
Potential Complementary Effect of Zinc and Alkalihalobacillus clausii on Gut Health and Immunity: A Narrative Review
 ,
,
Abstract
:1. Introduction
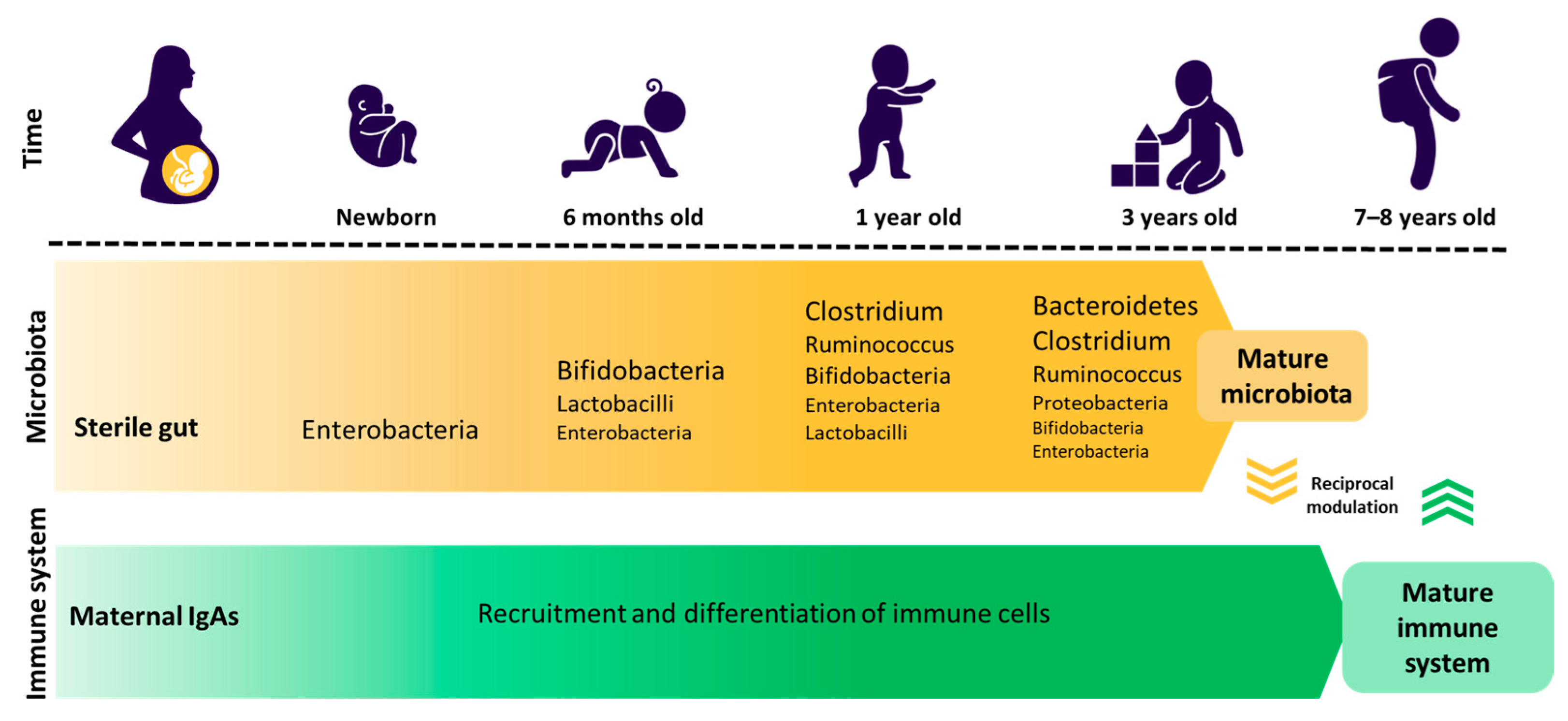
2. Gut Microbiota and Dysbiosis
2.1. Dysbiosis-Related Diseases and Conditions
2.2. Alkalihalobacillus clausii
3. Zinc
3.1. Dietary Needs and Biological Role
3.2. Prevalence and Health Consequences of Inadequate Zinc Intake and Zinc Deficiency
3.2.1. Prevalence of Inadequate Zinc Intake
3.2.2. Prevalence of Zinc Deficiency
3.2.3. Health Consequences of Inadequate Zinc Intake and Zinc Deficiency
3.3. Zinc Supplementation
3.3.1. Zinc Supplementation for Disease Treatment
3.3.2. Zinc Supplementation for Disease Prevention
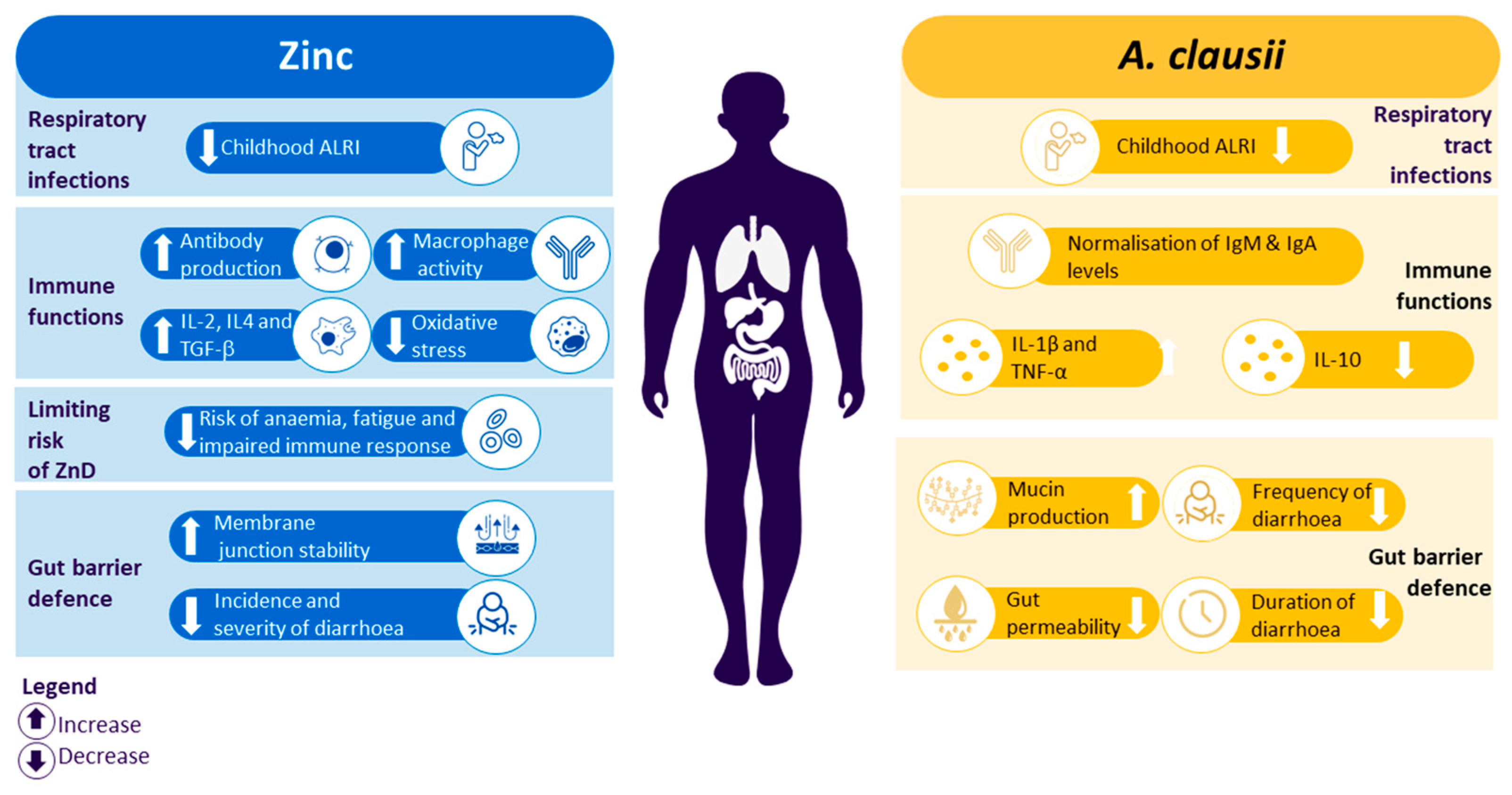
4. Benefits of the Combination of Zinc and A. clausii
4.1. Mechanism of Action: Potential Complementarity of Zinc and A. clausii
4.2. Potential Clinical Applications
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chiu, L.; Bazin, T.; Truchetet, M.E.; Schaeverbeke, T.; Delhaes, L.; Pradeu, T. Protective microbiota: From localized to long-reaching co-immunity. Front. Immunol. 2017, 8, 1678. [Google Scholar] [CrossRef]
- Wiertsema, S.P.; van Bergenhenegouwen, J.; Garssen, J.; Knippels, L.M.J. The interplay between the gut microbiome and the immune system in the context of infectious diseases throughout life and the role of nutrition in optimizing treatment strategies. Nutrients 2021, 13, 886. [Google Scholar] [CrossRef]
- Dukowicz, A.C.; Lacy, B.E.; Levine, G.M. Small intestinal bacterial overgrowth: A comprehensive review. Gastroenterol. Hepatol. 2007, 3, 112–122. [Google Scholar]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Lynn, D.J.; Benson, S.C.; Lynn, M.A.; Pulendran, B. Modulation of immune responses to vaccination by the microbiota: Implications and potential mechanisms. Nat. Rev. Immunol. 2022, 22, 33–46. [Google Scholar] [CrossRef]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lécuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G.; et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef]
- Gaufin, T.; Tobin, N.H.; Aldrovandi, G.M. The importance of the microbiome in pediatrics and pediatric infectious diseases. Curr. Opin. Pediatr. 2018, 30, 117–124. [Google Scholar] [CrossRef]
- Dupont, H.L.; Jiang, Z.D.; Dupont, A.W.; Utay, N.S. The intestinal microbiome in human health and disease. Trans. Am. Clin. Climatol. Assoc. 2020, 131, 178–197. [Google Scholar]
- Gagliardi, A.; Totino, V.; Cacciotti, F.; Iebba, V.; Neroni, B.; Bonfiglio, G.; Trancassini, M.; Passariello, C.; Pantanella, F.; Schippa, S. Rebuilding the gut microbiota ecosystem. Int. J. Environ. Res. Public. Health 2018, 15, 1679. [Google Scholar] [CrossRef]
- Fuller, R. Probiotics: The Scientific Basis; Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Boggio Marzet, C.; Burgos, F.; Del Compare, M.; Gerold, I.; Tabacco, O.; Vinderola, G. Approach to probiotics in pediatrics: The role of Lactobacillus rhamnosus GG. Arch. Argent. Pediatr. 2022, 120, e1–e7. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Franceschi, F.; Gasbarrini, A. Bacillus clausii and gut homeostasis: State of the art and future perspectives. Expert. Rev. Gastroenterol. Hepatol. 2016, 10, 943–948. [Google Scholar] [CrossRef]
- Wong-Chew, R.M.; de Castro, J.A.; Morelli, L.; Perez, M.; Ozen, M. Gut immune homeostasis: The immunomodulatory role of Bacillus clausii, from basic to clinical evidence. Expert. Rev. Clin. Immunol. 2022, 18, 717–729. [Google Scholar] [CrossRef]
- de Castro, J.A.; Guno, M.J.V.; Perez, M.O. Bacillus clausii as adjunctive treatment for acute community-acquired diarrhea among Filipino children: A large-scale, multicenter, open-label study (CODDLE). Trop. Dis. Travel. Med. Vaccines 2019, 5, 14. [Google Scholar] [CrossRef]
- Lahiri, K.R.; Singh, R.; Apte, M.; Patil, M.; Taksande, A.; Varona, R.; Chatterjee, G.; Verma, M.; Brette, S.; Perez, M.I. Efficacy and safety of Bacillus clausii (O/C, N/R, SIN, T) probiotic combined with oral rehydration therapy (ORT) and zinc in acute diarrhea in children: A randomized, double-blind, placebo-controlled study in India. Trop. Dis. Travel. Med. Vaccines 2022, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, B.; Guha, D.; Naik, A.K.; Banerjee, A.; Tambat, S.; Chawla, S.; Senapati, S.; Aich, P. Probiotics L. acidophilus and B. clausii modulate gut microbiota in Th1- and Th2-biased mice to ameliorate Salmonella typhimurium-induced diarrhea. Probiotics Antimicrob. Proteins 2018, 11, 887–904. [Google Scholar] [CrossRef] [PubMed]
