
Advances in Understanding the Human Gut Microbiota and Its Implication in Pediatric Celiac Disease—A Narrative Review
 , , , , , , ,
, , , , , , ,
Abstract
:1. Introduction
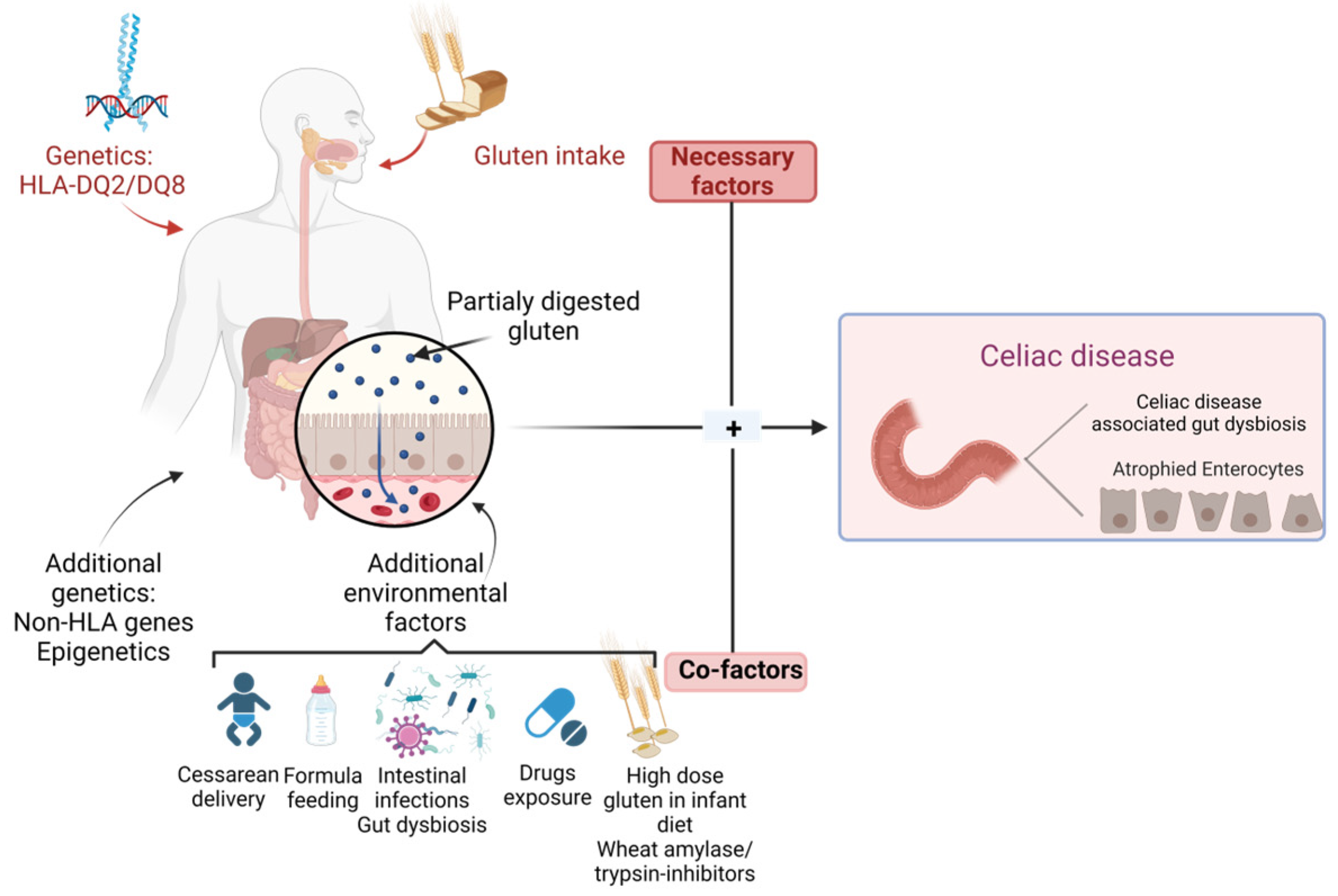
2. Genetic and Environmental Determinants of Celiac Disease
3. Gut Microbiota Profile in CD Progressors
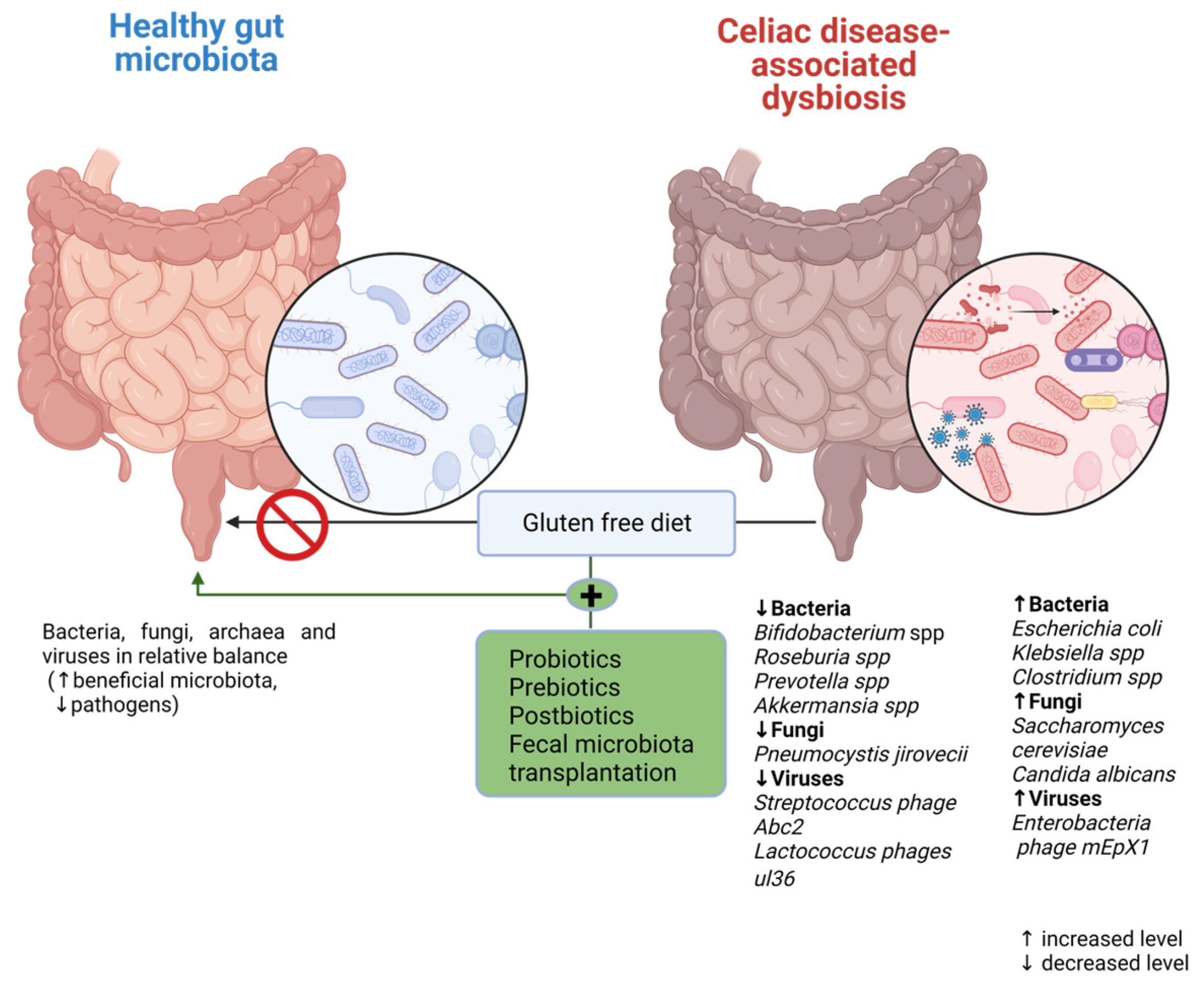
4. Gut Microbiota Profile at CD Onset
5. CD Prevention Strategies
6. Modulation of Gut Microbiota as a Potential Target in Celiac Disease
7. Limitations in the Study of Gut Microbiota and CD
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Withoff, S.; Li, Y.; Jonkers, I.; Wijmenga, C. Understanding Celiac Disease by Genomics. Trends Genet. 2016, 32, 295–308. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.; Singh, K.B.; Nnadozie, M.C.; Abdal, M.; Shrestha, N.; Abe, R.A.M.; Masroor, A.; Khorochkov, A.; Mohammed, L. New Evidence in the Pathogenesis of Celiac Disease and Type 1 Diabetes Mellitus: A Systematic Review. Cureus 2021, 13, e16721. [Google Scholar] [CrossRef] [PubMed]
- King, J.A.; Jeong, J.; Underwood, F.E.; Quan, J.; Panaccione, N.; Windsor, J.W.; Coward, S.; deBruyn, J.; Ronksley, P.E.; Shaheen, A.-A.; et al. Incidence of Celiac Disease Is Increasing Over Time: A Systematic Review and Meta-analysis. Am. J. Gastroenterol. 2020, 115, 507–525. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.E.; Morrison-Rees, S.; Thapar, N.; Benninga, M.A.; Borrelli, O.; Broekaert, I.; Dolinsek, J.; Martin-De-Carpi, J.; Mas, E.; Miele, E.; et al. Systematic review and meta-analysis: The incidence and prevalence of paediatric coeliac disease across Europe. Aliment. Pharmacol. Ther. 2021, 54, 109–128. [Google Scholar] [CrossRef] [PubMed]
- Van Berge-Henegouwen, G.P.; Mulder, C.J. Pioneer in the gluten free diet: Willem-Karel Dicke 1905–1962, over 50 years of gluten free diet. Gut 1993, 34, 1473–1475. [Google Scholar] [CrossRef]
- Paulley, J.W. Observation on the aetiology of idiopathic steatorrhoea; jejunal and lymphnode biopsies. BMJ 1954, 2, 1318–1321. [Google Scholar] [CrossRef]
- Berger, E.; Burgin-Wolff, A.; Freudenberg, E. Diagnostisehe Bewertung des Nachweises yon Gliadin-AntikSrpern bei Cöliakie. Klin. Wochenschr. 1964, 42, 788–790. [Google Scholar] [CrossRef]
- Falchuk, Z.M.; Rogentine, G.N.; Strober, W. Predominance of histocompatibility antigen HLA8 in patients with gluten-sensitive enteropathy. J. Clin. Investig. 1972, 51, 1602–1605. [Google Scholar] [CrossRef]
- Mearin, M.L.; Agardh, D.; Antunes, H.; Al-Toma, A.; Auricchio, R.; Castillejo, G.; Catassi, C.; Ciacci, C.; Discepolo, V.; Dolinsek, J.; et al. ESPGHAN Position Paper on Management and pandemic. Acta Paediatr. 2022, 111, 383–388. [Google Scholar]
- Meeuwisse, G.W. Diagnostic criteria in coeliac disease. Acta Paediatr. Scand. 1970, 59, 461–463. [Google Scholar]
- McNeish, A.S.; Harms, H.K.; Rey, J.; Shmerling, D.H.; Visakorpi, J.K.; A Walker-Smith, J. The diagnosis of coeliac disease. A commentary on the current practices of members of the European Society for Paediatric Gastroenterology and Nutrition (ESPGAN). Arch. Dis. Child. 1979, 54, 783–786. [Google Scholar] [CrossRef] [PubMed]
- Revised Criteria for Diagnosis of Coeliac Disease. Report of Working Group of European Society of Paediatric Gastroenterology and Nutrition. Arch. Dis. Child. 1990, 65, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Bozomitu, L.; Miron, I.; Raileanu, A.A.; Lupu, A.; Paduraru, G.; Marcu, F.M.; Buga, A.M.L.; Rusu, D.C.; Dragan, F.; Lupu, V.V. The Gut Microbiome and Its Implication in the Mucosal Digestive Disorders. Biomedicines 2022, 10, 3117. [Google Scholar] [CrossRef] [PubMed]
- Lupu, V.V.; Miron, I.C.; Raileanu, A.A.; Starcea, I.M.; Lupu, A.; Tarca, E.; Mocanu, A.; Buga, A.M.L.; Lupu, V.; Fotea, S. Difficulties in Adaptation of the Mother and Newborn via Cesarean Section versus Natural Birth-A Narrative Review. Life 2023, 13, 300. [Google Scholar] [CrossRef] [PubMed]
