.1. Study 1
2.3.2. Study 2
2.4. Barnes Maze Test
2.5. Passive Avoidance Test
2.6. Western Blot Analysis
2.7. Immunostaining
2.8. Microbiological Analysis
2.9. Statistical Analyses
3. Results
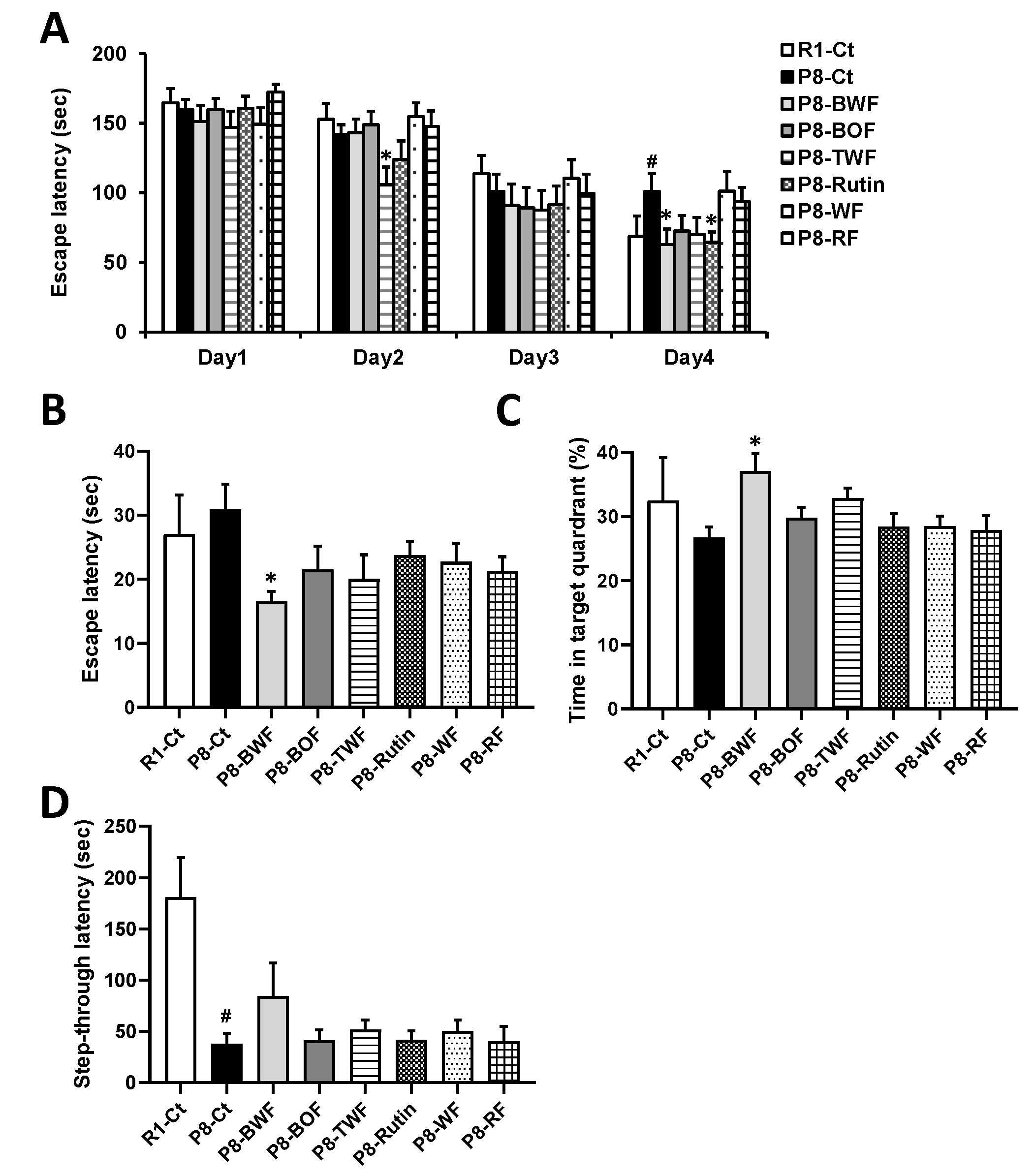
3.1. Effects of Buckwheat Flour on the Cognitive Function of SAMP8
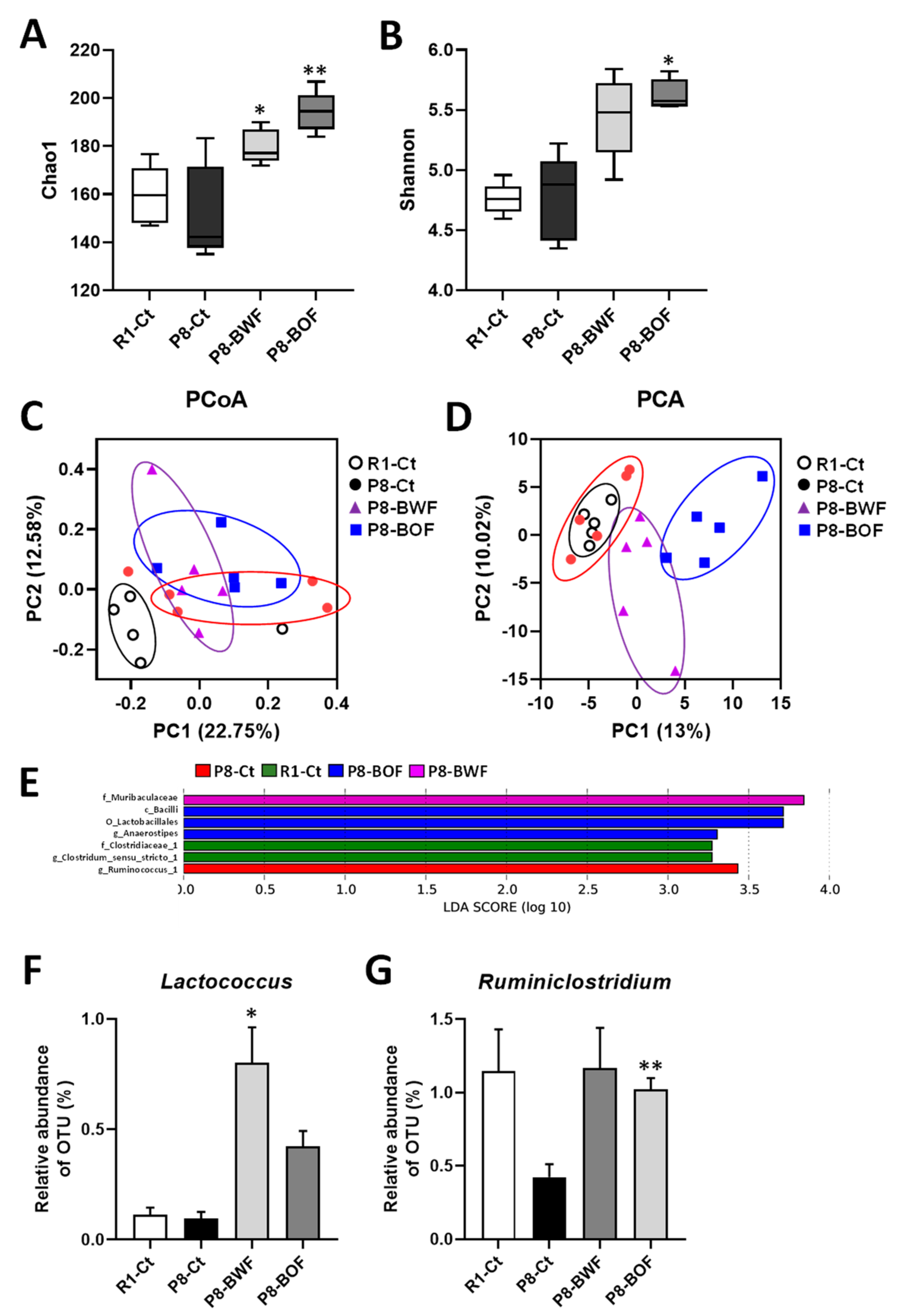
3.2. Effects of Buckwheat Flour on the Gut Microbiota Community of SAMP8
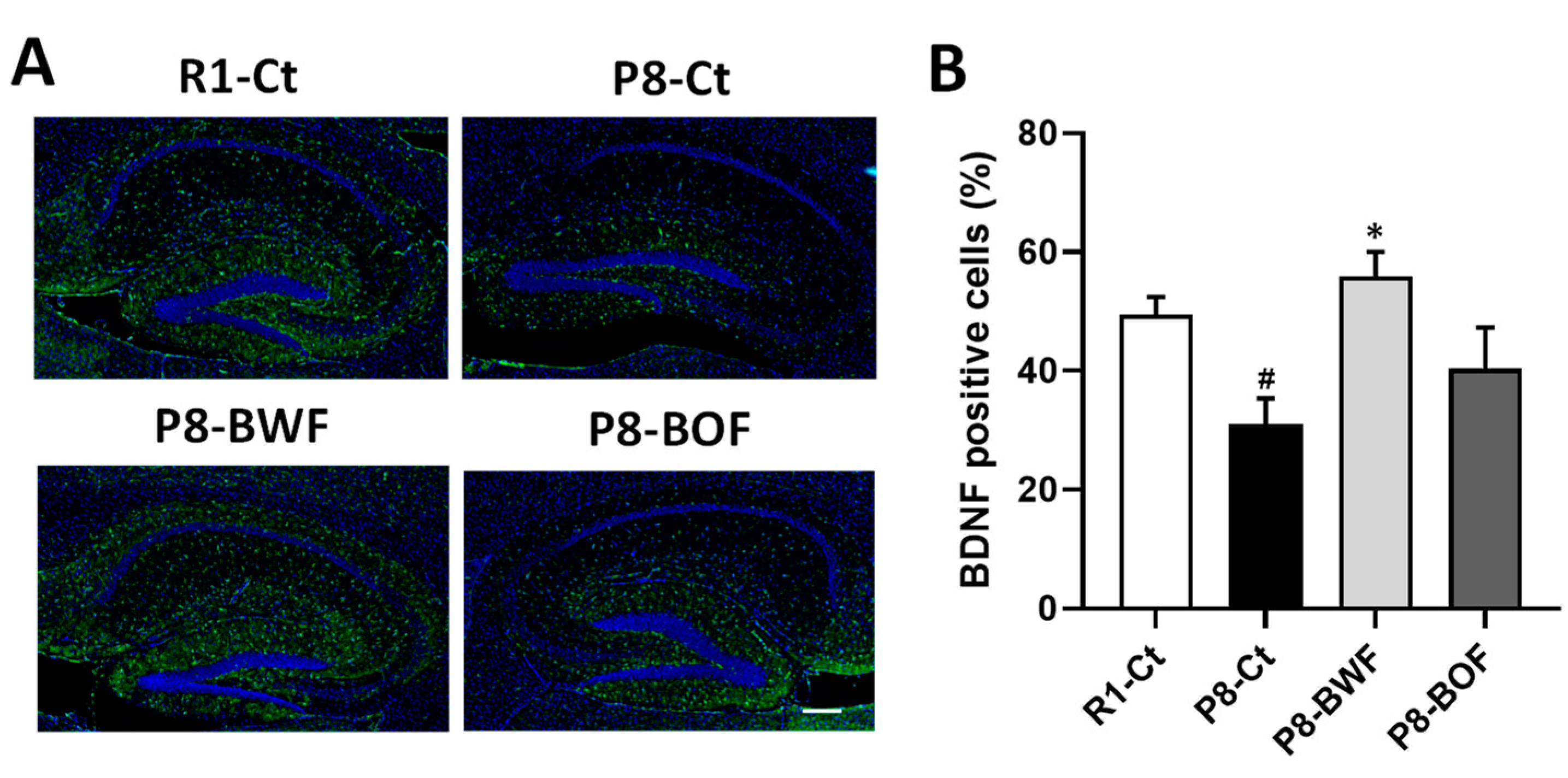
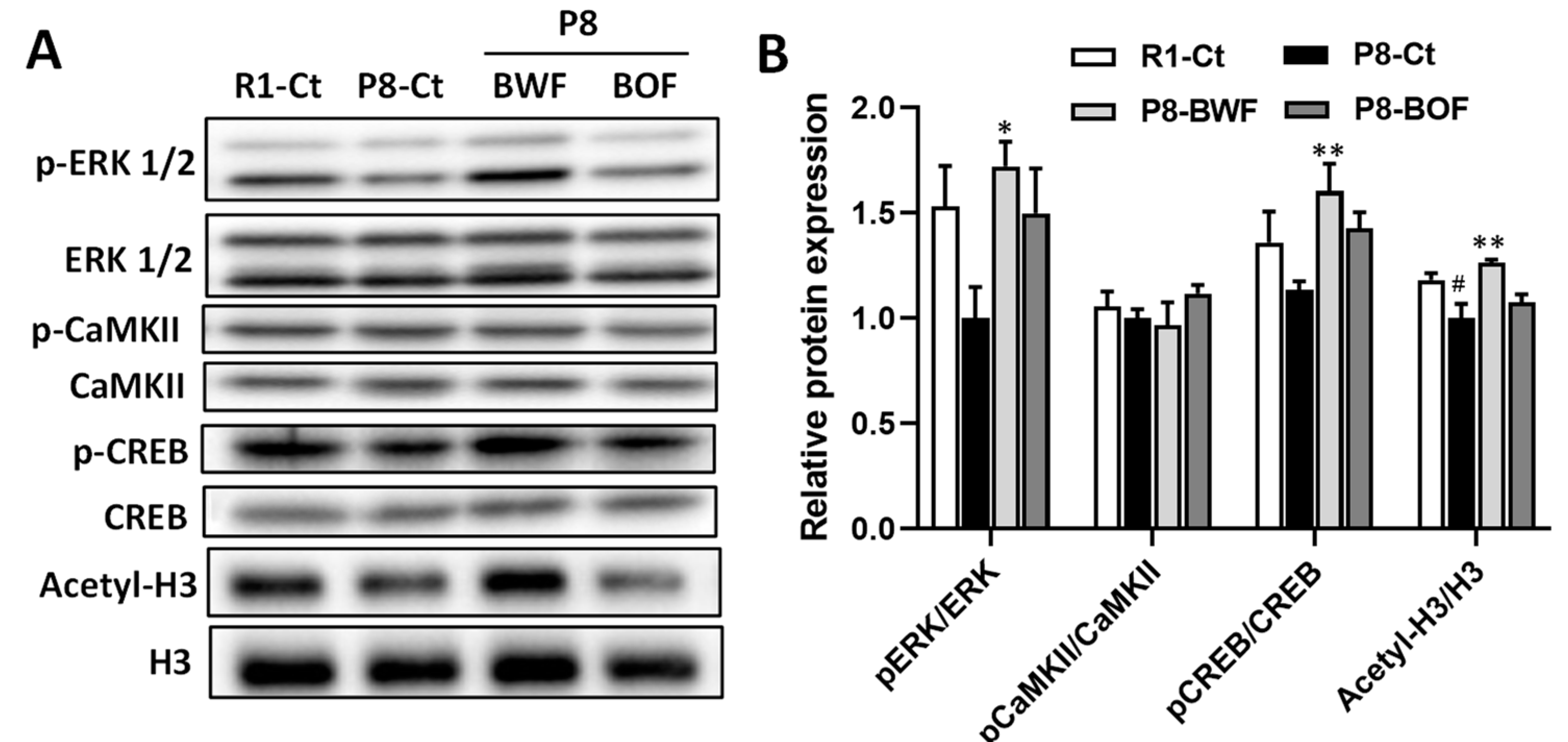
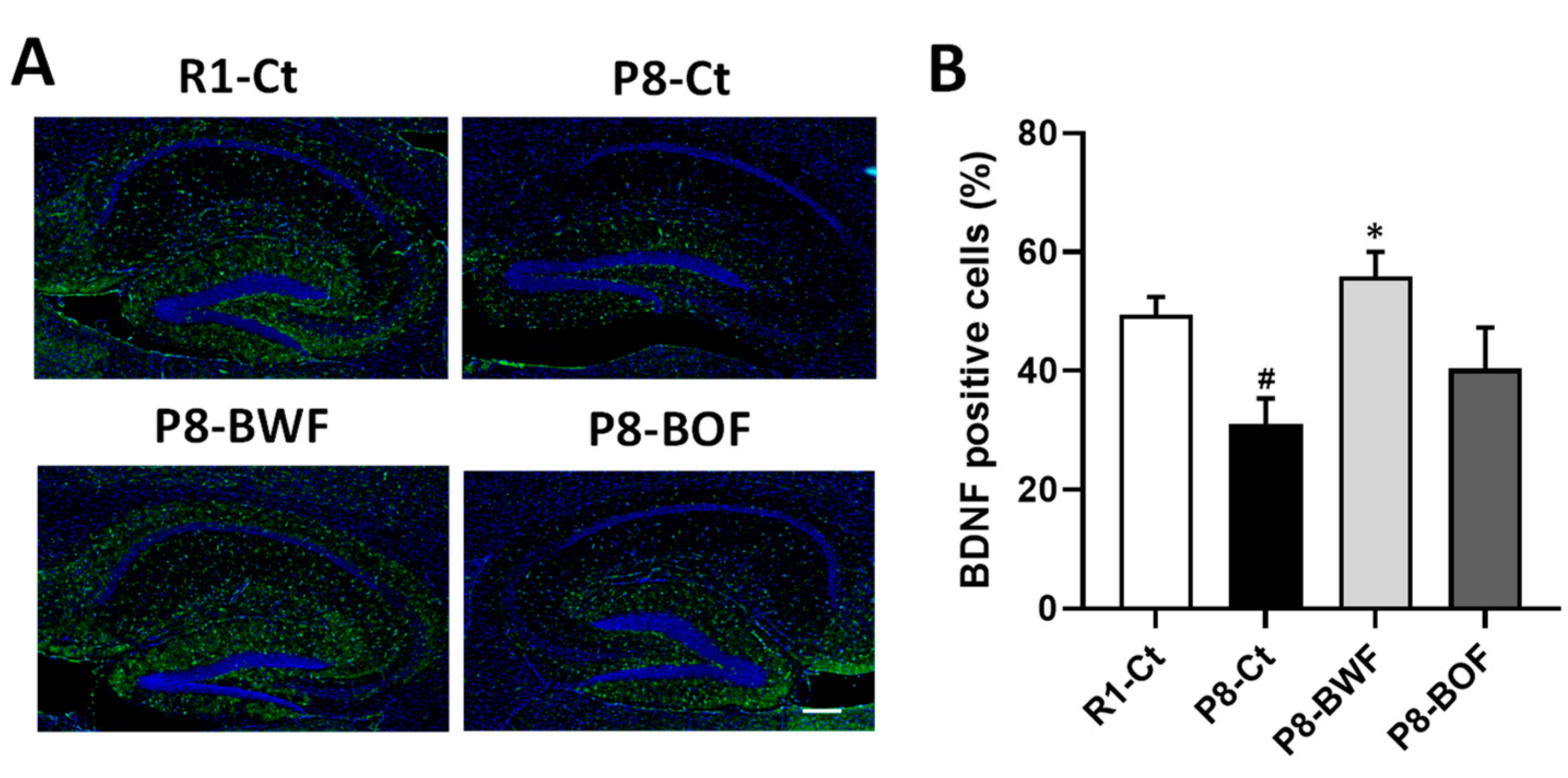
3.3. Regulatory Effects of Buckwheat Flour on Neurons and Neuroplasticity-Related Proteins
3.4. Effects of Buckwheat Starch on the Cognitive Function and Hippocampal BDNF Expression in SAMP8 Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wijngaard, H.; Arendt, E.K. Buckwheat. Cereal Chem. 2006, 83, 391–401. [Google Scholar] [CrossRef]
- Ikeda, K. Buckwheat composition, chemistry, and processing. Adv. Food Nutr. Res. 2002, 44, 395–434. [Google Scholar] [PubMed]
- Giménez-Bastida, J.A.; Piskuła, M.; Zieliński, H. Recent advances in development of gluten-free buckwheat products. Trends Food Sci. Technol. 2015, 44, 58–65. [Google Scholar] [CrossRef]
- Skrabanja, V.; Kreft, I.; Golob, T.; Modic, M.; Ikeda, S.; Ikeda, K.; Kreft, S.; Bonafaccia, G.; Knapp, M.; Kosmelj, K. Nutrient content in buckwheat milling fractions. Cereal Chem. 2004, 81, 172–176. [Google Scholar] [CrossRef]