- Ghelardi, E.; Abreu, Y.A.A.T.; Marzet, C.B.; Alvarez Calatayud, G.; Perez, M., 3rd; Moschione Castro, A.P. Current progress and future perspectives on the use of Bacillus clausii. Microorganisms 2022, 10, 1246. [Google Scholar] [CrossRef] [PubMed]
- Marseglia, G.L.; Tosca, M.; Cirillo, I.; Licari, A.; Leone, M.; Marseglia, A.; Castellazzi, A.M.; Ciprandi, G. Efficacy of Bacillus clausii spores in the prevention of recurrent respiratory infections in children: A pilot study. Ther. Clin. Risk Manag. 2007, 3, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Barone, M.; D'Amico, F.; Brigidi, P.; Turroni, S. Gut microbiome-micronutrient interaction: The key to controlling the bioavailability of minerals and vitamins? Biofactors 2022, 48, 307–314. [Google Scholar] [CrossRef]
- Chen, X.; Jiang, Y.; Wang, Z.; Chen, Y.; Tang, S.; Wang, S.; Su, L.; Huang, X.; Long, D.; Wang, L.; et al. Alteration in gut microbiota associated with zinc deficiency in school-age children. Nutrients 2022, 14, 2895. [Google Scholar] [CrossRef] [PubMed]
- Trumbo, P.; Yates, A.A.; Schlicker, S.; Poos, M. Dietary reference intakes: Vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc. J. Am. Diet. Assoc. 2001, 101, 294–301. [Google Scholar] [CrossRef]
- McCall, K.A.; Huang, C.; Fierke, C.A. Function and mechanism of zinc metalloenzymes. J. Nutr. 2000, 130, 1437s–1446s. [Google Scholar] [CrossRef]
- Kociova, S.; Dolezelikova, K.; Horky, P.; Skalickova, S.; Baholet, D.; Bozdechova, L.; Vaclavkova, E.; Belkova, J.; Nevrkla, P.; Skladanka, J.; et al. Zinc phosphate-based nanoparticles as alternatives to zinc oxide in diet of weaned piglets. J. Anim. Sci. Biotechnol. 2020, 11, 59. [Google Scholar] [CrossRef] [PubMed]
- Baltaci, A.K.; Yuce, K.; Mogulkoc, R. Zinc metabolism and metallothioneins. Biol. Trace Elem. Res. 2018, 183, 22–31. [Google Scholar] [CrossRef]
- Maret, W. Zinc biochemistry: From a single zinc enzyme to a key element of life. Adv. Nutr. 2013, 4, 82–91. [Google Scholar] [CrossRef]
- Maggini, S.; Pierre, A.; Calder, P. Immune function and micronutrient requirements change over the life course. Nutrients 2018, 10, 1531. [Google Scholar] [CrossRef]
- Wessels, I.; Maywald, M.; Rink, L. Zinc as a gatekeeper of immune function. Nutrients 2017, 9, 1286. [Google Scholar] [CrossRef]
- Salgueiro, M.J.; Zubillaga, M.B.; Lysionek, A.E.; Caro, R.A.; Weill, R.; Boccio, J.R. The role of zinc in the growth and development of children. Nutrition 2002, 18, 510–519. [Google Scholar] [CrossRef]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on dietary reference values for zinc. EFSA J. 2014, 12, 3844. [Google Scholar] [CrossRef]
- Wessells, K.R.; Singh, G.M.; Brown, K.H. Estimating the global prevalence of inadequate zinc intake from national food balance sheets: Effects of methodological assumptions. PLoS ONE 2012, 7, e50565. [Google Scholar] [CrossRef]
- Maxfield, L.; Shukla, S.; Crane, J.S. Zinc deficiency. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2023. [Google Scholar]
- Sugawara, D.; Makita, E.; Matsuura, M.; Ichihashi, K. The association between serum zinc levels and anthropometric measurements and nutritional indicators in children with idiopathic short stature. Cureus 2022, 14, e24906. [Google Scholar] [CrossRef]
- Imdad, A.; Rogner, J.; Sherwani, R.N.; Sidhu, J.; Regan, A.; Haykal, M.R.; Tsistinas, O.; Smith, A.; Chan, X.H.S.; Mayo-Wilson, E.; et al. Zinc supplementation for preventing mortality, morbidity, and growth failure in children aged 6 months to 12 years. Cochrane Database Syst. Rev. 2023, 3, CD009384. [Google Scholar] [CrossRef]
- Yakoob, M.Y.; Theodoratou, E.; Jabeen, A.; Imdad, A.; Eisele, T.P.; Ferguson, J.; Jhass, A.; Rudan, I.; Campbell, H.; Black, R.E.; et al. Preventive zinc supplementation in developing countries: Impact on mortality and morbidity due to diarrhea, pneumonia and malaria. BMC Public. Health 2011, 11 (Suppl. 3), S23. [Google Scholar] [CrossRef]
- Ciprandi, G.; Vizzaccaro, A.; Cirillo, I.; Tosca, M.A. Bacillus clausii exerts immuno-modulatory activity in allergic subjects: A pilot study. Eur. Ann. Allergy Clin. Immunol. 2005, 37, 129–134. [Google Scholar] [PubMed]
