
Foraging Wild Edibles: Dietary Diversity in Expanded Food Systems
by
 , and
, and
Anne C. Bellows
1,*,
Sudha Raj
1,
Ellen Pitstick
1,
Matthew R. Potteiger
2 and
Stewart A. W. Diemont
3 1
Department of Nutrition and Food Studies, Falk College, Syracuse University, Syracuse, NY 13244, USA
2
Department of Landscape Architecture, College of Environmental Science and Forestry, State University of New York, Syracuse, NY 13210, USA
3
Department of Environmental Biology, College of Environmental Science and Forestry, State University of New York, Syracuse, NY 13210, USA
*
Author to whom correspondence should be addressed.
Nutrients 2023, 15(21), 4630; https://doi.org/10.3390/nu15214630
Submission received: 11 September 2023
/
Revised: 18 October 2023
/
Accepted: 19 October 2023
/
Published: 31 October 2023
(This article belongs to the Section Phytochemicals and Human Health)
Abstract
:Human food foraging in community forests offers extensive and expandable sources of food and high-quality nutrition that support chronic disease prevention and management and are underrepresented in US diets. Despite severe gaps in non-commercial “wild food” data, research in Syracuse, NY, identified substantial amounts of five key antioxidant phytochemicals in locally available, forageable foods with the potential to augment local dietary diversity and quality. Findings endorse the need for micro- and macro-nutrient research on an expanded range of forageable foods, community nutrition education on those foods, an expanded study on antioxidant phytochemical function, and the inclusion of forageables in the food system definition.
1. Introduction
This paper makes the case that human food foraging in community forests encompasses extensive and expandable sources of food and high-quality nutrition, notably antioxidant phytochemicals, that are underrepresented in US diets. Current US food consumption patterns misalign with the Dietary Guidelines Advisory Committee counsel that promotes foods with protective roles in decreasing the risk of all-cause mortality and chronic disease risk [1] and additionally supports environmental sustainability [2,3]. Instead of nutrient-dense and relatively unprocessed foods spread across six core food groups, i.e., vegetables, fruits, grains, dairy, proteins, and oils [4], residents consume few fruits, vegetables, whole grains, and dairy, and too many refined grains and foods containing added sugars, saturated fats, or high levels of sodium [5]. To counter the contribution of our globalized and commodified food system on present day diets [6,7], alternative forms of high-quality, more localized, and low- or no-cost food access are called for [8,9]. The study joins two discrete research questions: (a) What forageable plants, “wild” and intentionally introduced and grown without cultivation practices for commercial or domestic use, are available in Syracuse, NY, USA? (b) What is their nutrient content, including the presence of bioactive compounds, specifically antioxidant phytochemicals? The findings present arguments that foraging supports a largely untapped capacity to augment a community’s affordable, nutrient-dense dietary alternatives.
Foraging is a biocultural practice of nurture and exchange between humans and their local environment [10,11] that bolsters community nutrition and well-being across the world [12,13,14]. Shackleton et al. [15] introduce foraging in cities and towns, which is performed across undeveloped and built environments alike, as “the practice of harvesting or gathering raw biological resources within urban and peri-urban settings primarily for direct consumption, decoration, crafts, barter, or small-scale sale” (p. 2). Bukowski and Munsell [16] describe the “big picture” of urban food forestry as
“The intentional use of perennial food-producing plants to improve the sustainability and resilience of urban communities. A wide range of activities and projects fit this definition, including community food forests, urban orchards, foraging in public urban forests, and community-wide gleaning programs. Even maintaining a small food forest on a private residential lot in a city is considered urban food forestry” (p. 6).
Plants provide humans with macro- and micronutrients that are essential for optimal health and wellness by contributing to nutrient density and dietary diversity in mindfully planned diets. Additionally, plants produce phytochemicals that are bioactive nutrient and non-nutrient compounds that are able to attenuate metabolic imbalances such as oxidative stress and chronic inflammation in a protective capacity [17].
These metabolic imbalances are major contributors to chronic degenerative diseases such as cancer, metabolic syndrome, and cardiovascular disease [18,19]. Conventional allopathic medicine utilizes a variety of pharmaceuticals to treat these diseases, often with limited success and co-existing with the impact of side effects and questions related to the safety of such products. Phytochemicals such as polyphenols, carotenoids, anthocyanins, alkaloids, glycosides, saponins, and terpenes, as well as dietary antioxidants such as vitamins A, C and E, from natural whole foods such as fruits, vegetables, and spices, are dietary sources that have therapeutic potential to lessen the oxidative stress and inflammation associated with degenerative diseases. Dietary antioxidants such as vitamins A, C and E and other nutrients such as selenium and riboflavin that are essential cofactors for antioxidant enzymes participate in the process of scavenging free radicals, thereby reducing their degenerative potential in the chronic disease progression [20,21]. The complexities of their biological action remain a field of significant scientific interest. This paper focuses on a selected subset of antioxidant phytochemicals to be identified in forageable plants (such as lambsquarters, serviceberry, and black walnut), for which the available epidemiological evidence verifies their synergistic value in offering protection against chronic disease. In other words, the full, and only partially understood, power of antioxidant phytochemicals can be realized through foraged food sources. These bioactives, independently and in certain complementary combinations (or “packages”), can offer additive and synergistic benefits that may be more than the sum of individual benefits [22]. It is, however, important to recognize that (a) the action of these phytochemical bioactives is influenced by individual nutritional status, genetic predispositions, and socio-cultural and ecological environments and that (b) not all complex “packages” offer benefits and that certain bioactives and their associated complexes may also be antagonistic and not beneficial [23]. Phytochemicals such as saponins, terpenes, glycoalkaloids, etc., are categorized as antinutrients because they can bind to minerals, compromising their bioavailability. At low concentrations, antinutrients such as phytates, lectins and tannins can act as antioxidants and glucose and insulin modulators [24]. Fermentation, germination, milling, autoclaving, and soaking are food preparation techniques that can reduce the level of antinutrients in food [25].
The focus of this paper is on nutrient content, not toxicity risk. Authors readily acknowledge that addressing toxicity is as vital to the discussion of wild edibles as it is to commercially available foods. Concerns about contaminants in wild and commercial foods are not new and have been addressed with calls for caution in both cases [26]. In wild foods, these concerns range from the consumption of poisonous or allergy-inducing plants or parts of plants to risks associated with the potential presence of pesticides, herbicides, heavy metals, and polycyclic aromatic hydrocarbons (PAHs), among others [27,28,29,30,31,32,33]. Broad community education is of course required. Risk-reduction strategies can help, e.g., in avoiding difficult-to-identify plants (note that mushroom species are omitted from this article’s review) or urging newcomers to wild edibles to learn from experienced foragers in the field. Foraging education must be extended to local governments so that appropriate environmental policy can adapt to public food harvests [32,34]. For example, herbicide use must be curtailed or warning signs posted in a manner similar to how many municipalities provide signage and education on risks associated with urban fishing. A closer review of toxic risks is outside the scope of this paper.
This article derives from inquiry generated by the Syracuse Urban Food Forest Project, started in 2019 with goals of identifying existing forageables in an urban riparian corridor and augmenting them with supplemental plantings. Three steps shape our objective of demonstrating the antioxidant phytochemical potential in local, accessible, and low- or no-cost forageable foods. The first step is to identify good sources of the selected antioxidant phytochemicals (vitamins A, C, and E, selenium, and riboflavin) in forageable foods growing in the urban forests in Syracuse, NY. Where data are available, we report all found forageables and their phytochemical availability. The second step is to introduce a method to characterize an antioxidant “package” consisting of the five selected phytochemicals reviewed with the intention of portraying synergistic interactions of co-existing nutrient and non-nutrient bioactives. This method is applied to forageables for which relevant data are available for all five selected phytochemicals, as well as to select commercial plants. The final step is to expose gaps in available data on antioxidant phytochemicals in wild, forageable foods.
2. Antioxidants and Health
2.1. Diet-Related Chronic Disease and Food Supply
Nutrition-related non-communicable chronic diseases are recognized as the most significant contributors to morbidity and mortality in the world today. More than 50% of all deaths can be attributed to degenerative, inflammation-related diseases such as ischemic heart disease, stroke, cancer, diabetes mellitus, chronic kidney disease, non-alcoholic fatty liver disease, and autoimmune and neurodegenerative conditions [35]. Altered body composition associated with pathophysiological and/or metabolic anomalies such as oxidative stress and chronic inflammation is one of the well-documented risk factors at the nexus of the global chronic disease epidemic. A potent contributor to this growing epidemic is “nutrition transition”, a global-scale phenomenon marked by far-reaching alterations in the food supply. Across the world, food systems have been moving away from culturally based, traditional, and indigenous food economies grounded at the local and subsistence scale and moving toward cash crop production. Industrialized economies are characterized by consolidation of food production, economies of scale, money, and cash markets that determine what is produced and sold in the global food system. Selective breeding techniques, fortification and enrichment strategies, the expansion of functional foods and nutraceuticals, novel food processing and packaging technologies, and information and communication technologies have increased agricultural commodities’ yields, encouraged product innovation, diversification, and the volume of production of processed and ultraprocessed foods [36]. Concomitant advancements in the retail, market, advertising, and transportation sectors further ensure guaranteed availability of such products to global populations worldwide. However, much heterogeneity exists within and between global countries on the consequences of this industrialization and globalization depending on the ecological, internationalization, political, and technological climates that prevail within them. As local food markets have expanded to include more industrialized and commodified food products, traditional indigenous foods have been replaced by ultraprocessed foods high in sugars, processed fats, and sodium [37,38,39]. Cultural foodways, dietary traditions, and related food practices have been altered by pervasive industrialization and urbanization. These structural conditions behind nutrition transition correlate with uneven socio-economic improvement, wealth–poverty polarization, and attendant purchasing power disparities, as well as injury to cultural landscape ecologies. Habitat destruction, loss of land tenure, and rural out-migration have undermined traditional food activities like foraging.
Under a nutrition transition, consumption is facilitated by the easy and often year-round accessibility of shelf-stable processed foods. At the same time, access to whole and nutrient-dense fruits, vegetables, and whole grains is limited for many segments of the population due to higher prices and sporadic availability. Further, traditional and Indigenous food access (e.g., food production for local markets, subsistence production, and foraging) and related cultural knowledge continue to decline, adding to the accessibility dilemma. Wild foraging has the capacity to provide consumers with nutritional variety, accessibility, and expanded food choice within their local food system. Altered dietary patterns along with concurrent lifestyle behaviors such as growing sedentarism have contributed to body composition changes, seen as overweight and obesity. Depending upon the degree and pace of these modifications, different parts of the globe have experienced a mix of positive and negative consequences. The net result, however, is population-scale conversion to poorer-quality diets characterized by more processed products that are rich in undesirable ingredients such as fats, oils, and sweeteners and often lacking nutrient-dense foods that can offer protection against chronic diseases [18,40].
At the global scale, we additionally observe a shift from undernutrition as the main nutrition problem to a triple burden of disease. The latter is characterized by overnutrition and undernutrition coupled with maternal mortality that is reflective of extensive population-scale food and nutrition insecurity, nutritional deficiency and excess in some cases, and the persisting burden of infectious diseases. Although distinct in terms of caloric excess, nutritional well-being has become increasingly limited or extinct in overnutrition cases. Overall, poor-quality diets marked by calorie- and energy-dense foods continue to replace nutrient-dense options, resulting in a dietary landscape wherein chronic diseases such as diabetes, cardiovascular disease, cancer, etc., prevail. Additionally, the poor micronutrient status of these diets affords little immunity against infectious diseases. Poverty, food insecurity, exposure to environmental toxins, and income and health access disparities further undermine the health of global populations. These factors, exacerbated by genetic predisposition and chronic stress, overburden the healthcare system’s capacity to simultaneously handle the dual burden of co-existing communicable and non-communicable diseases and lead to lowered immunity and chronic disease across the lifespan [41,42,43,44].
Researchers interested in the fetal origins of adult-onset disease view the triple burden of malnutrition in pregnancy and early fetal life as a major threat that increases the susceptibility to chronic metabolic diseases such as diabetes in later life [43,44]. Poor nutritional status accompanied by metabolic disturbances such as insulin resistance in pregnancy mediated by chronic inflammation and oxidative stress leads to a compromised placental nutrient supply to the fetus [45]. This can lead to developmental adaptations resulting in a compromised or altered organ structure, for instance, a pancreas with fewer islet cells and altered functional capacity accompanied by altered metabolic, endocrine, nutrient sensing and inflammatory signaling pathways [43,45,46,47,48].
2.2. Chronic Disease and the Underlying Dysfunctions in Oxidative Stress, Inflammation, and Detoxification
Current research suggests that chronic disease occurs when three inherent and necessary metabolic processes—namely oxidative stress, inflammation, and detoxification—in the human body become less resilient [41,49]. These processes, independently and synergistically, are integral for humans to function. For example, the acute inflammatory response that is critical to recovering from trauma and oxidative stress produces free radicals for pathogen destruction. However, these processes can become vicious and self-perpetuating under abnormal circumstances, such as stress impacting various physiological systems and ultimately causing disease. Restoring balance involves strengthening the physiological systems to better handle these processes by addressing genetic anomalies and the provision of good-quality nutrition, living environments, and positive lifestyle behaviors.
Oxidative stress occurs when there is an imbalance between the production of highly unstable free radicals such as reactive oxygen and nitrogen species with an unpaired electron and the body’s antioxidant defenses to address these free radicals [50,51,52]. Free radicals with one or more unpaired electrons are normally created in the body via a variety of physiological processes (e.g., respiration) and genetic abnormalities (e.g., inborn errors of metabolism) and as a result of exposure (e.g., pollution). Within limits, free radicals are critical in cellular processes such as cell signaling. Under normal physiological conditions, free radicals are dealt with by a combination of endogenous antioxidant systems produced within the body such as enzymes like catalase, superoxide dismutase, and exogenous antioxidants such as vitamins C, A, and E obtained from the diet. These systems individually and in combination neutralize the free radicals by giving up their own electrons and maintaining a steady supply of available antioxidants through the processes of oxidation and reduction [31,51,52]. However, adverse chronic environmental exposures and dysfunctional physiological processes can lead to a buildup of free radicals, resulting in oxidative stress. This situation is further exacerbated by an inadequate intake of antioxidants from food and/or insufficient or inefficient functioning of endogenous antioxidant systems. These condition results in both the consequent inhibition of the body’s protective defensive mechanisms and damage from free radicals. The uncontrolled production of free radicals can result in, for example, damage to the lipid-rich cellular membranes, the creation of nonfunctional proteins, and DNA damage causing mutagenesis [51]. This occurs because free radicals indiscriminately target biomolecules like lipids, proteins, and DNA in the hopes of securing their missing electron.
