

Protective Effects of Resveratrol on Adolescent Social Isolation-Induced Anxiety-Like Behaviors via Modulating Nucleus Accumbens Spine Plasticity and Mitochondrial Function in Female Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Animal Experiment
2.3. Drug Treatment
2.4. Sucrose Preference Test (SPT)
2.5. Open Field Test (OFT)
2.6. Elevated plus Maze (EPM)
2.7. Forced Swim Test (FST)
2.8. Social Interaction Test (SIT)
2.9. Viral Constructs and Microinjection
2.10. Dendritic Spine Analysis
2.11. RNA-Seq Analysis
2.12. Quantitative Real-Time PCR (qPCR)
2.13. Transmission Electron Microscopy
2.14. Cytochrome-c Oxidase (COX) Activity and Adenosine Triphosphate Content
2.15. Statistical Analysis
3. Results
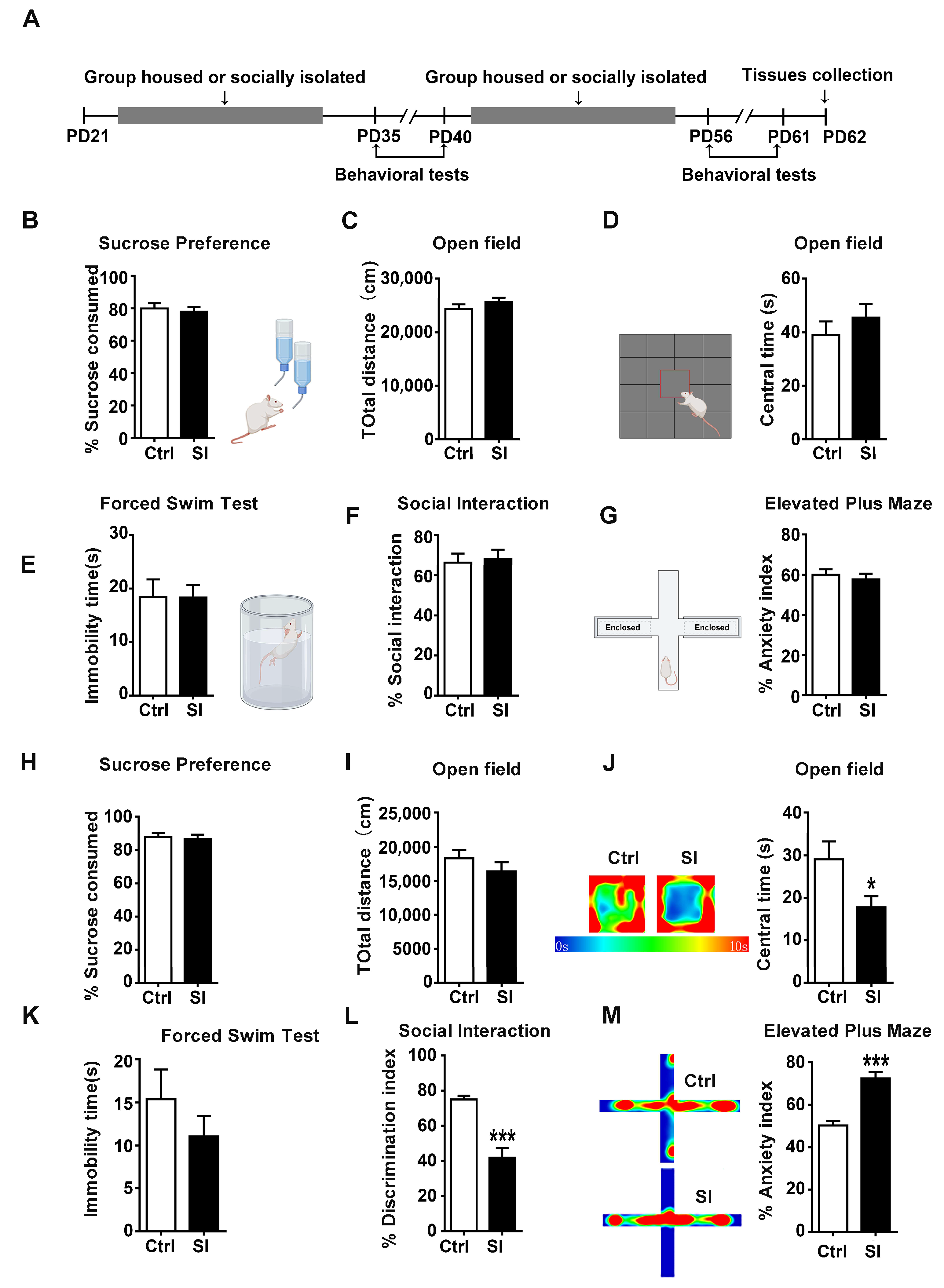
3.1. Social Isolation Induced Anxiety-Like Behavior and Social Dysfunction but Not Depression-Like Behavior at Late Adolescence in Female Rats
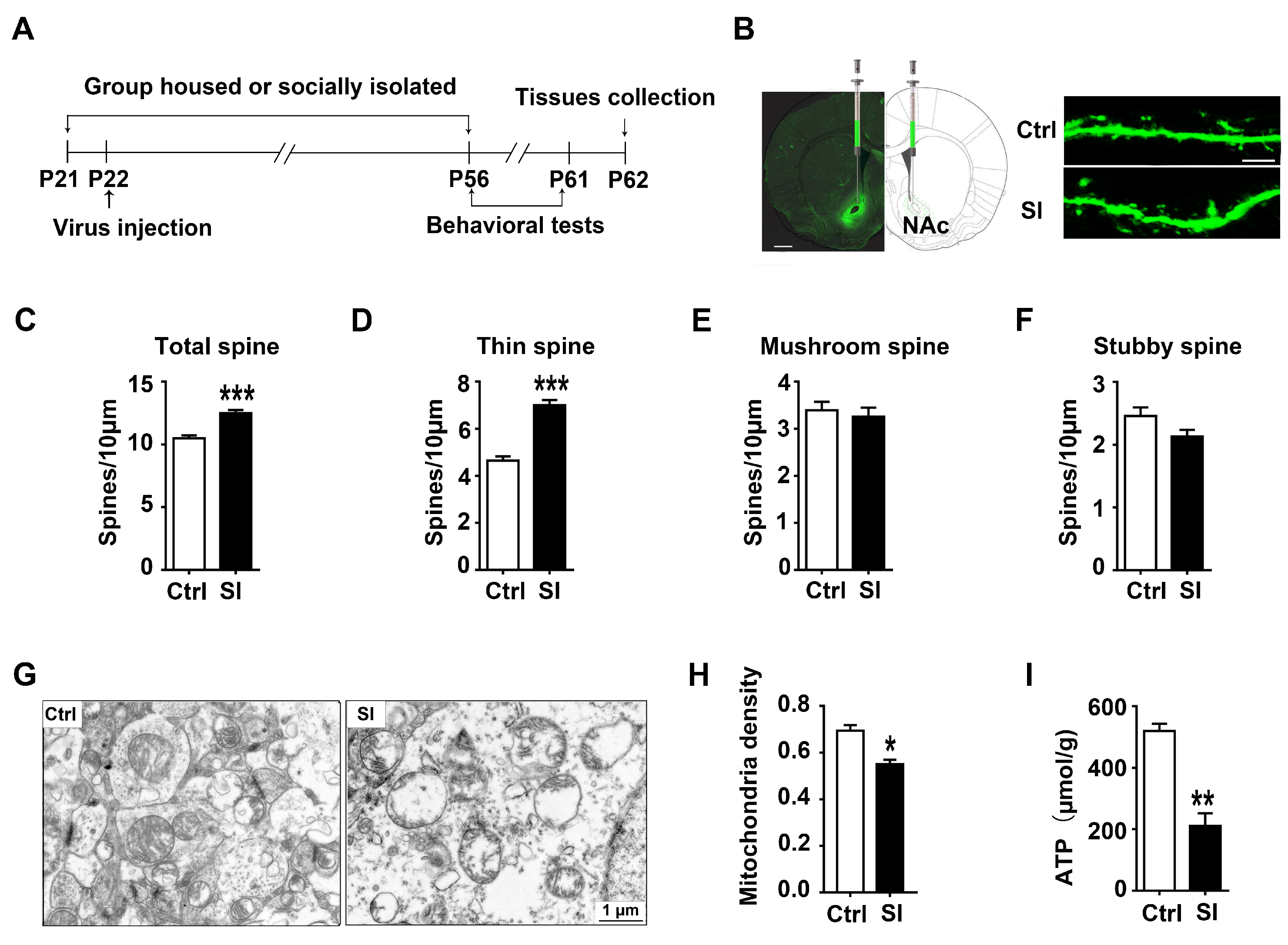
3.2. The Effect of Adolescent Social Isolation on Spine Remodelling and Mitochondrial Function in the NAc
3.3. The Effect of Adolescent Social Isolation on the Transcriptional Patterns in the NAc
3.4. The Protective Effects of Resveratrol on Adolescent Social-Isolation-Induced Behavioral Abnormalities in Female Rats
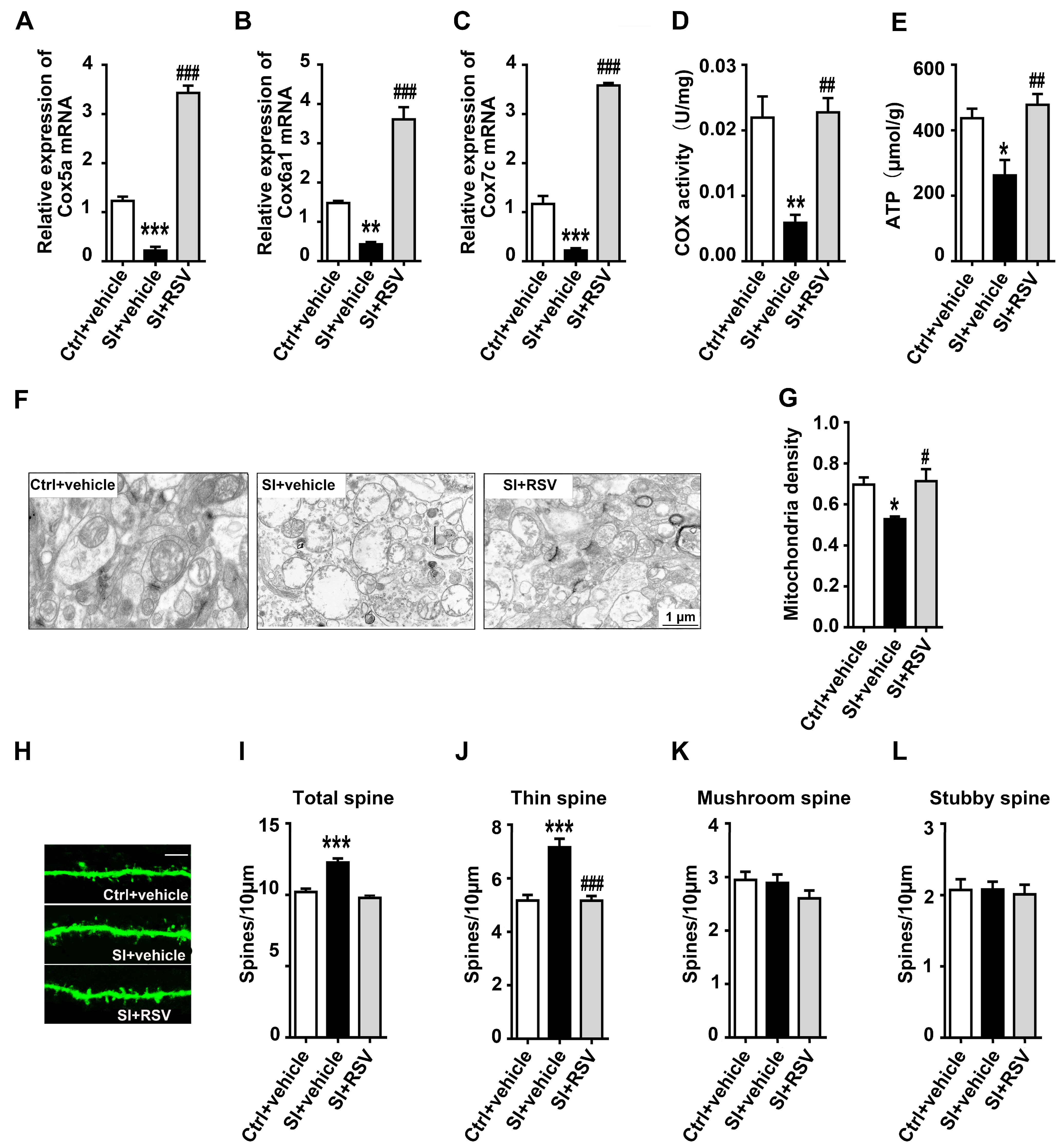
3.5. The Protective Effects of Resveratrol on Adolescent Social Isolation-Induced Mitochondrial and Spine Abnormalities in Female Rats
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanson, J.L.; Williams, A.V.; Bangasser, D.A.; Pena, C.J. Impact of Early Life Stress on Reward Circuit Function and Regulation. Front. Psychiatry 2021, 12, 744690. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Mental Disorders Collaborators. Global prevalence and burden of depressive and anxiety disorders in 204 countries and territories in 2020 due to the COVID-19 pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef]
- Magson, N.R.; Freeman, J.Y.A.; Rapee, R.M.; Richardson, C.E.; Oar, E.L.; Fardouly, J. Risk and Protective Factors for Prospective Changes in Adolescent Mental Health during the COVID-19 Pandemic. J. Youth Adolesc. 2021, 50, 44–57. [Google Scholar] [CrossRef]
