
Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS
Department of Immunology and Cell Biology, Faculty of Medicine and Health Sciences, University of Sherbrooke, Sherbrooke, QC J1H 5N4, Canada
*
Author to whom correspondence should be addressed.
Nutrients 2024, 16(5), 745; https://doi.org/10.3390/nu16050745
Submission received: 22 January 2024
/
Revised: 29 February 2024
/
Accepted: 1 March 2024
/
Published: 5 March 2024
(This article belongs to the Special Issue Nutrition, Lipoproteins and Cardiovascular Diseases)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Endocardial endothelium (EE) is a layer of cells covering the cardiac cavities and modulates cardiomyocyte function. This cell type releases several cardioactive factors, including Angiotensin II (Ang II). This octopeptide is known to induce cardiac hypertrophy. However, whether this circulating factor also induces EE hypertrophy is not known. Taurine is known to prevent cardiac hypertrophy. Whether this endogenous antioxidant prevents the effect of Ang II on human EE (hEE) will be verified. Using quantitative fluorescent probe imaging for calcium and reactive oxygen species (ROS), our results show that Ang II induces (10−7 M, 48 h treatment) an increase in hEE cell (hEEC) volume and its nucleus. Pretreatment with 20 mM of taurine prevents morphological remodeling and increases intracellular calcium and ROS. These results suggest that the reported Ang II induces cardiac hypertrophy is associated with hEEC hypertrophy. This later effect is prevented by taurine by reducing intracellular calcium and ROS overloads. Thus, taurine could be an excellent tool for preventing Ang II-induced remodeling of hEECs.
1. Introduction
The heart cavities are covered with a monolayer of endothelial cells called the endocardial endothelium (EE) [1,2,3]. These cells are highly active. One important property of EE is its anti-coagulant and anti-thrombotic activities. This aspect is important for cardiomyocyte intracellular calcium homeostasis. The latter level is essential for the synthesis and secretion of vasodilatory factors such as nitric oxide (NO) and prostacyclin [4,5]. On the other hand, the EE secretes several vasoconstrictor cardioactive substances, including endothelin-1 (ET-1), prostaglandins, and several components of the renin–angiotensin system (RAS), such as Angiotensin II (Ang II) [3,5,6]. They also possess receptors at the plasma and nuclear membranes for ET-1, NPY, and Ang II [3,5,6]. Ang II acts via its receptors at the surface and nuclear membranes of EE cells (EECs), inducing an elevation of intracellular calcium concentrations ([Ca2+]i). This elevation in [Ca2+]i may, in turn, induce remodeling of EE [6], hence its importance at the cardiac level. Moreover, a balance between the various factors secreted by EE is important for maintaining the intracellular ionic homeostasis and integrity of the endocardial endothelium.
Both EECs and vascular endothelial cells (VECs) regulate cardiac function [1]. However, although EECs and VECs have some similarities, there are significant differences [7], more particularly at the level of secretion of different circulating cardiovascular active factors [8]. The contribution of VECs to cardiac performance is due, at least in part, to the control of coronary blood flow supply to the myocardium [8]. However, the contribution of EECs to cardiac function is due to direct action on cardiomyocytes via the release and tuning of factors that affect not only cardiac function [1]. Thus, their close proximity to the adjacent cardiomyocytes affects the whole myocardium function.
Several studies have reported the presence of components of the renin–angiotensin II system (RAS) in endothelial cells, suggesting a prominent role for these cells in Ang II synthesis and secretion [9,10]. The presence of renin and its mRNA in ECs was reported, and their capacity to synthesize and secrete Ang II [11] was demonstrated. Angiotensin II converting enzyme (ACE) has also been identified in the endothelium’s luminal region at the cells’ plasma membrane [12]. In addition, ACE activity has been demonstrated in several endothelial cell preparations [11].
EECs release cardioactive factors such as Ang II in the extracellular matrix between these cells and ventricular cardiomyocytes [3]. Thus, they contribute to regulating cardiomyocyte function [3,7]. Evidence for the involvement of Ang II in cardiac hypertrophy is abundant [13,14,15]. This octapeptide, generated as part of the renin–angiotensin system, has been implicated in the pathophysiology of many cardiovascular diseases, such as peripheral artery disease, heart failure, hypertension, and coronary artery disease [16]. This circulating factor has been shown to induce hypertrophy of several cell types; however, nothing is known about the effect of Ang II on EEC hypertrophy, particularly those of human origin. It is reported that Ang II-induced cardiomyocyte hypertrophy is mediated via activation of intracellular signaling, particularly increasing ROS levels or decreasing anti-ROS production [17]. The increase in intracellular ROS by Ang II was reported to be mediated via the activation of c-Src causing an increase in NOX activity [17] and, more particularly, the Ca2+-dependent NOX5. In addition, taurine, a nonessential amino acid, is a well-known endogenous antioxidant and was recently reported to prevent the development of hypertrophy associated with heart failure and early death [18,19]. However, verifying whether taurine prevents Ang II’s effect on EECs via reducing Ang II-induced cell hypertrophy associated with increased intracellular ROS awaits verification.
Taurine is a well-known nonessential amino acid [19,20] that is very important for cell function. This circulating factor is produced mainly by the brain and the heart. Nutrients, particularly seafood [21,22,23,24,25,26], ensure its supplementation. Taurine’s beneficial effects are numerous, particularly at the cardiovascular level. Taurine was reported to be a cardiac anti-hypertrophic factor in vivo as well as in vitro [19,27]. Its overall beneficial effects were attributed in part to its anti-ROS properties [19,28] and in preventing intracellular calcium overload [3,19,27].
We took into consideration the following: 1—Ang II induces cardiac hypertrophy associated with an increase in cardiomyocyte intracellular Ca2+ and ROS [15]; 2—the first abnormal remodeling takes place at the EE during the development of hereditary cardiomyopathy, and it contributes to cardiovascular dysfunction [21]; 3—endothelial cell dysfunction is a complex mechanism [29] implicating ROS generation and overload that is also activated by Ang II [22,23,24,25]. Thus, in the present study, we wanted to verify whether Ang II induces hypertrophy of human EECs and whether this effect can be prevented by treatment with taurine.