Inflammation (acute and chronic) is a normal protective response that is critical to survival yet when unabated can become a threat to human health. The acute inflammatory response is seen in conditions characterized by tissue damage due to trauma, microbial invasion, or noxious compounds, as is the case when one gets a bruise or when a cut becomes infected. Under these circumstances, several immune system components participate in eliciting the inflammatory response. Acute inflammatory responses are typically resolved to baseline levels in a short period of time depending on the efficiency and functionality of the immune system. In chronic inflammation, the immune response is initiated in a similar manner, but the degree of resolution is modulated by genetic predispositions or environmental factors such as diet, physical inactivity, chronic stress, and lifestyle behaviors. A persistent hyperactivated stress response and/or a dysfunctional immune response accompanied by undesirable nutrition and lifestyle factors make the resolution of inflammation unachievable. Chronic inflammation is marked by increasing levels of pro-inflammatory mediators such as Interleukin-6 and TNF-alpha, cortisol buildup, and insulin resistance that eventually culminate in disease. The intensity and impact of the inflammatory response appear to be highly individualized [53]. The effective resolution of the inflammatory response is regulated by an arsenal of endogenous and dietary anti-inflammatory mediators. These mediators can synergistically and effectively inhibit or block actions that perpetuate inflammation, thereby preventing the progression of the chronic inflammatory state [54].
Detoxification is the third process by which the body identifies, neutralizes, and eliminates toxic substances. This process is also influenced by lifestyle and environmental factors, the total toxic load experienced by an individual determined by exposure to toxins, the efficiency of the body’s endogenous detoxification enzyme systems that involve the Phase I and Phase II cytochrome P450 enzyme systems, and the availability of nutrient and non-nutrient cofactors for the proper action of these enzyme systems [55,56].
2.3. Reducing Oxidative Stress and Chronic Inflammation
Dietary patterns rich in minimally processed, nutrient-dense foods and low in ultra-processed foods are associated with a reduced risk of unimpeded oxidative stress, chronic inflammation, and enhanced detoxification processes. Whole foods such as fruits, vegetables, whole grains, nuts, legumes, etc., contain a plethora of bioactive compounds that include antioxidant phytochemicals, macro- and micronutrients such as lipid bioactives, vitamins, minerals, and dietary fibers in natural ratios and proportions [57]. These compounds, when present in their innate food matrices, in contrast to industrially created dietary supplements, act in an independent, complementary, or synergistic manner to maximize disease prevention and offer a therapeutic potential beyond nutrition [58]. Food bioactives exert their therapeutic and protective potential in several ways. These include influencing cell signaling pathways, engaging in scavenging free radicals, interacting with transcription factors such as NF-kB and Nrf2 that are related to anti-inflammatory and antioxidant effects, as well as enhancing anti-inflammatory effects via the production of short-chain fatty acids by the gut microbiota. In this regard, whole foods, with their innate combinations of bioactive antioxidants and phytochemicals, can keep processes such as oxidative stress and chronic inflammation at bay. Although several thousand food bioactives have been identified, much remains to be learned about their bioavailability, mode of action, and dietary requirements [59].
There is no simple guide to an ideal composition of antioxidant phytochemicals that can ameliorate unimpeded oxidative stress and chronic inflammation and that can boost detoxification processes [60,61,62]. Yet there is substantial scientific evidence that indicates higher dietary intakes and/or blood concentrations of vitamin C, carotenoids and tocopherol are associated with reduced risk of cardiovascular disease, cancer and total mortality [63]. Researchers further highlight that these benefits are accrued not due to the individual antioxidants themselves but the combination of the various bioactive compounds that coexist within the food matrix, underscoring the importance of consistently higher dietary intakes of a diverse assortment of whole foods. This recommendation is made based on the premise that there is a constantly fluctuating balance between the proinflammatory and oxidative processes that occur in the body and the available exogenous dietary antioxidants and endogenous antioxidant enzyme systems to handle these processes. Additionally, while genetics determine the endogenous antioxidant profile, the availability of exogenous antioxidants varies with individual dietary patterns, dietary diversity, seasonality, the local availability and accessibility of whole foods, and appropriate behavior-modification efforts. This underscores the importance of increasing consumer awareness of the value of such products and encouraging their use in chronic disease prevention and management [64].
The argument is made that dietary patterns such as the Mediterranean diet featuring a predominance of whole foods provide a food bioactive package that offers significant health benefits and immune-boosting capabilities [65]. A steady supply of these bioactive compounds in an appropriately planned meal pattern can assist in overcoming the imbalance created by the free radicals and proinflammatory compounds. Through a variety of mechanisms such as anti-inflammatory and chemo-preventive effects, food bioactives can simultaneously provide various benefits that can offer additive and synergistic advantages [58]. A beneficial antioxidant bioactive “package” would then include both “direct antioxidants” (like vitamins A, C, and E that themselves participate in free radical scavenging) and “helper antioxidants” (like riboflavin and selenium that participate as cofactors for important anti-inflammatory endogenous enzyme systems such as glutathione peroxidase) [66,67], as well as efficiently support detoxification enzyme systems such as the Phase 1 and 2 enzymes [68,69].
3. Method
3.1. Case Study Background
3.1.1. Location
The Syracuse Urban Food Forest Project (SUFFP) aims to (a) learn how edible urban forests can be structured to optimize human and wildlife health and well-being; (b) work with communities to share and expand knowledge about the social benefits of public harvests in urban forests; and (c) recognize and respond to historical and contemporary infringements on rights of land tenure, public land access, and green space equity. To examine whether a local, forageable food system can enhance dietary diversity, we chose to examine total antioxidant and phytochemical availability as a first step. This strategy could reveal local foods that can potentially reduce chronic disease risk in the population. The SUFFP, initiated in 2019, focuses on a seven-mile corridor on the south and southwest side of Syracuse, NY. The riparian transect runs along Furnace Brook to its intersection with Onondaga Creek and northward through the city center to Lake Onondaga. The corridor is surrounded by public land, parks, schools, residential low-low-income and resurgent (identified by the city for needed improvements) neighborhoods, and two urban farms (see Figure 1) [70].
3.1.2. Poverty, Urban Food Infrastructure, and Health Nexus
Southside Syracuse experiences social and economic isolation and inequality that, in turn, precipitate poor health and nutrition outcomes. The land sits on unceded Onondaga Territory, but the actual number of tribal members of this branch of the Haudenosaunee living within the Syracuse city limits is quite small. As of 2020, the overall population of Syracuse was 148,620 [71], with an estimated 30.3% of Syracuse residents living below the poverty level (more recent US census data are available but do not include the American Indian and Alaskan Native population due to limited sample size) [72]. The city’s history of colonization, redlining, and environmental racism has had lasting effects visible through segregated poverty levels, where 33.3% of American Indian and Alaskan Native, 40.0% of Black or African American, and 42.9% of the Hispanic or Latino populations live below the poverty level, compared to only 23.3% of the White population [72,73]. In 2020, the estimated median income in Syracuse was USD 38,893. American Indian and Alaskan Native residents were estimated to earn a median income of USD 23,316, Black residents a median income of USD 30,142, and Hispanic or Latino residents a median income of USD 33,310, while the median White income was estimated at USD 46,177 [72]. The Southside neighborhood of Syracuse, where our study takes place, is predominantly home to people of color, particularly Black or African American residents [71]. The 2020 Syracuse Urban Forest Master Plan acknowledges greenspace alignments with higher income and more white neighborhoods and defines strategies and sets goals of “canopy equity” for City’s Division of Forestry [74].
Income and food access infrastructure influence food and nutrition security and, in turn, susceptibility to chronic disease [75]. The prevalence of chronic disease throughout Syracuse varies along neighborhood lines [76]. Model-based estimates of chronic disease rates across the United States were calculated via census tract as part of the 2019 Behavioral Risk Factor Surveillance System (BRFSS). In the census tracts along the SUFFP transect, chronic disease rates are generally higher than the City of Syracuse and the national average (Table 1). Reports also note limited availability of high-quality and full-service markets on the Southside or easy public transportation routes to access such food outside of the area [77]. Of the four census tracts that intersect with the Southside neighborhood (New York tracts 42, 53, 54, and 58), three are classified by the USDA as being food-insecure (Table 1). In other words, populations along the SUFFP transect experience high or higher-than-average rates of diet-related chronic disease, live in low-income households, and have limited access to healthy food needed to address related nutrition deficiencies.
Correlation of poverty, lack of healthy food access, and chronic disease incidence is known [75]. While we have not conclusively established these connections on Syracuse’s Southside, available data indicate that the availability of whole and healthy food alternatives would be welcome.
3.1.3. Forageable Species Selection and Antioxidant Phytochemical Data Collection
Syracuse is in the US Department of Agriculture Plant Hardiness Zone 5b, reflected by lowest annual temperatures of −15 to −10°F [79]. Under a US Forest Service, Great Lakes Restoration Initiative (GLRI) grant, SUFFP is expanding edible tree, shrub, and other ground foliage on public land, including parks and vacant lots. The SUFFP project objectives include modeling the impact of new planting projects on stormwater run-off and nutrition availability.
In 2019 and 2020, field testing in 12 unique sampling areas of upland forest, upland grassland/field, wetland, and riparian eco-zones along the seven-mile transect identified 196 native and non-native edible species growing “wild”, i.e., without standard practices for commercial cultivation or kitchen gardens. In 2020 and 2021, with the intention of a future “wild” edible planting effort, SUFFP assembled a list of plants to augment and complement the available edible species for its GLRI proposal. Plant selection attempted to embrace flora that the local population might already know and recognize, as well as less familiar ones.
In the literature above, plant-based sources of nutrition from “whole food” (i.e., less processed or unprocessed food) are identified for their potential to reduce, synergistically, the risk of chronic disease because of their high sources of antioxidant phytochemicals and the corresponding immune system and anti-inflammatory function. The literature also underscores that there is no perfect “package” of antioxidants, but that the ideal complement varies according to individual and environmental factors that are in constant fluctuation.
3.1.4. Antioxidant Phytochemical Data Collection
For this paper, we focused only on those plants found within the SUFFP transect for which we could find data on a combination of select direct and helper antioxidants (vitamins A, C, and E, selenium, and/or riboflavin) content. Of the 196 identified species, 38 species were selected and organized into five groups: fruits, herbs, nuts, vegetables, and other. For selection, ready availability in the food system, established evidence of effectiveness (as defined in the section above), and a manageable number were considered for this research paper. The primary source for species’ nutrient compositions was the USDA FoodData Central (FDC) database [80]. Strategies to locate missing data points included referrals to agricultural extension offices and outreach to local ethnobotanists and nutrition scientists (see Appendix A Table A1 for a full list and data sources). It is entirely possible that isolated data exist that researchers have not been able to locate. The authors welcome contact with further leads.
Commonly appearing vitamins, A, C, and E, as direct antioxidants, serve as radical scavengers interrupting oxidative stress processes [66,67]. As helper antioxidants, micronutrients such as selenium [68] and riboflavin [69] work as cofactors and co-enzymes for endogenous antioxidant enzymes such as thioredoxin reductase and glutathione peroxidase, respectively, that are involved in free radical scavenging, anti-inflammatory, and immune-boosting processes. Although selenium and riboflavin are not necessarily considered antioxidants, they are important because the activity of endogenous antioxidant enzymes cannot ensue without them.
Cases where possibly comparable food values were not applied. We did not use available antioxidant values for one species of a plant for other related species. For example, we did not apply the butternut values (Juglans cinerea) to heartnut (J. ailantifolia). We could also have used known commercial values for a comparable but different plant. For example, we could have used FDC values for pecan (Carya illinoinensis) to evaluate other hickories, including bitternut (C. cordiformis), pignut (C. glabra), shellbark (C. laciniosa), and shagbark (C. ovata). This project has not adopted that course for several reasons: to emphasize the need for more related nutrition research on wild edibles; the fact that wild edibles differ in unpredictable ways; and in part because the assigned score method discussed above already collapses some level of data.
Cases where possibly comparable food values were applied. Commercially available fruits, vegetables, nuts, and medicinals comprise both native and non-native species, all of which have been genetically altered over time to address size, productivity, taste, etc. Forageable plants in both urban and rural Central New York, as elsewhere, include both native and introduced non-native species, the latter of which might be found in the form of abandoned orchards or invasive but edible undergrowth like Japanese knotweed (Polygonum cuspidatum). For this reason, we have used FDC commercial plant values for forageables like apples (Malus domestica used for Malus spp.), pears (Pyrus communis used for Pyrus spp.), and watercress (Nasturtium officinale).
Comparable food values were also applied to those foods included in FDC data that lack specific species identification. For example, we used the FDC data for non-specific hazelnuts (Corylus spp.) to evaluate American hazelnuts (Corylus americana). This was also performed with gooseberry (Ribes spp. used for Ribes uva-crispa), blackberry (Rubus spp. used for Rubus fruticosus), and black elderberry (Sambucus spp. used for Sambucus canadensis).
3.2. Case Study Analysis
This paper next presents three stages of analysis on the antioxidant potential in the local forageable environment of southside Syracuse. First, we demonstrate the relative power of distinct antioxidants (vitamins A, C, E, selenium, and riboflavin) in forageable species along the Syracuse riparian corridor described above. In Stage 2, we introduce these species in terms of “antioxidant packages” by grouping the distinct antioxidants together (vitamins A + C + E + selenium + riboflavin), ranking the “packages” within organized categories of fruits, herbs, nuts, and vegetables. In Stage 3, we depict the forageable plant foods with the strongest-ranked (or one of the strongest, where equal) “antioxidant package” in each category (fruits, herbs, nuts, vegetables) with the most popular commercial food in each category.
3.2.1. Stage 1 Analysis: Antioxidant Phytochemical Concentration and FDA Daily Value by Individual Plant Species
Plant concentrations per 100 g of vitamins A, C, E, selenium, and riboflavin are re-interpreted through the National Institutes of Health’s (NIH) established recommended allowances for macro- and micronutrients. Recommended Dietary Allowances (RDAs) are designed to guide individuals in planning nutritionally adequate diets. RDAs vary according to age, sex, and pregnancy or lactation status [81]. To simplify public understanding, however, the US Food and Drug Administration (FDA) developed a single Daily Value (DV) per nutrient. It is noteworthy that the DV is based on the RDAs for males aged 19 or older [82], thereby possibly eliding specific needs of females, young males, females in various reproductive stages, and all older persons.
The FDA has also developed a system to identify diverse foods as being high, medium, or low (or no) nutrient sources of individual macro- and micronutrients in terms of their percent DV. As seen in Table 2, a food is a high source of a nutrient if it provides 20% or more of the DV per serving. Medium sources provide 5–20% of a nutrient’s DV per serving. Low sources contain 5% or less of the nutrient DV per serving [83]. “No value” is for foods with zero evidence of the nutrient in the food. Table 2 identifies the high, medium, and low threshold values for vitamins A, C, E, riboflavin, and selenium.
The authors further created assigned scores (0–3) that correspond to the FDA’s high, medium, and low sources for vitamins A, C, and E and selenium and riboflavin. Table 2 displays the DVs for vitamins A, C, E, selenium, and riboflavin and correlates the assigned scores with the FDA nutrient source threshold values. A value of “3” reflects a high nutrient source; “2”, a medium source; and “1”, a low source. Zero (0) describes a dearth of nutrient availability.
