
Short-Term Dietary Restriction Potentiates an Anti-Inflammatory Circulating Mucosal-Associated Invariant T-Cell Response
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Selection, Dietary Restriction, and Sample Collection
2.2. Peripheral Blood Mononuclear Cell Isolation from Blood
2.3. Blood Sample Thawing for Single-Cell Workflow
2.4. Library Generation and Sequencing
2.5. Data Preprocessing and Filtering
2.6. Annotation and MAIT Identification
2.7. Differential Expression and MAIT Activation
2.8. Functional Enrichment Analysis
2.9. Stool Sample Collection, Processing, Filtering, and Sequencing
2.10. Microbiome Taxonomic and Functional Pathway Analysis
3. Results
3.1. Participant Demographics
3.2. Filtering and Annotation
3.3. Differential Expression and MAIT Activation
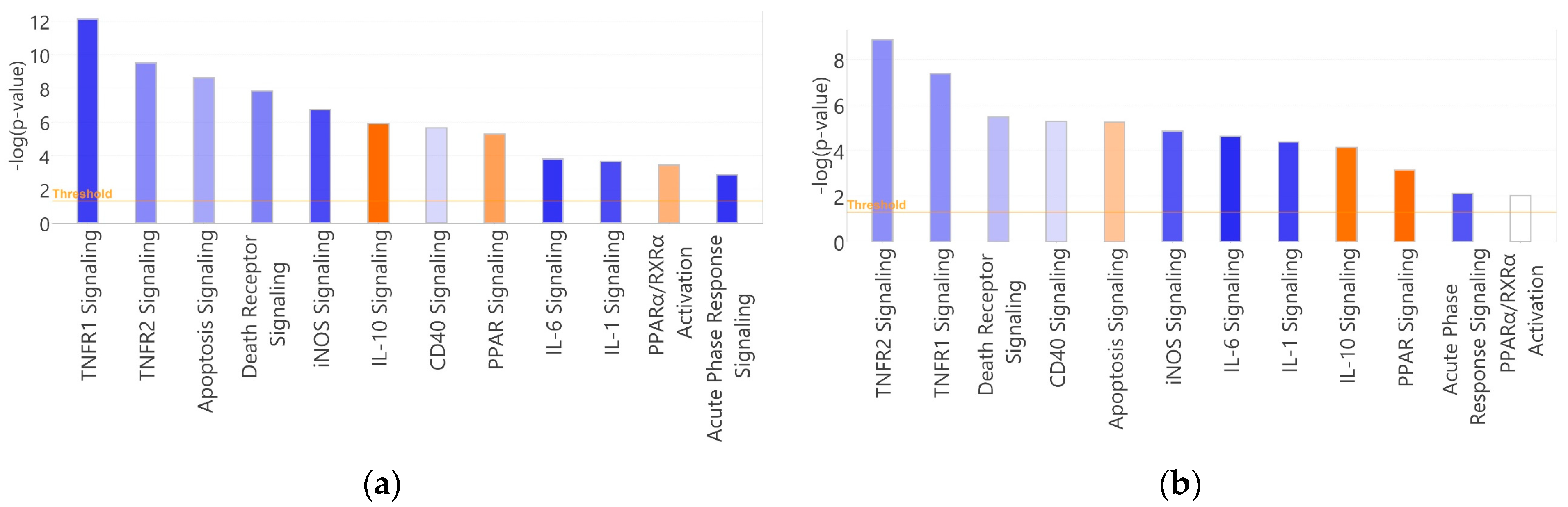
3.4. Canonical Pathways
3.5. Regulators
3.6. Species Abundance Analysis
3.7. Microbial Functional Pathway Analysis
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robertson, L.T.; Treviño-Villarreal, J.H.; Mejia, P.; Grondin, Y.; Harputlugil, E.; Hine, C.; Vargas, D.; Zheng, H.; Ozaki, C.K.; Kristal, B.S.; et al. Protein and Calorie Restriction Contribute Additively to Protection from Renal Ischemia Reperfusion Injury Partly via Leptin Reduction in Male Mice. J. Nutr. 2015, 145, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Varendi, K.; Airavaara, M.; Anttila, J.; Vose, S.; Planken, A.; Saarma, M.; Mitchell, J.R.; Andressoo, J.-O. Short-term Preoperative Dietary Restriction Is Neuroprotective in a Rat Focal Stroke Model. PLoS ONE 2014, 9, e93911. [Google Scholar] [CrossRef]
- Mauro, C.R.; Tao, M.; Yu, P.; Treviño-Villerreal, J.H.; Longchamp, A.; Kristal, B.S.; Ozaki, C.K.; Mitchell, J.R. Preoperative Dietary Restriction Reduces Intimal Hyperplasia and Protects from Ischemia-Reperfusion Injury. J. Vasc. Surg. 2014, 63, 500–509.e1. [Google Scholar] [CrossRef] [PubMed]
- Ruetenik, A.; Barrientos, A. Dietary Restriction, Mitochondrial Function and Aging: From Yeast to Humans. Biochim. Biophys. Acta (BBA) Bioenerg. 2015, 1847, 1434–1447. [Google Scholar] [CrossRef]
- Brandhorst, S.; Harputlugil, E.; Mitchell, J.R.; Longo, V.D. Protective Effects of Short-Term Dietary Restriction in Surgical Stress and Chemotherapy. Ageing Res. Rev. 2017, 39, 68–77. [Google Scholar] [CrossRef]
- Hine, C.; Harputlugil, E.; Zhang, Y.; Ruckenstuhl, C.; Lee, B.C.; Brace, L.; Longchamp, A.; Treviño-Villarreal, J.H.; Mejia, P.; Ozaki, C.K.; et al. Endogenous Hydrogen Sulfide Production Is Essential for Dietary Restriction Benefits. Cell 2014, 160, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Kip, P.; Trocha, K.M.; Tao, M.; O’leary, J.J.; Ruske, J.; Giulietti, J.M.; Trevino-Villareal, J.H.; MacArthur, M.R.; Bolze, A.; Burak, M.F.; et