- Zugravu, C.; Nanu, M.I.; Moldovanu, F.; Arghir, O.C.; Mihai, C.M.; Oțelea, M.R.; Cambrea, S.C. The influence of perinatal education on breastfeeding decision and duration. Int. J. Child Health Nutr. 2018, 7, 74–81. [Google Scholar] [CrossRef]
- Wu, X.; Qian, L.; Liu, K.; Wu, J.; Shan, Z. Gastrointestinal microbiome and gluten in celiac disease. Ann. Med. 2021, 53, 1797–1805. [Google Scholar] [CrossRef]
- Tomer, R.; Patiyal, S.; Dhall, A.; Raghava, G.P.S. Prediction of celiac disease associated epitopes and motifs in a protein. Front. Immunol. 2023, 14, 1056101. [Google Scholar] [CrossRef]
- Liang, C.P.; Geng, L.L.; Chen, P.Y.; Li, H.W.; Ren, L.; Gong, S.T. Celiac disease may be rare among children in South China. J. Int. Med. Res. 2022, 50, 3000605221076923. [Google Scholar] [CrossRef]
- Yuan, J.L.; Xu, J.; Shuai, H.; JinYan, G.; HongBing, C. Recent advances in celiac disease. J. Food Saf. Food Qual. 2015, 11, 4510–4515. Available online: https://www.cabdirect.org/cabdirect/abstract/20153441953 (accessed on 20 May 2023).
- Valitutti, F.; Cucchiara, S.; Fasano, A. Celiac Disease and the Microbiome. Nutrients 2019, 11, 2403. [Google Scholar] [CrossRef]
- Lupu, V.V.; Adam Raileanu, A.; Mihai, C.M.; Morariu, I.D.; Lupu, A.; Starcea, I.M.; Frasinariu, O.E.; Mocanu, A.; Dragan, F.; Fotea, S. The Implication of the Gut Microbiome in Heart Failure. Cells 2023, 12, 1158. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Zu, X.-P.; Xie, H.-S.; Jin, H.-Z.; Yang, N.; Liu, X.-R.; Zhang, W.-D. Research progress in mechanism of intestinal microorganisms in human diseases. Acta Pharm. Sin. 2016, 6, 843–852. [Google Scholar]
- Leonard, M.M.; Valitutti, F.; Karathia, H.; Pujolassos, M.; Kenyon, V.; Fanelli, B.; Troisi, J.; Subramanian, P.; Camhi, S.; Colucci, A.; et al. Microbiome signatures of progression toward celiac disease onset in at-risk children in a longitudinal prospective cohort study. Proc. Natl. Acad. Sci. USA 2021, 118, e2020322118. [Google Scholar] [CrossRef]
- Pozo-Rubio, T.; de Palma, G.; Mujico, J.R.; Olivares, M.; Marcos, A.; Acuña, M.D.; Polanco, I.; Sanz, Y.; Nova, E. Influence of early environmental factors on lymphocyte subsets and gut microbiota in infants at risk of celiac disease; the PROFICEL study. Nutr. Hosp. 2013, 28, 464–473. [Google Scholar]
- Leonard, M.M.; Karathia, H.; Pujolassos, M.; Troisi, J.; Valitutti, F.; Subramanian, P.; Camhi, S.; Kenyon, V.; Colucci, A.; Serena, G.; et al. Multi-omics analysis reveals the influence of genetic and environmental risk factors on developing gut microbiota in infants at risk of celiac disease. Microbiome 2020, 8, 130. [Google Scholar] [CrossRef] [PubMed]
- Hov, J.R.; Zhong, H.; Qin, B.; Anmarkrud, J.A.; Holm, K.; Franke, A.; Lie, B.A.; Karlsen, T.H. The influence of the autoimmunity-associated ancestral HLA haplotype AH8. 1 on the human gut microbiota: A cross-sectional study. PLoS ONE 2015, 10, e0133804. [Google Scholar] [CrossRef] [PubMed]
- De Palma, G.; Capilla, A.; Nadal, I.; Nova, E.; Pozo, T.; Varea, V.; Polanco, I.; Castillejo, G.; López, A.; Garrote, J.; et al. Interplay between human leukocyte antigen genes and the microbial colonization process of the newborn intestine. Curr. Issues Mol. Biol. 2010, 12, 1–10. [Google Scholar]
- Olivares, M.; Neef, A.; Castillejo, G.; De Palma, G.; Varea, V.; Capilla, A.; Palau, F.; Nova, E.; Marcos, A.; Polanco, I.; et al. The HLADQ2 genotype selects for early intestinal microbiota composition in infants at high risk of developing coeliac disease. Gut 2014, 64, 406–417. [Google Scholar] [CrossRef]
- Yoshida, N.; Emoto, T.; Yamashita, T.; Watanabe, H.; Hayashi, T.; Tabata, T.; Hoshi, N.; Hatano, N.; Ozawa, G.; Sasaki, N.; et al. Bacteroides vulgatus and Bacteroides dorei reduce gut microbial lipopolysaccharide production and inhibit atherosclerosis. Circulation 2018, 138, 2486–2498. [Google Scholar] [CrossRef]