- Greifenberg, D.M.; Perez, M. Zinc and Bacillus clausii: A promising combination to improve immunity in children? Evid. Self-Medicat. 2022, 2, 220084. [Google Scholar] [CrossRef]
- Maywald, M.; Rink, L. Zinc in human health and infectious diseases. Biomolecules 2022, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Agans, R.; Rigsbee, L.; Kenche, H.; Michail, S.; Khamis, H.J.; Paliy, O. Distal gut microbiota of adolescent children is different from that of adults. FEMS Microbiol. Ecol. 2011, 77, 404–412. [Google Scholar] [CrossRef]
- Arrieta, M.C.; Stiemsma, L.T.; Amenyogbe, N.; Brown, E.M.; Finlay, B. The intestinal microbiome in early life: Health and disease. Front. Immunol. 2014, 5, 427. [Google Scholar] [CrossRef]
- Mesa, M.D.; Loureiro, B.; Iglesia, I.; Fernandez Gonzalez, S.; Llurba Olive, E.; Garcia Algar, O.; Solana, M.J.; Cabero Perez, M.J.; Sainz, T.; Martinez, L.; et al. The evolving microbiome from pregnancy to early infancy: A comprehensive review. Nutrients 2020, 12, 133. [Google Scholar] [CrossRef]
- Yu, L.C.; Wang, J.T.; Wei, S.C.; Ni, Y.H. Host-microbial interactions and regulation of intestinal epithelial barrier function: From physiology to pathology. World J. Gastrointest. Pathophysiol. 2012, 3, 27–43. [Google Scholar] [CrossRef]
- Libertucci, J.; Young, V.B. The role of the microbiota in infectious diseases. Nat. Microbiol. 2019, 4, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Smith, P.M.; Garrett, W.S. The gut microbiota and mucosal T cells. Front. Microbiol. 2011, 2, 111. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.L.; Malfair, D.; Gray, D.; Doyle, J.S.; Jewell, L.D.; Fedorak, R.N. Interleukin-10 gene-deficient mice develop a primary intestinal permeability defect in response to enteric microflora. Inflamm. Bowel Dis. 1999, 5, 262–270. [Google Scholar] [CrossRef]
- Cao, Y.; Shen, J.; Ran, Z.H. Association between Faecalibacterium prausnitzii reduction and inflammatory bowel disease: A meta-analysis and systematic review of the literature. Gastroenterol. Res. Pract. 2014, 2014, 872725. [Google Scholar] [CrossRef]
- Michail, S.; Durbin, M.; Turner, D.; Griffiths, A.M.; Mack, D.R.; Hyams, J.; Leleiko, N.; Kenche, H.; Stolfi, A.; Wine, E. Alterations in the gut microbiome of children with severe ulcerative colitis. Inflamm. Bowel Dis. 2012, 18, 1799–1808. [Google Scholar] [CrossRef]
- Wang, Z.J.; Chen, X.F.; Zhang, Z.X.; Li, Y.C.; Deng, J.; Tu, J.; Song, Z.Q.; Zou, Q.H. Effects of anti-Helicobacter pylori concomitant therapy and probiotic supplementation on the throat and gut microbiota in humans. Microb. Pathog. 2017, 109, 156–161. [Google Scholar] [CrossRef]
- Stewart, C.J.; Marrs, E.C.; Magorrian, S.; Nelson, A.; Lanyon, C.; Perry, J.D.; Embleton, N.D.; Cummings, S.P.; Berrington, J.E. The preterm gut microbiota: Changes associated with necrotizing enterocolitis and infection. Acta Paediatr. 2012, 101, 1121–1127. [Google Scholar] [CrossRef]
- Abbrescia, A.; Palese, L.L.; Papa, S.; Gaballo, A.; Alifano, P.; Sardanelli, A.M. Antibiotic sensitivity of Bacillus clausii strains in commercial preparation. Curr. Med. Chem. 2014, 1, 102–110. [Google Scholar] [CrossRef]
- Courvalin, P. Antibiotic resistance: The pros and cons of probiotics. Dig. Liver Dis. 2006, 38 (Suppl. 2), S261–S265. [Google Scholar] [CrossRef]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Gueye, S.A.; Lupetti, A.; Senesi, S. Survival and persistence of Bacillus clausii in the human gastrointestinal tract following oral administration as spore-based probiotic formulation. J. Appl. Microbiol. 2015, 119, 552–559. [Google Scholar] [CrossRef]
- Rochín-Medina, J.J.; Ramírez-Medina, H.K.; Rangel-Peraza, J.G.; Pineda-Hidalgo, K.V.; Iribe-Arellano, P. Use of whey as a culture medium for Bacillus clausii for the production of protein hydrolysates with antimicrobial and antioxidant activity. Food Sci. Technol. Int. 2018, 24, 35–42. [Google Scholar] [CrossRef]
- Ilinskaya, O.N.; Ulyanova, V.V.; Yarullina, D.R.; Gataullin, I.G. Secretome of intestinal bacilli: A natural guard against pathologies. Front. Microbiol. 2017, 8, 1666. [Google Scholar] [CrossRef] [PubMed]
- Urdaci, M.C.; Bressollier, P.; Pinchuk, I. Bacillus clausii probiotic strains: Antimicrobial and immunomodulatory activities. J. Clin. Gastroenterol. 2004, 38, S86–S90. [Google Scholar] [CrossRef]