- Van Hung, P.; Maeda, T.; Tsumori, R.; Morita, N. Characteristics of fractionated flours from whole buckwheat grain using a gradual milling system and their application for noodle making. J. Sci. Food Agric. 2007, 87, 2823–2829. [Google Scholar] [CrossRef]
- Krkošková, B.; Mrazova, Z. Prophylactic components of buckwheat. Food Res. Int. 2005, 38, 561–568. [Google Scholar] [CrossRef]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T. Senescence-accelerated mouse (SAM) with special references to neurodegeneration models, SAMP8 and SAMP10 mice. Neurochem. Res. 2009, 34, 639–659. [Google Scholar] [CrossRef]
- Akiguchi, I.; Pallàs, M.; Budka, H.; Akiyama, H.; Ueno, M.; Han, J.; Yagi, H.; Nishikawa, T.; Chiba, Y.; Sugiyama, H. SAMP8 mice as a neuropathological model of accelerated brain aging and dementia: Toshio Takeda’s legacy and future directions. Neuropathology 2017, 37, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Katayama, S.; Imai, R.; Sugiyama, H.; Nakamura, S. Oral administration of soy peptides suppresses cognitive decline by induction of neurotrophic factors in SAMP8 mice. J. Agric. Food Chem. 2014, 62, 3563–3569. [Google Scholar] [CrossRef]
- Corpuz, H.M.; Arimura, M.; Chawalitpong, S.; Miyazaki, K.; Sawaguchi, M.; Nakamura, S.; Katayama, S. Oral administration of okara soybean by-product attenuates cognitive impairment in a mouse model of accelerated aging. Nutrients 2019, 11, 2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katayama, S.; Nakamura, S. Emerging roles of bioactive peptides on brain health promotion. Int. J. Food Sci. Technol. 