- Walker, D.M.; Cunningham, A.M.; Gregory, J.K.; Nestler, E.J. Long-Term Behavioral Effects of Post-weaning Social Isolation in Males and Females. Front. Behav. Neurosci. 2019, 13, 66. [Google Scholar] [CrossRef] [PubMed]
- Meherali, S.; Punjani, N.; Louie-Poon, S.; Abdul Rahim, K.; Das, J.K.; Salam, R.A.; Lassi, Z.S. Mental Health of Children and Adolescents Amidst COVID-19 and Past Pandemics: A Rapid Systematic Review. Int. J. Environ. Res. Public Health 2021, 18, 3432. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, A.N.; Rose, J.H.; Weiner, J.L.; Jones, S.R. Early-Life Social Isolation Stress Increases Kappa Opioid Receptor Responsiveness and Downregulates the Dopamine System. Neuropsychopharmacology 2016, 41, 2263–2274. [Google Scholar] [CrossRef]
- Novoa, J.; Rivero, C.J.; Perez-Cardona, E.U.; Freire-Arvelo, J.A.; Zegers, J.; Yarur, H.E.; Santiago-Marerro, I.G.; Agosto-Rivera, J.L.; González-Pérez, J.L.; Gysling, K.; et al. Social isolation of adolescent male rats increases anxiety and K+-induced dopamine release in the nucleus accumbens: Role of CRF-R1. Eur. J. Neurosci. 2021, 54, 4888–4905. [Google Scholar] [CrossRef] [PubMed]
- Cuesta, S.; Funes, A.; Pacchioni, A.M. Social Isolation in Male Rats During Adolescence Inhibits the Wnt/beta-Catenin Pathway in the Prefrontal Cortex and Enhances Anxiety and Cocaine-Induced Plasticity in Adulthood. Neurosci. Bull. 2020, 36, 611–624. [Google Scholar] [CrossRef]
- Walker, D.M.; Zhou, X.; Cunningham, A.M.; Lipschultz, A.P.; Ramakrishnan, A.; Cates, H.M.; Bagot, R.C.; Shen, L.; Zhang, B.; Nestler, E.J. Sex-Specific Transcriptional Changes in Response to Adolescent Social Stress in the Brain’s Reward Circuitry. Biol. Psychiatry 2022, 91, 118–128. [Google Scholar] [CrossRef]
- Kinley, B.L.; Kyne, R.F.; Lawton-Stone, T.S.; Walker, D.M.; Paul, M.J. Long-term consequences of peri-adolescent social isolation on social preference, anxiety-like behaviour, and vasopressin neural circuitry of male and female rats. Eur. J. Neurosci. 2021, 54, 7790–7804. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.; Sadananda, M. Anxiety- and depressive-like profiles during early- and mid-adolescence in the female Wistar Kyoto rat. Int. J. Dev. Neurosci. 2017, 56, 18–26. [Google Scholar] [CrossRef]