- Bayer, A.L.; Fraker, C.A. The folate cycle as a cause of natural killer cell dysfunction and viral etiology in type 1 diabetes. Front. Endocrinol. 2017, 8, 315. [Google Scholar] [CrossRef]
- Chua, H.H.; Chou, H.C.; Tung, Y.L.; Chiang, B.L.; Liao, C.C.; Liu, H.H.; Ni, Y.H. Intestinal dysbiosis featuring abundance of Ruminococcus gnavus associates with allergic diseases in infants. Gastroenterology 2018, 154, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Olshan, K.L.; Leonard, M.M.; Serena, G.; Zomorrodi, A.R.; Fasano, A. Gut microbiota in Celiac Disease: Microbes, metabolites, pathways and therapeutics. Expert Rev. Clin. Immunol. 2020, 16, 1075–1092. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Ishaq, S.L.; Zhao, F.-Q.; Wright, A.-D.G. Colonic inflammation accompanies an increase of β-catenin signaling and Lachnospiraceae/Streptococcaceae bacteria in the hind gut of high-fat diet-fed mice. J. Nutr. Biochem. 2016, 35, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Cabral, D.; Penumutchu, S.; Reinhart, E.M.; Zhang, C.; Korry, B.J.; Wurster, J.I.; Nilson, R.; Guang, A.; Sano, W.H.; Rowan-Nash, A.D.; et al. Microbial metabolism modulates antibiotic susceptibility within the murine gut microbiome. Cell Metab. 2019, 30, 800–823.e7. [Google Scholar] [CrossRef] [PubMed]
- Girdhar, K.; Dogru, Y.D.; Huang, Q.; Yang, Y.; Tolstikov, V.; Raisingani, A.; Chrudinova, M.; Oh, J.; Kelley, K.; Ludvigsson, J.; et al. Dynamics of the gut microbiome, IgA response, and plasma metabolome in the development of pediatric celiac disease. Microbiome 2023, 11, 9. [Google Scholar] [CrossRef]
- Maffeis, C.; Martina, A.; Corradi, M.; Quarella, S.; Nori, N.; Torriani, S.; Plebani, M.; Contreas, G.; Felis, G.E. Association between intestinal permeability and faecal microbiota composition in Italian children with beta cell autoimmunity at risk for type 1 diabetes. Diabetes Metab. Res. Rev. 2016, 32, 700–709. [Google Scholar] [CrossRef]
- Stewart, C.J.; Ajami, N.J.; O’brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Ye, Z.; Zhang, N.; Wu, C.; Zhang, X.; Wang, Q.; Huang, X.; Du, L.; Cao, Q.; Tang, J.; Zhou, C.; et al. A metagenomic study of the gut microbiome in Behcet’s disease. Microbiome 2018, 6, 135. [Google Scholar] [CrossRef]
- Kameyama, K.; Itoh, K. Intestinal colonization by a Lachnospiraceae bacterium contributes to the development of diabetes in obese mice. Microbes Environ. 2014, 29, 427–430. [Google Scholar] [CrossRef]
- Nakanishi, Y.; Sato, T.; Ohteki, T. Commensal Gram-positive bacteria initiates colitis by inducing monocyte/macrophage mobilization. Mucosal Immunol. 2015, 8, 152–160. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? eBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef] [PubMed]
- Lauwers, G.Y.; Fasano, A.; Brown, I.S. Duodenal lymphocytosis with no or minimal enteropathy: Much ado about nothing? Mod. Pathol. 2015, 28, S22–S29. [Google Scholar] [CrossRef]
- Rintala, A.; Riikonen, I.; Toivonen, A.; Pietilä, S.; Munukka, E.; Pursiheimo, J.-P.; Elo, L.L.; Arikoski, P.; Luopajärvi, K.; Schwab, U.; et al. Early fecal microbiota composition in children who later develop celiac disease and associated autoimmunity. Scand. J. Gastroenterol. 2018, 53, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Benítez-Páez, A.; De Palma, G.; Capilla, A.; Nova, E.; Castillejo, G.; Varea, V.; Marcos, A.; Garrote, J.A.; Polanco, I.; et al. Increased prevalence of pathogenic bacteria in the gut microbiota of infants at risk of developing celiac disease: The PROFICEL study. Gut Microbes 2018, 9, 551–558. [Google Scholar] [CrossRef]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef] [PubMed]