In the paper’s Results section, Table 3 reports antioxidant phytochemical values per 100 g of those species for which there are known antioxidant values. Additionally, the tables introduce the antioxidant concentration in terms of high, medium, low, or no concentration value (per FDA protocol) as well as an assigned score (3, 2, 1, 0, respectively, as outlined in Table 2). Plant species are organized from highest to lowest antioxidant concentration per 100 g.
3.2.2. Stage 2 Analysis: Plants Ranked by Power of Composite (or “Packaged”) Antioxidant Scores
In this stage of analysis, antioxidant scores (3, 2, 1, 0) are added together for all forageable plants for which all five reviewed antioxidants were known (Table 4). The resulting composite assigned score reveals the relative potential of diverse plants in terms of the number and power of their constituent complementary antioxidants. Table 4 groups these plants according to type (fruits, herbs, nuts, vegetables, and other) and ranks them within their type using a composite assigned score power.
3.2.3. Stage 3 Analysis: Reflections on Antioxidant Phytochemical Packages of High-Value Forageables and Popular Commercial Plant Foods according to Food Type (Fruits, Herbs, Nuts, and Vegetables)
To assess the contribution of forageable foods to antioxidant phytochemical consumption, Table 5 presents forageable and commercial foods side-by-side according to food type (fruits, herbs, nuts, and vegetables) using (a) the forageable food with the strongest (or one of the strongest, where equal) ranked “antioxidant package” in each food type and (b) the most popular commercial food in the same category. There is one exception. In the vegetables category, we include romaine lettuce (the most popular leafy green) instead of onion (the most popular vegetable overall) to be better portrayed alongside the forageable dandelion greens. The visualization of Table 5 data is further enhanced through the inclusion of radar charts (Figure 2 and Figure 3).
3.2.4. Limitations
The most critical limitation has been the lack of available antioxidant data for forageables. To the extent that nutrient data on wild edibles are at all extant, they are restricted to those plants comparable to commercial products (e.g., black walnuts or mulberries) and occasionally consumed in contemporary diets; they are not reflective of the broad spectrum of plants central to traditional and Indigenous diets.
We further emphasize that available data (e.g., Table 3) have not been validated under Syracuse conditions. Plants adapt to local conditions, affecting nutrient values that can vary according to soil type and quality, USDA plant hardiness zone, and the vast diversity of plant types known as cultivars, among other factors. Selenium (Se) in plant-based foods provides a good example, as availability varies according to the plant’s accumulation ability and the Se content of soils [84,85]. The United States is a Se-rich region with the Se content of most soil samples ranging from 0.1 to 2 mg/Kg and never exceeding 100 mg/kg [84,86]. The mean Se value in Onondaga County, where Syracuse is located, is 0.277 mg/kg [87]. FDC values do not reflect nutrient values with this level of plant specificity. Thus, available antioxidant values must be viewed as best estimates until more regionally and cultivar-specific data can be obtained.
An additional limitation concerns human nutrition impact assessments according to presumed plant quantities consumed. Nutrient availability is notably connected to quantities eaten, which is greatly determined in forageable and commercial plant foods by seasonality, cost, familiarity, accessibility, and preparation methods. Nutrient assessment does not, of course, lead to or predict access and incorporation into the diet. Like nutritious commercial foods, just because a nutritious “wild” food is available does not mean people recognize it, have the means to obtain it, know how to prepare it, or want to eat it. A discussion of the actual incorporation of forageable plants into the diet is outside the scope of this paper but certainly a topic for future investigation. We note that such a discussion must address policy and planning to expand and protect plant availability and safety, as well as to promote related knowledge and training to nutrition educators and diverse food programs. It requires addressing local foraging knowledge (including understanding of the ethics of foraging—see, e.g., Kimmerer 2013 [10]) and rudimentary plant-processing skills. Foraging for auxiliary nutrient support also presumes peoples’ awareness, time, interest, cultural food inclinations and preferences and the availability of foraging and cooking tools, among other factors which must be considered.
4. Results
The Results section reviews vitamins A, C, and E, selenium, and riboflavin concentrations in the edible portions of forageables found growing “wild” or planted as part of the SUFFP transect (Table 3). Table 4 presents composite scores for five nutrients according to whole forageable foods grouped according to fruits, herbs, nuts, vegetables, and others. Table 5 displays the highest-ranking composite scores for forageable foods according to group against the composite score for the most popular commercially available food according to group [88,89]. The radar charts that follow assist with visualizing the nutrient potential of forageables.
In Table 3, vitamin A concentrations (in mcg per 100 g) show values ranging from 1 to 1380 mcg. The DV for vitamin A is 900 mcg, with high sources containing at least 180 mcg per serving. High plant-based sources of vitamin A include leafy green vegetables, orange and yellow vegetables, and tomato products [90]. Our data on forageables show high vitamin A values in grape leaves, lambsquarters, dandelion greens, ground sage, spearmint leaves, and ostrich fern.
Vitamin C concentrations (in mg per 100 g) exhibit values ranging from 0.7 to 181 mg. The DV for vitamin C is 90 mg, with high sources of at least 18mg per serving and tending toward citrus fruits, potatoes, and tomato products [91]. Our data reveal high vitamin C content in blackcurrants, ramps, lambsquarters, quince, and persimmon, among many other vegetables, fruits, and herbs.
Foods naturally high in vitamin E include nuts, seeds, and leafy green vegetables [92]. The DV for vitamin E is 15 mg, with high sources containing at least 3 mg per serving. Our data on forageables (in mg per 100 g) range from 0.70 to 15 mg, with the highest sources identified as hazelnut, ground sage, and dandelion greens.
Table 3 displays selenium concentrations (in mcg per 100 g) ranging from 0 to 18.3 mcg. The DV for selenium is 55 mcg; the high sources contain at least 11 mcg per serving. While foods naturally high in selenium include muscle meats, cereals, and dairy products [85], our data reveal high sources in hackberry, butternut, and black walnut.
Riboflavin concentrations (in mg per 100 g) range in value from 0.02 to 3.54 mg. The DV for riboflavin is 1.3 mg, with high sources containing at least 0.26 mg per serving. Foods naturally high in riboflavin tend toward meats, eggs, and milk, including few vegetables, with rare exceptions like spinach [93]. Nevertheless, our data show high riboflavin sources in Saskatoon serviceberry, lambsquarters, grape leaves, ground sage, and dandelion greens.
As seen in Table 4 and based on the wild foods for which we have complete data, herbs and vegetables have stronger composite antioxidant phytochemical packages than other forageables. The “composite score” communicates strength pertaining to our selected package of antioxidant phytochemicals.
Table 5 displays composite scores for one forageable and one commercial plant in each category (fruits, herbs, nuts, and vegetables). The forageable plants chosen had the highest composite score in each category. The commercial plants selected for the fruits and nuts categories represent the most popular respective commercial consumables in 2021 according to USDA per capita availability [88,89]. Note that the USDA ERS does not maintain data on herbs, so the authors selected basil as the commercial plant in the herb category based on their own understanding of the popularity of herbs in the US. To better illustrate the antioxidant potential of dandelion greens in the vegetables category, the most popular commercial leafy green, romaine lettuce, was selected over the most popular commercial vegetable overall, i.e., the onion.
Table 5 shows that the selected forageable foods within each category have composite scores that are at least on par with (and sometimes better than) popular commercial foods for the same categories. This suggests that forageables can contribute successfully to dietary diversity through the consumption of varied and complementary antioxidant-rich foods.
The radar charts in Figure 2 are based on Table 5. Selected forageables and commercial foods provide a graphic illustration of the assessed composite antioxidant phytochemical profiles. Fuller profiles correspond with higher composite scores. Figure 3 presents the amalgamated antioxidant profiles of forageable and commercial foods, respectively. Measuring the synergistic (and possibly even antagonistic) interactions of these compound profiles is beyond the scope of this paper. Yet based on the literature cited earlier, one can expect that, when eaten together in whole or minimally processed form, complementary and positive antioxidant interactions associated with dietary diversity likely occur.
Composites for the selected foraged foods exhibit analogous or more complete profiles than do the composites for commercially popular foods in the same food category (e.g., with fruits, 2—black mulberry vs. 3—strawberry; with herbs: 2—sage vs. 3—basil). Again, this provides evidence for the potential of forageable foods to contribute to overall dietary diversity, quality, and quantity. Actual accessibility of any or all the selected foods varies dramatically according to many factors, including seasonality, availability in the local outdoors or local supermarkets, and eaters’ familiarity and experience with the different foods.
5. Discussion
Motivated by questions of foraging benefits in and around urban centers, this paper provides evidence of the nutritional value of antioxidant phytochemicals in existing and planned food forests in Syracuse, NY, USA. Our efforts led to an increased understanding of the value of locally available low- and no-cost high-quality dietary fortification against massive population-scale chronic disease prevalence (diabetes, heart disease, etc.). Piecing together this information revealed present and meaningful nutrient density in “wild” foods.
The project also exposed enormous gaps in nutrition data for non-commercial wild forageable foods in the US. While research in other countries on wild edible plants is bountiful [29,94,95,96,97,98,99,100], efforts within the United States are severely limited and warrant future study. The lack of complete data in our study could have been adjusted through the insertion of nutrient values of comparable foods to broaden the available plant data.
Globally, knowledge about nutritional diversity and native plant availability is embedded in local communities and their food systems. These knowledges of survival in, and co-existence with, the uncommercialized natural world is transmitted through generations. They value sustainable and symbiotic relationships between living beings in the environment, both human and non-human. Our findings, however, lead us to recognize and reflect on the lack of community knowledge of nutritional abundance in local food systems. “Modern” food systems rely on human relationships with their markets, undermining the cultural capital gained from reproducing time-tested nutritional well-being through intra-generational foraging practices.
5.1. Available Nutrition Data: Implications of What We Could Find
A variety of forageable plants in Syracuse (fruits, herbs, nuts, vegetables) contain the US FDA Daily Value recommendations of antioxidants A, C, E, selenium, and riboflavin at medium and high levels per 100 g of plant material. Leafy greens and herbs supply high values of vitamin A. Fruits and leafy greens provide high values of vitamin C. Select nuts, herbs, and leafy greens offer the highest concentrations of vitamin E. Some berries and nuts stand out for selenium content. A diversity of fruits, maple syrup, leafy greens, and herbs contain the highest riboflavin scores. This information unmistakably points to the existence of a diverse and untapped availability of nutrients in one region of a nation prone to poor diets and increasing incidence of chronic disease. Although improved diet quality can assist in addressing the underlying bodily dysfunction associated with the national chronic disease emergency, commercially available healthier, less processed, and whole foods cost much more than ultra-processed foods with negligible nutritional value [101]. This limits access to nutrient-dense whole foods by major population segments such as those residing in the Syracuse Southside community.
As noted in the paper’s literature review section, research on antioxidant phytochemicals is an expanding frontier in nutrition science. As much as there is to learn about individual nutrients, even more may yet unfold in investigations of the synergies resulting from nutrient–nutrient as well as nutrient–non-nutrient interactions within food matrices. It is important to recognize that no plant food is made up of only one nutrient, but rather is a complex assembly of nutrients and non-nutrients that interact physically and chemically to exert a myriad of therapeutically beneficial effects [102,103]. From a nutrition perspective, the value and functionality of these bioactives are to a large extent dependent on multiple factors: (a) the form it is consumed in, that is, whole food or as an individual component within a supplement, e.g., a whole orange versus a vitamin C supplement; (b) bio-accessibility, that is, the release of nutrients from the matrix, e.g., the release of minerals such as selenium from fibrous components; (c) the bioavailability of the bioactive, e.g., the availability of the antioxidant vitamins to the systemic circulation for functionality in the body; (d) the bioconversion of the bioavailable nutrient into an active form for functionality, e.g., retinol from provitamin A; (e) the bioactivity or specific effects of a compound in the body, e.g., antioxidant effects; and (f) bio-efficacy, or the production of a desirable or undesirable effect (e.g., radical scavenging activity) by the antioxidant fraction of an ingested, activated bioactive compound [103]. Food bioactive contents within plant foods is influenced by genotype, environmental factors (e.g., soil type, sun exposure, rainfall, climate change), environmental stress (e.g., exposure to light, agronomic factors like organic versus conventional cultivation), and processing (e.g., pasteurization, bleaching, roasting) [104]. This bioactive content is further shaped by the eater’s genetics, heritable predispositions, and environmental contexts of eating [105] that will ultimately impinge on the bioavailability and functionality of these compounds within the eater’s body. These myriad challenges to bioactive contents complicate the delineation of dietary recommendations, as well the predictability of potential interactions between these components in whole foods. Nevertheless, it is reasonable to conclude that if people have access to and consume a diverse plate of nutrient-dense whole foods, several health benefits can be gained through the coordinated activities of their countless components in the long run [58].
In line with available data, we have shown which plants appear to be the best sources of a complement of the studied antioxidants. We acknowledge that plants contain many more antioxidants than the five selected for this study. We also point out that nutrient data vary according to local soils, local climate, and other factors; some of our data did not even originate in the United States (yarrow data, for example, originated in Portugal and Spain). In this study, the best conceptual antioxidant “composite package” was noted in black mulberry among fruits; sage among herbs; black walnut among nuts; and dandelion greens among vegetables. Further, the conceptual forageable plant antioxidant packages present at least as strongly as, or better than, their popular commercial counterparts (strawberry, basil, almond, and romaine lettuce). Forageables present a low-cost, easily accessible opportunity for whole foods with nutrient-dense potential. This is critical, especially given the poor nutritional status nationwide made worse by poverty conditions such as in Syracuse. To that end, public health and social justice-focused policies are called for to address (a) the counter-productive high costs of accessing nutrient-dense whole foods; (b) the expansion of the popular understanding of wild forageables and their contribution to dietary diversity; and (c) the local geographies of healthy food systems. Additionally, but beyond the scope of this paper, municipalities should consider the availability of community commons for secure and tenured publicly accessible food foraging spaces.