- Zhao, J.; Ying, L.; Liu, Y.; Liu, N.; Tu, G.; Zhu, M.; Wu, Y.; Xiao, B.; Ye, L.; Li, J.; et al. Different roles of Rac1 in the acquisition and extinction of methamphetamine-associated contextual memory in the nucleus accumbens. Theranostics 2019, 9, 7051–7071. [Google Scholar] [CrossRef] [PubMed]
- Birnie, M.T.; Kooiker, C.L.; Short, A.K.; Bolton, J.L.; Chen, Y.; Baram, T.Z. Plasticity of the Reward Circuitry After Early-Life Adversity: Mechanisms and Significance. Biol. Psychiatry 2020, 87, 875–884. [Google Scholar] [CrossRef]
- Peña, C.J.; Smith, M.; Ramakrishnan, A.; Cates, H.M.; Bagot, R.C.; Kronman, H.G.; Patel, B.; Chang, A.B.; Purushothaman, I.; Dudley, J.; et al. Early life stress alters transcriptomic patterning across reward circuitry in male and female mice. Nat. Commun. 2019, 10, 5098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebara, E.; Zanoletti, O.; Ghosal, S.; Grosse, J.; Schneider, B.L.; Knott, G.; Astori, S.; Sandi, C. Mitofusin-2 in the Nucleus Accumbens Regulates Anxiety and Depression-like Behaviors Through Mitochondrial and Neuronal Actions. Biol. Psychiatry 2021, 89, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; McGee, S.; Czeczor, J.K.; Walker, A.J.; Kale, R.P.; Kouzani, A.Z.; Walder, K.; Berk, M.; Tye, S.J. Nucleus accumbens deep-brain stimulation efficacy in ACTH-pretreated rats: Alterations in mitochondrial function relate to antidepressant-like effects. Transl. Psychiatry 2016, 6, e842. [Google Scholar] [CrossRef] [Green Version]
- Van Der Kooij, M.A.; Hollis, F.; Lozano, L.; Zalachoras, I.; Abad, S.; Zanoletti, O.; Grosse, J.; De Suduiraut, I.G.; Canto, C.; Sandi, C. Diazepam actions in the VTA enhance social dominance and mitochondrial function in the nucleus accumbens by activation of dopamine D1 receptors. Mol. Psychiatry 2018, 23, 569–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, T.C. Editorial: Toward Neurobiological-Based Treatments of Depression and Anxiety: A Potential Case for the Nucleus Accumbens. J. Am. Acad. Child Adolesc. Psychiatry 2022, 61, 136–138. [Google Scholar] [CrossRef]