- Ripert, G.; Racedo, S.M.; Elie, A.M.; Jacquot, C.; Bressollier, P.; Urdaci, M.C. Secreted compounds of the probiotic Bacillus clausii strain O/C inhibit the cytotoxic effects induced by Clostridium difficile and Bacillus cereus toxins. Antimicrob. Agents Chemother. 2016, 60, 3445–3454. [Google Scholar] [CrossRef] [PubMed]
- Paparo, L.; Tripodi, L.; Bruno, C.; Pisapia, L.; Damiano, C.; Pastore, L.; Berni Canani, R. Protective action of Bacillus clausii probiotic strains in an in vitro model of Rotavirus infection. Sci. Rep. 2020, 10, 12636. [Google Scholar] [CrossRef] [PubMed]
- Pirozzi, C.; Opallo, N.; Coretti, L.; Lama, A.; Annunziata, C.; Comella, F.; Melini, S.; Buommino, E.; Mollica, M.P.; Aviello, G.; et al. Alkalihalobacillus clausii (formerly Bacillus clausii) spores lessen antibiotic-induced intestinal injury and reshape gut microbiota composition in mice. Biomed. Pharmacother. 2023, 163, 114860. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Cirillo, P.; Terrin, G.; Cesarano, L.; Spagnuolo, M.I.; De Vincenzo, A.; Albano, F.; Passariello, A.; De Marco, G.; Manguso, F.; et al. Probiotics for treatment of acute diarrhoea in children: Randomised clinical trial of five different preparations. BMJ 2007, 335, 340. [Google Scholar] [CrossRef]
- García, J.P.; Hoyos, J.A.; Alzate, J.A.; Cristancho, E. Bacteremia after Bacillus clausii administration for the treatment of acute diarrhea: A case report. Biomedica 2021, 41, 13–20. [Google Scholar] [CrossRef]
- Princess, I.; Natarajan, T.; Ghosh, S. When good bacteria behave badly: A case report of Bacillus clausii sepsis in an immunocompetant adult. Access Microbiol. 2020, 2, acmi000097. [Google Scholar] [CrossRef]
- Joshi, S.; Udani, S.; Sen, S.; Kirolikar, S.; Shetty, A. Bacillus clausii septicemia in a pediatric patient after treatment with probiotics. Pediatr. Infect. Dis. J. 2019, 38, e228–e230. [Google Scholar] [CrossRef]
- Vuralli, D.; Tumer, L.; Hasanoglu, A. Zinc deficiency in the pediatric age group is common but under evaluated. World J. Pediatr. 2017, 13, 360–366. [Google Scholar] [CrossRef]
- WHO. Trace Elements in Human Nutrition and Health; World Health Organization, International Atomic Energy Agency & Food and Agriculture Organization of the United Nations: Vienna, Austria, 1996. [Google Scholar]
- Meyers, L.D.; Hellwig, J.P.; Otten, J.J. Dietary Reference Intakes: The Essential Guide to Nutrient Requirements; National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Brown, K.H.; Rivera, J.A.; Bhutta, Z.; Gibson, R.S.; King, J.C.; Lönnerdal, B.; Ruel, M.T.; Sandtröm, B.; Wasantwisut, E.; Hotz, C. International Zinc Nutrition Consultative Group (IZiNCG) technical document #1. Assessment of the risk of zinc deficiency in populations and options for its control. Food Nutr. Bull. 2004, 25, S99–S203. [Google Scholar]
- Wuehler, S.; Lopez de Romaña, D.; Haile, D.; McDonald, C.M.; Brown, K.H. Reconsidering the tolerable upper levels of zinc intake among infants and young children: A systematic review of the available evidence. Nutrients 2022, 14, 1938. [Google Scholar] [CrossRef]
- Hojyo, S.; Fukada, T. Roles of zinc signaling in the immune system. J. Immunol. Res. 2016, 2016, 6762343. [Google Scholar] [CrossRef]
- Sadeghsoltani, F.; Mohammadzadeh, I.; Safari, M.-M.; Hassanpour, P.; Izadpanah, M.; Qujeq, D.; Moein, S.; Vaghari-Tabari, M. Zinc and respiratory viral infections: Important trace element in anti-viral response and immune regulation. Biol. Trace Elem. Res. 2022, 200, 2556–2571. [Google Scholar] [CrossRef] [PubMed]
- Pecora, F.; Persico, F.; Argentiero, A.; Neglia, C.; Esposito, S. The role of micronutrients in support of the immune response against viral infections. Nutrients 2020, 12, 3198. [Google Scholar] [CrossRef] [PubMed]
- Gammoh, N.Z.; Rink, L. Zinc in infection and inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef]
- Marreiro, D.D.; Cruz, K.J.; Morais, J.B.; Beserra, J.B.; Severo, J.S.; de Oliveira, A.R. Zinc and oxidative stress: Current mechanisms. Antioxidants 2017, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- José, J.; Souza, A.C.R.; Vasconcelos, A.R.; Prado, P.S.; Pereira, C.P.M. Zinc, vitamin D and vitamin C: Perspectives for COVID-19 with a focus on physical tissue barrier integrity. Front. Nutr. 2020, 7, 606398. [Google Scholar] [CrossRef]
- Ohashi, W.; Fukada, T. Contribution of zinc and zinc transporters in the pathogenesis of inflammatory bowel diseases. J. Immunol. Res. 2019, 2019, 8396878. [Google Scholar] [CrossRef]