2. Materials and Methods
2.1. Isolation and Culturing of Human Endocardial Endothelial Cells
The procedure for isolating and culturing hEECs was described previously. The procedures were performed in compliance with the institutional review committee’s requirements for using the human tissues of donors. In brief, hEECs were separated from the right ventricle of human fetal donor hearts. The right ventricle is open and cleaned from blood. For isolating the hEECs, the cardiac cavities were exposed to trypsin and then washed with the physiological ionic culture medium, the M199 solution containing 5% fetal bovine serum (v/v) (Life Technologies, Burlington, ON, Canada). The hEECs were gently isolated using a scalpel. Then, the cells were centrifuged at 200× g and added in the culture medium. As reported previously, isolated hEECs were cultured in Petri dishes. At confluence, hEECs were again isolated and recultured on glass coverslips placed in the culture dishes. The purity of cultured hEECs is verified using markers of these cell types.
2.2. Confocal Microscopy
As previously described, hEECs are studied using quantitative 3D confocal microscopy of a Bio-Rad system [26]. In summary, the laser line (9.0 mV) is directed to hEEC and is filtered to prevent photobleaching of the fluorescent dye. The confocal settings were kept unchanged in all the image recordings. The size between the cell sections (16–20 sections/cell depending on the value of the z line) is kept near zero to construct the real image of the cell in 3D. As reported previously, the nucleus is labeled with the fluorescent probe of nucleic acids, syto-11 (Molecular Probes, Eugene, OR, USA) [26]. Real 3D images are analyzed using an ImageSpace Rix version 6.5 analyzing system. This program allows quantitative 3D images to be obtained by measuring the volume of the cell (expressed in μm3).
2.3. Determination of the Cell Volume
Recorded images are transferred to an analysis station of Silicon Graphics equipped with Imagespace 3D analysis and reconstruction software from Molecular Dynamics. Images of cell volume are obtained and calcium measurements are performed on real three-dimensional reconstructions. The nucleus region, marked with Syto 11, is isolated from the rest of the cell by lowering the intensity threshold to delineate the pixels in this space. This method enables us to create a true three-dimensional reconstruction of the nucleus alone or of the cell without the nucleus. This method allows us to measure the fluorescence intensity values of the volume of the cell and the Fluo-4/Ca2+ complex of the nuclear region and the cytosolic region separately, eliminating any contribution from the other compartment.
2.4. Loading with the Calcium and ROS Fluorescent Probes
The cell membrane permeable Ca2+ fluorescence dye Fluo-4/AM (Molecular Probes, Eugene, OR, USA) is used to load Fluo-4 into the cytoplasm and the nucleoplasm of hEECs, as reported in our previous published work [26]. The calcium fluorescent probe Fluo-4 is homogeneously distributed in the hEECs [26] and can be expressed in free calcium concentration using a calibration method described previously. For ROS imaging studies, cells are loaded, respectively with the ROS/probe, 6-carboxy-2′,7′-dichlorodihydrofluorescein diacetate (carboxy-H2DCF-DA; Life Technologies, Burlington, ON, Canada). The methods have been developed and described previously.
2.5. Statistical Analyses
In this work, intracellular calcium and ROS levels are expressed as means ± SEM. n represents the number of hEECs from a minimum of three different experiments. N is the number of human donors. Student’s t-test and one-way repeated measures ANOVA with post hoc t-test (p < 0.05) are used along with the Tukey–Kramer or the Newman–Keuls and analysis by the program Graph Pad Prism version 8.4.3 (686).
3. Results
Effect of Ang II on Human Ventricular Endocardial Endothelial Cells
To verify if Ang II induces hypertrophy of hEECs, the cells were exposed for 48 h to 10−7 M of Ang II. Our group reported that this concentration increased intracellular free calcium in several types of cells. Then, the volume of the cells was assessed using the fluorescent probe syto-11 coupled to quantitative 3D confocal microscopy [3]. Figure 1 illustrates typical images of the effect of Ang II treatment on hEECs. Figure 1A,B show an apparent increase in the cell volume, including the nucleus following chronic treatment with Ang II (10−7 M, for 48 h) (Figure 1B) compared to control (Figure 1A). This apparent increase in hEECs volume by Ang II is highly significant at the whole cell (Figure 2A; p < 0.01), the cytosol (Figure 2B; p < 0.01) and the nucleus (Figure 2C, p < 0.01) levels. On the other hand, 48 h treatment with Ang II in the presence of 20 mM of taurine [19] prevents Ang II-induced increase in EEC apparent volume (Figure 1C). This effect of taurine was highly significant at the whole cell (p < 0.0001), the cytosol (p < 0.001), and the nucleus (p < 0.0001) levels.
In a second set of experiments, using the protocols described earlier, coupled to the calcium fluorescent probe Fluo-4, we examined if an increase in cytosolic and nuclear calcium accompanies the hypertrophy induced by a 48 h Ang II treatment. Figure 3 shows typical results. Figure 3A shows, as expected, a higher calcium level in the nucleoplasm than in the cytosol. Figure 3B shows that the hypertrophy induced by a 48 treatment with Ang II is associated with a very high increase in free calcium that is significant at the level of the whole cell (Figure 4A; p < 0.0001), the cytoplasm (Figure 4B; p < 0.01), and the nucleoplasm (Figure 4C; p < 0.001).
In the following experiments, we verified whether the prevention of Ang II-induced hypertrophy associated with increased intracellular calcium is prevented by treatment with taurine (20 mM). As seen in Figure 3C and Figure 4, treatment with taurine prevents Ang II-induced hypertrophy and the increase in the whole cell (Figure 4A; p < 0.01), cytosolic (Figure 4B; p < 0.05), and nuclear (Figure 4C; p < 0.05) calcium levels.