- Deutschmann, A.U.; Kirkland, J.M.; Briand, L.A. Adolescent social isolation induced alterations in nucleus accumbens glutamate signalling. Addict. Biol. 2022, 27, e13077. [Google Scholar] [CrossRef] [PubMed]
- Grinan-Ferre, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillon, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C.; et al. The pleiotropic neuroprotective effects of resveratrol in cognitive decline and Alzheimer’s disease pathology: From antioxidant to epigenetic therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef]
- Sahiner, M.; Yilmaz, A.S.; Gungor, B.; Ayoubi, Y.; Sahiner, N. Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources. Molecules 2022, 27, 6225. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, W.; Li, Y.; Xu, W.; Yuan, Y.; Zheng, V.; Zhang, H.; O’Donnell, J.M.; Xu, Y.; Yin, X. The antidepressant- and anxiolytic-like effects of resveratrol: Involvement of phosphodiesterase-4D inhibition. Neuropharmacology 2019, 153, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Ma, Y.; Wang, D.; Zhao, W.; Hu, X.; Chen, F.; Zhao, X. Protective Effects of Dietary Resveratrol against Chronic Low-Grade Inflammation Mediated through the Gut Microbiota in High-Fat Diet Mice. Nutrients 2022, 14, 1994. [Google Scholar] [CrossRef] [PubMed]
- Sahin, T.D.; Gocmez, S.S.; Eraldemir, F.C.; Utkan, T. Anxiolytic-Like and Antidepressant-Like Effects of Resveratrol in Streptozotocin-Induced Diabetic Rats. Noro Psikiyatr. Ars. 2019, 56, 144–149. [Google Scholar]
- Moore, A.; Beidler, J.; Hong, M.Y. Resveratrol and Depression in Animal Models: A Systematic Review of the Biological Mechanisms. Molecules 2018, 23, 2197. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Ma, Y.; Zhang, R.; Zhong, H.; Wang, L.; Zhao, J.; Yang, L.; Fan, X. Resveratrol ameliorates estrogen deficiency-induced depression- and anxiety-like behaviors and hippocampal inflammation in mice. Psychopharmacology 2019, 236, 1385–1399. [Google Scholar] [CrossRef]