- El Mouzan, M.; Al-Hussaini, A.; Serena, G.; Assiri, A.; Al Sarkhy, A.; Al Mofarreh, M.; Alasmi, M.; Fasano, A. Microbiota profile of new-onset celiac disease in children in Saudi Arabia. Gut Pathog. 2022, 14, 37. [Google Scholar] [CrossRef]
- Zafeiropoulou, K.; Nichols, B.; Mackinder, M.; Biskou, O.; Rizou, E.; Karanikolou, A.; Clark, C.; Buchanan, E.; Cardigan, T.; Duncan, H.; et al. Alterations in Intestinal Microbiota of Children with Celiac Disease at the Time of Diagnosis and on a Gluten-free Diet. Gastroenterology 2020, 159, 2039–2051.e20. [Google Scholar] [CrossRef]
- Han, T.; Li, J. Gut microbiota as a new player in children with celiac disease. J. Gastroenterol. Hepatol. 2021, 36, 39–40. [Google Scholar] [CrossRef]
- Wacklin, P.; Kaukinen, K.; Tuovinen, E.; Collin, P.; Lindfors, K.; Partanen, J.; Mäki, M.; Mättö, J. The duodenal microbiota composition of adult celiac disease patients is associated with the clinical manifestation of the disease. Inflamm. Bowel Dis. 2013, 19, 934–941. [Google Scholar] [CrossRef]
- Di Biase, A.R.; Marasco, G.; Ravaioli, F.; Dajti, E.; Colecchia, L.; Righi, B.; D’Amico, V.; Festi, D.; Iughetti, L.; Colecchia, A. Gut microbiota signatures and clinical manifestations in celiac disease children at onset: A pilot study. J. Gastroenterol. Hepatol. 2020, 36, 446–454. [Google Scholar] [CrossRef]
- El Mouzan, M.; Assiri, A.; Al Sarkhy, A. Gut microbiota predicts the diagnosis of celiac disease in Saudi children. World J. Gastroenterol. 2023, 29, 1994–2000. [Google Scholar] [CrossRef] [PubMed]
- Grandi, N.; Tramontano, E. Human Endogenous Retroviruses Are Ancient Acquired Elements Still Shaping Innate Immune Responses. Front. Immunol. 2018, 9, 2039. [Google Scholar] [CrossRef] [PubMed]
- El Mouzan, M.I.; Korolev, K.S.; Al Mofarreh, M.A.; Menon, R.; Winter, H.S.; Al Sarkhy, A.A.; Dowd, S.E.; Al Barrag, A.M.; Assiri, A.A. Fungal dysbiosis predicts the diagnosis of pediatric Crohn’s disease. World J. Gastroenterol. 2018, 24, 4510–4516. [Google Scholar] [CrossRef]
- Santelmann, H.; Howard, J.M. Yeast metabolic products, yeast antigens and yeasts as possible triggers for irritable bowel syndrome. Eur. J. Gastroenterol. Hepatol. 2005, 17, 21–26. [Google Scholar] [CrossRef] [PubMed]
- El Mouzan, M.; Al-Hussaini, A.; Fanelli, B.; Assiri, A.; AlSaleem, B.; Al Mofarreh, M.; Al Sarkhy, A.; Alasmi, M. Fungal Dysbiosis in Children with Celiac Disease. Dig. Dis. Sci. 2021, 67, 216–223. [Google Scholar] [CrossRef]
- Granito, A.; Zauli, D.; Muratori, P.; Grassi, A.; Bortolotti, R.; Petrolini, N.; Veronesi, L.; Gionchetti, P.; Bianchi, F.B.; Volta, U. Anti-Saccharomyces cerevisiae and perinuclear anti-neutrophil cytoplasmic antibodies in coeliac disease before and after gluten-free diet. Aliment. Pharmacol. Ther. 2005, 21, 881–887. [Google Scholar] [CrossRef]
- Mallant-Hent, R.C.; Mary, B.; Von Blomberg, E.; Yüksel, Z.; Wahab, P.J.; Gundy, C.; Meyer, G.A.; Mulder, C.J.J. Disappearance of anti-saccharomyces cerevisiae antibodies in coeliac disease during a gluten-free diet. Eur. J. Gastroenterol. Hepatol. 2006, 18, 75–78. [Google Scholar] [CrossRef]
- Nieuwenhuizen, W.F.; Pieters, R.H.; Knippels, L.M.; Jansen, M.C.; Koppelman, S.J. Is Candida albicans a trigger in the onset of coeliac disease? Lancet 2003, 361, 2152–2154. [Google Scholar] [CrossRef]
- Corouge, M.; Loridant, S.; Fradin, C.; Salleron, J.; Damiens, S.; Moragues, M.D.; Souplet, V.; Jouault, T.; Robert, R.; Dubucquoi, S.; et al. Humoral immunity links Candida albicans infection and celiac disease. PLoS ONE 2015, 10, e0121776. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Candida albicans in celiac disease: A wolf in sheep’s clothing. Autoimmune Rev. 2020, 19, 102621. [Google Scholar]
- El Mouzan, M.; Assiri, A.; Al Sarkhy, A.; Alasmi, M.; Saeed, A.; Al-Hussaini, A.; AlSaleem, B.; Al Mofarreh, M. Viral dysbiosis in children with new-onset celiac disease. PLoS ONE 2022, 17, e0262108. [Google Scholar] [CrossRef] [PubMed]