2019, 54, 1949–1955. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Godos, J.; Privitera, A.; Lanza, G.; Castellano, S.; Chillemi, A.; Bruni, O.; Ferri, R.; Caraci, F.; Grosso, G. Phenolic acids and prevention of cognitive decline: Polyphenols with a neuroprotective role in cognitive disorders and Alzheimer’s disease. Nutrients 2022, 14, 819. [Google Scholar] [CrossRef] [PubMed]
- Caracciolo, B.; Xu, W.; Collins, S.; Fratiglioni, L. Cognitive decline, dietary factors and gut–brain interactions. Mech. Ageing Dev. 2014, 136, 59–69. [Google Scholar] [CrossRef]
- Corpuz, H.M.; Ichikawa, S.; Arimura, M.; Mihara, T.; Kumagai, T.; Mitani, T.; Nakamura, S.; Katayama, S. Long-Term Diet Supplementation with Lactobacillus paracasei K71 Prevents Age-Related Cognitive Decline in Senescence-Accelerated Mouse Prone 8. Nutrients 2018, 10, 762. [Google Scholar] [CrossRef] [Green Version]
- Enogieru, A.B.; Haylett, W.; Hiss, D.C.; Bardien, S.; Ekpo, O.E. Rutin as a potent antioxidant: Implications for neurodegenerative disorders. Oxid. Med. Cell. Longev. 2018, 2018, 6241017. [Google Scholar] [CrossRef]
- von Bohlen und Halbach, O.; von Bohlen und Halbach, V. BDNF effects on dendritic spine morphology and hippocampal function. Cell Tissue Res. 2018, 373, 729–741. [Google Scholar] [CrossRef]