5.2. Non-Available Nutrition Data: Implications of What We Could Not Find
Food exists as a human right and the common heritage of all people [106]. As cultural capital, food shapes the health, well-being, and identity of communities. So much indigenous and traditional food and ecological knowledge that sustained the socioeconomic, nutritional, and pharmacological needs of people for eons has been lost to colonization and market-dominated economies [107,108]. Across the globe, Indigenous and local subsistence food systems are marked by traditions transmitted across generations that are instrumental in supporting local food culture and foodways. Enduring local food systems conserve and reproduce native plant varieties and their wild relatives; they offer food diversity and nutritional and medical security to local human and non-human communities while concomitantly maintaining agricultural and genetic biodiversity and environmental sustainability [109,110]. The loss of such systems and knowledges that protect and employ them, endangers food and nutrition security, as well as medical security, and erodes the cultural identity of local communities [111,112,113]. Foraging is an alternate food access strategy that can help (re)build a community’s cultural food capital, in part through restoration of, and edification about, bio-cultural capital [114,115]. Bio-cultural capital arises from regionally and culturally specific knowledge of how people and their local ecology can co-exist and sustain each other [116,117,118]. Bio-cultural capital’s Indigenous foundation confronts and variously absorbs food capitals—from root stock to foraging practices—from migrant, immigrant, and refugee populations [119]. Cultural capital, rooted in food-based relationships with the land, benefits from sustainable food practices, as well as social relationships based on collaboration and trust [120]. Lyson famously wrote about the capacity of local food systems and alternative food exchanges like community-supported agriculture to foster well-being in civil society [121]. Foraging amplifies his concept of “civic agriculture” through its intertwined relationship with nature, its reach for historical and Indigenous knowledges, and its greater disentanglement from the mainstream economy. With ethical foraging and honorable harvests, one gathers with consideration of plant regeneration and other human and non-human needs [10]. These practices, especially on public land, provide not only shared nutrition but also space to exchange bio-cultural information about health, well-being, and building resilience via ensuring food for humans, the animal and plant ecological infrastructure that sustain our environment [16]. When foraging practice wanes, critical ecological and cultural knowledge weakens, and community health becomes compromised.
We question why foods once central to the population’s dietary balance and health are not represented in popular nutrition data access [122]. Our efforts to provide antioxidant phytochemical data as a strategy to showcase the dietary value of “wild” foods play into a modernist mind frame that privileges technical evidence-based knowledge (EBK) over traditional ecological knowledge (TEK) [10,123]. Cultural and Indigenous food traditions nurtured centuries of humans before EBK identified the nutritional profiles of mac ‘n’ cheese and fruit punch. In such an environment TEK struggles for recognition in public policy. EBK data are expensive; the more informal and sometimes oral traditions of TEK cost less, little, or nothing. EBK data have been used largely for research oriented to the commodity scale, nutritional fortification, highly processed food, long shelf life, and the agro-business [124,125,126]. Foraging does not serve the marketplace. EBK and its presumed authority assign value to hundreds of chocolate bar varieties because of the profit margin, but not to the mayapples and chinkapin oak that are not sold in big box stores. The dominance of EBK overwhelms public confidence in TEK. It drives doubt into cultural knowledge and heritage, and it disturbs the sustainable ecological relationship that cultures build with TEK across time.
In presenting our findings, we follow the call of Kimmerer [10] for balanced respect of and support for EBK and TEK in our understanding of the world. Nutritional transition highlights the worldwide phenomenon of shifting patterns of human engagement in the food system. As societies change and food sourcing shifts from local territory to globalized supermarket shelves, they experience specific corresponding nutrition-related trauma of those arrangements [41]. It is true that dietary reliance on the most local of production can leave populations vulnerable to food insecurity and famine caused by routine seasonal and climate fluctuations, interferences with regular plant and animal reproduction, or social upheaval. But the dependency on the global supermarket coexists with conditions of uneven capital accumulation and speculation that have consistently led to polarized wealth and attendant poverty and food insecurity in rich and poor countries alike. Further, global food circulates with global capital and travels best in highly processed, shelf-stable forms that have been linked to the diminished consumption of local and traditional foods, loss of cultural food knowledge, less direct engagement with the food system, co-incidental sedentary lifestyles, and finally an epidemic of chronic disease. The agro-industrial pattern of food consumption and correlated nutritional deficiencies have been growing steadily in the Global North since the latter 19th century. In the Global South, this shift began more recently and is exploding in terms of both global market consumption, chronic disease prevalence, and loss of traditional food and nutritional knowledge in the attempt to “catch up” to the technology-driven (and unevenly distributed) wealth of the Global North.
In summary, we have compiled evidence demonstrating the potential antioxidant contribution of forageable foods to address oxidative stress and inflammation that are major players in the incidence of chronic disease. Where existing data allowed, we introduced a method to profile the theoretical synergistic potential of multiple antioxidants working together in select whole, unprocessed foods. We regret that the available nutrition data on forageables are incomplete, and we regard this as a violation of a public right to information about preventive health and well-being. We question why foods’ “nutritional content” is controlled by market participants who can afford expensive chemical analyses because they are “repaid” in profit. Public policy must prioritize, fund, and promote mechanisms to learn more about the nutritional benefits of wild, forageable foods, as well as expanded and non-commercialized food systems. Such efforts will support community and cultural food capital and help to realize food and nutrition rights associated with empowered community self-reliance.
6. Conclusions
This paper shows that forageable foods in a Central New York State city contain substantial amounts of five key antioxidant phytochemicals, indicating that “wild” foods have the potential to augment local dietary diversity and quality. These findings could be amplified in several ways. Most importantly, USDA and other researchers must expand and report micro- and macronutrient research on non- and less- commercialized forageable foods. Secondly, future studies should consider a review of a broader array of antioxidant phytochemicals, especially as nutritional biochemistry expands our understanding of how these bioactives function. Thirdly, comparable studies should be developed in diverse local ecosystems with alternate selections of forageable foods.
Public and private expansion of urban and rural food forests have implications for not only food and nutrient reserves but also other issues such as urban greening, climate change mitigation, riparian conservation, and sustainability. Support for food forests requires attention to safe and clean water and soils, including monitoring, pesticide-free maintenance, and public notice of risks and safety. Public and professional training is necessary. Nutritionists need expanded datasets for their community nutrition and research repertoire. Nutrition education can raise consumer awareness and encourage the incorporation of more non-commodified foods. Landscape professionals should reconsider design configurations that incorporate edibility. Food studies and food systems experts must magnify their vision of food access beyond commercial food systems, and even beyond backyard and allotment gardens. Public knowledge about and experience with safe and productive foraging needs to be advanced. And perhaps most of all, Indigenous knowledges about ethical foraging and the honorable harvest that sustain wild food ecologies must be respected.
Author Contributions
Conceptualization, A.C.B. and S.R.; writing—original draft preparation, A.C.B., S.R. and E.P.; field data acquisition, A.C.B., M.R.P. and S.A.W.D.; methodology, A.C.B., S.R. and E.P.; writing—review and editing, A.C.B., S.R., E.P., M.R.P. and S.A.W.D.; analysis, A.C.B., S.R. and E.P.; resources, A.C.B., S.R. and E.P.; supervision and project administration, A.C.B.; funding acquisition A.C.B., M.R.P. and S.A.W.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the USDA Forest Service under Grant 22-DG-11094200-028.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki, and approved by the Institutional Review Board (or Ethics Committee) of Syracuse University (protocol code IRB #20-091, approved 24 March 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
For information on data used to support this study, contact Anne C Bellows at [email protected].
Acknowledgments
The Syracuse Urban Food Forest Project has benefited from funding from the US Forest Service, Great Lakes Restoration Initiative. The authors thank Jennifer Wilkins, Moira Tidball, Rick Welsh, and Marguerite Leek for support, review, and constructive critique and Julia Wilson for map design.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

{kind=link}
{kind=link}
{kind=link}
Table A1.
Forageable species on public land on the Syracuse southside including (1) already growing species identified by the Syracuse Urban Food Forest Project (SUFFP; 2019–2021) and (2) SUFFP planted edibles (2021–2022). All Species. (All values are reported per 100 g of “edible part”).
Table A1.
Forageable species on public land on the Syracuse southside including (1) already growing species identified by the Syracuse Urban Food Forest Project (SUFFP; 2019–2021) and (2) SUFFP planted edibles (2021–2022). All Species. (All values are reported per 100 g of “edible part”).
| Scientific Name | Common Name | Other Names | Edible Part | Vit A (RAE mcg) | Vit C (mg) | Vit E (mg) | Se (mcg) | Riboflavin (mg) | Source a |
|---|---|---|---|---|---|---|---|---|---|
| Berries | |||||||||
| Amelanchier alnifolia | saskatoon serviceberry | juneberry, shadbush | raw fruit | 10.91 | 3.55 | 1.12 | 3.54 | [127] | |
| Amelanchier arborea | downy serviceberry | sarvis-berry, shadblow, sugarplum, Indian cherry | fruit | ||||||
| Amelanchier canadensis | Canadian serviceberry | sarvice, wild currant | fruit | ||||||
| Amelanchier laevis | Allegheny serviceberry | fruit | |||||||
| Aronia melanocarpa | black chokeberry | fruit | |||||||
| Celtis occidentalis | hackberry | beaverwood, false elm, nettle tree, sugar berry | raw fruit | 18.3 | [128] | ||||
| Morus alba | white mulberry | raw fruit | 15.2 | 0.088 | [129] | ||||
| Morus nigra | black mulberry | raw fruit | 1 | 36.4 | 0.87 | 0.6 | 0.101 | FDC 169913 | |
| Morus rubra | red mulberry | fruit | |||||||
| Ribes nigrum | blackcurrant | raw fruit | 12 | 181 | 1 | 0.05 | FDC 173963 | ||
| Ribes rubrum | redcurrant | raw fruit | 2 | 41 | 0.1 | 0.6 | 0.05 | FDC 173964 | |
| Ribes uva-crispa | gooseberry | raw fruit | 15 | 27.7 | 0.37 | 0.6 | 0.03 | FDC 173030 | |
| Rubus caesius | European dewberry | fruit | |||||||
| Rubus fruticosus | blackberry | raw fruit | 11 | 21 | 1.17 | 0.4 | 0.026 | FDC 173946 | |
| Rubus idaeus | red raspberry | raw fruit | 2 | 26.4 | 0.57 | 0.08 | FDC 168997 | ||
| Rubus occidentalis | black raspberry | fruit | |||||||
| Sambucus canadensis | black elderberry | flor sauco, tree of music, Danewort, Walewort | raw fruit | 30 | 36 | 0.06 | 0.06 | FDC 171727 | |
| Other Fruits (excluding berries) | |||||||||
| Asimina triloba | pawpaw | false banana, custard apple, white plum | raw fruit | 8.6 | 18.3 | 0.09 | [130] | ||
| Chaenomeles speciosa | flowering quince | raw fruit | 72.15 | [131] | |||||
| Cornus kousa | kousa dogwood | Chinese/Korean/Japanese dogwood | raw fruit | 93 | [132] | ||||
| Diospyros virginiana | persimmon b | possumwood, American ebony, white ebony, bara-bara, boa-wood, butterwood | raw fruit | 66 | FDC 169943 | ||||
| Malus spp. | apple | raw fruit | 3 | 4.6 | 0.18 | 0 | 0.026 | FDC 171688 | |
| Malus spp. | crabapple | raw fruit | 2 | 8 | 0.02 | FDC 171721 | |||
| Podophyllum peltatum | mayapple | Indian apple, wild mandrake, pomme de mai, podophylle pelt | fruit | ||||||
| Prunus americana | American plum | wild plum, August plum, sand cherry, sloe | raw fruit | 173 | 10.3 | 0.53 | 0.042 | FDC 169820 | |
| Prunus avium | sweet cherry | wild cherry | raw fruit | 3 | 7 | 0.07 | 0 | 0.033 | FDC 171719 |
| Prunus tomentosa | nanking cherry | Korean/Chinese cherry | fruit | ||||||
| Pyrus spp. | pear | raw fruit | 1 | 4.3 | 0.12 | 0.1 | 0.026 | FDC 169118 | |
| Viburnum prunifolium | blackhaw | smooth blackhaw, stagbush | fruit | ||||||
| Vitis spp. | grape | raw fruit | 5 | 4 | 0.19 | 0.1 | 0.057 | FDC 174682 | |
| Herbs | |||||||||
| Achillea millefolium | yarrow c | western yarrow, milfoil | leaves | 0.95 | [133] | ||||
| Agastache foeniculum | anise hyssop | blue giant/lavender hyssop | leaves, flowers | ||||||
| Baptisia australis | blue false indigo | plains/rattlebush wild indigo, baptisia, rattlepod | leaves, roots | ||||||
| Ceanothus americanus | New Jersey tea | redroot | roots | ||||||
| Cercis canadensis | eastern redbud | Judas tree | flowers, seed pods | ||||||
| Echinacea purpurea | purple coneflower | Kansas snakeroot, scurvy root, Indian head, comb flower, hedge hog | leaves | ||||||
| Hamamelis ovalis | witch hazel | leaves, twigs | |||||||
| Juniperus spp. | juniper | fruit | |||||||
| Leonurus cardiaca | common motherwort | whole plant | |||||||
| Lindera benzoin | spicebush | wild allspice, Benjaminbush | leaves, twigs, fruits | ||||||
| Melissa officinalis | common balm | lemon balm | leaves | ||||||
| Mentha spicata | spearmint | fresh leaves | 203 | 13.3 | 0.175 | FDC 173475 | |||
| Monarda punctata | beebalm | spotted beebalm, horsemint | leaves, shoot tips, flowers | ||||||
| Picea abies | Norway spruce | needles | |||||||
| Picea glauca | white spruce | Canada spruce, skunk spruce | young tips gum | ||||||
| Picea pungens | blue spruce | white/silver spruce | needles | ||||||
| Rhus spp. | sumac | fruit | |||||||
| Salvia officinalis | sage | ground herb | 295 | 32.4 | 7.48 | 3.7 | 0.336 | FDC 170935 | |
| Symphytum spp. | comfrey | leaves, young shoots, roots | |||||||
| Nuts | |||||||||
| Carya cordiformis | bitternut hickory | nut | |||||||
| Carya glabra | pignut hickory | sweet hickory, smoothbark hickory, switch/switchbud hickory | nut | ||||||
| Carya illinoinensis | pecan | pecan hickory | raw nut | 3 | 1.1 | 1.4 | 3.8 | 0.13 | FDC 170182 |
| Carya laciniosa | shellbark hickory | bigleaf shagbark hickory, kingnut | nut | ||||||
| Carya ovata | shagbark hickory | nut | |||||||
| Castanea dentata | American chestnut | nut | |||||||
| Castanea pumila | Allegheny chinkapin | American/Ozark chinquapin, dwarf chestnut | nut | ||||||
| Corylus americana | hazelnut | American filbert | raw nut | 1 | 6.3 | 15 | 2.4 | 0.113 | FDC 170581 |
| Juglans ailantifolia | heartnut | nut | |||||||
| Juglans cinerea | butternut | white walnut, oilnut | dried nut | 6 | 3.2 | 17.2 | 0.148 | FDC 170570 | |
| Juglans nigra | black walnut | dried nut | 2 | 1.7 | 2.08 | 17 | 0.13 | FDC 170186 | |
| Juglans regia | English walnut | dried nut | 1 | 1.3 | 0.7 | 4.9 | 0.15 | FDC 170187 | |
| Quercus alba | white oak | nut | |||||||
| Quercus coccinea | scarlet oak | nut | |||||||
| Quercus macrocarpa | bur oak | blue oak | nut | ||||||
| Quercus muehlenbergii | chinkapin oak | chestnut/rock chestnut oak | nut | ||||||
| Quercus palustris | pin oak | nut | |||||||
| Quercus rubra | northern red oak | red oak | nut | ||||||
| Quercus velutina | black oak | nut | |||||||
| Vegetables | |||||||||
| Alliaria petiolata | garlic mustard | fresh leaves | 26.1 | [134] | |||||
| Allium tricoccum | ramps | wild leeks | leaves, bulbs | 80 | [135] | ||||
| Amaranthus albus | prostrate pigweed | leaves, stems | |||||||
| Asclepias tuberosa | butterfly milkweed | pleurisy root, orange milkweed, chigger flower | flowers, flower buds, seed pods, root, seed oil | ||||||
| Chenopodium album | lambsquarters | raw vegetable | 580 | 80 | 0.9 | 0.44 | FDC 169244 | ||
| Matteuccia struthiopteris | ostrich fern | fiddlehead fern | raw fronds | 181 | 26.6 | 0.21 | FDC 169405 | ||
| Nasturtium officinale | watercress | fresh leaves | 160 | 43 | 1 | 0.9 | 0.12 | FDC 170068 | |
| Polygonum cuspidatum | Japanese knotweed | shoots, leaves, roots | |||||||
| Portulaca oleracea | purslane | little hogweed | raw vegetable | 21 | 0.9 | 0.112 | FDC 169274 | ||
| Taraxacum spp. | dandelion | raw greens | 508 | 35 | 3.44 | 0.5 | 0.26 | FDC 169226 | |
| Typha latifolia | broadleaf cattail | flags, rushes, bulbrushes | young shoots | 1 | 0.7 | 0.6 | 0.025 | FDC 168994 | |
| Vitis spp. | grape | leaves | 1380 | 11.1 | 2 | 0.9 | 0.354 | FDC 168575 | |
| Other | |||||||||
| Acer saccharum | sugar maple | rock maple, hard maple | sap | 0 | 0 | 0 | 0.6 | 1.27 | FDC 169661 |
| Asarum canadense | wild ginger | Canadian wild ginger, Canadian snakeroot | roots | ||||||
| Robinia pseudoacacia | black locust | false acacia; yellow locust | flowers | ||||||
| Viola spp. | violet | flowers | |||||||
a USDA FoodData Central (FDC) uses unique ID numbers to identify food items [81]; b Persimmon is an example of a food for which the USDA provides values for macronutrients but limited micronutrients. c Yarrow (Achillea millefolium) samples were collected in Spain and Portugal.