al. Insights From a Short-Term Protein–Calorie Restriction Exploratory Trial in Elective Carotid Endarterectomy Patients. Vasc. Endovasc. Surg. 2019, 53, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Kip, P.; Sluiter, T.J.; Moore, J.K.; Hart, A.; Ruske, J.; O’leary, J.; Jung, J.; Tao, M.; MacArthur, M.R.; Heindel, P.; et al. Short-Term Pre-Operative Protein Caloric Restriction in Elective Vascular Surgery Patients: A Randomized Clinical Trial. Nutrients 2021, 13, 4024. [Google Scholar] [CrossRef]
- Osterholt, T.; Gloistein, C.; Todorova, P.; Becker, I.; Arenskrieger, K.; Melka, R.; Koehler, F.C.; Faust, M.; Wahlers, T.; Benzing, T.; et al. Preoperative Short-Term Restriction of Sulfur-Containing Amino Acid Intake for Prevention of Acute Kidney Injury After Cardiac Surgery: A Randomized, Controlled, Double-Blind, Translational Trial. J. Am. Heart Assoc. 2022, 11, e025229. [Google Scholar] [CrossRef]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 7618. [Google Scholar] [CrossRef]
- Ma, W.; Nguyen, L.H.; Song, M.; Wang, D.D.; Franzosa, E.A.; Cao, Y.; Joshi, A.; Drew, D.A.; Mehta, R.; Ivey, K.L.; et al. Dietary Fiber Intake, the Gut Microbiome, and Chronic Systemic Inflammation in a Cohort of Adult Men. Genome Med. 2021, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.J.; Inniss, S.; Sebepos-Rogers, G.; Rahman, F.Z.; Smith, A.M. The Role of the Microbiome in Gastrointestinal Inflammation. Biosci. Rep. 2021, 41, BSR20203850. [Google Scholar] [CrossRef] [PubMed]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The Role of the Gut Microbiome in Systemic Inflammatory Disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef] [PubMed]
- Brandsma, E.; Kloosterhuis, N.J.; Koster, M.; Dekker, D.C.; Gijbels, M.J.; van der Velden, S.; Ríos-Morales, M.; van Faassen, M.J.; Loreti, M.G.; de Bruin, A.; et al. A Proinflammatory Gut Microbiota Increases Systemic Inflammation and Accelerates Atherosclerosis. Circ. Res. 2019, 124, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Oh, L.; Ab Rahman, S.; Dubinsky, K.; Azanan, M.S.; Ariffin, H. Manipulating the Gut Microbiome as a Therapeutic Strategy to Mitigate Late Effects in Childhood Cancer Survivors. Technol. Cancer Res. Treat. 2023, 22, 15330338221149800. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Rozowsky, J.M.; Fazzone, B.J.; Schmidt, E.A.; Stevens, B.R.; O’malley, K.A.; Scali, S.T.; Berceli, S.A. Temporal Dynamics of the Intestinal Microbiome Following Short-Term Dietary Restriction. Nutrients 2022, 14, 2785. [Google Scholar] [CrossRef] [PubMed]
- Toubal, A.; Nel, I.; Lotersztajn, S.; Lehuen, A. Mucosal-Associated Invariant T Cells and Disease. Nat. Rev. Immunol. 2019, 19, 643–657. [Google Scholar] [CrossRef] [PubMed]