- Kedrov, A.V.; Durymanov, M.; Anokhin, K.V. The Arc gene: Retroviral heritage in cognitive functions. Neurosci. Biobehav. Rev. 2019, 99, 275–281. [Google Scholar] [CrossRef]
- Han, K.; Kim, E. Synaptic adhesion molecules and PSD-95. Prog. Neurobiol. 2008, 84, 263–283. [Google Scholar] [CrossRef]
- Ying, S.-W.; Futter, M.; Rosenblum, K.; Webber, M.J.; Hunt, S.P.; Bliss, T.V.; Bramham, C.R. Brain-derived neurotrophic factor induces long-term potentiation in intact adult hippocampus: Requirement for ERK activation coupled to CREB and upregulation of Arc synthesis. J. Neurosci. 2002, 22, 1532–1540. [Google Scholar] [CrossRef] [Green Version]
- Vecsey, C.G.; Hawk, J.D.; Lattal, K.M.; Stein, J.M.; Fabian, S.A.; Attner, M.A.; Cabrera, S.M.; McDonough, C.B.; Brindle, P.K.; Abel, T. Histone deacetylase inhibitors enhance memory and synaptic plasticity via CREB: CBP-dependent transcriptional activation. J. Neurosci. 2007, 27, 6128–6140. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Ganai, S.; Ramadoss, M.; Mahadevan, V. Histone Deacetylase (HDAC) Inhibitors-emerging roles in neuronal memory, learning, synaptic plasticity and neural regeneration. Curr. Neuropharmacol. 2016, 14, 55–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berding, K.; Carbia, C.; Cryan, J.F. Going with the grain: Fiber, cognition, and the microbiota-gut-brain-axis. Exp. Biol. Med. 2021, 246, 796–811. [Google Scholar] [CrossRef]