References
- Boushey, C.J. Dietary Guidelines for Americans Committee: Process and Updates. Dietary Guidelines for Americans 2020–2025. 2020. Available online: https://mchnutritionpartners.ucla.edu/wp-content/uploads/2021/04/NLN-DGA-Boushey.pdf (accessed on 23 August 2023).
- Nelson, M.E.; Hamm, M.W.; Hu, F.B.; Abrams, S.A.; Griffin, T.S. Alignment of Healthy Dietary Patterns and Environmental Sustainability: A Systematic Review. Adv. Nutr. 2016, 7, 1005–1025. [Google Scholar] [CrossRef]
- Chen, C.; Chaudhary, A.; Mathys, A. Dietary Change Scenarios and Implications for Environmental, Nutrition, Human Health and Economic Dimensions of Food Sustainability. Nutrients 2019, 11, 856. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture (USDA); U.S. Department of Health and Human Services (HHS. Dietary Guidelines for Americans, 2020–2025. 2020. Available online: https://www.dietaryguidelines.gov/sites/default/files/2020-12/Dietary_Guidelines_for_Americans_2020-2025.pdf (accessed on 23 August 2023).
- U.S. Department of Agriculture (USDA); Food and Nutrition Service & Center for Nutrition Policy and Promotion. Average Healthy Eating Index-2015 Scores for All Americans by Age Groups. What We Eat in America, NHANES 2017–2018. 2022. Available online: https://fns-prod.azureedge.us/sites/default/files/media/file/HEIScores_AllAmericans_byAgeGroup_NHANES2017-2018.pdf (accessed on 23 August 2023).
- Fanzo, J.; Davis, C. Global Food Systems, Diets, and Nutrition: Linking Science, Economics, and Policy; Palgrave Macmillan: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Bai, Y. Quantifying the Cost of Nutritious Diets and Dietary Impacts on Health: Economic Approaches to Global Food Systems and Nutrition Transition. Ph.D. Thesis, Tufts University, Gerald J. and Dorothy R. Friedman School of Nutrition Science and Policy, Somerville, MA, USA, 2021. Available online: https://www.proquest.com/docview/2489532200/abstract/30F0C8C12DC64F34PQ/1 (accessed on 23 August 2023).
- Mason, P.; Lang, T. Sustainable Diets: How Ecological Nutrition Can Transform Consumption and the Food System; Routledge, Taylor & Francis Group: London, UK, 2017. [Google Scholar]
- Morland, K.B. (Ed.) Local Food Environments: Food Access in America, 1st ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar] [CrossRef]
- Kimmerer, R.W. Braiding Sweetgrass: Indigenous Wisdom, Scientific Knowledge and the Teachings of Plants, 1st ed.; Milkweed Editions: Minneapolis, MN, USA, 2013. [Google Scholar]
- Khan, S.; Hussain, W.; Sulaiman Shah, S.; Hussain, H.; Altyar, A.E.; Ashour, M.L.; Pieroni, A. Overcoming Tribal Boundaries: The Biocultural Heritage of Foraging and Cooking Wild Vegetables among Four Pathan Groups in the Gadoon Valley, NW Pakistan. Biology 2021, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- Nisbet, C.; Lestrat, K.E.; Vatanparast, H. Food Security Interventions among Refugees around the Globe: A Scoping Review. Nutrients 2022, 14, 522. [Google Scholar] [CrossRef] [PubMed]
- Potteiger, M. Eating Ecologies: Integraging productive ecologies and foraging at the landscape scale. In Localizing Urban Food Strategies. Farming Cities and Performing Rurality; Cinà, G., Dansero, E., Eds.; Politecnico di Torino: Torino, Italy, 2015; pp. 131–145. Available online: http://www.aesoptorino2015.it/content/download/408/2229/version/1/file/10_T1C_672_potteiger_A.pdf (accessed on 23 August 2023).
- McGrath, N. Foraging for Food Security. Thought for Food Blog. 2014. Available online: https://www.ifis.org/blog/2014/food-science-and-technology/foraging-for-food-security (accessed on 23 August 2023).
- Shackleton, C.M.; Hurley, P.T.; Dahlberg, A.C.; Emery, M.R.; Nagendra, H. Urban Foraging: A Ubiquitous Human Practice Overlooked by Urban Planners, Policy, and Research. Sustainability 2017, 9, 10. [Google Scholar] [CrossRef]
- Bukowski, C.; Munsell, J.F. The Community Food Forest Handbook: How to Plan, Organize, and Nurture Edible Gathering Places; Chelsea Green Publishing: London, UK, 2018. [Google Scholar]
- Yoo, S.; Kim, K.; Nam, H.; Lee, D. Discovering Health Benefits of Phytochemicals with Integrated Analysis of the Molecular Network, Chemical Properties and Ethnopharmacological Evidence. Nutrients 2018, 10, 1042. [Google Scholar] [CrossRef] [PubMed]
- Wallinga, D.; Maizes, V. Foraging for healthy food in the global economy: Ten steps we can all take. Explore 2008, 4, 385–388. [Google Scholar] [CrossRef]
- Silveira Rossi, J.L.; Barbalho, S.M.; Reverete de Araujo, R.; Bechara, M.D.; Sloan, K.P.; Sloan, L.A. Metabolic syndrome and cardiovascular diseases: Going beyond traditional risk factors. Diabetes Metab. Res. Rev. 2022, 38, e3502. [Google Scholar] [CrossRef]
- Zhang, Y.-J.; Gan, R.-Y.; Li, S.; Zhou, Y.; Li, A.-N.; Xu, D.-P.; Li, H.-B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 12. [Google Scholar] [CrossRef]
- Forman, H.J.; Zhang, H. Author Correction: Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 8. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Zhang, B.; Deng, Z. The synergistic and antagonistic antioxidant interactions of dietary phytochemical combinations. Crit. Rev. Food Sci. Nutr. 2022, 62, 5658–5677. [Google Scholar] [CrossRef] [PubMed]
- Efferth, T.; Koch, E. Complex interactions between phytochemicals. The multi-target therapeutic concept of phytotherapy. Curr. Drug Targets 2011, 12, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Habtamu, F.G.; Negussie, R. Antinutritional Factors in Plant Foods: Potential Health Benefits and Adverse Effects. Int. J. Nutr. Food Sci. 2014, 3, 284–289. [Google Scholar] [CrossRef]
- Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process Nutr. 2020, 6, 2. [Google Scholar] [CrossRef]
- Säumel, I.; Kotsyuk, I.; Hölscher, M.; Lenkereit, C.; Weber, F.; Kowarik, I. How healthy is urban horticulture in high traffic areas? Trace metal concentrations in vegetable crops from plantings within inner city neighbourhoods in Berlin, Germany. Environ. Pollut. 2012, 165, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Felix, D.; Alcantara, F.; Zaslavsky, I.; Work, A.; Watson, P.L.; Pezzoli, K.; Yu, Q.; Zhu, D.; Scavo, A.J.; et al. Monitoring and mitigation of toxic heavy metals and arsenic accumulation in food crops: A case study of an urban community garden. Plant Direct 2020, 4, e00198. [Google Scholar] [CrossRef] [PubMed]
- Dimitrijević, M.D.; Nujkić, M.M.; Alagić, S.Č.; Milić, S.M.; Tošić, S.B. Heavy metal contamination of topsoil and parts of peach-tree growing at different distances from a smelting complex. Int. J. Environ. Sci. Technol. 2016, 13, 615–630. [Google Scholar] [CrossRef]
- Stark, P.B.; Miller, D.; Carlson, T.J.; De Vasquez, K.R. Open-source food: Nutrition, toxicology, and availability of wild edible greens in the East Bay. PLoS ONE 2019, 14, e0202450. [Google Scholar] [CrossRef]
- Guil, J.L.; Rodrigues-Garcia, I.; Torija, E. Nutritional and toxic factors in selected wild edible plants. Plant Foods Hum. Nutr. 1997, 51, 99–107. [Google Scholar] [CrossRef]
- Samsøe-Petersen, L.; Larsen, E.H.; Larsen, P.B.; Bruun, P. Uptake of Trace Elements and PAHs by Fruit and Vegetables from Contaminated Soils. Environ. Sci. Technol. 2002, 36, 3057–3063. [Google Scholar] [CrossRef]
- Von Hoffen, L.P.; Säumel, I. Orchards for edible cities: Cadmium and lead content in nuts, berries, pome and stone fruits harvested within the inner city neighbourhoods in Berlin, Germany. Ecotoxicol. Environ. Saf. 2014, 101, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Zeiner, M.; Cindrić, I. Harmful Elements (Al, Cd, Cr, Ni, and Pb) in Wild Berries and Fruits Collected in Croatia. Toxics 2018, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Dabady, S.; Stark, P.B. Urban Foraging in Municipal Parks and Public Schools: Opportunities for Policymakers. Berkley Food Institute. 2017. Available online: https://forage.berkeley.edu/wp-content/uploads/2017/08/UrbanForagingPolicyBrief-2017-08-29.pdf (accessed on 4 October 2023).