- Amini, A.; Pang, D.; Hackstein, C.-P.; Klenerman, P. MAIT Cells in Barrier Tissues: Lessons from Immediate Neighbors. Front. Immunol. 2020, 11, 584521. [Google Scholar] [CrossRef] [PubMed]
- Legoux, F.; Salou, M.; Lantz, O. MAIT Cell Development and Functions: The Microbial Connection. Immunity 2020, 53, 710–723. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Koay, H.-F.; McCluskey, J.; Gherardin, N.A. The Biology and Functional Importance of MAIT Cells. Nat. Immunol. 2019, 20, 1110–1128. [Google Scholar] [CrossRef]
- Eckle, S.B.G.; Corbett, A.J.; Keller, A.N.; Chen, Z.; Godfrey, D.I.; Liu, L.; Mak, J.Y.W.; Fairlie, D.P.; Rossjohn, J.; McCluskey, J. Recognition of Vitamin B Precursors and Byproducts by Mucosal Associated Invariant T Cells. J. Biol. Chem. 2015, 290, 30204–30211. [Google Scholar] [CrossRef]
- Tastan, C.; Karhan, E.; Zhou, W.; Fleming, E.; Voigt, A.Y.; Yao, X.; Wang, L.; Horne, M.; Placek, L.; Kozhaya, L.; et al. Tuning of Human MAIT Cell Activation by Commensal Bacteria Species and MR1-Dependent T-Cell Presentation. Mucosal Immunol. 2018, 11, 1591–1605. [Google Scholar] [CrossRef] [PubMed]
- Chiba, A.; Murayama, G.; Miyake, S. Mucosal-Associated Invariant T Cells in Autoimmune Diseases. Front. Immunol. 2018, 9, 1333. [Google Scholar] [CrossRef]
- Cibrián, D.; Sánchez-Madrid, F. CD69: From Activation Marker to Metabolic Gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef]
- Legoux, F.; Bellet, D.; Daviaud, C.; El Morr, Y.; Darbois, A.; Niort, K.; Procopio, E.; Salou, M.; Gilet, J.; Ryffel, B.; et al. Microbial Metabolites Control the Thymic Development of Mucosal-Associated Invariant T Cells. Science 2019, 366, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Paffenbarger, R.S., Jr.; Wing, A.L.; Hyde, R.T. Physical activity as an index of heart attack risk in college alumni. Am. J. Epidemiol. 1978, 108, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, B.E.; Leon, A.S.; Richardson, M.T.; Jacobs, D.R.; Paffenbarger, R. Accuracy of the College Alumnus Physical Activity Questionnaire. J. Clin. Epidemiol. 1993, 46, 1403–1411. [Google Scholar] [CrossRef]
- Mifflin, M.D.; St Jeor, S.T.; Hill, L.A.; Scott, B.J.; Daugherty, S.A.; Koh, Y.O. A new predictive equation for resting energy expenditure in healthy individuals. Am. J. Clin. Nutr. 1990, 51, 241–247. [Google Scholar] [CrossRef]
- Ianevski, A.; Giri, A.K.; Aittokallio, T. Fully-Automated and Ultra-Fast Cell-Type Identification Using Specific Marker Combinations from Single-Cell Transcriptomic Data. Nat. Commun. 2022, 13, 1246. [Google Scholar] [CrossRef]
- Vorkas, C.K.; Krishna, C.; Li, K.; Aubé, J.; Fitzgerald, D.W.; Mazutis, L.; Leslie, C.S.; Glickman, M.S. Single-Cell Transcriptional Profiling Reveals Signatures of Helper, Effector, and Regulatory MAIT Cells during Homeostasis and Activation. J. Immunol. 2022, 208, 1042–1056. [Google Scholar] [CrossRef]