- van Hylckama Vlieg, J.E.; Rademaker, J.L.; Bachmann, H.; Molenaar, D.; Kelly, W.J.; Siezen, R.J. Natural diversity and adaptive responses of Lactococcus lactis. Curr. Opin. Biotechnol. 2006, 17, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, J.B.; Spiazzi, C.C.; Bicca, D.F.; Rodrigues, J.F.; Sehn, C.P.; da Silva, W.P.; Cibin, F.W.S. Beneficial effects of Lactococcus lactis subsp. cremoris LL95 treatment in an LPS-induced depression-like model in mice. Behav. Brain Res. 2022, 426, 113847. [Google Scholar]
- Gao, K.; Farzi, A.; Ke, X.; Yu, Y.; Chen, C.; Chen, S.; Yu, T.; Wang, H.; Li, Y. Oral administration of Lactococcus lactis WHH2078 alleviates depressive and anxiety symptoms in mice with induced chronic stress. Food Funct. 2022, 13, 957–969. [Google Scholar] [CrossRef]
- Lim, S.-M.; Lee, N.-K.; Paik, H.-D. Potential neuroprotective effects of heat-killed Lactococcus lactis KC24 using SH-SY5Y cells against oxidative stress induced by hydrogen peroxide. Food Sci. Biotechnol. 2020, 29, 1735–1740. [Google Scholar] [CrossRef]
- Sherwin, E.; Sandhu, K.V.; Dinan, T.G.; Cryan, J.F. May the force be with you: The light and dark sides of the microbiota–gut–brain axis in neuropsychiatry. CNS Drugs 2016, 30, 1019–1041. [Google Scholar] [CrossRef] [Green Version]
- Kato, N.; Kayashita, J.; Sasaki, M. Physiological functions of buckwheat protein and sericin as resistant proteins. J. Jpn. Soc. Food Nutr. 2000, 53, 71–75. [Google Scholar] [CrossRef]