- Mullin, J.M.; Skrovanek, S.M.; Valenzano, M.C. Modification of tight junction structure and permeability by nutritional means. Ann. N. Y. Acad. Sci. 2009, 1165, 99–112. [Google Scholar] [CrossRef] [PubMed]
- López-Moreno, M.; Garcés-Rimón, M.; Miguel, M. Antinutrients: Lectins, goitrogens, phytates and oxalates, friends or foe? J. Funct. Foods 2022, 89, 104938. [Google Scholar] [CrossRef]
- Dao, D.T.; Anez-Bustillos, L.; Cho, B.S.; Li, Z.; Puder, M.; Gura, K.M. Assessment of micronutrient status in critically ill children: Challenges and opportunities. Nutrients 2017, 9, 1185. [Google Scholar] [CrossRef]
- Killilea, D.W.; Schultz, K. Pre-analytical variables influence zinc measurement in blood samples. PLoS ONE 2023, 18, e0286073. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.H.; Wuehler, S.E.; Peerson, J.M. The importance of zinc in human nutrition and estimation of the global prevalence of zinc deficiency. Food Nutr. Bull. 2001, 22, 113–125. [Google Scholar] [CrossRef]
- Wuehler, S.E.; Peerson, J.M.; Brown, K.H. Use of national food balance data to estimate the adequacy of zinc in national food supplies: Methodology and regional estimates. Public. Health Nutr. 2005, 8, 812–819. [Google Scholar] [CrossRef]
- De Benoist, B.; Darnton-Hill, I.; Davidsson, L.; Fontaine, O. Report of a WHO/UNICEF/IAEA/IZiNCG interagency meeting on zinc status indicators, held in IAEA headquarters, Vienna, December 9, 2005-Foreword. Food Nutr. Bull. 2007, 28, S401–S402. [Google Scholar] [CrossRef]
- Chao, H.C. Zinc deficiency and therapeutic value of zinc supplementation in pediatric gastrointestinal diseases. Nutrients 2023, 15, 4093. [Google Scholar] [CrossRef]
- Marcos, J.M.; Perlas, L.A.; Gironella, G.M.P. Risk factors associated with zinc status of Filipino preschool and school-aged children. Philipp. J. Sci. 2019, 2, 225–236. [Google Scholar]
- Wieser, S.; Plessow, R.; Eichler, K.; Malek, O.; Capanzana, M.V.; Agdeppa, I.; Bruegger, U. Burden of micronutrient deficiencies by socio-economic strata in children aged 6 months to 5 years in the Philippines. BMC Public. Health 2013, 13, 1167. [Google Scholar] [CrossRef]
- Palanog, A.D.; Calayugan, M.I.C.; Descalsota-Empleo, G.I.; Amparado, A.; Inabangan-Asilo, M.A.; Arocena, E.C.; Sta. Cruz, P.C.; Borromeo, T.H.; Lalusin, A.; Hernandez, J.E.; et al. Zinc and iron nutrition status in the Philippines population and local soils. Front. Nutr. 2019, 6, 81. [Google Scholar] [CrossRef]
- Laillou, A.; Pham, T.V.; Tran, N.T.; Le, H.T.; Wieringa, F.; Rohner, F.; Fortin, S.; Le, M.B.; Tran, D.T.; Moench-Pfanner, R.; et al. Micronutrient deficits are still public health issues among women and young children in Vietnam. PLoS ONE 2012, 7, e34906. [Google Scholar] [CrossRef]
- Hung, N.; Thi, P.; Huong, T.; Tran Thi Tra, P.; Phuong, T.; Truong, T.; Mai, T.; Nguyen, T.; Lam, N.; Luong, H.; et al. The current situation of anemia and zinc deficiency of children ages 6–59 months with anorexia WHO examined at national institute of nutrition. Sys. Rev. Pharm. 2021, 12, 1422–1425. [Google Scholar]
- Roos, N.; Ponce, M.C.; Doak, C.M.; Dijkhuizen, M.; Polman, K.; Chamnan, C.; Khov, K.; Chea, M.; Prak, S.; Kounnavong, S.; et al. Micronutrient status of populations and preventive nutrition interventions in South East Asia. Matern. Child. Health 2019, 23, 29–45. [Google Scholar] [CrossRef] [PubMed]
- Bouglé, D.; Laroche, D.; Bureau, F. Zinc and iron status and growth in healthy infants. Eur. J. Clin. Nutr. 2000, 54, 764–767. [Google Scholar] [CrossRef]
- Taylor, A.; Redworth, E.W.; Morgan, J.B. Influence of diet on iron, copper, and zinc status in children under 24 months of age. Biol. Trace Elem. Res. 2004, 97, 197–214. [Google Scholar] [CrossRef] [PubMed]
- Persson, L.A.; Lundström, M.; Lönnerdal, B.; Hernell, O. Are weaning foods causing impaired iron and zinc status in 1-year-old Swedish infants? A cohort study. Acta Paediatr. 1998, 87, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Vreugdenhil, M.; Akkermans, M.D.; van der Merwe, L.F.; van Elburg, R.M.; van Goudoever, J.B.; Brus, F. Prevalence of zinc deficiency in healthy 1-3-year-old children from three Western European Countries. Nutrients 2021, 13, 3713. [Google Scholar] [CrossRef] [PubMed]