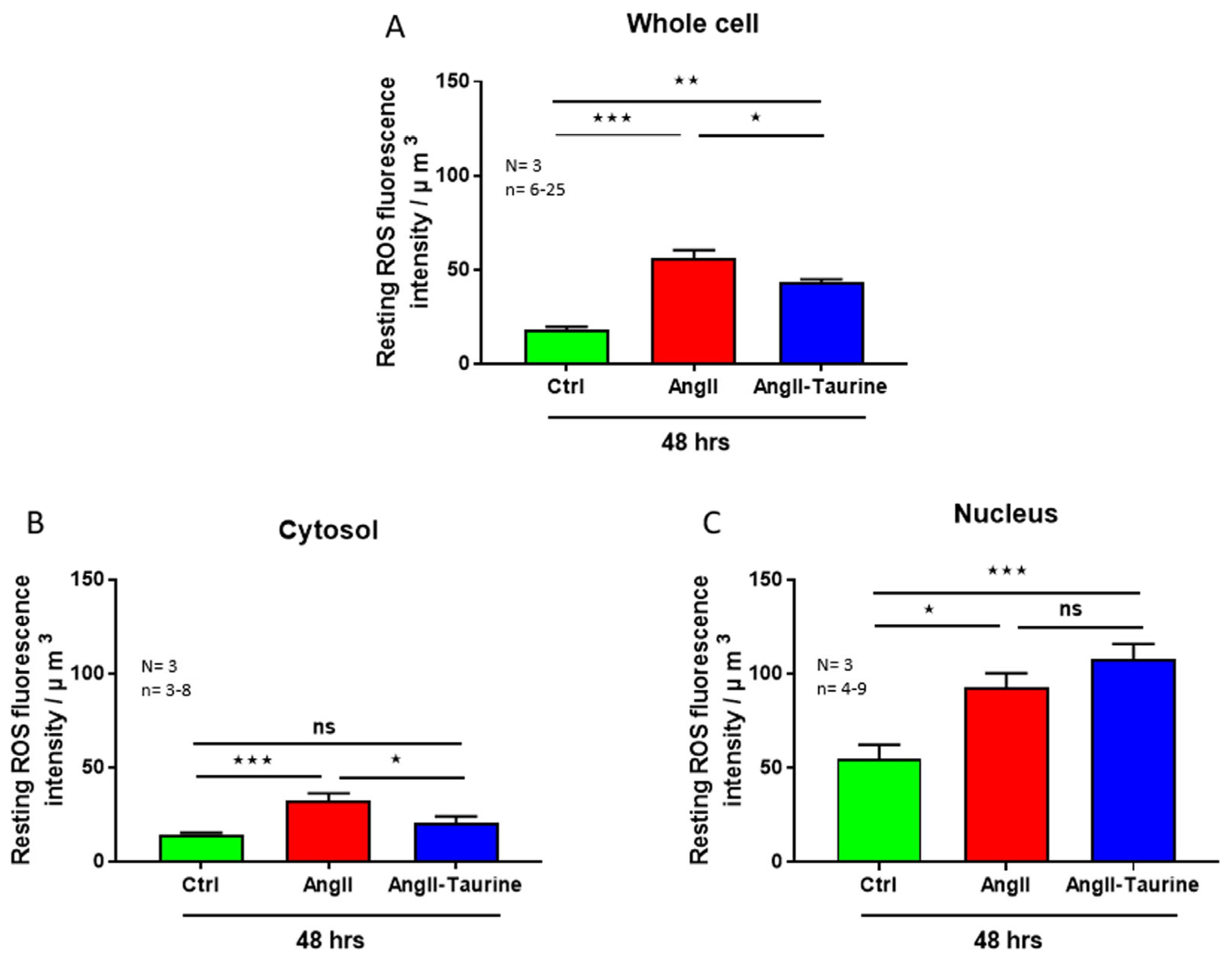
Using the same protocol described previously, we verified whether the Ang II-induced hEEC hypertrophy and intracellular calcium overload are associated with increased cytosolic and nuclear ROS overload. Figure 5 shows typical examples. As seen in this figure, intracellular ROS is distributed non-homogenously (Figure 5A). Figure 5B shows that Ang II-induced hEEC hypertrophy is accompanied by an apparent increase in ROS throughout the cells. As seen in Figure 5B,C, in the presence of Ang II, taurine apparently decreased ROS levels throughout the cell except for the nucleus. This ROS increase is statistically significant in the whole cell (Figure 6A; p < 0.001), the cytoplasm (Figure 6B; p < 0.001), and the nucleoplasm (Figure 6C; p < 0.05). In the presence of Ang II, treatment with 20 mM of taurine partially prevents the Ang II-induced increase in ROS at the whole cell level (Figure 6A; p < 0.05). However, it completely prevented the increase in cytoplasmic ROS level compared to the control ROS level (Figure 6B; ns) and did not affect nuclear ROS level (Figure 6C; ns).
4. Discussion
The dialogue between EECs and the cardiomyocytes occurs during the fetal formation of the heart. This continues during fetal cardiac development and cardiomyocytes differentiation [8]. Signaling, including secretion of EECs contributes to the development and maturation of ventricular trabecula that characterize the heart ventricles. Several reports have demonstrated that the contribution of EECs to heart function continues during an individual’s life. This is due to the fact that EECs are essential for the survival of postnatal cardiomyocytes and for the integrity of the heart [30]. In addition, EECs influence myocardial performance [31]. Brutsaert et al. were the first to report that in the absence of EECs, the contraction of cat cardiac papillary muscles resulted in a shortening of twitch contractions and a decrease in peak twitch force [8,32]. Similar work was reported by Fort et al.’s group using the whole heart [33]. The contractile performance of the heart depends primarily on the level of increase in transient intracellular Ca2+ [8].
Furthermore, EECs modulate the affinity of contractile proteins to an intracellular elevation of Ca2+ [8] in chronic high-circulating Ang II. Two mechanisms have been proposed to explain the modulatory effect of Ang II on EE and its regulation of myocardial performance: 1—upon stimulation with Ang II, EECs secrete several factors that alter the contractile state of the subjacent myocytes (excitation–secretion–contraction coupling) [8]; 2—the EE may act as a physicochemical barrier which controls the ionic constitution of the interstitial milieu surrounding the cardiomyocytes, and consequently, it modulates their performance (EE as a blood–heart barrier) [2,8,32]. Thus, modulation of EE function by Ang II will affect its excitation–secretion coupling and, consequently, will modulate the adjacent cardiomyocytes’ excitation–contraction coupling and EE morphological remodeling, leading to cardiac hypertrophy. Our results, using quantitative 3D confocal microscopy, demonstrate that long-term treatment with Ang II (48 h) increased the volume of hEECs. This increase is associated with an increase in the volume of the nucleus. The latter directly indicates that the increase in volume is indeed hypertrophy [34]. These findings demonstrate that Ang II-induced cardiac hypertrophy [13,14] is associated with hEEC hypertrophy. The latter will promote the release of hEECs growth factors such as ET-1 and Ang II, which contribute to the development of the well-known cardiac hypertrophy [13,14]. Ang II-induced hypertrophy could be partly due to the activation of AT1 and AT2 receptors. The latter promotes the Ang II-induced morphological remodeling of all cardiac cell types, including EECs. Our results also show that the Ang II-induced hypertrophy of hEECs is associated with increased intracellular calcium. Such an effect is similar to those effects reported in cardiomyocytes, vascular endothelial, and smooth muscle cells [3,35].