- Ravachol, J.; Borne, R.; Meynial-Salles, I.; Soucaille, P.; Tardif, C.; Fierobe, H.-P. Combining free and aggregated cellulolytic systems in the cellulosome-producing bacterium Ruminiclostridium cellulolyticum. Biotechnol. Biofuels 2015, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.-D.; Cai, L.; Ni, Y.-S.; Tian, S.-Y.; Lu, Y.-Q.; Wang, L.-N.; Chen, L.-L.; Ma, W.-Y.; Deng, S.-P. Comparisons of effects on intestinal short-chain fatty acid concentration after exposure of two glycosidase inhibitors in mice. Biol. Pharm. Bull. 2018, 41, 1024–1033. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Su, L.; Zhu, H.; Li, X.; Guo, Y.; Du, X.; Zhang, L.; Qin, C. Gut microbiota regulation and their implication in the development of neurodegenerative disease. Microorganisms 2021, 9, 2281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Buckwheat starch: Structures, properties, and applications. Trends Food Sci. Technol. 2016, 49, 121–135. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (g/100 g) | Lipid (g/100 g) | Carbohydrate (g/100 g) | Dietary Fiber (g/100 g) | Rutin (mg/100 g) | |
|---|---|---|---|---|---|
| BWF | 13.8 | 3.3 | 68.1 | 4.5 | 6.6 |
| BOF | 33.6 | 7.1 | 44.3 | 16.7 | 13.0 |
| TWF | 12.8 | 3.3 | 69.4 | 3.6 | 808 |
| WF | 11.5 | 1.4 | 73.6 | 2.6 | 0.037 |
| RF | 6.2 | 0.9 | 82.0 | 0.9 | ND |