- Shi, J.; Zhou, J.; Zhang, X.; Hu, W.; Zhao, J.-F.; Wang, S.; Wang, F.-S.; Zhang, J.-Y. Single-Cell Transcriptomic Profiling of MAIT Cells in Patients with COVID-19. Front. Immunol. 2021, 12, 700152. [Google Scholar] [CrossRef] [PubMed]
- Yanai, I.; Benjamin, H.; Shmoish, M.; Chalifa-Caspi, V.; Shklar, M.; Ophir, R.; Bar-Even, A.; Horn-Saban, S.; Safran, M.; Domany, E.; et al. Genome-Wide Midrange Transcription Profiles Reveal Expression Level Relationships in Human Tissue Specification. Bioinformatics 2004, 21, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. Voom: Precision Weights Unlock Linear Model Analysis Tools for RNA-Seq Read Counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.V.D.; Perraudeau, F.; Soneson, C.; Love, M.I.; Risso, D.; Vert, J.-P.; Robinson, M.D.; Dudoit, S.; Clement, L. Observation Weights Unlock Bulk RNA-Seq Tools for Zero Inflation and Single-Cell Applications. Genome Biol. 2018, 19, 24. [Google Scholar] [CrossRef]
- Matsuyama, H.; Isshiki, T.; Chiba, A.; Yamaguchi, T.; Murayama, G.; Akasaka, Y.; Eishi, Y.; Sakamoto, S.; Homma, S.; Miyake, S. Activation of Mucosal-Associated Invariant T Cells in the Lungs of Sarcoidosis Patients. Sci. Rep. 2019, 9, 13181. [Google Scholar] [CrossRef] [PubMed]
- Radulovic, K.; Niess, J.H. CD69 Is the Crucial Regulator of Intestinal Inflammation: A New Target Molecule for IBD Treatment? J. Immunol. Res. 2015, 2015, 497056. [Google Scholar] [CrossRef] [PubMed]
- Rea, I.; McNerlan, S.; Alexander, H. CD69, CD25, and HLA-DR Activation Antigen Expression on CD3+ Lymphocytes and Relationship to Serum TNF-Alpha, IFN-Gamma, and SIL-2R Levels in Aging. Exp. Gerontol. 1999, 34, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Hinks, T.S.C.; Zhang, X.-W. MAIT Cell Activation and Functions. Front. Immunol. 2020, 11, 1014. [Google Scholar] [CrossRef]
- Deschler, S.; Kager, J.; Erber, J.; Fricke, L.; Koyumdzhieva, P.; Georgieva, A.; Lahmer, T.; Wiessner, J.R.; Voit, F.; Schneider, J.; et al. Mucosal-Associated Invariant T (MAIT) Cells Are Highly Activated and Functionally Impaired in COVID-19 Patients. Viruses 2021, 13, 241. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Kim, D.; Song, L.; Breitwieser, F.P.; Salzberg, S.L. Centrifuge: Rapid and sensitive classification of metagenomic sequences. Genome Res. 2016, 26, 1721–1729. [Google Scholar] [CrossRef]
- Carolan, E.; Tobin, L.M.; Mangan, B.A.; Corrigan, M.; Gaoatswe, G.; Byrne, G.; Geoghegan, J.; Cody, D.; O’connell, J.; Winter, D.C.; et al. Altered Distribution and Increased IL-17 Production by Mucosal-Associated Invariant T Cells in Adult and Childhood Obesity. J. Immunol. 2015, 194, 5775–5780. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, I.; Pingris, K.; Poitou, C.; Bessoles, S.; Venteclef, N.; Kiaf, B.; Beaudoin, L.; Da Silva, J.; Allatif, O.; Rossjohn, J.; et al. Mucosal-Associated Invariant T Cell Alterations in Obese and Type 2 Diabetic Patients. J. Clin. Investig. 2015, 125, 1752–1762. [Google Scholar] [CrossRef] [PubMed]