The increase in hEECs nuclear calcium by Ang II could be due to Ang II activation of the following: 1—calcium influx through the nuclear membranes ionic transporters; 2—release of calcium from endoplasmic IP3 sensitive pools [3]; and 3—release of calcium from nucleoplasmic reticulum IP3 [13]. Since endothelial cells generally possess mainly R-type calcium channels [15,23], the increase in cytosolic and nuclear calcium could also be due to the activation of plasma and nuclear membranes’ R-type calcium channels, as reported previously.
Our results also show that, as for other types of cells [3], Ang II-induced hEEC hypertrophy is also associated with increased cytosolic and nuclear ROS levels. This ROS increase occurs in several types of hypertrophies, including that induced by Ang II [13,14]. The increase in cytosolic and nuclear ROS could be due, at least in part, to activation of the calcium-depended NOX5.
Our results show that, as in hypertrophy associated with cardiomyopathy [19], taurine prevents Ang II-induced hEEC hypertrophy. This suggests that the reported preventive effect of taurine in hypertrophic cardiomyopathy [19] could be due at least in part to beneficial effects in the prevention of EEC hypertrophy. This should be verified in the future. In addition, the prevention of Ang II-induced hypertrophy and increase in intracellular free calcium could be due to the long-term effect of taurine in reducing calcium overload [18,19]. Our results also show that taurine prevents Ang II-induced increase in cytosolic ROS without affecting the increase in ROS at the nucleoplasmic level. These results suggest that ROS generation at the cytosolic level is differently generated in the cytosol when compared to the nucleoplasm. This could be due to differences in localization and density of NOXs (1–5) at the nuclear level compared to the cytosol. Since taurine prevents Ang II from inducing an increase in nuclear volume and calcium without affecting nuclear ROS levels in hEECs, this suggests that the latter does not contribute to the hypertrophy induced by Ang II.
Taurine supplementation also increases the proteomic and mRNA levels of protein kinase A-cAMP response element-binding protein (PKA-CREB) [36,37]. This transcription factor prevents cardiovascular cells’ morphological remodeling and explains, at least in part, taurine’s anti-trophic action on Ang II-induced EE hypertrophy. It is known that an increase in Ang II modulates the activity of the Na/Ca exchanger. This promotes calcium entry via this exchanger, accumulating calcium at both the cytosolic and nuclear levels [19,27,38]. It is possible that taurine indirectly blocks the activity of this exchanger via its inhibition of calcium-dependent kinases such as protein kinase C (PKC) [39,40]. It is unlikely that taurine blocks the activity of Ang II-activated receptors. On the other hand, as indicated above, it is likely that, at least in part, taurine’s anti-hypertrophic effect in EE is due to its inhibition of Ang II signalization cascades, such as those responsible for activation of the PKA-CREB and intracellular ROS generation pathways [17]. Thus, the prevention of Ang II-induced hypertrophy in hEECs by taurine could be mainly due to taurine blunting the increase in oxidative stress induced by Ang II [17,19].
We have to mention that the nucleus also acts as a cell within a cell. The nuclear membranes possess ACE and thus can generate Ang II, which will activate its receptors AT1 and AT2 at the nuclear membranes’ levels. The activation of these nuclear membrane receptors by Ang II-induced calcium influx through the nuclear envelope membranes is independent of the increase in cytosolic Ca2+. The internalization of Ang II promotes the activation of the nuclear membranes’ AT1 and AT2 receptors. The activation of these receptors stimulates Ca2+ influx through the nuclear membranes’ R-type Ca2+ channels. The latter may explain, at least in part, the increase in nuclear Ca2+ by Ang II in EECs. Since taurine influx takes place via its plasma membrane symporters, which increases its intracellular level, the latter may directly act at the nuclear level, thus preventing remodeling of the nucleus by Ang II.
5. Conclusions
In conclusion, our results clearly show that Ang II does induce hypertrophy of EECs, and these types of cells may contribute to the overall effect of Ang II in the heart. In addition, taurine’s prevention of Ang II-induced EEC hypertrophy demonstrates that the preventive effect of this nonessential amino acid on cardiac hypertrophy in hereditary cardiomyopathy [18,19] is due, at least in part, to its prevention of EEC hypertrophy.
Author Contributions
D.J. and G.B. contributed equally to this work. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Sciences and Engineering Research Council of Canada (NSERC, RGPIN-2017-05508 and RGPIN-2016-04414).
Institutional Review Board Statement
This work was carried out according the Comité d'éthique de la recherche du CIUSSS de l’Estrie-CHUS (protocol number 2005-31, 04-031, 10 March 2023).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflicts of interest.