- Shen, J.-D.; Zhang, Y.-W.; Wang, B.-Y.; Bai, L.; Lu, S.-F.; Zhu, L.-L.; Bai, M.; Li, Y.-C.; Xu, E.-P. Effects of resveratrol on the levels of ATP, 5-HT and GAP-43 in the hippocampus of mice exposed to chronic unpredictable mild stress. Neurosci. Lett. 2020, 735, 135232. [Google Scholar] [CrossRef]
- Finnell, J.E.; Lombard, C.M.; Melson, M.N.; Singh, N.P.; Nagarkatti, M.; Nagarkatti, P.; Fadel, J.R.; Wood, C.S.; Wood, S.K. The protective effects of resveratrol on social stress-induced cytokine release and depressive-like behavior. Brain Behav. Immun. 2017, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Hollis, F.; Pope, B.S.; Gorman-Sandler, E.; Wood, S.K. Neuroinflammation and Mitochondrial Dysfunction Link Social Stress to Depression. Curr. Top. Behav. Neurosci. 2022, 54, 59–93. [Google Scholar]
- Taheri Zadeh, Z.; Rahmani, S.; Alidadi, F.; Joushi, S.; Esmaeilpour, K. Depresssion, anxiety and other cognitive consequences of social isolation: Drug and non-drug treatments. Int. J. Clin. Pract. 2021, 75, e14949. [Google Scholar] [CrossRef]
- Burke, A.R.; McCormick, C.M.; Pellis, S.M.; Lukkes, J.L. Impact of adolescent social experiences on behavior and neural circuits implicated in mental illnesses. Neurosci. Biobehav. Rev. 2017, 76, 280–300. [Google Scholar] [CrossRef] [PubMed]
- Lukkes, J.L.; Watt, M.J.; Lowry, C.A.; Forster, G.L. Consequences of post-weaning social isolation on anxiety behavior and related neural circuits in rodents. Front. Behav. Neurosci. 2009, 3, 18. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Cao, K.; Cui, S.; Cui, Y.; Mo, H.; Wen, W.; Dong, Z.; Lin, H.; Bai, S.; Yang, L.; et al. SiNiSan ameliorates depression-like behavior in rats by enhancing synaptic plasticity via the CaSR-PKC-ERK signaling pathway. Biomed. Pharmacother. 2020, 124, 109787. [Google Scholar] [CrossRef]
- Karkhanis, A.N.; Locke, J.L.; McCool, B.A.; Weiner, J.L.; Jones, S.R. Social isolation rearing increases nucleus accumbens dopamine and norepinephrine responses to acute ethanol in adulthood. Alcohol Clin. Exp. Res. 2014, 38, 2770–2779. [Google Scholar] [CrossRef] [Green Version]