- Di Renzo, L.; Gualtieri, P.; De Lorenzo, A. Diet, Nutrition and Chronic Degenerative Diseases. Nutrients 2021, 13, 1372. [Google Scholar] [CrossRef]
- Bilali, H.E.; Allahyari, M.S. Transition towards sustainability in agriculture and food systems: Role of information and communication technologies. Inf. Process. Agric. 2018, 5, 456–464. [Google Scholar] [CrossRef]
- Hawkes, C. Uneven dietary development: Linking the policies and processes of globalization with the nutrition transition, obesity and diet-related chronic diseases. Glob. Health 2006, 2, 4. [Google Scholar] [CrossRef]
- Monteiro, C.A.; Cannon, G. The Impact of Transnational “Big Food” Companies on the South: A View from Brazil. PLoS Med. 2012, 9, e1001252. [Google Scholar] [CrossRef]
- Baker, P.; Friel, S. Food systems transformations, ultra-processed food markets and the nutrition transition in Asia. Glob. Health 2016, 12, 80. [Google Scholar] [CrossRef] [PubMed]
- Willey, J.; Wakefield, M.; Silver, H.J. Exploring the Diets of Adults with Obesity and Type II Diabetes from Nine Diverse Countries: Dietary Intakes, Patterns, and Quality. Nutrients 2020, 12, 2027. [Google Scholar] [CrossRef]
- Raj, S. Influences of the Nutrition Transition on Chronic Disease. In Integrative and Functional Medical Nutrition Therapy: Principles and Practices; Noland, D., Drisko, J.A., Wagner, L., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 17–29. [Google Scholar] [CrossRef]
- Mapesi, H.; Paris, D.H. Non-Communicable Diseases on the Rise in Sub-Saharan Africa, the Underappreciated Threat of a Dual Disease Burden. Praxis 2019, 108, 997–1005. [Google Scholar] [CrossRef]
- Kopp, W. How Western Diet And Lifestyle Drive The Pandemic Of Obesity And Civilization Diseases. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 2221–2236. [Google Scholar] [CrossRef]
- Lechner, K.; von Schacky, C.; McKenzie, A.L.; Worm, N.; Nixdorff, U.; Lechner, B.; Kränkel, N.; Halle, M.; Krauss, R.M.; Scherr, J. Lifestyle factors and high-risk atherosclerosis: Pathways and mechanisms beyond traditional risk factors. Eur. J. Prev. Cardiol. 2020, 27, 394–406. [Google Scholar] [CrossRef]
- Perrone, S.; Tataranno, M.L.; Santacroce, A.; Bracciali, C.; Riccitelli, M.; Alagna, M.G.; Longini, M.; Belvisi, E.; Bazzini, F.; Buonocore, G. Fetal Programming, Maternal Nutrition, and Oxidative Stress Hypothesis. J. Pediatr. Biochem. 2016, 6, 96–102. [Google Scholar] [CrossRef]
- Kuiper-Makris, C.; Selle, J.; Nüsken, E.; Dötsch, J.; Alejandre Alcazar, M.A. Perinatal Nutritional and Metabolic Pathways: Early Origins of Chronic Lung Diseases. Front. Med. 2021, 8, 667315. [Google Scholar] [CrossRef] [PubMed]
- Carpinello, O.J.; DeCherney, A.H.; Hill, M.J. Developmental Origins of Health and Disease: The History of the Barker Hypothesis and Assisted Reproductive Technology. Semin. Reprod. Med. 2018, 36, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, V.; Cardoso, R.C.; Puttabyatappa, M. Developmental Programming, a Pathway to Disease. Endocrinology 2016, 157, 1328–1340. [Google Scholar] [CrossRef]
- MacDonald Baker, S. The Metaphor of Oceanic Disease. Integr. Med. 2008, 7, 40–45. [Google Scholar]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Dayem, A.A.; Cho, S.G. Correlation between Oxidative Stress, Nutrition, and Cancer Initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed]
- Bosma-den Boer, M.M.; van Wetten, M.-L.; Pruimboom, L. Chronic inflammatory diseases are stimulated by current lifestyle: How diet, stress levels and medication prevent our body from recovering. Nutr. Metab. 2012, 9, 32. [Google Scholar] [CrossRef]
- Li, Q.-F.; Hao, H.; Tu, W.-S.; Guo, N.; Zhou, X.-Y. Maresins: Anti-inflammatory pro-resolving mediators with therapeutic potential. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 7442–7453. [Google Scholar] [CrossRef] [PubMed]
- Liska, D.J. The detoxification enzyme systems. Altern. Med. Rev. A J. Clin. Ther. 1998, 3, 187–198. [Google Scholar]
- Palrasu, M.; Siddavaram, N. Cytochrome P450 Structure, Function and Clinical Significance: A Review. Curr. Drug Targets 2018, 18, 38–54. [Google Scholar] [CrossRef]
- Pingitore, A.; Lima, G.P.P.; Mastorci, F.; Quinones, A.; Iervasi, G.; Vassalle, C. Exercise and oxidative stress: Potential effects of antioxidant dietary strategies in sports. Nutrition 2015, 31, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D.R.; Gross, M.D.; Tapsell, L.C. Food synergy: An operational concept for understanding nutrition. Am. J. Clin. Nutr. 2009, 89, 1543S–1548S. [Google Scholar] [CrossRef] [PubMed]
- Barba, F. (Ed.) Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Woodhead Publishing: Sawston, UK, 2019. [Google Scholar]
- Petroski, W.; Minich, D.M. Is There Such a Thing as “Anti-Nutrients”? A Narrative Review of Perceived Problematic Plant Compounds. Nutrients 2020, 12, E2929. [Google Scholar] [CrossRef]
- Foyer, C.H.; Kyndt, T.; Hancock, R.D. Vitamin C in Plants: Novel Concepts, New Perspectives, and Outstanding Issues. Antioxid. Redox Signal. 2020, 32, 463–485. [Google Scholar] [CrossRef]
- Webb, G.P. Nutritional supplements and conventional medicine; what should the physician know? Proc. Nutr. Soc. 2007, 66, 471–478. [Google Scholar] [CrossRef]
- Aune, D.; Keum, N.; Giovannucci, E.; Fadnes, L.T.; Boffetta, P.; Greenwood, D.C.; Tonstad, S.; Vatten, L.J.; Riboli, E.; Norat, T. Dietary intake and blood concentrations of antioxidants and the risk of cardiovascular disease, total cancer, and all-cause mortality: A systematic review and dose-response meta-analysis of prospective studies. Am. J. Clin. Nutr. 2018, 108, 1069–1091. [Google Scholar] [CrossRef]
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2016, 22, 6701–6715. [Google Scholar] [CrossRef]
- Augimeri, G.; Montalto, F.I.; Giordano, C.; Barone, I.; Lanzino, M.; Catalano, S.; Andò, S.; De Amicis, F.; Bonofiglio, D. Nutraceuticals in the Mediterranean Diet: Potential Avenues for Breast Cancer Treatment. Nutrients 2021, 13, 2557. [Google Scholar] [CrossRef] [PubMed]
- Grosso, G. Dietary Antioxidants and Prevention of Non-Communicable Diseases. Antioxidants 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, N.; Victor, V.M. Molecular strategies for targeting antioxidants to mitochondria: Therapeutic implications. Antioxid. Redox Signal. 2015, 22, 686–729. [Google Scholar] [CrossRef] [PubMed]
- Kiełczykowska, M.; Kocot, J.; Paździor, M.; Musik, I. Selenium—A fascinating antioxidant of protective properties. Adv. Clin. Exp. Med. Off. Organ. Wroc. Med. Univ. 2018, 27, 245–255. [Google Scholar] [CrossRef]
- Suwannasom, N.; Kao, I.; Pruß, A.; Georgieva, R.; Bäumler, H. Riboflavin: The Health Benefits of a Forgotten Natural Vitamin. Int. J. Mol. Sci. 2020, 21, E950. [Google Scholar] [CrossRef]
- Our City. In 2020 State of The City, Syracuse Mayor Ben Walsh Sees “Window of Opportunity” to Revitalize Neighborhoods and Continue Progress on Syracuse Surge. Our City. 2020. Available online: https://ourcity.syrgov.net/2020/01/in-2020-state-of-the-city-syracuse-mayor-ben-walsh-sees-window-of-opportunity-to-revitalize-neighborhoods-and-continue-progress-on-syracuse-surge/ (accessed on 23 August 2023).
- U.S. Census Bureau. 2020 Census Redistricting Data (Public Law 94-171). United States Census Bureau. 2020. Available online: https://data.census.gov/cedsci/table?q=Syracuse%20city,%20New%20York&tid=DECENNIALPL2020.P1 (accessed on 23 August 2023).
- U.S. Census Bureau. 2020 American Community Survey 5-Year Estimates. United States Census Bureau. 2020. Available online: https://data.census.gov/cedsci/all?q=american%20community%20survey (accessed on 23 August 2023).
- Jargowsky, P.A. The Architecture of Segregation: Civil Unrest, the Concentration of Poverty, and Public Policy. The Century Foundation. 2015. Available online: https://tcf.org/content/report/architecture-of-segregation/ (accessed on 23 August 2023).
- Davey Resource Group, Inc. Syracuse Urban Forest Master Plan; Forestry Department City of Syracuse: Syracuse, NY, USA, 2020.
- Pine, A. Ambient struggling: Food, chronic disease, and spatial isolation among the urban poor. Agric. Hum. Values 2022, 40, 1105–1116. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. PLACES: Local Data for Better Health. 2022. Available online: https://www.cdc.gov/places/index.html (accessed on 23 August 2023).
- Potteiger, M.; Weissman, E. FoodPlanCNY. 2021. Available online: https://agriculture.ongov.net/wp-content/uploads/2021/03/Food-Plan-CNY.pdf (accessed on 23 August 2023).
- Economic Research Service (ERS); U.S. Department of Agriculture (USDA). Food Access Research Atlas. 2019. Available online: https://www.ers.usda.gov/data-products/food-access-research-atlas/ (accessed on 23 August 2023).
- United States Department of Agriculture. USDA Plant Hardiness Zone Map. USDA. 2012. Available online: https://planthardiness.ars.usda.gov/ (accessed on 23 August 2023).
- United States Department of Agriculture, Agricultural Research Service. FoodData Central. 2019. Available online: https://fdc.nal.usda.gov/ (accessed on 23 August 2023).
- National Institutes of Health Office of Dietary Supplements. Nutrient Recommendations and Databases. National Institutes of Health, n.d. Available online: https://ods.od.nih.gov/HealthInformation/nutrientrecommendations.aspx (accessed on 23 August 2023).
- Food and Drug Administration. Food Labeling: Revision of the Nutrition and Supplement Facts Labels and Serving Sizes of Foods That Can Reasonably Be Consumed at One Eating Occasion; Dual-Column Labeling; Updating, Modifying, and Establishing Certain Reference Amounts Customarily Consumed; Serving Size for Breath Mints; and Technical Amendments; Proposed Extension of Compliance Dates. 2017. Available online: https://www.federalregister.gov/documents/2017/10/02/2017-21019/food-labeling-revision-of-the-nutrition-and-supplement-facts-labels-and-serving-sizes-of-foods-that (accessed on 23 August 2023).
- Center for Food Safety and Applied Nutrition. Daily Value on the New Nutrition and Supplement Facts Labels. U.S. Food and Drug Administration. FDA. 2022. Available online: https://www.fda.gov/food/new-nutrition-facts-label/daily-value-new-nutrition-and-supplement-facts-labels (accessed on 23 August 2023).
- National Research Council (US) Subcommittee on Selenium. Distribution. In Selenium in Nutrition: Revised Edition; National Academies Press (US): Washington, DC, USA, 1983. Available online: https://www.ncbi.nlm.nih.gov/books/NBK216733/ (accessed on 23 August 2023).
- National Institutes of Health Office of Dietary Supplements. Selenium: Fact Sheet for Health Professionals. National Institutes of Health. 2021. Available online: https://ods.od.nih.gov/factsheets/Selenium-HealthProfessional/ (accessed on 23 August 2023).
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2017, 7, 2074. Available online: https://www.frontiersin.org/articles/10.3389/fpls.2016.02074 (accessed on 23 August 2023). [CrossRef]
- United States Geological Survey. Average concentrations of elements in Onondaga County, New York. United States Geological Survey. 2008. Available online: https://mrdata.usgs.gov/geochem/county.php?place=f36067&el=Se&rf=northeastern (accessed on 23 August 2023).
- Economic Research Service (ERS). Vegetables, Fresh Market (Including Mushrooms): Selected U.S. Per Capita Availability; [Data set]; USDA Economic Research Service: Washington, DC, USA, 2022.
- Economic Research Service (ERS). U.S. Per Capita Use of Selected, Commercially Produced, Fresh, and Processing Fruit and Tree Nuts, 1976 to Date; [Data set]; USDA Economic Research Service: Washington, DC, USA, 2022.
- National Institutes of Health Office of Dietary Supplements. Vitamin A and Carotenoids: Fact Sheet for Health Professionals. National Institutes of Health. 2022. Available online: https://ods.od.nih.gov/factsheets/VitaminA-HealthProfessional/ (accessed on 23 August 2023).
- National Institutes of Health Office of Dietary Supplements. Vitamin C: Fact Sheet for Health Professionals. National Institutes of Health. 2021. Available online: https://ods.od.nih.gov/factsheets/VitaminC-HealthProfessional/ (accessed on 23 August 2023).
- National Institutes of Health Office of Dietary Supplements. Vitamin E: Fact Sheet for Health Professionals. National Institutes of Health. 2021. Available online: https://ods.od.nih.gov/factsheets/VitaminE-HealthProfessional/ (accessed on 23 August 2023).
- National Institutes of Health Office of Dietary Supplements. Riboflavin: Fact Sheet for Health Professionals. National Institutes of Health. 2022. Available online: https://ods.od.nih.gov/factsheets/Riboflavin-HealthProfessional/ (accessed on 23 August 2023).
- Ao, T.; Deb, C.R. Nutritional and antioxidant potential of some wild edible mushrooms of Nagaland, India. J. Food Sci. Technol. 2019, 56, 1084–1089. [Google Scholar] [CrossRef]
- Duguma, H.T. Wild Edible Plant Nutritional Contribution and Consumer Perception in Ethiopia. Int. J. Food Sci. 2020, 2020, e2958623. [Google Scholar] [CrossRef]
- Motti, R. Wild Edible Plants: A Challenge for Future Diet and Health. Plants 2022, 11, 344. [Google Scholar] [CrossRef]
- Pinela, J.; Carvalho, A.M.; Ferreira, I.C.F.R. Wild edible plants: Nutritional and toxicological characteristics, retrieval strategies and importance for today’s society. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 110, 165–188. [Google Scholar] [CrossRef] [PubMed]
- Queirós, B.; Barreira, J.C.M.; Sarmento, A.C.; Ferreira, I.C.F.R. In search of synergistic effects in antioxidant capacity of combined edible mushrooms. Int. J. Food Sci. Nutr. 2009, 60, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Tharmabalan, R.T. Nutritional Analysis of Five Wild Edible Vegetables Traditionally Consumed by the Orang Asli in Perak. Int. J. Food Sci. 2021, 8823565. [Google Scholar] [CrossRef]
- Ullah, I.; Gul, S.; Rehman, H.U.; Ahmad, N.; Ullah, I.; Jawad, S.M.; Ali, J.; Ahmad, A.; Akbar, M.U. Analysis of nutrients and minerals of some wild edible plants. Int. J. Fauna Biol. Stud. 2017, 4, 35–39. [Google Scholar]
- Monteiro, C.A. Nutrition and health. The issue is not food, nor nutrients, so much as processing. Public. Health Nutr. 2009, 12, 729–731. [Google Scholar] [CrossRef] [PubMed]
- Crowe, K. Designing Functional Foods with Bioactive Polyphenols: Highlighting Lessons Learned from Original Plant Matrices. 2014. Available online: https://www.semanticscholar.org/paper/Designing-Functional-Foods-with-Bioactive-%3A-Lessons-Crowe/dea27cdd60e5ebf8319d3651dac58e298aafb033 (accessed on 23 August 2023).
- Aguilera, J.M. The food matrix: Implications in processing, nutrition and health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef]
- Bolling, B.W.; Chen, C.-Y.O.; McKay, D.L.; Blumberg, J.B. Tree nut phytochemicals: Composition, antioxidant capacity, bioactivity, impact factors. A systematic review of almonds, Brazils, cashews, hazelnuts, macadamias, pecans, pine nuts, pistachios and walnuts. Nutr. Res. Rev. 2011, 24, 244–275. [Google Scholar] [CrossRef]
- Virgen-Carrillo, C.A.; Martínez Moreno, A.G.; Valdés Miramontes, E.H. Potential Hypoglycemic Effect of Pomegranate Juice and Its Mechanism of Action: A Systematic Review. J. Med. Food 2020, 23, 1–11. [Google Scholar] [CrossRef]
- Bellows, A.C.; Valente, F.L.S.; Lemke, S.; de Lara, M.D.N.B. (Eds.) Gender, Nutrition, and the Human Right to Adequate Food: Toward an Inclusive Framework; Routledge: London, UK, 2017. [Google Scholar] [CrossRef]
- Kurashima, N.; Jeremiah, J.; Ticktin, T.; Ka Wā, I. Ma Mua: The Value of a Historical Ecology Approach to Ecological Restoration in Hawaii 1. Pac. Sci. 2017, 71, 437–456. [Google Scholar] [CrossRef]
- Waldman, J. On natural abundances: Remembering not to forget. In Running Silver: Restoring Atlantic Rivers and Their Great Fish Migrations, 1st ed.; Lyons Press: Lyon, France, 2013; pp. 63–71. [Google Scholar]
- Ojha, S.N.; Anand, A.; Sundriyal, R.C.; Arya, D. Traditional Dietary Knowledge of a Marginal Hill Community in the Central Himalaya: Implications for Food, Nutrition, and Medicinal Security. Front. Pharmacol. 2021, 12, 789360. [Google Scholar] [CrossRef]
- Johns, T.; Sthapit, B.R. Biocultural Diversity in the Sustainability of Developing-Country Food Systems. Food Nutr. Bull. 2004, 25, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Spring, A.; Neyelle, M.; Bezha, W.; Simmons, D.; Blay-Palmer, A. Learning from the past to deal with the future: Using different knowledges to ensure food security in the Tsá Tué biosphere reserve (Northwest Territories, Canada). Front. Sustain. Food Syst. 2023, 6, 984290. [Google Scholar] [CrossRef]
- Biedenweg, K.; Harguth, H.; Stiles, K. The science and politics of human well-being: A case study in cocreating indicators for Puget Sound restoration. Ecol. Soc. 2017, 22, 11. [Google Scholar] [CrossRef]
- Reyes-García, V.; Paneque-Gálvez, J.; Luz, A.C.; Gueze, M.; Macía, M.J.; Orta-Martínez, M.; Pino, J. Cultural change and traditional ecological knowledge. An empirical analysis from the Tsimane’ in the Bolivian Amazon. Hum. Organ. 2014, 73, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Pape, T. Futuristic restoration as a policy tool for environmental justice objectives. Restor. Ecol. 2022, 30, e13629. [Google Scholar] [CrossRef]
- Pilgrim, S.; Samson, C.; Pretty, J. Rebuilding Lost Connections: How Revitalisation Projects Contribute to Cultural Continuity and Improve the Environment; Semantic Scholar: Seattle, WA, USA, 2023. [Google Scholar]
- Maillard, K.N. On Remote Farms and in City Gardens, a Native American Movement Grows. The New York Times. 26 August 2022. Available online: https://www.nytimes.com/2022/08/26/dining/native-american-agriculture.html (accessed on 23 August 2023).