- Bergin, R.; Kinlen, D.; Kedia-Mehta, N.; Hayes, E.; Cassidy, F.C.; Cody, D.; O’shea, D.; Hogan, A.E. Mucosal-Associated Invariant T Cells Are Associated with Insulin Resistance in Childhood Obesity and Disrupt Insulin Signaling via IL-17. Diabetologia 2022, 65, 1012–1017. [Google Scholar] [CrossRef]
- Berod, L.; Friedrich, C.; Nandan, A.; Freitag, J.; Hagemann, S.; Harmrolfs, K.; Sandouk, A.; Hesse, C.; Castro, C.N.; Bähre, H.; et al. De Novo Fatty Acid Synthesis Controls the Fate between Regulatory T and T Helper 17 Cells. Nat. Med. 2014, 20, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Cluxton, D.; Petrasca, A.; Moran, B.; Fletcher, J.M. Differential Regulation of Human Treg and Th17 Cells by Fatty Acid Synthesis and Glycolysis. Front. Immunol. 2019, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Naimimohasses, S.; O’gorman, P.; Wright, C.; Ni Fhloinn, D.; Holden, D.; Conlon, N.; Monaghan, A.; Kennedy, M.; Gormley, J.; Beddy, P.; et al. Differential Effects of Dietary versus Exercise Intervention on Intrahepatic MAIT Cells and Histological Features of NAFLD. Nutrients 2022, 14, 2198. [Google Scholar] [CrossRef] [PubMed]
- Leeming, E.R.; Johnson, A.J.; Spector, T.D.; Le Roy, C.I. Effect of Diet on the Gut Microbiota: Rethinking Intervention Duration. Nutrients 2019, 11, 2862. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Cintoni, M.; Raoul, P.; Ianiro, G.; Laterza, L.; Lopetuso, L.R.; Ponziani, F.R.; Gasbarrini, A.; Mele, M.C. Gut Microbiota during Dietary Restrictions: New Insights in Non-Communicable Diseases. Microorganisms 2020, 8, 1140. [Google Scholar] [CrossRef] [PubMed]
- Pataky, Z.; Genton, L.; Spahr, L.; Lazarevic, V.; Terraz, S.; Gaïa, N.; Rubbia-Brandt, L.; Golay, A.; Schrenzel, J.; Pichard, C. Impact of Hypocaloric Hyperproteic Diet on Gut Microbiota in Overweight or Obese Patients with Nonalcoholic Fatty Liver Disease: A Pilot Study. Dig. Dis. Sci. 2016, 61, 2721–2731. [Google Scholar] [CrossRef]
- Schubert, K.; Karkossa, I.; Schor, J.; Engelmann, B.; Steinheuer, L.M.; Bruns, T.; Rolle-Kampczyk, U.; Hackermüller, J.; von Bergen, M. A Multi-Omics Analysis of Mucosal-Associated-Invariant T Cells Reveals Key Drivers of Distinct Modes of Activation. Front. Immunol. 2021, 12, 616967. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-M.; Bothwell, A.L.M. The Nuclear Receptor PPARs as Important Regulators of T-Cell Functions and Autoimmune Diseases. Mol. Cells 2012, 33, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, M.; Lefebvre, P.; Staels, B. Molecular mechanism of PPARα action and its impact on lipid metabolism, inflammation and fibrosis in non-alcoholic fatty liver disease. J. Hepatol. 2015, 62, 720–733. [Google Scholar] [CrossRef] [PubMed]