- Schiavi, S.; Iezzi, D.; Manduca, A.; Leone, S.; Melancia, F.; Carbone, C.; Petrella, M.; Mannaioni, G.; Masi, A.; Trezza, V. Reward-Related Behavioral, Neurochemical and Electrophysiological Changes in a Rat Model of Autism Based on Prenatal Exposure to Valproic Acid. Front. Cell. Neurosci. 2019, 13, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, G.; Ying, L.; Ye, L.; Zhao, J.; Liu, N.; Li, J.; Liu, Y.; Zhu, M.; Wu, Y.; Xiao, B.; et al. Dopamine D(1) and D(2) Receptors Differentially Regulate Rac1 and Cdc42 Signaling in the Nucleus Accumbens to Modulate Behavioral and Structural Plasticity After Repeated Methamphetamine Treatment. Biol. Psychiatry 2019, 86, 820–835. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Ma, C.; Lu, L.; Zhang, L.; Luo, X. Determination of dietary iron requirements by full expression of iron-containing cytochrome c oxidase in the heart of broilers from 22 to 42 d of age. Br. J. Nutr. 2017, 118, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Sun, H.; Li, K.; Shang, L.; Liang, X.; Yang, H.; Dong, Z.; Xiaokereti, J.; Shang, S.; Zhou, Q.; et al. Cholinergic Elicitation Prevents Ventricular Remodeling via Alleviations of Myocardial Mitochondrial Injury Linked to Inflammation in Ischemia-Induced Chronic Heart Failure Rats. Mediat. Inflamm. 2021, 2021, 4504431. [Google Scholar] [CrossRef]
- Moreno-Martinez, S.; Tendilla-Beltran, H.; Sandoval, V.; Flores, G.; Terron, J.A. Chronic restraint stress induces anxiety-like behavior and remodeling of dendritic spines in the central nucleus of the amygdala. Behav. Brain Res. 2022, 416, 113523. [Google Scholar] [CrossRef]
- Amiri, S.; Amini-Khoei, H.; Haj-Mirzaian, A.; Rahimi-Balaei, M.; Naserzadeh, P.; Dehpour, A.R.; Mehr, S.E.; Hosseini, M.-J. Tropisetron attenuated the anxiogenic effects of social isolation by modulating nitrergic system and mitochondrial function. Biochim. Biophys. Acta 2015, 1850, 2464–2475. [Google Scholar] [CrossRef]
- Cunatova, K.; Reguera, D.P.; Houstek, J.; Mracek, T.; Pecina, P. Role of cytochrome c oxidase nuclear-encoded subunits in health and disease. Physiol. Res. 2020, 69, 947–965. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Wang, Y.; Zhang, H.; Li, M.; Chen, G.; Hao, J.; Xie, M. Involvement of purinergic P2Y1R in antidepressant-like effects of electroacupuncture treatment on social isolation stress mice. Purinergic Signal. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-J.; Shwani, T.; Liu, J.; Zhong, P.; Yang, F.; Schatz, K.; Zhang, F.; Pralle, A.; Yan, Z. Molecular and cellular mechanisms for differential effects of chronic social isolation stress in males and females. Mol. Psychiatry 2022, 27, 3056–3068. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Flashner, B.; Chiu, M.; ver Hoeve, E.; Luz, S.; Bhatnagar, S. Social isolation in adolescence alters behaviors in the forced swim and sucrose preference tests in female but not in male rats. Physiol. Behav. 2012, 105, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.P.; McGovern, A.J.; Melo, T.G.; Barron, A.; Nolan, Y.M.; O’Leary, O.F. Juvenile stress exerts sex-independent effects on anxiety, antidepressant-like behaviours and dopaminergic innervation of the prelimbic cortex in adulthood and does not alter hippocampal neurogenesis. Behav. Brain Res. 2022, 421, 113725. [Google Scholar] [CrossRef]
- Logue, J.; Schoepfer, K.; Guerrero, A.B.; Zhou, Y.; Kabbaj, M. Sex-specific effects of social isolation stress and ketamine on hippocampal plasticity. Neurosci. Lett. 2022, 766, 136301. [Google Scholar] [CrossRef]
- Chen, J.-J.; Shen, J.-X.; Yu, Z.-H.; Pan, C.; Han, F.; Zhu, X.-L.; Xu, H.; Xu, R.-T.; Wei, T.-Y.; Lu, Y.-P. The Antidepressant Effects of Resveratrol are Accompanied by the Attenuation of Dendrite/Dendritic Spine Loss and the Upregulation of BDNF/p-cofilin1 Levels in Chronic Restraint Mice. Neurochem. Res. 2021, 46, 660–674. [Google Scholar] [CrossRef]
- Parekh, P.K.; Johnson, S.B.; Liston, C. Synaptic Mechanisms Regulating Mood State Transitions in Depression. Annu. Rev. Neurosci. 2022, 45, 581–601. [Google Scholar] [CrossRef]
- Muir, J.; Tse, Y.C.; Iyer, E.S.; Biris, J.; Cvetkovska, V.; Lopez, J.; Bagot, R.C. Ventral Hippocampal Afferents to Nucleus Accumbens Encode Both Latent Vulnerability and Stress-Induced Susceptibility. Biol. Psychiatry 2020, 88, 843–854. [Google Scholar] [CrossRef]