- Galetti, V. Zinc deficiency and stunting. In Handbook of Famine, Starvation, and Nutrient Deprivation; Springer: Cham, Switzerland, 2019; pp. 1969–1991. [Google Scholar] [CrossRef]
- Wang, X.; Valenzano, M.C.; Mercado, J.M.; Zurbach, E.P.; Mullin, J.M. Zinc supplementation modifies tight junctions and alters barrier function of CACO-2 human intestinal epithelial layers. Dig. Dis. Sci. 2012, 58, 77–87. [Google Scholar] [CrossRef]
- Willoughby, J.L.; Bowen, C.N. Zinc deficiency and toxicity in pediatric practice. Curr. Opin. Pediatr. 2014, 26, 579–584. [Google Scholar] [CrossRef]
- Reed, S.; Neuman, H.; Moscovich, S.; Glahn, R.P.; Koren, O.; Tako, E. Chronic zinc deficiency alters chick gut microbiota composition and function. Nutrients 2015, 7, 9768–9784. [Google Scholar] [CrossRef]
- Oyagbemi, A.A.; Ajibade, T.O.; Aboua, Y.G.; Gbadamosi, I.T.; Adedapo, A.D.A.; Aro, A.O.; Adejumobi, O.A.; Thamahane-Katengua, E.; Omobowale, T.O.; Falayi, O.O.; et al. Potential health benefits of zinc supplementation for the management of COVID-19 pandemic. J. Food Biochem. 2021, 45, e13604. [Google Scholar] [CrossRef]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The role of zinc in antiviral immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef]
- Saleh, N.Y.; Abo El Fotoh, W.M.M. Low serum zinc level: The relationship with severe pneumonia and survival in critically ill children. Int. J. Clin. Pract. 2018, 72, e13211. [Google Scholar] [CrossRef]
- Nakamura, T.; Nishiyama, S.; Futagoishi-Suginohara, Y.; Matsuda, I.; Higashi, A. Mild to moderate zinc deficiency in short children: Effect of zinc supplementation on linear growth velocity. J. Pediatr. 1993, 123, 65–69. [Google Scholar] [CrossRef]
- Skalny, A.V.; Aschner, M.; Lei, X.G.; Gritsenko, V.A.; Santamaria, A.; Alekseenko, S.I.; Prakash, N.T.; Chang, J.-S.; Sizova, E.A.; Chao, J.C.J.; et al. Gut microbiota as a mediator of essential and toxic effects of zinc in the intestines and other tissues. Int. J. Mol. Sci. 2021, 22, 13074. [Google Scholar] [CrossRef] [PubMed]
- Shankar, A.H.; Prasad, A.S. Zinc and immune function: The biological basis of altered resistance to infection. Am. J. Clin. Nutr. 1998, 68, 447s–463s. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Urakami, T.; Kuwabara, R.; Morioka, I. Zinc deficiency in Japanese children with idiopathic short stature. JPEM 2019, 32, 1083–1087. [Google Scholar] [CrossRef] [PubMed]
- Hamza, R.T.; Hamed, A.I.; Sallam, M.T. Effect of zinc supplementation on growth hormone insulin growth factor axis in short Egyptian children with zinc deficiency. Ital. J. Pediatr. 2012, 38, 21. [Google Scholar] [CrossRef] [PubMed]
- Petkovic, V.; Miletta, M.C.; Eblé, A.; Iliev, D.I.; Binder, G.; Flück, C.E.; Mullis, P.E. Effect of zinc binding residues in growth hormone (GH) and altered intracellular zinc content on regulated GH secretion. Endocrinology 2013, 154, 4215–4225. [Google Scholar] [CrossRef]
- Dørup, I.; Flyvbjerg, A.; Everts, M.E.; Clausen, T. Role of insulin-like growth factor-1 and growth hormone in growth inhibition induced by magnesium and zinc deficiencies. Br. J. Nutr. 1991, 66, 505–521. [Google Scholar] [CrossRef]
- Jeng, S.-S.; Chen, Y.-H. Association of zinc with anemia. Nutrients 2022, 14, 4918. [Google Scholar] [CrossRef]
- Chen, Y.H.; Feng, H.L.; Jeng, S.S. Zinc supplementation stimulates red blood cell formation in rats. Int. J. Mol. Sci. 2018, 19, 2824. [Google Scholar] [CrossRef]
- Osawa, M.; Yamaguchi, T.; Nakamura, Y.; Kaneko, S.; Onodera, M.; Sawada, K.; Jegalian, A.; Wu, H.; Nakauchi, H.; Iwama, A. Erythroid expansion mediated by the Gfi-1B zinc finger protein: Role in normal hematopoiesis. Blood 2002, 100, 2769–2777. [Google Scholar] [CrossRef] [PubMed]
- Van Nhien, N.; Khan, N.C.; Ninh, N.X.; Van Huan, P.; Hople, T.; Lam, N.T.; Ota, F.; Yabutani, T.; Hoa, V.Q.; Motonaka, J.; et al. Micronutrient deficiencies and anemia among preschool children in rural Vietnam. Asia Pac. J. Clin. Nutr. 2008, 17, 48–55. [Google Scholar]
- Semrad, C.E. Zinc and intestinal function. Curr. Gastroenterol. Rep. 1999, 1, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Lazzerini, M.; Wanzira, H. Oral zinc for treating diarrhoea in children. Cochrane Database Syst. Rev. 2016, 12, CD005436. [Google Scholar] [CrossRef] [PubMed]
- WHO; UNICEF. Clinical Management of Acute Diarrhoea. 2004. Available online: https://www.who.int/publications/i/item/WHO_FCH_CAH_04.7 (accessed on 16 October 2023).