| Ingredients (%) | Standard Diet | BWF | BOF | TWF | Rutin | WF | RF |
|---|---|---|---|---|---|---|---|
| Casein | 14.00 | 13.31 | 12.32 | 13.36 | 14.00 | 13.43 | 13.69 |
| L-cystine | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 |
| Corn starch | 46.57 | 42.65 | 44.44 | 42.55 | 46.57 | 42.34 | 41.97 |
| α-cornstarch | 15.50 | 15.50 | 15.50 | 15.50 | 15.50 | 15.50 | 15.50 |
| Sucrose | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Soybean oil | 4.00 | 3.84 | 3.65 | 3.84 | 4.00 | 3.93 | 3.96 |
| Cellulose | 5.00 | 4.78 | 4.17 | 4.82 | 5.00 | 4.87 | 4.96 |
| Mineral mix | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 |
| Vitamin mix | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline bitartrate | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| tert-butylhydroquinone | 0.0008 | 0.0008 | 0.0008 | 0.0008 | 0.0008 | 0.0008 | 0.0008 |
| Tested sample | 0.00 | 5.00 | 5.00 | 5.00 | 0.05 | 5.00 | 5.00 |
| Ingredients (%) | Standard Diet | BWF | BS | WS | RS |
|---|---|---|---|---|---|
| Casein | 14.00 | 13.31 | 13.97 | 13.99 | 13.99 |
| L-cystine | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 |
| Corn starch | 46.57 | 42.65 | 42.64 | 42.61 | 42.60 |
| α-cornstarch | 15.50 | 15.50 | 15.50 | 15.50 | 15.50 |
| Sucrose | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Soybean oil | 4.00 | 3.84 | 3.98 | 3.98 | 3.98 |
| Cellulose | 5.00 | 4.78 | 4.98 | 5.00 | 5.00 |