- Bagot, R.C.; Parise, E.M.; Pena, C.J.; Zhang, H.X.; Maze, I.; Chaudhury, D.; Persaud, B.; Cachope, R.; Bolanos-Guzmán, C.A.; Cheer, J.F.; et al. Ventral hippocampal afferents to the nucleus accumbens regulate susceptibility to depression. Nat. Commun. 2015, 6, 7062. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Li, N.; Xue, X.; Shao, F.; Wang, W. Early social isolation disrupts latent inhibition and increases dopamine D2 receptor expression in the medial prefrontal cortex and nucleus accumbens of adult rats. Brain Res. 2012, 1447, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Fosnocht, A.Q.; Lucerne, K.E.; Ellis, A.S.; Olimpo, N.A.; Briand, L.A. Adolescent social isolation increases cocaine seeking in male and female mice. Behav. Brain Res. 2019, 359, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Chen, S.; Gao, F.; Hu, B.; Wang, Y.; Ni, S.; Kou, H.; Song, Z.; Qing, X.; Wang, S.; et al. Resveratrol-enhanced SIRT1-mediated osteogenesis in porous endplates attenuates low back pain and anxiety behaviors. FASEB J. 2021, 35, e21414. [Google Scholar] [CrossRef] [PubMed]
- Lee, I. Regulation of Cytochrome c Oxidase by Natural Compounds Resveratrol, (−)-Epicatechin, and Betaine. Cells 2021, 10, 1346. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Ye, L.; Liu, Z.; Cui, Y.; Deng, D.; Bai, S.; Yang, L.; Shi, Y.; Liu, Z.; Zhang, R. Protective Effects of Resveratrol on Adolescent Social Isolation-Induced Anxiety-Like Behaviors via Modulating Nucleus Accumbens Spine Plasticity and Mitochondrial Function in Female Rats. Nutrients 2022, 14, 4542. https://doi.org/10.3390/nu14214542
Zhao J, Ye L, Liu Z, Cui Y, Deng D, Bai S, Yang L, Shi Y, Liu Z, Zhang R. Protective Effects of Resveratrol on Adolescent Social Isolation-Induced Anxiety-Like Behaviors via Modulating Nucleus Accumbens Spine Plasticity and Mitochondrial Function in Female Rats. Nutrients. 2022; 14(21):4542. https://doi.org/10.3390/nu14214542
Chicago/Turabian StyleZhao, Jinlan, Lihong Ye, Zuyi Liu, Yongfei Cui, Di Deng, Shasha Bai, Lei Yang, Yafei Shi, Zhongqiu Liu, and Rong Zhang. 2022. "Protective Effects of Resveratrol on Adolescent Social Isolation-Induced Anxiety-Like Behaviors via Modulating Nucleus Accumbens Spine Plasticity and Mitochondrial Function in Female Rats" Nutrients 14, no. 21: 4542. https://doi.org/10.3390/nu14214542