- Duszka, K.; Gregor, A.; Guillou, H.; König, J.; Wahli, W. Peroxisome Proliferator-Activated Receptors and Caloric Restriction—Common Pathways Affecting Metabolism, Health, and Longevity. Cells 2020, 9, 1708. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject | Sex | Race/Ethnicity | Age | BMI | Medications | Tobacco Use | Daily Energy Requirement (kcal) |
|---|---|---|---|---|---|---|---|
| 1 | F | Asian | 22 | 23.4 | Oral Contraceptive | No | 1940 |
| 2 | M | White | 30 | 28.9 | No | 3267 | |
| 3 | M | White | 26 | 26.9 | Magnesium, Krill Oil | No | 2983 |
| 4 | M | White | 28 | 22.4 | No | 2884 | |
| 5 | F | Hispanic | 23 | 32.2 | Oral Contraceptive | No | 2116 |
| 6 | F | White | 19 | 18.9 | Oral Contraceptive | No | 2221 |
| 7 | M | White | 29 | 27.1 | No | 3118 |
| Day 2 | Day 4 | ||||
|---|---|---|---|---|---|
| Regulator | Function | Z-Score | Regulator | Function | Z-Score |
| CD3 | TCR co-receptor | −2.1 | CD3 | −3.3 | |
| NFκB Complex | Cell stress response | −3.7 | NFκB Complex | −4.4 | |
| CD40LG | Costimulation | −2.7 | CD40LG | −3.2 | |
| GM-CSF | Proinflammatory cytokine | −2.2 | GM-CSF | −3.4 | |
| IL2 | T-cell growth | −3.0 | IL2 | −2.6 | |
| KLF6 | Tumor suppressor gene | −3.1 | KLF6 | −3.1 | |
| TNF | Proinflammatory cytokine | −3.9 | TCR | TCR complex | −2.5 |
| IL1B | Proinflammatory cytokine protein | −3.6 | RELA | NFκB complex subunit | −3.6 |
| CD40 | Costimulation | −2.2 | CD28 | Costimulation | −2.8 |
| EPHA2 | Protein kinase | −2.1 | IFNγ | Proinflammatory cytokine | −2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazzone, B.; Anderson, E.M.; Rozowsky, J.M.; Yu, X.; O’Malley, K.A.; Robinson, S.; Scali, S.T.; Cai, G.; Berceli, S.A. Short-Term Dietary Restriction Potentiates an Anti-Inflammatory Circulating Mucosal-Associated Invariant T-Cell Response. Nutrients 2024, 16, 1245. https://doi.org/10.3390/nu16081245
Fazzone B, Anderson EM, Rozowsky JM, Yu X, O’Malley KA, Robinson S, Scali ST, Cai G, Berceli SA. Short-Term Dietary Restriction Potentiates an Anti-Inflammatory Circulating Mucosal-Associated Invariant T-Cell Response. Nutrients. 2024; 16(8):1245. https://doi.org/10.3390/nu16081245
Chicago/Turabian StyleFazzone, Brian, Erik M. Anderson, Jared M. Rozowsky, Xuanxuan Yu, Kerri A. O’Malley, Scott Robinson, Salvatore T. Scali, Guoshuai Cai, and Scott A. Berceli. 2024. "Short-Term Dietary Restriction Potentiates an Anti-Inflammatory Circulating Mucosal-Associated Invariant T-Cell Response" Nutrients 16, no. 8: 1245. https://doi.org/10.3390/nu16081245