| Mineral mix | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 |
| Vitamin mix | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline bitartrate | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| tert-butylhydroquinone | 0.0008 | 0.0008 | 0.0008 | 0.0008 | 0.0008 |
| Tested sample | 0.00 | 5.00 | 4.00 | 4.00 | 4.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katayama, S.; Okahata, C.; Onozato, M.; Minami, T.; Maeshima, M.; Ogihara, K.; Yamazaki, S.; Takahashi, Y.; Nakamura, S. Buckwheat Flour and Its Starch Prevent Age-Related Cognitive Decline by Increasing Hippocampal BDNF Production in Senescence-Accelerated Mouse Prone 8 Mice. Nutrients 2022, 14, 2708. https://doi.org/10.3390/nu14132708
Katayama S, Okahata C, Onozato M, Minami T, Maeshima M, Ogihara K, Yamazaki S, Takahashi Y, Nakamura S. Buckwheat Flour and Its Starch Prevent Age-Related Cognitive Decline by Increasing Hippocampal BDNF Production in Senescence-Accelerated Mouse Prone 8 Mice. Nutrients. 2022; 14(13):2708. https://doi.org/10.3390/nu14132708
Chicago/Turabian StyleKatayama, Shigeru, Chizuru Okahata, Masashi Onozato, Takaaki Minami, Masanaga Maeshima, Kazuaki Ogihara, Shinya Yamazaki, Yuta Takahashi, and Soichiro Nakamura. 2022. "Buckwheat Flour and Its Starch Prevent Age-Related Cognitive Decline by Increasing Hippocampal BDNF Production in Senescence-Accelerated Mouse Prone 8 Mice" Nutrients 14, no. 13: 2708. https://doi.org/10.3390/nu14132708