- Acevedo-Murillo, J.A.; García León, M.L.; Firo-Reyes, V.; Santiago-Cordova, J.L.; Gonzalez-Rodriguez, A.P.; Wong-Chew, R.M. Zinc supplementation promotes a Th1 response and improves clinical symptoms in fewer hours in children with pneumonia younger than 5 years old. A randomized controlled clinical trial. Front. Pediatr. 2019, 7, 431. [Google Scholar] [CrossRef]
- Park, S.G.; Choi, H.N.; Yang, H.R.; Yim, J.E. Effects of zinc supplementation on catch-up growth in children with failure to thrive. Nutr. Res. Pract. 2017, 11, 487–491. [Google Scholar] [CrossRef]
- Mozaffari-Khosravi, H.; Shakiba, M.; Eftekhari, M.-H.; Fatehi, F. Effects of zinc supplementation on physical growth in 2–5-year-old children. Biol. Trace Elem. Res. 2009, 128, 118–127. [Google Scholar] [CrossRef]
- Su, T.H.; Kao, J.H. The clinical manifestations and management of COVID-19-related liver injury. J. Formos. Med. Assoc. 2020, 119, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Li, C.; Zhao, D.; Xu, L.; Kuang, M.; Yao, X.; Hu, H. The Impact of zinc supplementation on critically Ill patients with acute kidney injury: A propensity score matching analysis. Front. Nutr. 2022, 9, 894572. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Ajami, M.; Abdollahi, Z.; Kalantari, N.; Houshiarrad, A.; Fozouni, F.; Fallahrokni, A.; Mazandarani, F.S. Zinc supplementation is an effective and feasible strategy to prevent growth retardation in 6 to 24 month children: A pragmatic double blind, randomized trial. Heliyon 2019, 5, e02581. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.H.; Peerson, J.M.; Baker, S.K.; Hess, S.Y. Preventive zinc supplementation among infants, preschoolers, and older prepubertal children. Food Nutr. Bull. 2009, 30, S12–S40. [Google Scholar] [CrossRef]
- Vlieg-Boerstra, B.; de Jong, N.; Meyer, R.; Agostoni, C.; De Cosmi, V.; Grimshaw, K.; Milani, G.P.; Muraro, A.; Oude Elberink, H.; Pali-Schöll, I.; et al. Nutrient supplementation for prevention of viral respiratory tract infections in healthy subjects: A systematic review and meta-analysis. Allergy 2022, 77, 1373–1388. [Google Scholar] [CrossRef]
- Abdollahi, M.; Abdollahi, Z.; Fozouni, F.; Bondarianzadeh, D. Oral zinc supplementation positively affects linear growth, but not weight, in children 6–24 months of age. Int. J. Prev. Med. 2014, 5, 280. [Google Scholar] [PubMed]
- Ceballos-Rasgado, M.; Lowe, N.M.; Mallard, S.; Clegg, A.; Moran, V.H.; Harris, C.; Montez, J.; Xipsiti, M. Adverse effects of excessive zinc intake in infants and children aged 0–3 years: A systematic review and meta-analysis. Adv. Nutr. 2022, 13, 2488–2518. [Google Scholar] [CrossRef]
- Mayo-Wilson, E.; Junior, J.A.; Imdad, A.; Dean, S.; Chan, X.H.; Chan, E.S.; Jaswal, A.; Bhutta, Z.A. Zinc supplementation for preventing mortality, morbidity, and growth failure in children aged 6 months to 12 years of age. Cochrane Database Syst. Rev. 2014, 5, CD009384. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Summary of Tolerable Upper Intake Levels—Version 4 (September 2018). Available online: https://www.efsa.europa.eu/sites/default/files/assets/UL_Summary_tables.pdf (accessed on 8 August 2023).