- Lang, M. Buen vivir as a territorial practice. Building a more just and sustainable life through interculturality. Sustain. Sci. 2022, 17, 1287–1299. [Google Scholar] [CrossRef]
- Kimmerer, R.W. Restoration and Reciprocity: The Contributions of Traditional Ecological Knowledge to the Philosophy and Practice of Ecological Restoration. In Human Dimensions of Ecological Restoration: Integrating Science, Nature, and Culture; Egan, D., Hjerpe, E.E., Abrams, J., Eds.; Island Press: Washington, DC, USA, 2011; pp. 257–276. [Google Scholar]
- Gingell, T.; Murray, K.; Correa-Velez, I.; Gallegos, D. Determinants of food security among people from refugee backgrounds resettled in high-income countries: A systematic review and thematic synthesis. PLoS ONE 2022, 17, e0268830. [Google Scholar] [CrossRef]
- Pasquier Merino, A.G.; Torres Salcido, G.; Monachon, D.S.; Villatoro Hernández, J.G. Alternative Food Networks, Social Capital, and Public Policy in Mexico City. Sustainability 2022, 14, 23. [Google Scholar] [CrossRef]
- Lyson, T.A. Civic Agriculture: Reconnecting Farm, Food, and Community; University Press of New England: Lebanon, NH, USA, 2004; Available online: https://press.uchicago.edu/ucp/books/book/distributed/C/bo44309143.html (accessed on 23 August 2023).
- Tidball, M.M.; Tidball, K.G.; Curtis, P. The Absence of Wild Game and Fish Species from the USDA National Nutrient Database for Standard Reference: Addressing Information Gaps in Wild Caught Foods. Ecol. Food Nutr. 2014, 53, 142–148. [Google Scholar] [CrossRef]
- Little Bear, L. Jagged Worldviews Colliding. In Reclaiming Indigenous Voice and Vision; Battiste, M., Ed.; University of British Columbia Press: Vancouver, BC, Canada, 2000; Available online: https://press.uchicago.edu/ucp/books/book/distributed/R/bo69994495.html (accessed on 23 August 2023).
- Nestle, M. Unsavory Truth, 1st ed.; Basic Books: New York, NY, USA, 2018. [Google Scholar]
- Scrinis, G. Nutritionism: The Science and Politics of Dietary Advice. In Nutritionism; Columbia University Press: Vancouver, BC, Canada, 2013. [Google Scholar] [CrossRef]
- Scrinis, G. Ultra-processed foods and the corporate capture of nutrition-an essay by Gyorgy Scrinis. BMJ 2020, 371, m4601. [Google Scholar] [CrossRef]
- Mazza, G. Compositional and Functional Properties of Saskatoon Berry and Blueberry. Int. J. Fruit. Sci. 2005, 5, 101–120. [Google Scholar] [CrossRef]
- Demır, F.; Doğan, H.; Özcan, M.; Haciseferoğullari, H. Nutritional and physical properties of hackberry (Celtis australis L.). J. Food Eng. 2002, 54, 241–247. [Google Scholar] [CrossRef]
- Imran, M.; Khan, H.; Shah, M.; Khan, R.; Khan, F. Chemical composition and antioxidant activity of certain Morus species. J. Zhejiang Univ. Sci. B 2010, 11, 973–980. [Google Scholar] [CrossRef]
- Peterson, R.N.; Cherry, J.P.; Simmons, J.G. Composition of pawpaw (Asimina triloba) fruit [Flavor and aroma, eastern United States]. Annu. Rep. North. Nut Grow. Assoc. 1982, 77, 97–106. [Google Scholar]
- Yang, L.; Ahmed, S.; Stepp, J.R.; Zhao, Y.; Zeng, M.J.; Pei, S.; Xue, D.; Xu, G. Cultural Uses, Ecosystem Services, and Nutrient Profile of Flowering Quince (Chaenomeles speciosa) in the Highlands of Western Yunnan, China. Econ. Bot. 2015, 69, 273–283. [Google Scholar] [CrossRef]
- Schmitzer, V.; Sircelj, H.; Stampar, F.; Slatnar, A. Physico-chemical characterization of Cornus kousa Burg. Fruit: Determining optimal maturity for fresh consumption. J. Sci. Food Agric. 2021, 101, 778–785. [Google Scholar] [CrossRef]
- Dias, M.I.; Barros, L.; Dueñas, M.; Pereira, E.; Carvalho, A.M.; Alves, R.C.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Chemical composition of wild and commercial Achillea millefolium L. and bioactivity of the methanolic extract, infusion and decoction. Food Chem. 2013, 141, 4152–4160. [Google Scholar] [CrossRef] [PubMed]
- Guil-Guerrero, J.L.; Giménez-Martínez, J.J.; Torija-Isasa, M.E. Nutritional Composition of Wild Edible Crucifer Species. J. Food Biochem. 1999, 23, 283–294. [Google Scholar] [CrossRef]
- Zennie, T.; Ogzewalla, D. Ascorbic acid and Vitamin A content of edible wild plants of Ohio and Kentucky. Econ. Bot. 1977, 31, 76–79. [Google Scholar] [CrossRef]
Figure 1.
Syracuse Urban Food Forest Project (SUFFP), Riparian Forest Corridor along Furnace Brook and Onondaga Creek on Southside of Syracuse, NY.
Figure 1.
Syracuse Urban Food Forest Project (SUFFP), Riparian Forest Corridor along Furnace Brook and Onondaga Creek on Southside of Syracuse, NY.
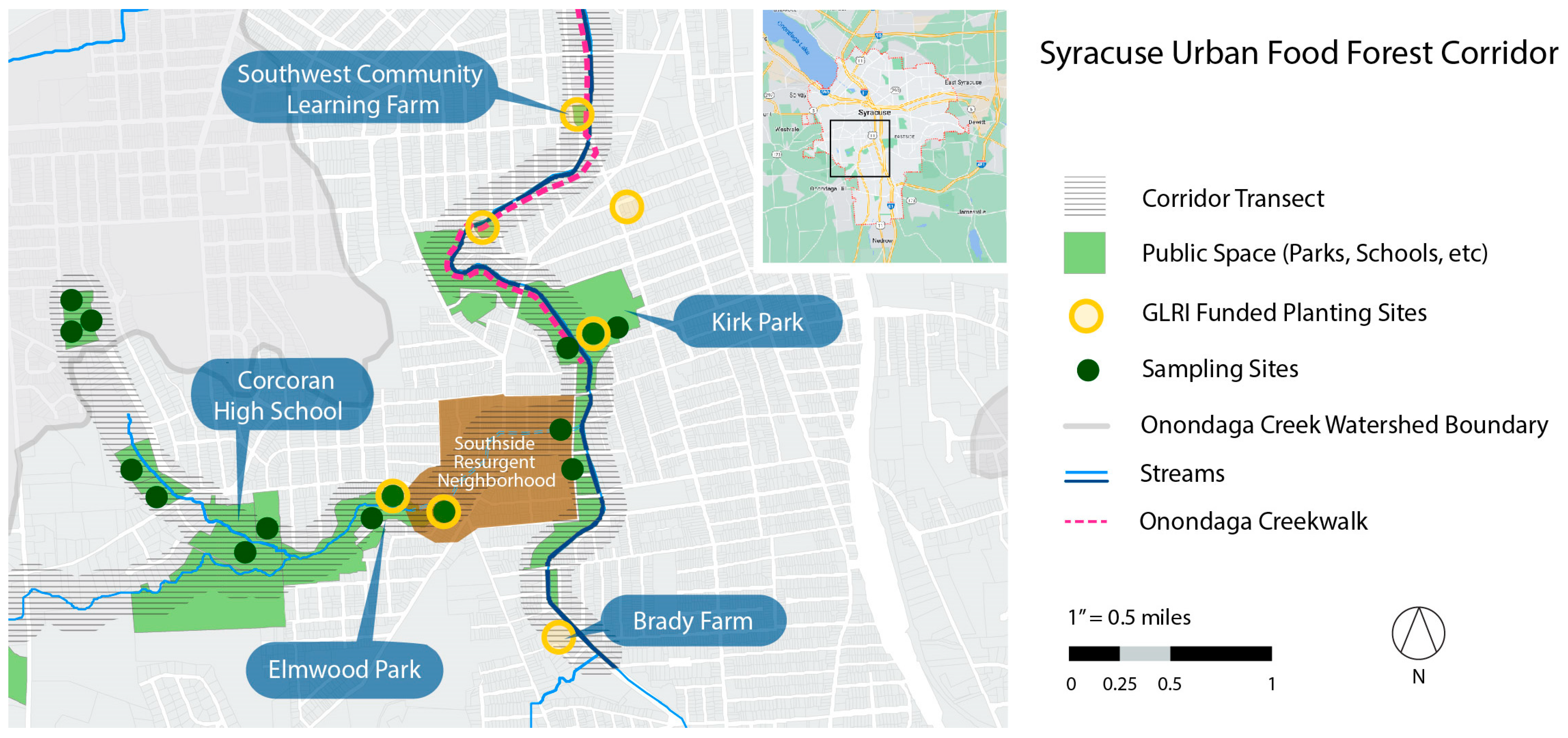
Figure 2.
Radar charts of antioxidant scores of select forageable and commercial plant foods. Note: The individual antioxidants are shown on the “spoke” axes. The antioxidant “scores” (0–3), representative of the FDA’s classification of foods as no-concentration to high sources of an antioxidant, are shown on the “ring” axes.
Figure 2.
Radar charts of antioxidant scores of select forageable and commercial plant foods. Note: The individual antioxidants are shown on the “spoke” axes. The antioxidant “scores” (0–3), representative of the FDA’s classification of foods as no-concentration to high sources of an antioxidant, are shown on the “ring” axes.
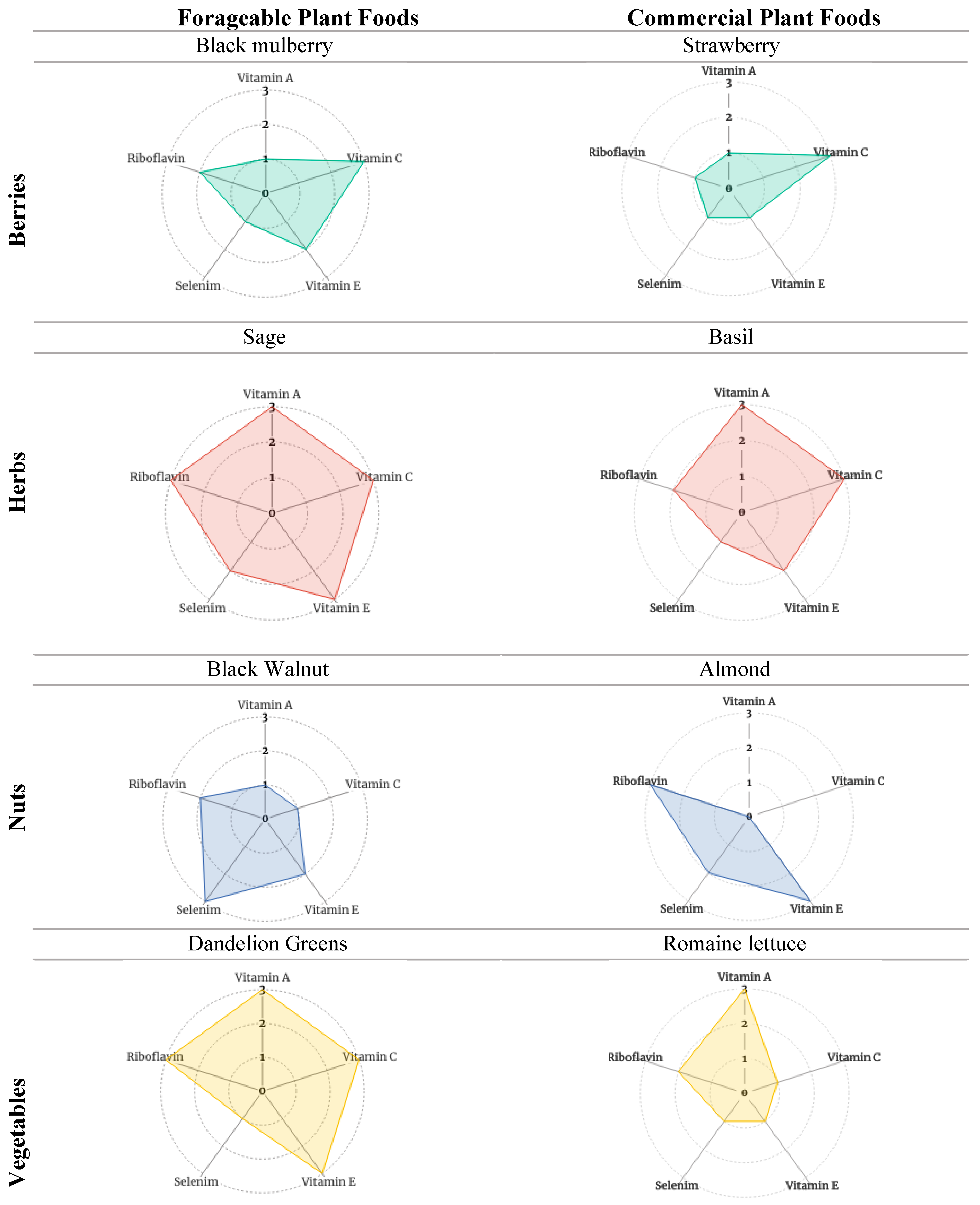
Figure 3.
Combined antioxidant score profiles for select forageable and commercial plant foods Note: The individual antioxidants are shown on the “spoke” axes. The antioxidant “scores” (0–3), representative of the FDA’s classification of foods as no-concentration to high sources of an antioxidant, are shown on the “ring” axes.
Figure 3.
Combined antioxidant score profiles for select forageable and commercial plant foods Note: The individual antioxidants are shown on the “spoke” axes. The antioxidant “scores” (0–3), representative of the FDA’s classification of foods as no-concentration to high sources of an antioxidant, are shown on the “ring” axes.