References
- Kuruvilla, L.; Kartha, C.C. Molecular mechanisms in endothelial regulation of cardiac function. Mol. Cell. Biochem. 2003, 253, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Brutsaert, D.L.; De Keulenaer, G.W.; Fransen, P.; Mohan, P.; Kaluza, G.L.; Andries, L.J.; Rouleau, J.L.; Sys, S.U. The cardiac endothelium: Functional morphology, development, and physiology. Prog. Cardiovasc. Dis. 1996, 39, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Jacques, D. Morphological and Functional Remodeling of Vascular Endothelium in Cardiovascular Diseases. Int. J. Mol. Sci. 2023, 24, 1998. [Google Scholar] [CrossRef] [PubMed]
- Lüscher, T.F.; Barton, M. Biology of the endothelium. Clin. Cardiol. 1997, 20 (Suppl. S2), II-3–II-10. [Google Scholar] [CrossRef]
- Mombouli, J.V.; Vanhoutte, P.M. Endothelial dysfunction: From physiology to therapy. J. Mol. Cell. Cardiol. 1999, 31, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Droogmans, G. Ion channels and their functional role in vascular endothelium. Physiol. Rev. 2001, 81, 1415–1459. [Google Scholar] [CrossRef]
- Shah, A.M.; Grocott-Mason, R.M.; Pepper, C.B.; Mebazaa, A.; Henderson, A.H.; Lewis, M.J.; Paulus, W.J. The cardiac endothelium: Cardioactive mediators. Prog. Cardiovasc. Dis. 1996, 39, 263–284. [Google Scholar] [CrossRef]
- Brutsaert, D.L. Cardiac endothelial-myocardial signaling: Its role in cardiac growth, contractile performance, and rhythmicity. Physiol. Rev. 2003, 83, 59–115. [Google Scholar] [CrossRef]
- Xiao, F.; Puddefoot, J.R.; Vinson, G.P. The expression of renin and the formation of angiotensin II in bovine aortic endothelial cells. J. Endocrinol. 2000, 164, 207–214. [Google Scholar] [CrossRef]
- Baudin, B.; Berard, M.; Carrier, J.L.; Legrand, Y.; Drouet, L. Vascular origin determines angiotensin I-converting enzyme expression in endothelial cells. Endothelium 1997, 5, 73–84. [Google Scholar] [CrossRef]
- Carley, W.W.; Tanoue, L.; Merker, M.; Gillis, C.N. Isolation of rabbit pulmonary microvascular endothelial cells and characterization of their angiotensin converting enzyme activity. Pulm. Pharmacol. 1990, 3, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.S.; Ryan, J.W.; Whitaker, C.; Chiu, A. Localization of angiotensin converting enzyme (kininase II). II. Immunocytochemistry and immunofluorescence. Tissue Cell 1976, 8, 125–145. [Google Scholar] [CrossRef]
- Yamazaki, T.; Yazaki, Y. Molecular basis of cardiac hypertrophy. Z. Kardiol. 2000, 89, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, S.K.; Dhalla, N.S. Adaptive and maladaptive roles of different angiotensin receptors in the development of cardiac hypertrophy and heart failure. Can. J. Physiol. Pharmacol. 2024, 102, 86–104. [Google Scholar] [CrossRef] [PubMed]
- Jacques, D.B.G. Cardiovascular physiopathology of angiotensin II and its plasma and nuclear envelop mem-branes’ receptors. In The Renin Angiotensin System in Cardiovascular Disease. Advances in Biochemistry in Health and Disease; Dhalla, N.S., Bhullar, S.K., Shah, A.K., Eds.; Springer: Cham, Switzerland, 2023; pp. 63–80. [Google Scholar]
- Vandemark, C.; Nguyen, J.; Zhao, Z.Q. Cardiovascular Protection with a Long-Acting GLP-1 Receptor Agonist Liraglutide: An Experimental Update. Molecules 2023, 28, 1369. [Google Scholar] [CrossRef]
- Hussain, M.; Ikram, W.; Ikram, U. Role of c-Src and reactive oxygen species in cardiovascular diseases. Mol. Genet. Genom. 2023, 298, 315–328. [Google Scholar] [CrossRef]
- Bkaily, G.; Simon, Y.; Normand, A.; Jazzar, A.; Najibeddine, H.; Khalil, A.; Jacques, D. Short-Communication: Short-Term Treatment with Taurine Prevents the Development of Cardiac Hypertrophy and Early Death in Hereditary Cardiomyopathy of the Hamster and Is Sex-Dependent. Nutrients 2022, 14, 3287. [Google Scholar] [CrossRef]
- Bkaily, G.; Jazzar, A.; Normand, A.; Simon, Y.; Al-Khoury, J.; Jacques, D. Taurine and cardiac disease: State of the art and perspectives. Can. J. Physiol. Pharmacol. 2020, 98, 67–73. [Google Scholar] [CrossRef]
- Ahmed, K.; Choi, H.N.; Park, J.S.; Kim, Y.G.; Bae, M.K.; Yim, J.E. Taurine supplementation alters gene expression profiles in white adipose tissue of obese C57BL/6J mice: Inflammation and lipid synthesis perspectives. Heliyon 2024, 10, e23288. [Google Scholar] [CrossRef]
- Padovan, J.C.; Dourado, T.M.H.; Pimenta, G.F.; Bruder-Nascimento, T.; Tirapelli, C.R. Reactive Oxygen Species Are Central Mediators of Vascular Dysfunction and Hypertension Induced by Ethanol Consumption. Antioxidants 2023, 12, 1813. [Google Scholar] [CrossRef]
- Braun, H.; Hauke, M.; Petermann, M.; Eckenstaler, R.; Ripperger, A.; Schwedhelm, E.; Ludwig-Kraus, B.; Bernhard Kraus, F.; Jalal Ahmed Shawon, M.; Dubourg, V.; et al. Deletion of vascular thromboxane A(2) receptors and its impact on angiotensin II-induced hypertension and atherosclerotic lesion formation in the aorta of Ldlr-deficient mice. Biochem. Pharmacol. 2024, 219, 115916. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.; Zhang, Y.; Xing, D.; Lei, Z.; Jiao, X. Angiotensin II type 1a receptor knockout ameliorates high-fat diet-induced cardiac dysfunction by regulating glucose and lipid metabolism. Acta Biochim. Biophys. Sin. 2023, 55, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Gwathmey, J.K.; Xie, L.H. Oxidative stress-mediated effects of angiotensin II in the cardiovascular system. World J. Hypertens. 2012, 2, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Domenighetti, A.A. Adaptive and maladaptive remodeling of cardiomyocyte excitation-contraction coupling by angiotensin II. Trends Cardiovasc. Med. 2010, 20, 78–85. [Google Scholar] [CrossRef]