- Schippa, S.; Iebba, V.; Barbato, M.; Di Nardo, G.; Totino, V.; Checchi, M.P.; Longhi, C.; Maiella, G.; Cucchiara, S.; Conte, M.P. A distinctive ‘microbial signature’ in celiac pediatric patients. BMC Microbiol. 2010, 10, 175. [Google Scholar] [CrossRef]
- Sample, D.M.; Fouhse, J.; King, S.M.; Huynh, H.Q.M.; Dieleman, L.A.M.; Willing, B.P.; Turner, J.M. Baseline Fecal Microbiota in Pediatric Patients with Celiac Disease Is Similar to Controls but Dissimilar After 1 Year on the Gluten-Free Diet. JPGN Rep. 2021, 2, e127. [Google Scholar] [CrossRef]
- Sánchez, E.; Donat, E.; Ribes-Koninckx, C.; Fernández-Murga, M.L.; Sanz, Y. Duodenal-mucosal bacteria associated with celiac disease in children. Appl. Environ. Microbiol. 2013, 79, 5472–5479. [Google Scholar] [CrossRef]
- Logan, K.; Perkin, M.R.; Marrs, T.; Radulovic, S.; Craven, J.; Flohr, C.; Bahnson, H.T.; Lack, G. Early gluten introduction and celiac disease in the EAT study: A prespecified analysis of the EAT randomized clinical trial. JAMA Pediatr. 2020, 174, 1041–1047. [Google Scholar] [CrossRef]
- Hummel, S.; Pflüger, M.; Hummel, M.; Bonifacio, E.; Ziegler, A.G. Primary dietary intervention study to reduce the risk of islet autoimmunity in children at increased risk for type 1 diabetes: The BABYDIET study. Diabetes Care 2011, 34, 1301–1305. [Google Scholar] [CrossRef] [PubMed]
- Beyerlein, A.; Chmiel, R.; Hummel, S.; Winkler, C.; Bonifacio, E.; Ziegler, A.G. Timing of gluten introduction and islet autoimmunity in young children: Updated results from the BABYDIET study. Diabetes Care 2014, 37, e194–e195. [Google Scholar] [CrossRef] [PubMed]
- Sellitto, M.; Bai, G.; Serena, G.; Fricke, W.F.; Sturgeon, C.; Gajer, P.; White, J.R.; Koenig, S.S.K.; Sakamoto, J.; Boothe, D.; et al. Proof of concept of microbiome-metabolome analysis and delayed gluten exposure on celiac disease autoimmunity in genetically at-risk infants. PLoS ONE 2012, 7, e33387. [Google Scholar] [CrossRef]
- Lionetti, E.; Castellaneta, S.; Francavilla, R.; Pulvirenti, A.; Tonutti, E.; Amarri, S.; Barbato, M.; Barbera, C.; Barera, G.; Bellantoni, A.; et al. Introduction of gluten, HLA status, and the risk of celiac disease in children. N. Engl. J. Med. 2014, 371, 1295–1303. [Google Scholar] [CrossRef]
- Vriezinga, S.L.; Auricchio, R.; Bravi, E.; Castillejo, G.; Chmielewska, A.; Escobar, P.C.; Kolaček, S.; Koletzko, S.; Korponay-Szabo, I.R.; Mummert, E.; et al. Randomized feeding intervention in infants at high risk for celiac disease. N. Engl. J. Med. 2014, 371, 1304–1315. [Google Scholar] [CrossRef]
- Aronsson, C.A.; Lee, H.-S.; Hårdaf Segerstad, E.M.; Uusitalo, U.; Yang, J.; Koletzko, S.; Liu, E.; Kurppa, K.; Bingley, P.J.; Toppari, J.; et al. Association of gluten intake during the first 5 years of life with incidence of celiac disease autoimmunity and celiac disease among children at increased risk. JAMA 2019, 322, 514–523. [Google Scholar] [CrossRef]
- Mårild, K.; Dong, F.; Lund-Blix, N.A.; Seifert, J.; Barón, A.E.; Waugh, K.C.; Taki, I.; Størdal, K.; Tapia, G.; Stene, L.C.; et al. Gluten intake and risk of celiac disease: Long-term follow-up of an At-risk birth cohort. Am. J. Gastroenterol. 2019, 114, 1307–1314. [Google Scholar] [CrossRef] [PubMed]
- Lund-Blix, N.A.; Marild, K.; Tapia, G.; Norris, J.M.; Stene, L.C.; Stordal, K. Gluten intake in early childhood and risk of celiac disease in childhood: A nationwide cohort study. Am. J. Gastroenterol. 2019, 114, 1299–1306. [Google Scholar] [CrossRef]
- Aronsson, C.A.; Agardh, D. Intervention strategies in early childhood to prevent celiac disease-a mini-review. Front. Immunol. 2023, 14, 1106564. [Google Scholar] [CrossRef]
- Hakansson, A.; Andren Aronsson, C.; Brundin, C.; Oscarsson, E.; Molin, G.; Agardh, D. Effects of lactobacillus plantarum and lactobacillus paracasei on the peripheral immune response in children with celiac disease autoimmunity: A randomized, double blind, placebo-controlled clinical trial. Nutrients 2019, 11, 1925. [Google Scholar] [CrossRef] [PubMed]