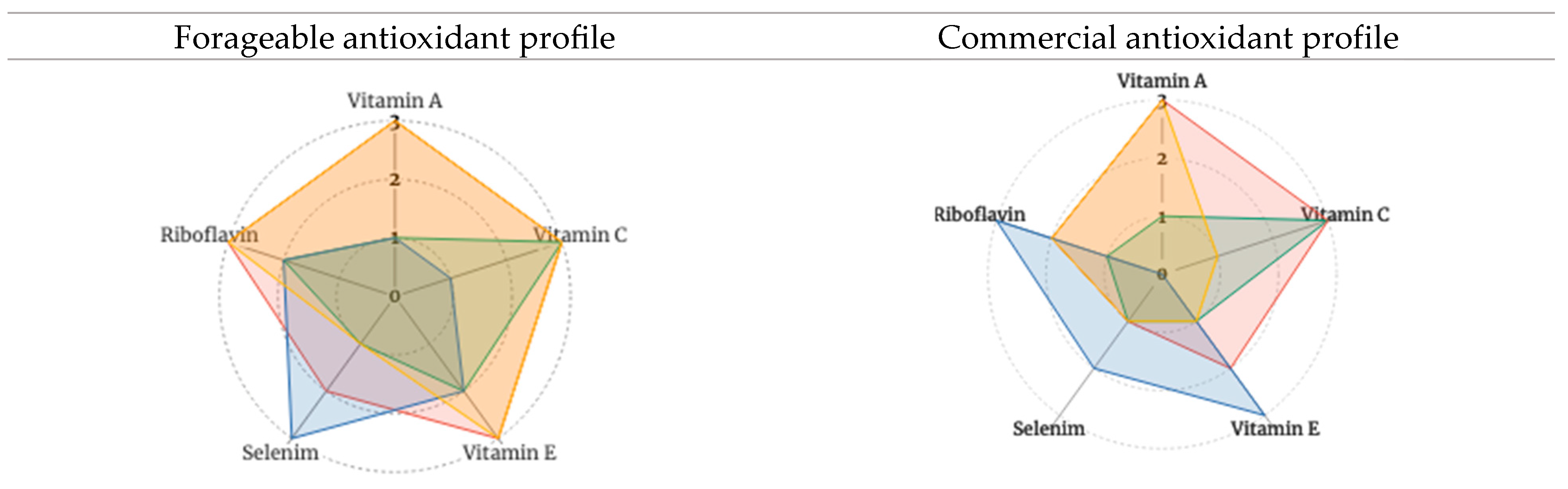
Table 1.
Chronic Disease Rates and Food Insecurity.
| Location | Cancer a (%) | Coronary Heart Disease (%) | Diabetes (%) | High Blood Pressure (%) | Kidney Disease (%) | Obesity (%) | Food Insecurity b LI = Low-Income LA = Low-Access c | |
|---|---|---|---|---|---|---|---|---|
| United States | 7.5 | 8.2 | 12.7 | 37.1 | 3.5 | 36.0 | ||
| Onondaga County | 7.4 | 6.3 | 9.7 | 30.7 | 2.9 | 31.9 | ---- | |
| City of Syracuse | 5.5 | 6.4 | 10.9 | 30.9 | 3.2 | 35.9 | ---- | |
| Southside (mean of 4 Census Tracts) | 5.2 | 8.9 | 19.0 | 44.1 | 5.2 | 49.1 | ---- | |
| Census Tract 42 | 4.3 | 9.1 | 20.1 | 44.0 | 5.7 | 52.3 | Not LI and/or LA | |
| Census Tract 53 | 6.4 | 11.0 | 20.3 | 46.7 | 5.8 | 47.9 | LI and LA at 0.5 miles | |
| Census Tract 54 | 5.2 | 7.7 | 17.7 | 42.6 | 4.6 | 47.3 | LI and LA at 0.5 miles | |
| Census Tract 58 | 5.0 | 7.6 | 17.9 | 43.2 | 4.6 | 48.9 | LI and LA at 0.5 miles | |
Table 2.
Food and Drug Administration Daily Values a and nutrient source thresholds b.
| Assigned Score | Vitamin A (DV = 900 mcg) | Vitamin C (DV = 90 mg) | Vitamin E (DV = 15 mg) | Selenium (DV = 55 mcg) | Riboflavin (DV = 1.3 mg) | |
|---|---|---|---|---|---|---|
| High source (≥20% DV) | 3 | ≥180 mcg | ≥18 mg | ≥3 mg | ≥11 mcg | ≥0.26 mg |
| Medium source (20–5% DV) | 2 | 45–180 mcg | 4.5–18 mg | 0.75– 3 mg | 2.75–11mcg | 0.065–0.26 mg |
| Low source (≤5% DV) | 1 | ≤45 mcg | ≤4.5 mg | ≤0.75 mg | ≤2.75 mcg | ≤0.065 mg |
| No value | 0 | 0 | 0 | 0 | 0 | 0 |
Table 3.
Known antioxidant concentrations (high, medium, low) in edible, forageable plants a.
| Scientific Name | Common Name | Edible Part | Vitamin A (RAE mcg) | Vitamin C (mg) | Vitamin E (mg) | Se (mcg) | Riboflavin (mg) |
|---|---|---|---|---|---|---|---|
| Fruits | |||||||
| Amelanchier alnifolia | Saskatoon serviceberry | raw fruit | 10.91 | 3.55 | 1.12 | 3.54 | |
| Asimina triloba | pawpaw | raw fruit | 8.6 | 18.3 | 0.09 | ||
| Celtis occidentalis | hackberry | raw fruit | 18.3 | ||||
| Chaenomeles speciosa | flowering quince | raw fruit | 72.15 | ||||
| Cornus kousa | kousa dogwood | raw fruit | 93 | ||||
| Diospyros virginiana | persimmon b | raw fruit | 66 | ||||
| Malus spp. | apple | raw fruit | 3 | 4.6 | 0.18 | 0 | 0.026 |
| Malus spp. | crabapple | raw fruit | 2 | 8 | 0.02 | ||
| Morus alba | white mulberry | raw fruit | 15.2 | 0.088 | |||
| Morus nigra | black mulberry | raw fruit | 1 | 36.4 | 0.87 | 0.6 | 0.101 |
| Prunus americana | American plum | raw fruit | 173 | 10.3 | 0.53 | 0.042 | |
| Prunus avium | sweet cherry | raw fruit | 3 | 7 | 0.07 | 0 | 0.033 |
| Pyrus spp. | pear | raw fruit | 1 | 4.3 | 0.12 | 0.1 | 0.026 |
| Ribes nigrum | blackcurrant | raw fruit | 12 | 181 | 1 | 0.05 | |
| Ribes rubrum | redcurrant | raw fruit | 2 | 41 | 0.1 | 0.6 | 0.05 |
| Ribes uva-crispa | gooseberry | raw fruit | 15 | 27.7 | 0.37 | 0.6 | 0.03 |
| Rubus fruticosus | blackberry | raw fruit | 11 | 21 | 1.17 | 0.4 | 0.026 |
| Rubus idaeus | red raspberry | raw fruit | 2 | 26.4 | 0.57 | 0.08 | |
| Sambucus canadensis | black elderberry | raw fruit | 30 | 36 | 0.06 | 0.06 | |
| Vitis spp. | grape | raw fruit | 5 | 4 | 0.19 | 0.1 | 0.057 |
| Herbs | |||||||
| Achillea millefolium | yarrow c | leaves | 0.95 | ||||
| Mentha spicata | spearmint | fresh leaves | 203 | 13.3 | 0.175 | ||
| Salvia officinalis | sage | ground herb | 295 | 32.4 | 7.48 | 3.7 | 0.336 |
| Nuts | |||||||
| Carya illinoinensis | pecan | raw nut | 3 | 1.1 | 1.4 | 3.8 | 0.13 |
| Corylus americana | hazelnut | raw nut | 1 | 6.3 | 15 | 2.4 | 0.113 |
| Juglans cinerea | butternut | dried nut | 6 | 3.2 | 17.2 | 0.148 | |
| Juglans nigra | black walnut | dried nut | 2 | 1.7 | 2.08 | 17 | 0.13 |
| Juglans regia | English walnut | dried nut | 1 | 1.3 | 0.7 | 4.9 | 0.15 |
| Vegetables | |||||||
| Alliaria petiolata | garlic mustard | fresh leaves | 26.1 | ||||
| Allium tricoccum | ramps | whole plant | 80 | ||||
| Chenopodium album | lambsquarters | raw vegetable | 580 | 80 | 0.9 | 0.44 | |
| Matteuccia struthiopteris | ostrich fern | raw fronds | 181 | 26.6 | 0.21 | ||
| Nasturtium officinale | watercress | fresh leaves | 160 | 43 | 1 | 0.9 | 0.12 |
| Portulaca oleracea | purslane | raw vegetable | 21 | 0.9 | 0.112 | ||
| Taraxacum spp. | dandelion | raw greens | 508 | 35 | 3.44 | 0.5 | 0.26 |
| Typha latifolia | broadleaf cattail | young shoots | 1 | 0.7 | 0.6 | 0.025 | |
| Vitis spp. | grape | leaves | 1380 | 11.1 | 2 | 0.9 | 0.354 |
| Other | |||||||
| Acer saccharum | sugar maple | sap | 0 | 0 | 0 | 0.6 | 1.27 |
Note. All values are reported per 100 g of “edible part.” Color coding corresponds to the scores described in Table 2, where 3 is a “high source” (green), 2 is a “medium source” (peach), 1, “low source “and 0, “no value” are grouped together (blue). a USDA FoodData Central (FDC) uses unique ID numbers to identify food items [81]; b Persimmon is an example of a food for which the USDA provides values for macronutrients but limited micronutrients. c Yarrow (Achillea millefolium) samples were collected in Spain and Portugal.
Table 4.
Composite Antioxidant Scores of Forageable Plants.
| Scientific Name | Common Name | Edible Part | Vit A Score | Vit C Score | Vit E Score | Se Score | Riboflavin Score | Composite Score |
|---|---|---|---|---|---|---|---|---|
| Fruits | ||||||||
| Morus nigra | black mulberry | raw fruit | 1 | 3 | 2 | 1 | 2 | 9 |
| Rubus fruticosus | blackberry | raw fruit | 1 | 3 | 2 | 1 | 1 | 8 |
| Ribes uva | gooseberry | raw fruit | 1 | 3 | 1 | 1 | 1 | 7 |
| Ribes rubrum | redcurrant | raw fruit | 1 | 3 | 1 | 1 | 1 | 7 |
| Prunus avium | sweet cherry | raw fruit | 1 | 2 | 1 | 0 | 1 | 5 |
| Pyrus spp. | pear | raw fruit | 1 | 1 | 1 | 1 | 1 | 5 |
| Vitis spp. | grape | raw fruit | 1 | 1 | 1 | 1 | 1 | 5 |
| Malus spp. | apple | raw fruit | 1 | 1 | 1 | 0 | 1 | 4 |
| Herbs | ||||||||
| Salvia officinalis | sage | ground herb | 3 | 3 | 3 | 2 | 3 | 14 |
| Nuts | ||||||||
| Juglans nigra | black walnut | dried nut | 1 | 1 | 2 | 3 | 2 | 9 |
| Corylus americana | hazelnut | raw nut | 1 | 2 | 3 | 1 | 2 | 9 |
| Carya illinoinensis | pecan | raw nut | 1 | 1 | 2 | 2 | 2 | 8 |
| Juglans regia | English walnut | dried nut | 1 | 1 | 1 | 2 | 2 | 7 |
| Vegetables | ||||||||
| Taraxacum spp. | dandelion | raw greens | 3 | 3 | 3 | 1 | 3 | 13 |
| Vitis spp. | grape | leaves | 3 | 2 | 2 | 1 | 3 | 11 |
| Nasturtium officinale | watercress | fresh leaves | 2 | 3 | 2 | 1 | 2 | 10 |
| Other | ||||||||
| Acer saccharum | sugar maple | sap | 0 | 0 | 0 | 1 | 3 | 4 |
Note. Scores and color-coding correspond to the scores described in Table 2, where 3 is a “high source” (green), 2 is a “medium source” (peach), 1 is a “low source” and 0 is “no value” (blue).
Table 5.
Composite Antioxidant Scores for Forageable and Popular Commercial Plants.
| Scientific Name | Common Name | Edible Part | Vit A Score | Vit C Score | Vit E Score | Se Score | Riboflavin Score | Composite Score |
|---|---|---|---|---|---|---|---|---|
| Fruits | ||||||||
| Morus nigra | black mulberry | raw fruit | 1 | 3 | 2 | 1 | 2 | 9 |
| Fragaria spp. | strawberry a | raw fruit | 1 | 3 | 1 | 1 | 1 | 7 |
| (1 mcg/100 g) | (58.8 mg/100 g) | (0.29 mg/100 g) | (0.4 mcg/100 g) | (0.022 mg/100 g) | ||||
| Herbs | ||||||||
| Salvia officinalis | sage | ground herb | 3 | 3 | 3 | 2 | 3 | 14 |
| Ocimum basilicum | basil b | fresh herb | 3 | 3 | 2 | 1 | 2 | 11 |
| (264 mcg/100 g) | (18 mg/100 g) | (0.8 mg/100 g) | (0.3 mcg/100 g) | (0.076 mg/100 g) | ||||
| Nuts | ||||||||
| Juglans nigra | black walnut | dried nut | 1 | 1 | 2 | 3 | 2 | 9 |
| Prunus dulcis | almond c | raw nut | 0 | 0 | 3 | 2 | 3 | 8 |
| (0 mcg/100 g) | (0 mg/100 g) | (25.6 mg/100 g) | (4.1 mcg/100 g) | (1.14 mg/100 g) | ||||
| Vegetables | ||||||||
| Taraxacum spp. | dandelion | raw greens | 3 | 3 | 3 | 1 | 3 | 13 |
| Lactuca sativa | romaine lettuce d | raw leaves | 3 | 1 | 1 | 1 | 2 | 8 |
| (436 mcg/100 g) | (4 mg/100 g) | (0.13 mg/100 g) | (0.4 mcg/100 g) | (0.067 mg/100 g) | ||||
a FDC 167762. b FDC 172232. c FDC 170567. d FDC 169247.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Bellows, A.C.; Raj, S.; Pitstick, E.; Potteiger, M.R.; Diemont, S.A.W. Foraging Wild Edibles: Dietary Diversity in Expanded Food Systems. Nutrients 2023, 15, 4630. https://doi.org/10.3390/nu15214630
AMA Style
Bellows AC, Raj S, Pitstick E, Potteiger MR, Diemont SAW. Foraging Wild Edibles: Dietary Diversity in Expanded Food Systems. Nutrients. 2023; 15(21):4630. https://doi.org/10.3390/nu15214630
Chicago/Turabian StyleBellows, Anne C., Sudha Raj, Ellen Pitstick, Matthew R. Potteiger, and Stewart A. W. Diemont. 2023. "Foraging Wild Edibles: Dietary Diversity in Expanded Food Systems" Nutrients 15, no. 21: 4630. https://doi.org/10.3390/nu15214630
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.