- Bkaily, G.; Al-Khoury, J.; Simon, Y.; Jacques, D. Intracellular Free Calcium Measurement Using Confocal Imaging. Methods Mol. Biol. 2017, 1527, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Takahahsi, K.; Azuma, M.; Baba, A.; Schaffer, S.; Azuma, J. Taurine improves angiotensin II-induced hypertrophy of cultured neonatal rat heart cells. Adv. Exp. Med. Biol. 1998, 442, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Hsu, C.N. Protective Role of Taurine on Rat Offspring Hypertension in the Setting of Maternal Chronic Kidney Disease. Antioxidants 2023, 12, 2059. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.H.; Liu, J.C.; Lin, H.; Shih, N.L.; Chen, Y.L.; Huang, M.T.; Chan, P.; Cheng, C.F.; Chen, J.J. Inhibitory effect of resveratrol on angiotensin II-induced cardiomyocyte hypertrophy. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, K.; Segers, V.F.; Demolder, M.; De Keulenaer, G.W. Role of neuregulin-1/ErbB2 signaling in endothelium-cardiomyocyte cross-talk. J. Biol. Chem. 2006, 281, 19469–19477. [Google Scholar] [CrossRef]
- Abelanet, A.; Camoin, M.; Rubin, S.; Bougaran, P.; Delobel, V.; Pernot, M.; Forfar, I.; Guilbeau-Frugier, C.; Galès, C.; Bats, M.L.; et al. Increased Capillary Permeability in Heart Induces Diastolic Dysfunction Independently of Inflammation, Fibrosis, or Cardiomyocyte Dysfunction. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 745–763. [Google Scholar] [CrossRef]
- Brutsaert, D.L.; Fransen, P.; Andries, L.J.; De Keulenaer, G.W.; Sys, S.U. Cardiac endothelium and myocardial function. Cardiovasc. Res. 1998, 38, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Fort, S.; Lewis, M.J.; Shah, A.M. The role of endocardial endothelium in the modulation of myocardial contraction in the isolated whole heart. Cardioscience 1993, 4, 217–223. [Google Scholar] [PubMed]
- Smiljic, S. The clinical significance of endocardial endothelial dysfunction. Medicina 2017, 53, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Abou Abdallah, N.; Simon, Y.; Jazzar, A.; Jacques, D. Vascular smooth muscle remodeling in health and disease. Can. J. Physiol. Pharmacol. 2021, 99, 171–178. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Wang, X.F.; Chen, H.; Yang, N. Antenatal taurine supplementation improves cerebral neurogenesis in fetal rats with intrauterine growth restriction through the PKA-CREB signal pathway. Nutr. Neurosci. 2013, 16, 282–287. [Google Scholar] [CrossRef]
- Fang, Q.; Liu, J.; Chen, L.; Chen, Q.; Ke, J.; Zhang, J.; Liu, Y.; Fu, W. Taurine improves the differentiation of neural stem cells in fetal rats with intrauterine growth restriction via activation of the PKA-CREB-BDNF signaling pathway. Metab. Brain Dis. 2021, 36, 969–981. [Google Scholar] [CrossRef]
- Qaradakhi, T.; Gadanec, L.K.; McSweeney, K.R.; Abraham, J.R.; Apostolopoulos, V.; Zulli, A. The Anti-Inflammatory Effect of Taurine on Cardiovascular Disease. Nutrients 2020, 12, 2847. [Google Scholar] [CrossRef]
- Schaffer, S.; Solodushko, V.; Pastukh, V.; Ricci, C.; Azuma, J. Possible cause of taurine-deficient cardiomyopathy: Potentiation of angiotensin II action. J. Cardiovasc. Pharmacol. 2003, 41, 751–759. [Google Scholar] [CrossRef]
- Azuma, M.; Takahashi, K.; Fukuda, T.; Ohyabu, Y.; Yamamoto, I.; Kim, S.; Iwao, H.; Schaffer, S.W.; Azuma, J. Taurine attenuates hypertrophy induced by angiotensin II in cultured neonatal rat cardiac myocytes. Eur. J. Pharmacol. 2000, 403, 181–188. [Google Scholar] [CrossRef]
Figure 1.
An example of real 3D top view confocal microscopy images showing the apparent increase in hEECs volume and its prevention by taurine. Images (A–C) show syto-11 labeled cells and their nucleus: (A) in the absence of angiotensin II (Ang II), (B) in the presence 48 h of treatment with 10−7 M of Ang II, and (C) in the presence of Ang II and taurine (Tau, 20 mM). The calibration of the white bar is in μm. Images (A–C) represent different cells.
Figure 1.
An example of real 3D top view confocal microscopy images showing the apparent increase in hEECs volume and its prevention by taurine. Images (A–C) show syto-11 labeled cells and their nucleus: (A) in the absence of angiotensin II (Ang II), (B) in the presence 48 h of treatment with 10−7 M of Ang II, and (C) in the presence of Ang II and taurine (Tau, 20 mM). The calibration of the white bar is in μm. Images (A–C) represent different cells.

Figure 2.
In human endocardial endothelial cells, treatment for 48 h with 10−7 M Ang II-induced an increase in the whole cell volume ((A), red color), the cytosol ((B), red color), and the nucleus ((C), red color). Treatment with taurine for 48 h induced a partial but significant prevention of Ang II-induced increase in cell volume at the whole cell ((A), blue color), the cytosolic ((B), blue color), and the nucleoplasmic ((C), blue color) levels. The results are presented as mean ± SEM; n is the number of cells from five donors (N). ** p < 0.01; **** p < 0.0001. Results are presented in µm3.
Figure 2.
In human endocardial endothelial cells, treatment for 48 h with 10−7 M Ang II-induced an increase in the whole cell volume ((A), red color), the cytosol ((B), red color), and the nucleus ((C), red color). Treatment with taurine for 48 h induced a partial but significant prevention of Ang II-induced increase in cell volume at the whole cell ((A), blue color), the cytosolic ((B), blue color), and the nucleoplasmic ((C), blue color) levels. The results are presented as mean ± SEM; n is the number of cells from five donors (N). ** p < 0.01; **** p < 0.0001. Results are presented in µm3.