- Ahrén, I.L.; Berggren, A.; Teixeira, C.; Martinsson Niskanen, T.; Larsson, N. Evaluation of the efficacy of Lactobacillus plantarum HEAL9 and Lactobacillus paracasei 8700:2 on aspects of common cold infections in children attending daycare: A randomised, double-blind, placebo-controlled clinical study. Eur. J. Nutr. 2020, 59, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Lindfors, K.; Lin, J.; Lee, H.-S.; Hyoty, H.; Nykter, M.; Kurppa, K.; Liu, E.; Koletzko, S.; Rewers, M.; Hagopian, W.; et al. Metagenomics of the faecal virome indicate a cumulative effect of enterovirus and gluten amount on the risk of coeliac disease autoimmunity in genetically at risk children: The TEDDY study. Gut 2019, 69, 1416–1422. [Google Scholar] [CrossRef] [PubMed]
- Stene, L.C.; Honeyman, M.C.; Hoffenberg, E.; Haas, J.E.; Sokol, R.J.; Emery, L.; Taki, I.; Norris, J.M.; Erlich, H.A.; Eisenbarth, G.S.; et al. Rotavirus infection frequency and risk of celiac disease autoimmunity in early childhood: A longitudinal study. Am. J. Gastroenterol. 2006, 101, 2333–2340. [Google Scholar] [CrossRef] [PubMed]
- Kemppainen, K.M.; Lynch, K.F.; Liu, E.; Lönnrot, M.; Simell, V.; Briese, T.; Koletzko, S.; Hagopian, W.; Rewers, M.; She, J.X.; et al. Factors that increase risk of celiac disease autoimmunity after a gastrointestinal infection in early life. Clin. Gastroenterol. Hepatol. 2017, 15, 694–702.e5. [Google Scholar] [CrossRef] [PubMed]
- Quagliariello, A.; Aloisio, I.; Cionci, N.B.; Luiselli, D.; D’Auria, G.; Martinez-Priego, L.; Pérez-Villarroya, D.; Langerholc, T.; Primec, M.; Mičetić-Turk, D.; et al. Effect of Bifidobacterium breve on the Intestinal Microbiota of Coeliac Children on a Gluten Free Diet: A Pilot Study. Nutrients 2016, 8, 660. [Google Scholar] [CrossRef] [PubMed]
- Klemenak, M.; Dolinšek, J.; Langerholc, T.; Di Gioia, D.; Mičetić-Turk, D. Administration of Bifidobacterium breve Decreases the Production of TNF-α in Children with Celiac Disease. Dig. Dis. Sci. 2015, 60, 3386–3392. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Castillejo, G.; Varea, V.; Sanz, Y. Double-blind, randomised, placebo-controlled intervention trial to evaluate the effects of Bifidobacterium longum CECT 7347 in children with newly diagnosed coeliac disease. Br. J. Nutr. 2014, 112, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Primec, M.; Klemenak, M.; Di Gioia, D.; Aloisio, I.; Cionci, N.B.; Quagliariello, A.; Gorenjak, M.; Mičetić-Turk, D.; Langerholc, T. Clinical intervention using Bifidobacterium strains in celiac disease children reveals novel microbial modulators of TNF-α and short-chain fatty acids. Clin. Nutr. 2018, 38, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Drabińska, N.; Jarocka-Cyrta, E.; Markiewicz, L.H.; Krupa-Kozak, U. The Effect of Oligofructose-Enriched Inulin on Faecal Bacterial Counts and Microbiota-Associated Characteristics in Celiac Disease Children Following a Gluten-Free Diet: Results of a Randomized, Placebo-Controlled Trial. Nutrients 2018, 10, 201. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Study | Subjects | Sample and Techniques | Microbiota Alterations |
|---|---|---|---|
| Pozo-Rubio et al. [24] | 55 infants | Blood-sample-flow cytometry analysis Fecal sample quantitative PCR analysis | -Infants born through cesarean delivery: ↓ B. catenulatum ↑B. angulatum -Antibiotic use during pregnancy ↓B. angulatum -Formula feeding ↓B. angulatum -Antibiotic use during first the 4 months of life ↑Bacteroides fragilis ↑B. angulatum ↓Bifidobacterium spp. B. longum -Rotavirus vaccine ↓Bacteroides fragilis -Allergy and dermatitis ↓B. angulatum |