- Institute of Medicine, F.; Nutrition, B. The National Academies Collection: Reports funded by National Institutes of Health. In Dietary Reference Intakes: A Risk Assessment Model for Establishing Upper Intake Levels for Nutrients; National Academies Press (US), National Academy of Sciences: Washington, DC, USA, 2020. [Google Scholar]
- Cai, Y.; Wang, X.; Li, C.; Li, F.; Yan, Z.; Ma, N.; Sun, M. Probiotics combined with zinc and selenium preparation in the treatment of child rotavirus enteritis. Am. J. Transl. Res. 2022, 14, 1043–1050. [Google Scholar]
- Lahiri, K.; Jadhav, K.; Gahlowt, P.; Najmuddin, F. Bacillus clausii as an adjuvant therapy in acute childhood diarrhoea. IOSR 2015, 14, 74–76. [Google Scholar]
- Malik, A.; Haldar, P.; Ray, A.; Shet, A.; Kapuria, B.; Bhadana, S.; Santosham, M.; Ghosh, R.S.; Steinglass, R.; Kumar, R. Introducing rotavirus vaccine in the Universal Immunization Programme in India: From evidence to policy to implementation. Vaccine 2019, 37, 5817–5824. [Google Scholar] [CrossRef]
- Shamir, R.; Makhoul, I.R.; Etzioni, A.; Shehadeh, N. Evaluation of a diet containing probiotics and zinc for the treatment of mild diarrheal illness in children younger than one year of age. J. Am. Coll. Nutr. 2005, 24, 370–375. [Google Scholar] [CrossRef]
- Tarantino, G.; Finelli, C. Systematic review on intervention with prebiotics/probiotics in patients with obesity-related nonalcoholic fatty liver disease. Fut. Microb. 2015, 10, 889–902. [Google Scholar] [CrossRef]
- Zhang, B.; Guo, Y. Supplemental zinc reduced intestinal permeability by enhancing occludin and zonula occludens protein-1 (ZO-1) expression in weaning piglets. Br. J. Nutr. 2009, 102, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, E.; Balsiger, L.M.; Maurizi, V.; Rinninella, E.; Gasbarrini, A.; Giostra, N.; Santori, P.; Abenavoli, L.; Rasetti, C. Zinc and gut microbiota in health and gastrointestinal disease under the COVID-19 suggestion. Biofactors 2022, 48, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Vahjen, W.; Neumann, K.; Van Kessel, A.G.; Zentek, J. Dose-dependent effects of dietary zinc oxide on bacterial communities and metabolic profiles in the ileum of weaned pigs. J. Anim. Physiol. Anim. Nutr. 2012, 96, 825–833. [Google Scholar] [CrossRef]
- Velasco, E.; Wang, S.; Sanet, M.; Fernández-Vázquez, J.; Jové, D.; Glaría, E.; Valledor, A.F.; O'Halloran, T.V.; Balsalobre, C. A new role for Zinc limitation in bacterial pathogenicity: Modulation of α-hemolysin from uropathogenic Escherichia coli. Sci. Rep. 2018, 8, 6535. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.J.; Sarker, P.; Roy, S.K.; Ahmad, S.M.; Chisti, J.; Azim, T.; Mathan, M.; Sack, D.; Andersson, J.; Raqib, R. Effects of zinc supplementation as adjunct therapy on the systemic immune responses in shigellosis. Am. J. Clin. Nutr. 2005, 81, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Raqib, R.; Roy, S.K.; Rahman, M.J.; Azim, T.; Ameer, S.S.; Chisti, J.; Andersson, J. Effect of zinc supplementation on immune and inflammatory responses in pediatric patients with shigellosis. Am. J. Clin. Nutr. 2004, 79, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Sazawal, S.; Jalla, S.; Mazumder, S.; Sinha, A.; Black, R.E.; Bhan, M.K. Effect of zinc supplementation on cell-mediated immunity and lymphocyte subsets in preschool children. Indian Pediatr. 1997, 34, 589–597. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EFSA | IOM | IZiNCG | WHO | ||||
|---|---|---|---|---|---|---|---|
| Age, Sex | RDA, mg/day | Age, Sex | RDA, mg/day | Age, Sex | RDA, mg/day | Age, Sex a | RDA, mg/day |
| – | 0–6 months | 2.0 b | – | 0–6 months | 2.8 | ||
| 7–11 months | 2.9 | 7–12 months | 3.0 | 6–11 months | 4.0 | 7–12 months | 4.1 |
| 1–3 years | 4.3 | 1–3 years | 3.0 | 1–3 years | 3.0 | 1–3 years | 4.1 |
| 4–6 years | 5.5 | 4–8 years | 5.0 | 4–8 years | 4.0 | 4–6 years | 4.8 |
| 7–10 years | 7.4 | 7–9 years | 5.6 | ||||
| 11–14 years | 9.4 | 9–13 years | 8.0 | 9–13 years | 6.0 | – | |
| 15–17 years, M | 12.5 | 14–18 years, M | 11.0 | 14–18 years, M | 10.0 | 10–18 years, M | 8.6 |
| 15–17 years, F | 10.4 | 14–18 years, F | 9.0 | 14–18 years, F | 9.0 | 10–18 years, F | 7.2 |
| Region at High Risk of Zinc Deficiency | Country | Estimated Prevalence of Inadequate Zinc Intake (%) |
|---|---|---|
| South Asia | Indonesia | 31 |
| Sri Lanka | 31 | |
| India | 31 | |
| Sub-Saharan Africa | Guinea-Bissau | 27 |
| Kenya | 25 | |
| Liberia | 35 | |
| Malawi | 41 | |
| Zambia | 45 | |
| Zimbabwe | 48 | |
| Central America | El Salvador | 28 |
| Guatemala | 30 | |
| Nicaragua | 29 |
| EFSA | IOM | |
|---|---|---|
| Age, Years | ULs, mg/day | |
| 0–6 months | - | 4 |
| 7–11 months | - | 5 |
| 1–3 years | 7 | - |
| 4–6 years | 10 | - |
| 4–8 years | - | 12 |
| 7–10 years | 10 | - |
| 9–13 years | - | 23 |
| 11–14 years | 18 | - |
| 14–18 years | - | 34 |
| 15–17 years | 22 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong-Chew, R.M.; Nguyen, T.V.H.; Rogacion, J.M.; Herve, M.; Pouteau, E. Potential Complementary Effect of Zinc and Alkalihalobacillus clausii on Gut Health and Immunity: A Narrative Review. Nutrients 2024, 16, 887. https://doi.org/10.3390/nu16060887
Wong-Chew RM, Nguyen TVH, Rogacion JM, Herve M, Pouteau E. Potential Complementary Effect of Zinc and Alkalihalobacillus clausii on Gut Health and Immunity: A Narrative Review. Nutrients. 2024; 16(6):887. https://doi.org/10.3390/nu16060887
Chicago/Turabian StyleWong-Chew, Rosa María, Thi Viet Ha Nguyen, Jossie M. Rogacion, Maxime Herve, and Etienne Pouteau. 2024. "Potential Complementary Effect of Zinc and Alkalihalobacillus clausii on Gut Health and Immunity: A Narrative Review" Nutrients 16, no. 6: 887. https://doi.org/10.3390/nu16060887