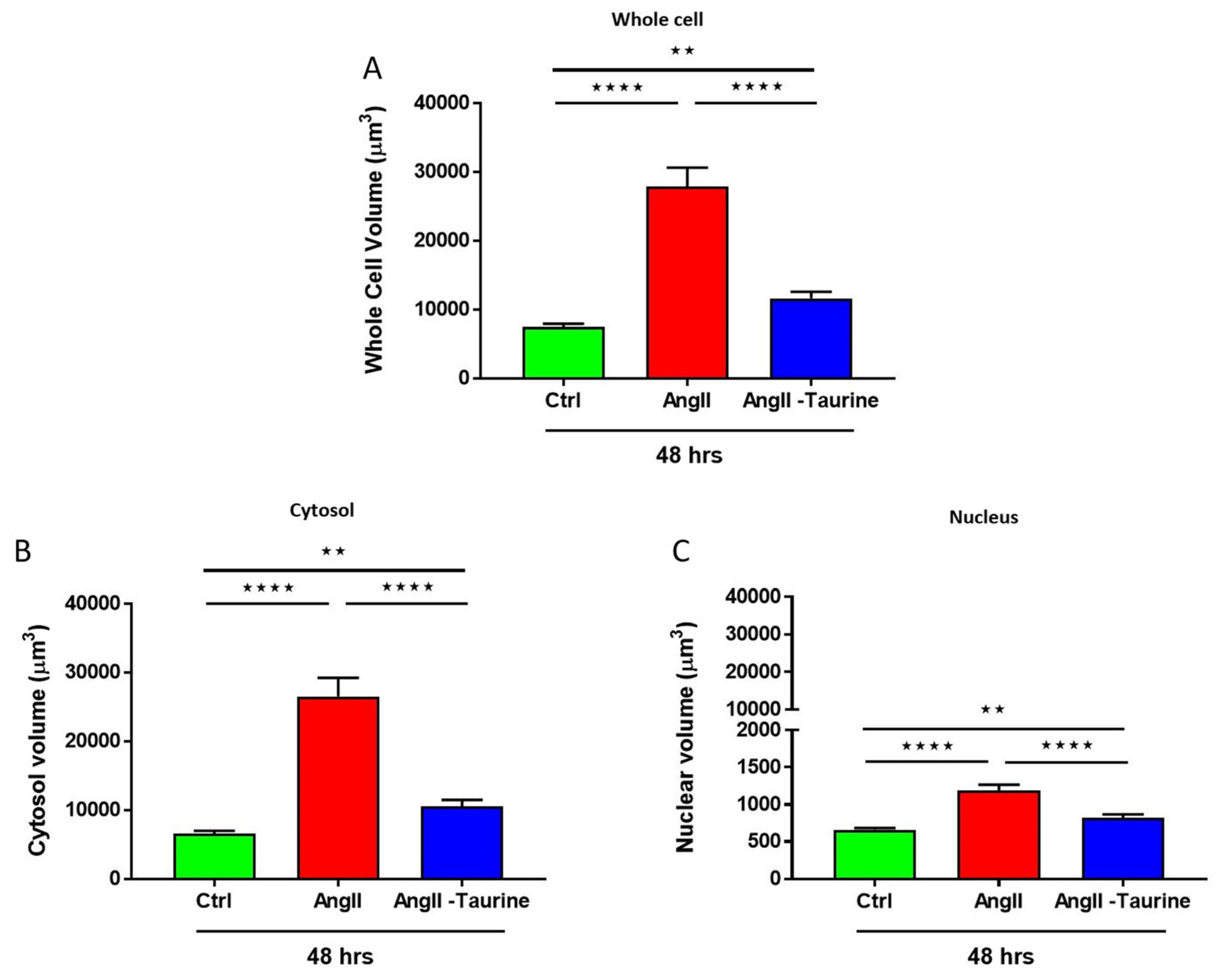
Figure 3.
An example of a quantitative 3D fishnet plot representation showing the apparent cytoplasmic and nucleoplasmic free Ca2+ levels in hEECs. Image (A) represents hVSMCs Ca2+ levels and distribution in a normal control medium. Image (B) represents hEECs Ca2+ levels and distribution in the presence of Ang II (10−7 M) for 48 h. Image (C) represents hEECs Ca2+ levels and distribution in the presence of 48 h of treatment with Ang II (10−7 M) and taurine (20 mM). Insert panels (A–C) in green color (color has no measurable meaning) represent syto-11 labeled nucleus in the absence of angiotensin II (Ang II) (A), in the presence 48 h treatment with Ang II (B), and in the presence of Ang II and taurine (C). The white bar scale is in μm. Images (A–C) represent different cells. The color calibration bar represents the free Ca2+ fluorescence level from 0 (black color) to 255 (white color).
Figure 3.
An example of a quantitative 3D fishnet plot representation showing the apparent cytoplasmic and nucleoplasmic free Ca2+ levels in hEECs. Image (A) represents hVSMCs Ca2+ levels and distribution in a normal control medium. Image (B) represents hEECs Ca2+ levels and distribution in the presence of Ang II (10−7 M) for 48 h. Image (C) represents hEECs Ca2+ levels and distribution in the presence of 48 h of treatment with Ang II (10−7 M) and taurine (20 mM). Insert panels (A–C) in green color (color has no measurable meaning) represent syto-11 labeled nucleus in the absence of angiotensin II (Ang II) (A), in the presence 48 h treatment with Ang II (B), and in the presence of Ang II and taurine (C). The white bar scale is in μm. Images (A–C) represent different cells. The color calibration bar represents the free Ca2+ fluorescence level from 0 (black color) to 255 (white color).

Figure 4.
In hECCs, treatment for 48 h with Ang II (10−7 M) elevated intracellular free Ca2+ in the whole cell ((A), red color), the cytoplasm ((B), red color), and the nucleoplasm ((C), red color). Treatment with taurine for 48 h induced partial but statistically significative prevention of Ang II-induced increase in Ca2+ free levels in the whole cell ((A), blue color), and completely prevented the Ang II-induced increase in Ca2+ free levels in the cytoplasm ((B), blue color) and the nucleoplasm ((C), blue color). The results are expressed as mean ± SEM, and n represents the number of cells from five donors (N). * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; ns: not significant. Results are presented per µm3 of the cell volume.
Figure 4.
In hECCs, treatment for 48 h with Ang II (10−7 M) elevated intracellular free Ca2+ in the whole cell ((A), red color), the cytoplasm ((B), red color), and the nucleoplasm ((C), red color). Treatment with taurine for 48 h induced partial but statistically significative prevention of Ang II-induced increase in Ca2+ free levels in the whole cell ((A), blue color), and completely prevented the Ang II-induced increase in Ca2+ free levels in the cytoplasm ((B), blue color) and the nucleoplasm ((C), blue color). The results are expressed as mean ± SEM, and n represents the number of cells from five donors (N). * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001; ns: not significant. Results are presented per µm3 of the cell volume.