| Leonard et al. [25] | 21 genetically predisposed infants 5 genetically non-predisposed infants | Fecal sample metagenomic analysis | -Standard and high risk of CD: ↓Streptococcus spp. ↓Coprococcus spp. ↓Veillonella spp. ↓Parabacteroides spp. ↓Clostridium perfringens ↑Bacteroides spp. ↑Enterococcus spp. -Infants born through cesarean delivery: ↓ Bacteroides spp. ↓Parabacteroides spp. ↑Enterococcus faecalis -Formula feeding ↓Bifidobacterium breve ↓Staphylococcus epidermis ↑Bifidobacterium adolescentis ↑Ruminococcus gnavus ↑Lachnospiraceae bacterium -Infant antibiotic exposure ↑Bacteroides thetaiotaomicron ↑Propionibacterium spp. ↑Subdoligranulum spp. ↓Bifidobacterium merycicum ↓Streptococcus lutetiensis |
| de Palma et al. [27] | 20 infants | Fecal sample fluorescence in situ hybridization analysis | -High-risk infants ↑Gram-negative bacteria ↑Bacteroides-Prevotella group ↑E. coli ↑Streptococcus-Lactococcus spp. ↑E. rectale-C. coccoides -Sulfate-reducing bacteria ↑C. lituseburense ↑C. histolyticum |
| Olivares et al. [28] | 22 infants | Fecal sample 16S rRNA gene pyrosequencing and real-time quantitative PCR analysis | -High-risk infants ↑ Bacillota phylum ↑Pseudomonadota phylum ↑Corynebacterium genus ↑Gemella genus ↑Clostridium sensu stricto ↑Escherichia/Shigella ↓Actinomycetota phylum ↓Bifidobacterium spp |
| Leonard et al. [23] | 20 infants | Fecal sample analysis using shotgun sequencing and metabolomic profiling | ↓Bacteroides vulgatus str_3775_S_1080 Branch ↓Bacteroides uniformis_ -American Type Culture Collection (ATCC)_8492 ↓Streptococcus thermophiles ↓Faecalibacterium prausnitzii ↓Clostridium clostridioforme ↓Veillonella parvula ↑Dialister invisus strain DSM_15470 ↑Parabacteroides species and strains ↑Lachnospiraceae bacterium ↑Bifidobacterium longum ↑Bifidobacterium breve ↑Escherichia coli ↑Clostridium hathewayi ↑Eubacterium eligens |
| Ghirdhar et al. [35] | 33 children | Fecal sample 16S rRNA sequencing and flow cytometry analysis Blood plasma sample metabolomic analysis | ↑IgA-coated bacteria and unique targets of IgA in their gut microbiota |
| Rintala et al. [43] | 27 infants | Fecal sample 16S rRNA sequencing | No statistically significant differences in early microbiota composition between children that later developed CD and healthy controls were found |
| Olivares et al. [44] | 127 infants | Fecal sample 16S rRNA sequencing | -High risk of CD, both formula and breastfeeding ↑ETEC -Formula feeding ↑C. perfringens ↑C. difficile |
| Study | Subjects | Sample and Techniques | Microbiota Alterations | Other Findings |
|---|---|---|---|---|
| El Mouzan et al. [46] | 20 CD children 20 fecal controls 19 mucosal controls | Duodenal samples of CD ↑Pseudomonadota phylum ↑Lactobacillus acidophilus, Neisseria spp. ↑Coprococcus spp. Fecal samples of CD ↑Verucomicrobia spp. ↑Clostridium spp. ↑Escherichia spp. ↑Lachnospiraceae_bacterium_oral ↓Bifidobacterium genus ↓Bacteroides spp. | Fecal samples were more diverse and richer in bacteria compared with mucosal samples Bacillota and Bacteroidota were the most abundant phyla in both fecal and mucosal samples | |
| Zafeiropoulou et al. [47] | 20 CD children 45 CD under GFD 57 healthy controls 19 children at risk of CD | Fecal sample 16S rRNA sequencing | Untreated CD ↓Clostridium sensu stricto 1 genus ↓Ruminococcus genus | Microbial dysbiosis was not reported in CD compared to healthy controls Alistipes was correlated with the presence of symptoms of CD |
| Di Base et al. [50] | 21 CD children 16 healthy controls | Fecal samples Duodenal sample 16S rRNA sequencing | Duodenal samples of CD ↑Enterobacteriaceae family ↑Bacteroidetes/Streptococcus spp. Fecal samples of CD ↓ Bacteroides–Prevotella ↓Akkermansia spp. ↓Staphylococcaceae family | Patients with abdominal pain ↑Bacillaceae family ↑Enterobaeriaceae family Patients with diarrhea ↓Clostridium cluster XIVa ↓Akkermansia ↑Bacillaceae ↑Fusobacterium |
| Schippa et al. [62] | 20 CD children, before and after GFD 10 healthy controls | Duodenal sample 16S ribosomal DNA analysis compared with TTGE | In CD patients vs. controls ↑Bacteroides vulgatus ↑Escherichia coli Active CD vs. Inactive CD prevalence B. vulgatus (80% vs. 90%) Clostridium coccoides group (50% vs. 90%) Bifidobacterium spp (20% vs. 40%) | Mean interindividual similarity index: 54.9% ± 14.9% Active CD 55.6% ± 15.7% remission state 21.8% ± 30.16% controls Similarity index between CD children before and after GFD: 63.9% ± 15.8% |
| Sample et al. [63] | 22 CD children, before and after GFD 17 healthy controls | Fecal sample16S ribosomal RNA sequencing | Active CD vs. Controls ↑Haemophilus genera ↑Alistipes genera ↑Bacteroides genera | |
| El Mouzan et al. [51] | 40 CD children 39 controls | Fecal samples Duodenal sample metagenomic analysis of microbial DNA | Fecal samples of CD ↓Bacteroides intestinalis ↓Burkholderiales bacterium 1-1-47 Mucosal samples of CD ↓Human_endogenous_retrovirus_K) | |
| El Mouzan et al. [61] | 40 CD children 39 controls | Fecal samples Duodenal sample metagenomic analysis of microbial DNA | Fecal samples ↑ Human polyomavirus 2, Enterobacteria phage mEpX1, Enterobacteria phage mEpX2 | Mucosal samples—no association with CD |
| El Mouzan et al. [55] | 40 CD children 39 controls | Fecal samples Duodenal sample metagenomic analysis of microbial DNA | Fecal samples of CD ↓Pichiaceae family ↓Pichia kudriavzevii ↑Saccharomycetes family ↑Saccharomyces cerevisiae ↑Tricholomataceae family ↑Mucosal samples of CD ↑ Saccharomycetaceae family ↑Candida spp. ↓Pneumocystis spp. ↓Pneumocystis jirovecii | Fecal fungal communities were more abundant than those observed in mucosal samples |
| Sanchez et al. [64] | 32 active CD on GFD 17 8 healthy controls | Duodenal mucosa sample 16S ribosomal RNA sequencing | Active CD ↑Pseudomonadota phylum ↑Enterobacteriaceae family ↑Klebsiella oxytoca ↑Staphylococcus epidermidis ↑Staphylococcus pasteuri ↓Bacillota phylum ↓Streptococcaceae family | Non-active CD ↑Streptococcus mitis group |
| Study | Subjects | Sample and Techniques | Microbiota Alterations | Other Findings |
|---|---|---|---|---|
| Di Cagno [45] | 19 CD children under GFD (T-CD) 15 non-celiac controls | Fecal samples Duodenal samples,-both on PCR and DGGE analysis | Duodenal biopsy ↑Eubacteria in T-CD Fecal samples of T-CD ↑Bacteroides spp. ↑Staphylococcus spp. ↑Salmonella spp. ↑Shigella spp. ↑Klebsiella spp. | |
| Sample et al. [63] | 22 CD children before and after GFD 17 healthy controls | Fecal sample 16S ribosomal RNA sequencing | CD after GFD vs. controls ↑Haemophilus genera ↑Alistipes genera ↑Bacteroides genera ↑Holdemania genera ↑Blautia genera | Faecalibacterium and Roseburia were enriched in patients whose aTTG levels did not normalize after GFD |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupu, V.V.; Trandafir, L.M.; Raileanu, A.A.; Mihai, C.M.; Morariu, I.D.; Starcea, I.M.; Mocanu, A.; Butnariu, L.I.; Stoleriu, G.; Salaru, D.L.; et al. Advances in Understanding the Human Gut Microbiota and Its Implication in Pediatric Celiac Disease—A Narrative Review. Nutrients 2023, 15, 2499. https://doi.org/10.3390/nu15112499
Lupu VV, Trandafir LM, Raileanu AA, Mihai CM, Morariu ID, Starcea IM, Mocanu A, Butnariu LI, Stoleriu G, Salaru DL, et al. Advances in Understanding the Human Gut Microbiota and Its Implication in Pediatric Celiac Disease—A Narrative Review. Nutrients. 2023; 15(11):2499. https://doi.org/10.3390/nu15112499
Chicago/Turabian StyleLupu, Vasile Valeriu, Laura Mihaela Trandafir, Anca Adam Raileanu, Cristina Maria Mihai, Ionela Daniela Morariu, Iuliana Magdalena Starcea, Adriana Mocanu, Lacramioara Ionela Butnariu, Gabriela Stoleriu, Delia Lidia Salaru, and et al. 2023. "Advances in Understanding the Human Gut Microbiota and Its Implication in Pediatric Celiac Disease—A Narrative Review" Nutrients 15, no. 11: 2499. https://doi.org/10.3390/nu15112499