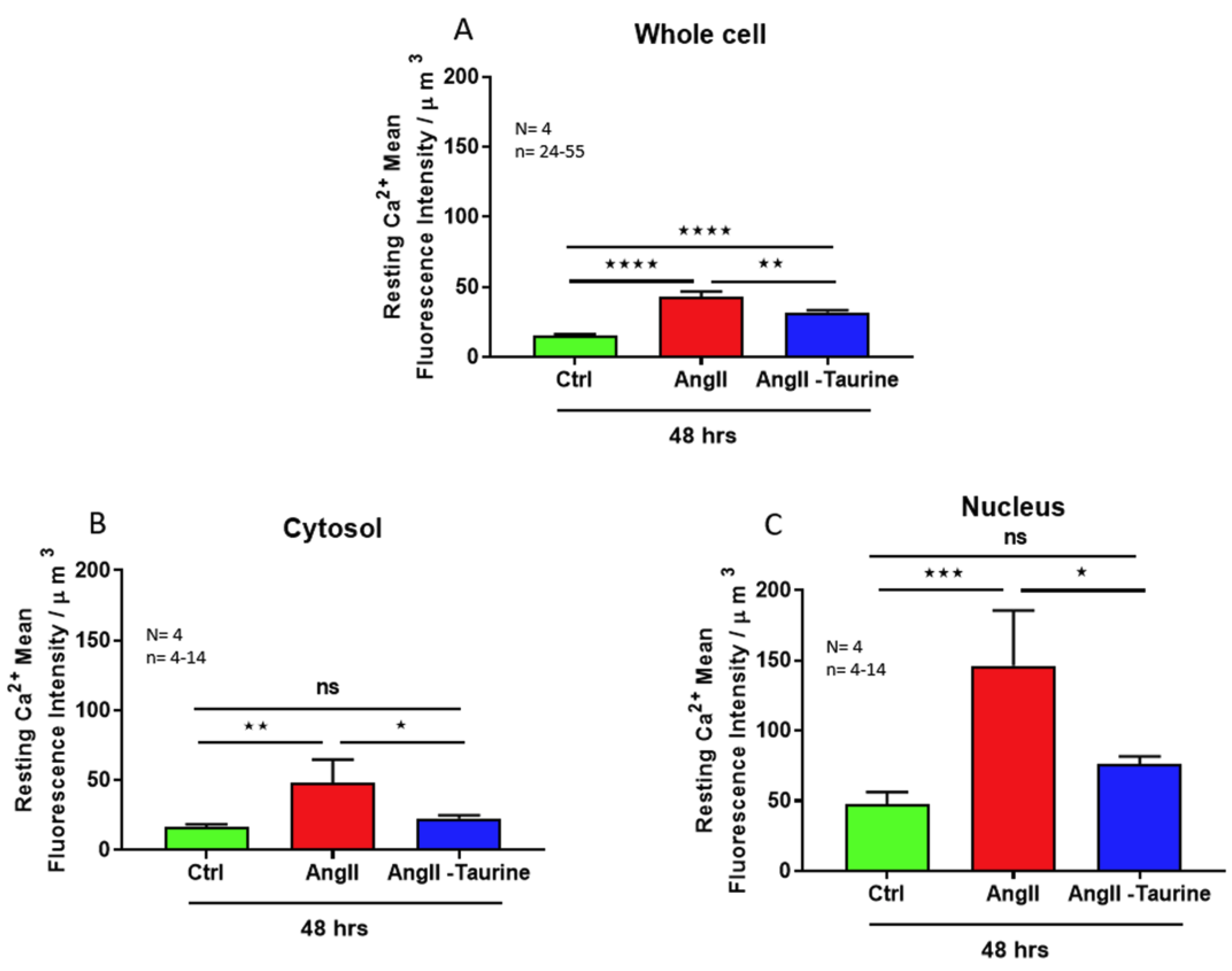
Figure 5.
An example of quantitative 3D confocal microscopy images (3D fishnet plot representation) showing the apparent cytoplasmic and nucleoplasmic ROS levels in hEECs. Image (A) represents hEECs ROS levels and distribution in a normal control medium. Image (B) represents hEECs ROS levels and distribution in the presence of Ang II (10−7 M) for 48 h. Image (C) represents hEECs ROS levels and distribution in the presence of 48 h of treatment with Ang II (10−7 M) and taurine (20 mM). Insert panels (A–C) in green color (color has no measurable meaning) represent syto-11-labeled nucleus in the absence of angiotensin II (Ang II) (A), in the presence of 48 h treatment with Ang II (B), and in the presence of Ang II and taurine (C). The white bar calibration is in μm, and (A–C) are from different cells. The color calibration bar is ROS fluorescence levels ranging from 0 (black color) to 255 (white).
Figure 5.
An example of quantitative 3D confocal microscopy images (3D fishnet plot representation) showing the apparent cytoplasmic and nucleoplasmic ROS levels in hEECs. Image (A) represents hEECs ROS levels and distribution in a normal control medium. Image (B) represents hEECs ROS levels and distribution in the presence of Ang II (10−7 M) for 48 h. Image (C) represents hEECs ROS levels and distribution in the presence of 48 h of treatment with Ang II (10−7 M) and taurine (20 mM). Insert panels (A–C) in green color (color has no measurable meaning) represent syto-11-labeled nucleus in the absence of angiotensin II (Ang II) (A), in the presence of 48 h treatment with Ang II (B), and in the presence of Ang II and taurine (C). The white bar calibration is in μm, and (A–C) are from different cells. The color calibration bar is ROS fluorescence levels ranging from 0 (black color) to 255 (white).

Figure 6.
In hEECs, treatment for 48 h with Ang II (10−7 M)-induced elevation in ROS levels in the whole cell ((A), red color), the cytoplasm ((B), red color), and the nucleoplasm ((C), red color). Treatment with taurine for 48 h partially but significantly prevented the Ang II-induced increase in ROS levels in the whole cell ((A), blue color), and in the cytoplasm ((B), blue color), but not in the nucleus ((C), blue color). The results are expressed as mean ± SEM, and n is the number of cells from three donors (N). * p < 0.05; ** p < 0.01; *** p < 0.001; ns: not significant. Results are presented per µm3 of the cell volume.
Figure 6.
In hEECs, treatment for 48 h with Ang II (10−7 M)-induced elevation in ROS levels in the whole cell ((A), red color), the cytoplasm ((B), red color), and the nucleoplasm ((C), red color). Treatment with taurine for 48 h partially but significantly prevented the Ang II-induced increase in ROS levels in the whole cell ((A), blue color), and in the cytoplasm ((B), blue color), but not in the nucleus ((C), blue color). The results are expressed as mean ± SEM, and n is the number of cells from three donors (N). * p < 0.05; ** p < 0.01; *** p < 0.001; ns: not significant. Results are presented per µm3 of the cell volume.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jacques, D.; Bkaily, G. Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS. Nutrients 2024, 16, 745. https://doi.org/10.3390/nu16050745
AMA Style
Jacques D, Bkaily G. Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS. Nutrients. 2024; 16(5):745. https://doi.org/10.3390/nu16050745
Chicago/Turabian StyleJacques, Danielle, and Ghassan Bkaily. 2024. "Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS" Nutrients 16, no. 5: 745. https://doi.org/10.3390/nu16050745
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.